sensory receptors. range from simple neurons to complex sense organs types: chemoreceptors,...
Post on 22-Dec-2015
220 views
TRANSCRIPT

Sensory Receptors

Sensory Receptors
Range from simple neurons to complex sense organs
Types: chemoreceptors, mechanoreceptors, photoreceptors, electroreceptors, magnetoreceptors, thermoreceptors
All transduce incoming stimuli into changes in membrane potential
Figure 7.1

Sensory Receptors
Figure 7.1

Classification of Sensory Receptors
Based on stimulus location Telereceptors – detect distant stimuli, e.g.,
vision and hearing Exteroceptors – detect stimuli on the outside
of the body, e.g., pressure and temperature Interoceptors – detect stimuli inside the body,
e.g., blood pressure and blood oxygen

Classification of Sensory Receptors
Based on type of stimuli the receptors can detect (stimulus modality) Chemoreceptors – chemicals, e.g., smell and taste Mechanoreceptors – pressure and movement, e.g.,
touch, hearing, balance, blood pressure Photoreceptors – light, e.g., vision; detect photons Electroreceptors – electrical fields Magnetoreceptors – magnetic fields Thermoreceptors - temperature

Receptors and stimulus
Location: Can distinguish the location of the stimulus (touch, light or odour)
Duration: Determine length of stimulus by responding to the stimulus for the duration of the stimulus.
Intensity: Increase in action potential frequency or increase in neurotransmitter release.

Sensitivity to Multiple Modalities
• Adequate stimulus – preferred or most sensitive stimulus modality
• Many receptors can also be excited by other stimuli, if sufficiently large, e.g., pressure on eyelid perceive bright light
• Polymodal receptors – naturally sensitive to more than one stimulus modality, e.g., ampullae of Lorenzini in sharks
• Nociceptors – sensitive to strong stimuli, e.g., pain; many are polymodal receptors

Stimulus Encoding
All stimuli are ultimately converted into action potentials in the primary afferent neurons
How can organisms differentiate among stimuli or detect the strength of the signal?
Sensory receptors must encode four types of information Stimulus modality Stimulus location Stimulus intensity Stimulus duration

Dynamic Range
• Action potentials code stimulus intensity through changes in frequency, e.g., strong stimuli high frequency
• Dynamic range – range of intensities for which receptors can encode stimuli
• Threshold detection – weakest stimulus that produces a response in a receptor 50% of the time
• Saturation – top of the dynamic range; all available proteins have been stimulated
Figure 7.4a

Range Fractionation
Relationships between stimulus intensity and AP frequency• Linear across large range of
intensities: large change in stimulus causes a small change in AP frequency large dynamic range, poor sensory discrimination
• Linear across small range of intensities: small change in stimulus causes a large change in AP frequency small dynamic range, high sensory discrimination
Range fractionation – groups of receptors work together to increase dynamic range without decreasing sensory discrimination
Figure 7.4b-c

Tonic and Phasic Receptors
Two classes of receptors that encode stimulus duration• Phasic – produce APs only at the beginning or end of the
stimulus encode changes in stimulus, but not stimulus duration
• Tonic – produce APs as long as the stimulus continues• Receptor adaptation – AP frequency decreases if stimulus
intensity is maintained at the same level

Tonic and Phasic Receptors, Cont.
Figure 7.5

Pain
• Pain and itching are mediated by Nocireceptors• Itch comes form Nocireceptors in the skin. Higher
pathways for itch are not well understood• Pain is s subjective perception

Chemoreception
• Most cells can sense incoming chemical signals• Animals have many types of chemoreceptors• Multicellular organisms typically use taste and smell• Olfaction – sense of smell
• Detection of chemicals carried in air• Gustation – sense of taste
• Detection of chemicals emitted from ingested food• Distinct due to structural criteria• Performed by different sense organs• Use different signal transduction mechanisms• Are processed in different integrating centers

The Olfactory System
Evolved independently in vertebrates and insects
Vertebrate olfactory system
• Can distinguish thousands of odorants
• Located in the roof of the nasal cavity
• Mucus layer to moisten olfactory epithelium
• Odorant binding proteins – allow lipophilic odorants to dissolve in mucus
• Receptor cells are bipolar neurons and are covered in cilia
• Odorant receptor proteins are located in the cilia

Odorant Receptors are G Proteins
• Each olfactory neuron expresses only one odorant receptor protein
• Each odorant receptor can recognize more than one odorant
Figure 7.7

Pheromones
Vomeronasal organ – detects pheromones
Structurally and molecularly distinct from the primary olfactory epithelium
• Location• Base of nasal cavity near the
septum in mammals• Palate in reptiles
• Transduction• Activates a phospholipase C-
based signal transduction system; adenylate cyclase-cAMP in other olfactory receptors
Figure 7.8

Taste Buds in Vertebrates
Group of taste receptor cells
Located on tongue, soft palate, larynx, and esophagus; external surface of the body in some fish

Taste Buds in Vertebrates
50 to 150 taste cells Epithelial cells that have apical and basal sides and joined by tight
junctions Life span of 10-14 days Basal stem cells divide to regenerate taste cells Microvilli on its apical surface that project into the mucus of the tongue Taste receptor proteins are found in the microvilli Chemicals are soluble and diffuse to the bind to their receptors Different cells in the same bud can detect NaCl, sucrose, H+ and quinine
(bitter) Taste cell forms a chemical synapse with a sensory neuron that projects
to the brain from the tongue

Taste buds and peripheral innervation
Figure 7.11c-d

A generic taste cell.
Apical surface: both channels and G-protein-coupled receptors that are activated by chemical stimuli
Basolateral surface: voltage-gated Na+, K+, and Ca2+ channels, as well as all the machinery for synaptic transmission mediated by serotonin
The increase in intracellular Ca2+ is either by the activation of voltage-gated Ca2+ channels or via the release from intracellular stores causes synaptic vesicles to fuse and release their transmitter onto receptors on primary sensory neurons
Each cell contains the standard complement of neuronal proteins including Na+/K+ ATPase at the basal level, voltage-gated Na+ and Ca2+ channels, leak K+ channel

A generic taste cell…cont.
The response to the chemical is mediated by the expression of receptors for that chemical in the microvilli
The response is a depolarization of the cell sometimes enough to generate an action potential
The signaling of the cell to the sensory neuron depends on a sufficient depolarization to open the voltage-gated Ca2+ channels necessary for vesicle fusion and neurotransmitter release.

Transduction mechanisms
