seeing the forest for the invasives: mapping … 2012 - seeing the...original paper seeing the...
TRANSCRIPT

ORIGINAL PAPER
Seeing the forest for the invasives: mapping buckthornin the Oak Openings
Richard H. Becker • Kirk A. Zmijewski •
Todd Crail
Received: 15 November 2011 / Accepted: 7 July 2012
� Springer Science+Business Media B.V. 2012
Abstract Landsat TM and ETM? satellite images
from 2001 to 2011 were used to map the extent and
change of the invasive shrubs common and glossy
buckthorn (Frangula alnus and Rhamnus cathartica)
at Irwin Prairie State Nature Preserve (IPSNP), and
throughout Oak Openings, a 1,500 km2 region,
located in NW Ohio, USA and SE Michigan near
Lake Erie. In the Oak Openings, buckthorn directly
threatens native biodiversity and habitat health of this
globally rare ecosystem. Buckthorn that forms as
dense shrub thicket in the understory is often obscured
from satellite view by other canopy and is not
spectrally dissimilar enough to be characterized using
multispectral images. To address this, time series
tasseled cap greenness images of land surface areas
dominated by buckthorn thicket (which exhibit early
leaf-out, late senescence) was used to identify areas
covered by thicket with a heterogeneous background.
A time series of vegetation index values was calcu-
lated from 49 Landsat images and combined with
in-situ observations to define the land surface phenol-
ogy of buckthorn thicket and other vegetation types.
The phenological differences among land surfaces
dominated by distinct vegetation types in the Oak
Openings Region were used to map the extent of
buckthorn thicket using a supervised classification
method. Buckthorn thicket was identified in 0.43 % of
the classified pixels (940 ha) in the 2007–2011
imagery and in 0.31 % (690 ha) of the 2001–2006
images. A Kappa test of the 2007–2011 classification
yielded a value of 0.73 with 88 % overall accuracy of
presence or absence of thicket based on 60 samples
throughout the Oak Openings. The areal extent of
buckthorn thicket increased by 39 % (255 ha) in the
study area over the time period from 2001 to 2011.
Keywords Remote sensing � Buckthorn � Rhamnus
cathartica � Frangula alnus � Multi-temporal �Phenology � Invasive species � Landsat �Oak Openings
Introduction
Non-native invasive species can pose significant
threats to unique ecosystems, threatening native
species, ecosystem properties, and even regional
economies. In 2000 in the US, the estimated cost of
damages associated with management and control of
these invasive species was estimated to be at least
$137 billion per year (Pimentel et al. 2000). When
total value of biodiversity reduction, ecosystem ser-
vice loss, species extinctions and aesthetic decline are
included, the cost of damage increases drastically
(Nunes and van den Bergh 2001).
Understory vegetation has a significant role in
forest ecosystems. Changes in the structure and
R. H. Becker (&) � K. A. Zmijewski � T. Crail
Department of Environmental Sciences, University of
Toledo, 2801 Bancroft Ave, Toledo, OH 43606, USA
e-mail: [email protected]
123
Biol Invasions
DOI 10.1007/s10530-012-0288-8
species composition of understory plants, including
both native and invasive species, can affect the health
and biodiversity and change forest succession
through physical, chemical, and biological mecha-
nisms (Royo and Carson 2006). Alteration of the
normal species composition may lead to the forma-
tion of dense monoculture thickets of non-native
invasive shrubs due to a reduction in recruitment of
native species (Frappier et al. 2003; Fagan and Peart
2004). This can disrupt forest ecosystems as well as
ecosystems that may be adjacent to forests such as
wet prairies. Invasive plant species, such as common
buckthorn (Rhamnus cathartica L.), have the poten-
tial to alter ecosystems by changing carbon, nitrogen,
and pH of soils as well as soil moisture (Heneghan
et al. 2006).
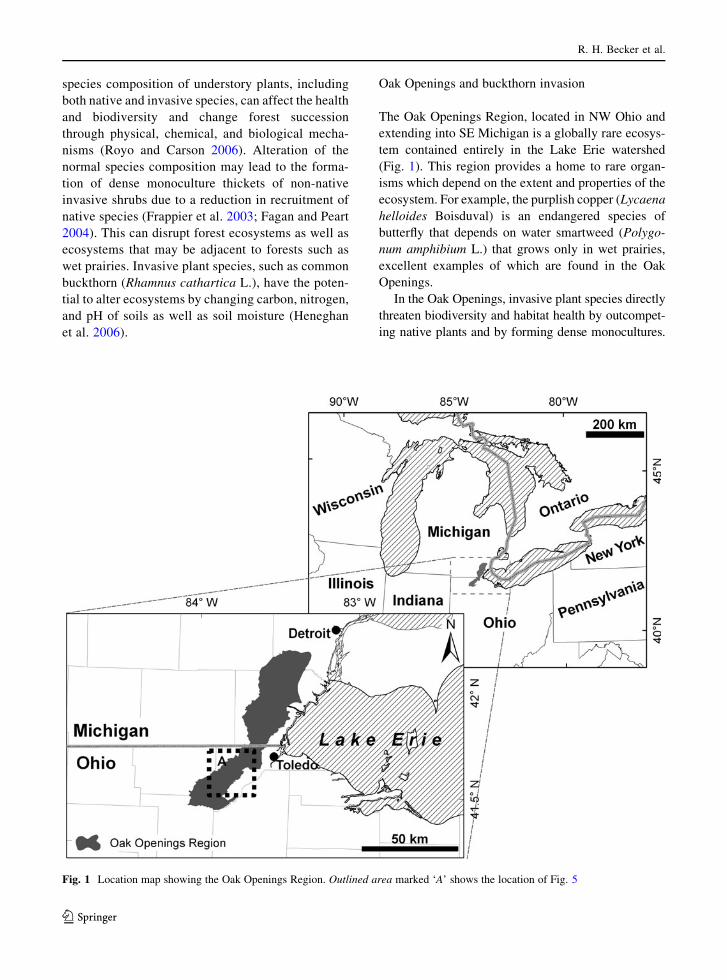
Oak Openings and buckthorn invasion
The Oak Openings Region, located in NW Ohio and
extending into SE Michigan is a globally rare ecosys-
tem contained entirely in the Lake Erie watershed
(Fig. 1). This region provides a home to rare organ-
isms which depend on the extent and properties of the
ecosystem. For example, the purplish copper (Lycaena
helloides Boisduval) is an endangered species of
butterfly that depends on water smartweed (Polygo-
num amphibium L.) that grows only in wet prairies,
excellent examples of which are found in the Oak
Openings.
In the Oak Openings, invasive plant species directly
threaten biodiversity and habitat health by outcompet-
ing native plants and by forming dense monocultures.
Fig. 1 Location map showing the Oak Openings Region. Outlined area marked ‘A’ shows the location of Fig. 5
R. H. Becker et al.
123

Other invasive shrubs which form similar thickets in the
region include honeysuckles (Lonicera sp.) and autumn
olive (Elaeagnus umbellata). At locations throughout
the Oak Openings, buckthorn has emerged as a great
threat to the native plant communities such as: oak
woodland, mesic tallgrass prairie, twig rush prairie and
oak flatwood wetlands. The native plant communities
are adapted to low nutrients and periodic fires;
invasions are successful when this regime is disrupted
either through suppression of fires or an influx of
nutrients (Abella et al. 2001; Zouhar et al. 2008).
Recent studies suggest that areas with higher biodiver-
sity such as the Oak Openings, more frequently
experience invasions than do lower diversity areas
(Stohlgren et al. 2003). This region is covered by a
variety of heterogeneous land uses, including suburban,
agricultural, forested, prairie, roadway, with Toledo
Express airport also located in this area. Within the
Ohio Oak Openings Region there are multiple large
metroparks and nature preserves including Oak Open-
ings Metropark, Secor Metropark, Wildwood Preserve
Metropark, Irwin Prairie State Nature Preserve (IPS-
NP), and the Kitty Todd Nature Preserve (shown in
Fig. 5). All of these areas are actively managed for
invasive species, of which glossy and common buck-
thorn are included.
Common and glossy buckthorn (Rhamnus cathar-
tica and Frangula alnus) are woody shrubs of
European origin. They form dense thickets, and have
been used to form hedgerows. Both species of
buckthorn were introduced in the early to mid 1800s,
but did not reach Ohio until the 1920s (Possessky et al.
2000; Torrey 1824; Knight et al. 2007). However,
plant surveys in the Oak Openings listed both buck-
thorn species to be ‘‘rare’’ or ‘‘infrequent’’ as recently
as 1978 (Easterly 1979; Moseley 1928). Contempora-
neous introduction of invasive birds such as the
European starling also contributed to the spread of
these plants throughout the continent (Howell and
Blackwell 1977).
Buckthorn has high fecundity and germination rates
upwards of 85 % with as many as 90 % of the berries
falling directly under the parent tree (Frappier et al.
2003; Knight et al. 2007). Frappier et al. (2003) found
dispersal in excess of six meters per year without the
assistance of birds. The berries can survive inundation
for multiple weeks and also can germinate after
digestion by birds such as the European starling and
American robin (Knight et al. 2007; Skinner et al.
2005; Lafleur et al. 2007). Some of the berries also
float in surface water, further contributing to expan-
sion of the plant extent (Howell and Blackwell 1977).
All of these factors contribute to high rates of
spreading after the initial invasion has been
established.
Buckthorn also may facilitate the invasion and
spread of other noxious invasive species. The Asian
soybean aphid (Aphis glycines) has emerged in the
past decade as a top agricultural pest threatening
soybean crops by providing a host for overwintering
(Venette and Ragsdale 2004; Voegtlin et al. 2005).
Invasive earthworms also pose a significant problem
by removing forest floor leaf litter and other organic
material (buckthorn does not germinate well in thick
organic layers) and in turn enriching the soil with
nitrogen which buckthorn may utilize more readily
than native plants (Hale et al. 2006; Heimpel et al.
2010). Buckthorn thickets provide shade on the forest
floor and large amounts of high quality leaf litter
which earthworms preferentially consume providing a
positive feedback loop for both invasive species
(Madritch and Lindroth 2009; Heneghan et al. 2007).
When woody shrub plants (such as buckthorn) exceed
approximately 30 % cover in woodlands, they reduce
the species richness and suppress the growth of tree
seedlings (Woods 1993). Glossy buckthorn also alters
the properties of the soil, increasing the soil nitrogen,
carbon, pH and soil moisture.
Past methods of identifying invasives
from satellite
Previous methods of identifying non-native plant
species using remote sensing have included using
moderate spatial and spectral resolution images such
as Landsat TM over very short temporal windows
associated with specific phenological differences
(Resasco et al. 2007); a combination of vegetation
response to rainfall and early season phenology over a
short time window(Bradley and Mustard 2006); or
using phenology metrics such as accumulated degree
days (De Beurs and Henebry 2005), minimum relative
humidity (Brown and de Beurs 2008), length of
season, or leaf-out day of year (Tuanmu et al. 2010). In
addition, high spatial resolution images such as color
infrared aerial photography and IKONOS or Quick-
bird imagery (Mullerova et al. 2005) have been used.
High temporal resolution low spatial resolution
Mapping buckthorn in the Oak Openings
123

remote sensing such as a AVHRR and MODIS has
been used to examine variations in the phenology over
large spatial extents with changing climate conditions
(e.g. Justice et al. 1985; Morisette et al. 2009); and
hyperspectral remote sensing to identify variations in
spectral absorption between plant species (e.g. Under-
wood et al. 2003). Other methods have focused on
single image acquisitions, and examining how sensi-
tive a variety of different vegetation indices are to the
vegetation type to discriminate between vegetation
types (Kandwal et al. 2009). Many of these methods
assess the variation of an invasive against a homoge-
neous background canopy or vegetation cover. Unfor-
tunately, a homogeneous background is not present in
the Oak Openings. As a whole, these methods either
sacrifice temporal resolution for spatial resolution or
sacrifice spatial resolution for temporal resolution.
Recently, Landsat images have become available
en masse at no cost, so it is no longer necessary to try to
discriminate phenology using moderate resolution
(30 9 30 m) images, while limiting analyses to only
2–4 scenes in a year. By taking advantage of a high
number of scenes in a multi-year image stack, we can
more fully utilize the variations in phenology to
discriminate among land covers and identify species
of interest.
Phenology
The phenology of buckthorn species is different from
other vegetation species in the study area. Buckthorn
leafs out earlier and remains green longer than a
majority of the tree canopy (shown in Fig. 2). The
NDVI and tasseled cap greenness of other land covers
either remains fairly constant throughout the year
(pine plantation, water, urban), starts later and ends
earlier (oak savanna, oak blueberry forest), or is not as
high (wet prairie, grasslands). Agricultural fields
exhibit land surface phenologies related to time of
planting and crop type. As a result, pixels dominated
by buckthorn species exhibit a distinct land surface
phenology which can be used to discriminate these
pixels from those with other land covers utilizing only
a time series of vegetation indices throughout the
course of a year.
Theoretical approach
In the Oak Openings Region, buckthorn appears in the
understory of forested areas, as a dense monoculture
encroaching over prairie, and along edges of disturbed
areas, such as agricultural drainage ditches adjoining
fields and pasture lands. Any method to identify these
thickets must be able to identify the plants both nested
in the understory and when they appear in dense
monoculture environments. In addition, the high
heterogeneity of the landscape relative to the scale
of lower resolution sensors means that methods
utilizing lower spatial resolution sensors (e.g. Justice
et al. 1985; Reed et al. 1994), such as MODIS, MERIS
and AVHRR, are not sufficient for identifying these
thickets. Buckthorn thickets may occur on the spatial
scale ranging from 50 m across up to several hectares
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
100 150 200 250 300 350
Tass
eled
-Cap
Gre
nn
ess
Ind
ex
Day of Year
Land Surface Phenologies
Buckthorn Thicket Wet Prairie Oak Savanna Oak Forest
Fig. 2 Land surface
phenology derived from
Landsat TC-G images from
2007 to 2011, showing high
early season greenness
index of buckthorn thicket.
Dashed lines represent 1
standard deviation of TC-G
in that land surface type
R. H. Becker et al.
123

in extent. Thickets as small as 50–100 m across may
be identified using Landsat resolution.
The spectral characteristics of buckthorn are not
sufficiently different from oak species (Quercus
spp.) for them to be uniquely discriminated by
spectral means using an instrument with moderate
spectral resolution, such as Landsat TM/ETM?,
SPOT or ASTER. In addition, high spectral resolu-
tion imagery is spatially and temporally limited, and
expensive.
Previous techniques using moderate resolution data
which either used specific senescence dates (Resasco
et al. 2007), or a regression between phenology at
multiple dates (Peterson 2008) were found not to work
in this environment. In order to discriminate between a
the wide variety of heterogeneous land cover types
found in the Oak Opening Region, a full-year phenol-
ogy was found in this study to be far more effective
than simply using two dates. The different land covers
which exhibit distinct satellite-based land surface
phenologies for the area include: buckthorn thicket,
twig rush wet prairie, pine plantation, maintained
(irrigated) golf course, un-irrigated pasture, farm fields
(corn, soy, alfalfa), and non-vegetated cover including
blacktop and water (sample land surface phenology
curves for pixels representing these land cover types
are shown in Fig. 2).
In the lower Great Lakes region, cloud cover can be
a significant issue in the acquisition of satellite
imagery. This cloud cover, combined with the low
temporal repeat of Landsat (14 days), means that a
single year of data can rarely be used to reveal a
complete phenological cycle of any of the plant
communities. To overcome this shortcoming of the
annual data set, we made use of a multi-year data set,
atmospherically corrected, spatially georeferenced,
and arranged by day of year to both define different
phenology types, and to classify image pixels. Because
the image data are acquired at discrete intervals in time,
the phenologic curve is interpolated between measure-
ments to give a continuous annual phenology. This
interpolated continuous curve can be resampled to
coincide with a different time series covering a
different set of years, and acquired on different days
of those years. Once the time series of vegetation index
images is created, these can be temporally resampled to
match the new image data stack, and then classified
using a supervised classification method based on the
phenology curve.
Methods
Satellite image processing
To generate the Landsat based land surface phenology,
and classify areas for presence/absence of buckthorn
thicket, the following steps were conducted.
Forty-nine images were acquired from the USGS
Eros Data Center. Images from Landsat 5 TM and
Landsat 7 ETM? were acquired covering path 20 and
row 31 (Table 1). Scenes were selected to minimize
cloud cover over the area. To eliminate inter-annual
variation due to drought, data was not selected for the
image stacks during years in which the monthly
historical Palmer Drought Severity Index (PDSI) for
the study area showed moderate to severe drought
conditions (PDSI below -2.0) for more than two
Table 1 Landsat image acquisition dates used in both the
2001–2006 and 2007–2011 stacks
2001–2006 Sensor type 2007–2011 Sensor type
14-Mar-01 ETM? 7 4-Mar-09 ETM? 7
22-Mar-01 TM 5 18-Apr-08 ETM? 7
13-Apr-03 TM 5 24-Apr-07 TM 5
18-Apr-05 TM 5 5-May-11 TM 5
29-Apr-03 TM 5 10-May-10 TM 5
1-May-01 ETM? 7 18-May-07 ETM? 7
4-May-05 ETM? 7 20-May-08 ETM? 7
23-May-06 TM 5 28-May-08 TM 5
16-Jun-03 TM 5 6-Jun-11 TM 5
24-Jun-06 TM 5 11-Jun-07 TM 5
7-Jul-02 ETM? 7 24-Jun-09 ETM? 7
15-Jul-02 TM 5 7-Jul-08 ETM? 7
23-Jul-05 TM 5 15-Jul-08 TM 5
31-Jul-05 ETM? 7 16-Aug-08 TM 5
8-Aug-05 TM 5 1-Sep-08 TM 5
21-Aug-01 ETM? 7 15-Sep-10 TM 5
1-Sep-02 TM 5 25-Sep-08 ETM? 7
6-Sep-04 TM 5 1-Oct-10 TM 5
17-Sep-02 TM 5 9-Oct-10 ETM? 7
30-Sep-04 ETM? 7 17-Oct-10 ETM? 7
8-Oct-01 ETM? 7 19-Oct-08 TM 5
24-Oct-01 ETM? 7 2-Nov-10 ETM? 7
12-Nov-05 TM 5 7-Nov-09 TM 5
20-Nov-05 ETM? 7 1-Dec-09 ETM? 7
1-Dec-03 ETM? 7
The sensor used to acquire the image is also listed
Mapping buckthorn in the Oak Openings
123

consecutive spring and summer months (Palmer
1965).
Images were atmospherically corrected using the
FLAASH atmospheric correction package imple-
mented in the ENVI software (Cooley et al. 2002).
Visibility at the Toledo Express airport was obtained for
the hour closest to scene acquisition for model input
(Roche 2011). Toledo Express is located at the center of
the scene of interest. Orbital parameters, scene center
acquisition date/time, latitude and longitude of scene
center were acquired from the image headers.
Images were co-registered and resampled to a
common base image. In order to allow for the highest
quality vegetation index generation, nearest neighbor
resampling was used to preserve the original spectra.
The co-registered, atmospherically corrected land
surface reflectances were used to calculate Tasseled
Cap Greenness (TC-G) transformation (Crist and
Kauth 1986) using the formula:
G ¼ �0:2728TM1� 0:2174TM2� 0:5508TM3
þ 0:7221TM4þ 0:0733TM5� 0:1648TM7
Enhanced Vegetation Index (EVI) and Normalized
Difference Vegetation Index (NDVI) were also cal-
culated for the same scenes based on the same co-
registered atmospherically corrected images.
Locations with known-type land surface coverage
were identified in the field, including buckthorn
thicket, turf grass, evergreen forest, open water,
asphalt, oak forest, wet prairie, and oak savanna. An
area of interest, with a minimum of 15 pixels and a
maximum of 300 pixels, was defined for each of
these land cover types. TC-G, EVI and NDVI images
spanning 2007–2011 were stacked in an image stack
indexed by day of year, generating a land-surface
phenology curve for each pixel location throughout
the entire region. For each of the land cover types, a
time series of each vegetation index was extracted
using the z-profile tool in ENVI based on the area of
interest chosen for each of the designated type land
surface coverages.
An image stack for 2007–2011 was generated for
each vegetation index, indexed by day of year
within the image stack. The parallelepiped method
of image classification was employed on each time
series stack and locations with buckthorn thicket
were identified using training sets that were obtained
from field sites that were identified as buckthorn
thicket. In addition, the maximum likelihood clas-
sification method was used, using phenology of
buckthorn thicket, turf grass, evergreen forest, open
water, asphalt, oak forest, wet prairie, and oak
savanna, as the land cover classification types. The
accuracy assessment used for each image classifica-
tion based on field validation data was also assessed
and is discussed in Sect. 3.3.
Image stacks covering the periods 2001–2006, and
2007–2011 were generated. Based on the accuracy
assessment results, the TC-G was selected as the best
vegetation index, and the parallelepiped classification
method as producing the most accurate results as
determined by the kappa test. The land surface
phenology curves were resampled to the day of year
indicies that were used in each image stack. The TC-
G image stacks were classified using the resampled
phenology curves with the parallelepiped classifica-
tion method.
Field characterization of vegetation
To provide a basis for validation of the remotely sensed
classification of buckthorn species thicket, the follow-
ing field measurements were conducted at IPSNP and
Bumpus Pond, a property owned by Toledo Area
Metroparks. Both of these properties have extensive
buckthorn thickets and are actively managed to remove
buckthorn and restore native wet prairie.
Areas of buckthorn thicket were sampled for stem
density, species composition, and canopy closure to
determine the definition and variability of thickets at a
set of ten P-Plot locations (Fig. 4). Nested vegetation
sampling has been widely used as a means to provide
an accurate representation of both plant species
richness and diversity at multiple spatial scales from
1 to 1,000 m2 plot sizes (Stohlgren et al. 1995). Pixel
nested plot (P-Plot) sampling utilizes a similar sam-
pling style as a modified-Whittaker plot but at a scale
that is similar to the nominal ground resolution of
sensors such as Landsat TM and ETM? (Kalkhan
et al. 2007). Due to the impenetrable nature of
buckthorn thickets, a 30 9 30 m P-Plot (a hybrid of
a pixel nested plot and a modified Whittaker plot) was
used. Figure 3 shows the layout of the plot. Because
graminoids and non-woody herbaceous species in this
R. H. Becker et al.
123

study area make a minimal contribution to overall
reflectance values at the 30 m Landsat scale, their
presence was noted in each sub-plot, but only woody
plants were considered during sampling. Ten
1 m x 1 m plots and two 3 m 9 3 m (A and B) were
sampled for stem density, species composition, and
canopy closure. All species within the 10 m 9
10 m sub-plot (C) were noted for presence. The larger
30 m 9 30 m plot was surveyed for species that were
not found in any of the smaller plots. A spherical den-
sitometer was used to determine canopy coverage
(Lemmon 1956). Each subplot was averaged to
determine species composition, density, and canopy
coverage for the entire 30 m 9 30 m plot. Based on
these ten P-plot locations, distributed over a 6.5 km2
area in a variety of buckthorn densities, a character-
ization of low, medium, and high buckthorn coverage
was determined and used for a rapid assessment of
larger numbers of locations.
In addition, 50 sample locations throughout the Oak
Openings in a 35 km2 area were selected for method
validation. Based on the measurement of buckthorn
density at the ten P-plot locations, a measure of ‘‘low’’,
‘‘medium’’ or ‘‘high’’ was assigned to samples at 50
other selected sample locations (Table 2).
Accuracy assessment
The image classification results and field measure-
ments were compared to determine the accuracy of
products derived using the above method. The
classifications of buckthorn derived from each of
the vegetation (TC-G, EVI, NDVI) index stacks
using each classification method on the 2007–2011
image stacks were checked against the results of
field measurements. Measurements at the ten P-plot
sites and the 50 other field validation sites were
used to assess the accuracy. Kappa values were
calculated for each image interpretation method
(Congalton 1991) for the presence or absence of
buckthorn thicket at locations throughout the
images (Table 3).
Fig. 3 Configuration of P-Plot canopy coverage sampling
scheme used to determine buckthorn thicket density of locations
in Fig. 4
Table 2 Ten 30 9 30 m
P-plots used in the study
(marked as stars on Fig. 4)
showing the percentage of
buckthorn plants classified
as overstory, the percentage
of buckthorn plants
compared to overall species
composition, total canopy
coverage (late May 2011),
and density classification
(None-High) based on the
number of buckthorn plants
per area
The latitude and longitude
of the plots is also listed
Plot
ID
Buckthorn
percentages of
overstory species
composition
Buckthorn overall
percentage of
species
composition
Canopy
coverage
Buckthorn
density
Latitude Longitude
A 23 44 92 LOW 41.6563 -83.7809
B 97 92 93 HIGH 41.6527 -83.7753
C 96 76 93 HIGH 41.6356 -83.7919
D 60 86 94 MED 41.6363 -83.7903
E 94 91 90 HIGH 41.6359 -83.7912
F 23 74 83 LOW 41.6529 -83.7696
G 0 0 87 NONE 41.6577 -83.7713
H 78 79 86 MED 41.6568 -83.7784
I 90 96 95 HIGH 41.6522 -83.7765
J 86 87 88 HIGH 41.6577 -83.7807
Mapping buckthorn in the Oak Openings
123

Results and discussion
Time series images of vegetation indexes from 2007 to
2011 were used to produce presence absence maps of
buckthorn species in the Oak Openings Region. With a
kappa test value of 0.73, the TC-G image stack
processed with the parallelepiped classification
method was found to have the highest accuracy of
buckthorn species thicket identification at 88 %
(Figs. 4, 5). The phenological differences are suffi-
cient to successfully discriminate buckthorn species
dominated thicket from various heterogeneous back-
grounds including wet prairie, pine plantation, main-
tained (irrigated) golf course, non-irrigated pasture,
farm fields (corn, soy, alfalfa), and non-vegetated
cover including blacktop and water.
Thickets of buckthorn species were identified
throughout the entire region—present in 0.43 %
(945 ha) of the land cover in the Oak Openings
Region in 2007–2011 (Fig. 5). Larger areal extents of
thicket were identified in clusters surrounded by or
adjacent to agricultural drainage ditches. These
ditches were observed to deliver high sediment,
nutrient rich waters to areas adjoining them, such as
Fig. 4 Map of buckthorn
thicket classified from 2007
to 2011 Landsat TC-G
images in the areas near
IPSNP and Bumpus Pond.
Stars mark the location of
P-plots used in the study.
Locations marked with x’s
show confirmation samples
Table 3 Confusion matrix showing accuracy assessment of
buckthorn thicket classification of the 2007–2011 Landsat
images
Class Buckthorn No buckthorn Row total
Buckthorn 15 4 19
No buckthorn 3 38 41
Column total 18 42 60
Overall accuracy 88.33 %
Kappa hat 0.7266
Buckthorn No buckthorn
Ommission error 16.67 9.52
Commission error 21.05 7.32
R. H. Becker et al.
123

the IPSNP, during spring thaw and flooding poten-
tially contributing to the success of the buckthorn
species.
Using the two time steps of 2001–2006, and
2007–2011, we were able to document the increase
in the extent of buckthorn thicket. This change in land
cover showed a consistent increase throughout the Oak
Openings Region from 0.31 % (690 ha) to 0.43 %
(945 ha) of pixels in the study area.
Examination of the time series images at IPSNP
shows that results from these images agree with
documented management practices at IPSNP. For
example, in 2002–2003, a 11.5-ha management unit in
the south-central portion of the nature preserve
received multiple treatment methods to remove buck-
thorn (Moser et al. 2005). In Fig. 6, area ‘C’,
buckthorn is absent in both the 2001–2006 and
2007–2011 image stack (Fig. 6, area ‘C’). An addi-
tional 10 ha area in the north central portion of the
preserve was remediated by removal using a hydro-ax
in 2007, and this area shows buckthorn present in the
2001–2006 image stack, but not in the 2007–2011
image stack (Fig. 6, area ‘A’) A final area was
remediated in the fall of 2010, and this area shows a
presence of buckthorn in an image stack from 2007 to
2009, but absent in a stack from 2007 to 2011 (Fig. 6,
area ‘B’).
This method is of land surface phenology extraction
from multi-year Landsat data, and classification of
species type is applicable for other areas or species
where a distinct phenological difference is notable.
The technique could prove particularly useful in other
areas where a species of interest present in understory
leafs out earlier than the overstory canopy.
Fig. 5 Regional
distribution map of Landsat
derived, non-native
buckthorn thicket locations
in the Oak Openings. Large
regions of buckthorn thicket
are linked to areas with
significant agricultural
drainage ditches
Mapping buckthorn in the Oak Openings
123

Acknowledgments This research was funded by the through
the Ohio Lake Erie Commission (OLEC) and the Lake Erie
Protection Fund (LEPF) Small Grant #416-11. The LEPF is
supported by the citizens of Ohio through their purchase of the
Lake Erie License Plate. We thank both Toledo Area
Metroparks and Irwin Prairie State Nature Preserve for
allowing us access to their property to conduct this research,
and Jonathon Sanders for field assistance. This is contribution
#2012-10 from the Lake Erie Research Center.
References
Abella SR, Jaeger JF, Gehring DH, Jacksy RG, Menard KS,
High KA (2001) Restoring historic plant communities in
the Oak Openings region of Northwestern Ohio. Ecol
Restor 19(3):155–160
Bradley BA, Mustard JF (2006) Characterizing the landscape
dynamics of an invasive plant and risk of invasion using
remote sensing. Ecol Appl 16(3):1132–1147
Fig. 6 Local change in
Landsat derived buckthorn
thicket locations at IPSNP
overlain on a 2006 aerial
photograph. Area A was
remediated in 2006–2007,
area B was remediated in
2010–2011, and area C was
remediated in 2002–2003
before a majority of image
acquisitions occurred
R. H. Becker et al.
123

Brown ME, de Beurs KM (2008) Evaluation of multi-sensor
semi-arid crop season parameters based on NDVI and
rainfall. Remote Sens Environ 112(5):2261–2271. doi:
10.1016/j.rse.2007.10.008
Congalton RG (1991) A review of assessing the accuracy of
classifications of remotely sensed data. Remote Sens
Environ 37(1):35–46
Cooley T, Anderson G, Felde G, Hoke M, Ratkowski A, Chet-
wynd J, Gardner J, Adler-Golden S, Matthew M, Berk A
(2002) FLAASH, a MODTRAN4-based atmospheric cor-
rection algorithm, its application and validation. IEEE
1413:1414–1418
Crist E, Kauth R (1986) The Tasseled Cap de-mystified (trans-
formations of MSS and TM data). Photogram Eng Remote
Sens 52:81–86
De Beurs KM, Henebry GM (2005) Land surface phenology and
temperature variation in the international geosphere–bio-
sphere program high-latitude transects. Glob Change Biol
11(5):779–790. doi:10.1111/j.1365-2486.2005.00949.x
Easterly NW (1979) Non-indigenous plant species in the Oak
Openings of Northwestern Ohio. Castanea 44(3):142–149
Fagan M, Peart D (2004) Impact of the invasive shrub glossy
buckthorn (Rhamnus frangula L.) on juvenile recruitment
by canopy trees. For Ecol Manage 194(1–3):95–107
Frappier B, Lee TD, Olson KF, Eckert RT (2003) Small-scale
invasion pattern, spread rate, and lag-phase behavior of
Rhamnus frangula L. For Ecol Manage 186:1–6
Hale CM, Frelich LE, Reich PB (2006) Changes in hardwood
forest understory plant communities in response to Euro-
pean earthworm invasions. Ecology 87(7):1637–1649
Heimpel GE, Frelich LE, Landis DA, Hopper KR, Hoelmer KA,
Sezen Z, Asplen MK, Wu K (2010) European buckthorn
and Asian soybean aphid as components of an extensive
invasional meltdown in North America. Biol Invasions
12(9):2913–2931
Heneghan L, Fatemi F, Umek L, Grady K, Fagen K, Workman
M (2006) The invasive shrub European buckthorn
(Rhamnus cathartica L.) alters soil properties in Mid-
western US woodlands. Appl Soil Ecol 32(1):142–148.
doi:10.1016/j.apsoil.2005.03.009
Heneghan L, Steffen J, Fagen K (2007) Interactions of an
introduced shrub and introduced earthworms in an Illinois
urban woodland: impact on leaf litter decomposition.
Pedobiologia 50(6):543–551
Howell JA, Blackwell WH (1977) The history of Rhamnusfrangula (Glossy Buckthorn) in the Ohio flora. Castanea
42(2):111–115
Justice CO, Townshend JRG, Holben BN, Tucker CJ (1985)
Analysis of the phenology of global vegetation using mete-
orological satellite data. Int J Remote Sens 6(8):1271–1318
Kalkhan MA, Stafford EJ, Stohlgren TJ (2007) Rapid plant
diversity assessment using a pixel nested plot design: a case
study in Beaver Meadows, Rocky Mountain National Park,
Colorado, USA. Divers Distribut 13(4):379–388
Kandwal R, Jeganathan C, Tolpekin V, Kushwaha SPS (2009)
Discriminating the invasive species, ‘Lantana’ using
vegetation indices. J Indian Soc Remote Sens 37(2):
275–290
Knight KS, Kurylo JS, Endress AG, Stewart JR, Reich PB
(2007) Ecology and ecosystem impacts of common
buckthorn (Rhamnus cathartica): a review. Biol Invas
9:925–937
Lafleur NE, Rubega MA, Elphick CS (2007) Invasive fruits,
novel foods, and choice: an investigation of European
starling and American Robin frugivory. Wilson J Ornithol
119(3):429–438. doi:10.1676/05-115.1
Lemmon PE (1956) A spherical densiometer for estimating
forest overstory density. Forensic Sci 2(4):314–320
Madritch MD, Lindroth RL (2009) Removal of invasive shrubs
reduces exotic earthworm populations. Biol Invas 11(3):
663–671
Morisette JT, Richardson AD, Knapp AK, Fisher JI, Graham
EA, Abatzoglou J, Wilson BE, Breshears DD, Henebry
GM, Hanes JM, Liang L (2009) Tracking the rhythm of the
seasons in the face of global change: phenological research
in the 21st century. Front Ecol Environ 7(5):253–260. doi:
10.1890/070217
Moseley EL (1928) Flora of the Oak Openings. Ohio Acad Sci
Spec Pap 20:79–134
Moser M, Arbour T, Schneider G (2005) Ecological restoration
of Irwin State Nature Preserve: control of glossy buckthorn
in a unique Oak-Openings habitat. Paper presented at the
Ohio invasive plant research conference: bridging the gap
between land management and research
Mullerova J, Pysek P, Jarosik V, Pergl J (2005) Aerial photographs
as a tool for assessing the regional dynamics of the invasive
plant species Heracleum mantegazzianum. J Appl Ecol
42(6):1042–1053. doi:10.1111/j.1365-2664.2005.01092.x
Nunes PALD, van den Bergh JCJM (2001) Economic valuation
of biodiversity: sense or nonsense? Ecol Econ 39(2):
203–222
Palmer WC (1965) Meterological drought. Weather Bureau,
NOAA Library and Information services Division, Wash-
ington, DC
Peterson EB (2008) Invasion of the exotic grasses: mapping
their progression via satellite. In: Kitchen SG, Pendleton
RL, Monaco TA, Vernon J (eds) Shrublands under fire:
disturbance and recovery in a changing world. Usda Forest
Service Rocky Mountain research station proceedings, vol
52, pp 33–39
Pimentel D, Lach L, Zuniga R, Morrison D (2000) Environ-
mental and economic costs of nonindigenous species in the
United States. Bioscience 50(1):53–65
Possessky SL, Williams C, Moriarity W (2000) Glossy buck-
thorn, Rhamnus frangula L.: a threat to riparian plantcommunities of the Northern Allegheny Plateau (USA).
Nat Areas J 20(3):290–292
Reed BC, Brown JF, Vanderzee D, Loveland TR, Merchant JW,
Ohlen DO (1994) Measuring phenological variability from
satellite imagery. J Veg Sci 5(5):703–714
Resasco J, Hale AN, Henry MC, Gorchov DL (2007) Detecting
an invasive shrub in a deciduous forest understory using
late-fall Landsat sensor imagery. Int J Remote Sens 28(16):
3739–3745
Roche T (2011) Weather underground. Weather Underground,
Inc. http://www.weatherunderground.com. Accessed 6/5/
2011
Royo AA, Carson WP (2006) On the formation of dense under-
story layers in forests worldwide: consequences and impli-
cations for forest dynamics, biodiversity, and succession.
Mapping buckthorn in the Oak Openings
123

Can J For Res-Rev Can Rech For 36(6):1345–1362. doi:
10.1139/x06-025
Skinner L, Blossey B, Nuzzo V, Maerz J, Davalos A, Gerber E,
Hinz H, Landis D, Davis A, Schemske D, Katovich E,
Becker R, Ragsdale D, Evans J, Knight K, Stewart J,
Heneghan L, Wyckoff P, Jansen R, Patten R, Moriarty J,
Gassmann A, Veldhuizen M (2005) Biology, ecology, and
management of garlic mustard (Alliaria petiolata) and
European buckthorn (Rhamnus cathartica). In: Skinner LC
(ed) Symposium on the biology, ecology, and management
of garlic mustard (Alliaria petiolata) and European buck-
thorn (Rhamnus cathartica). St. Paul, Minnesota. USDA
Forest Service, p 71
Stohlgren TJ, Falkner M, Schell L (1995) A modified-Whittaker
nested vegetation sampling method. Plant Ecol 117(2):
113–121
Stohlgren TJ, Barnett DT, Kartesz JT (2003) The rich get richer:
patterns of plant invasions in the United States. Front Ecol
Environ 1(1):11–14
Torrey J (1824) A flora of the Northern and Middle Sections of
the United States. Swords
Tuanmu MN, Vina A, Bearer S, Xu WH, Ouyang ZY, Zhang
HM, Liu JG (2010) Mapping understory vegetation using
phenological characteristics derived from remotely sensed
data. Remote Sens Environ 114(8):1833–1844. doi:
10.1016/j.rse.2010.03.008
Underwood E, Ustin S, DiPietro D (2003) Mapping nonnative
plants using hyperspectral imagery. Remote Sens Environ
86(2):150–161. doi:10.1016/s0034-4257(03)00096-8
Venette R, Ragsdale D (2004) Assessing the invasion by soy-
bean aphid (Homoptera: Aphididae): where will it end?
Ann Entomol Soc Am 97(2):219–226
Voegtlin DJ, O’Neil RJ, Graves WR, Lagos D, Yoo HJS (2005)
Potential winter hosts of soybean aphid. Ann Entomol Soc
Am 98(5):690–693
Woods KD (1993) Effects of Invasion by Lonicera-Tatarica L
on Herbs and Tree Seedlings in 4 New-England Forests.
Am Midl Nat 130(1):62–74
Zouhar K, Smith JK, Sutherland S, Brooks ML (2008) Wildland
fire in ecosystems: fire and nonnative invasive plants, vol 6.
Rocky Mountain Research Station, Ogden
R. H. Becker et al.
123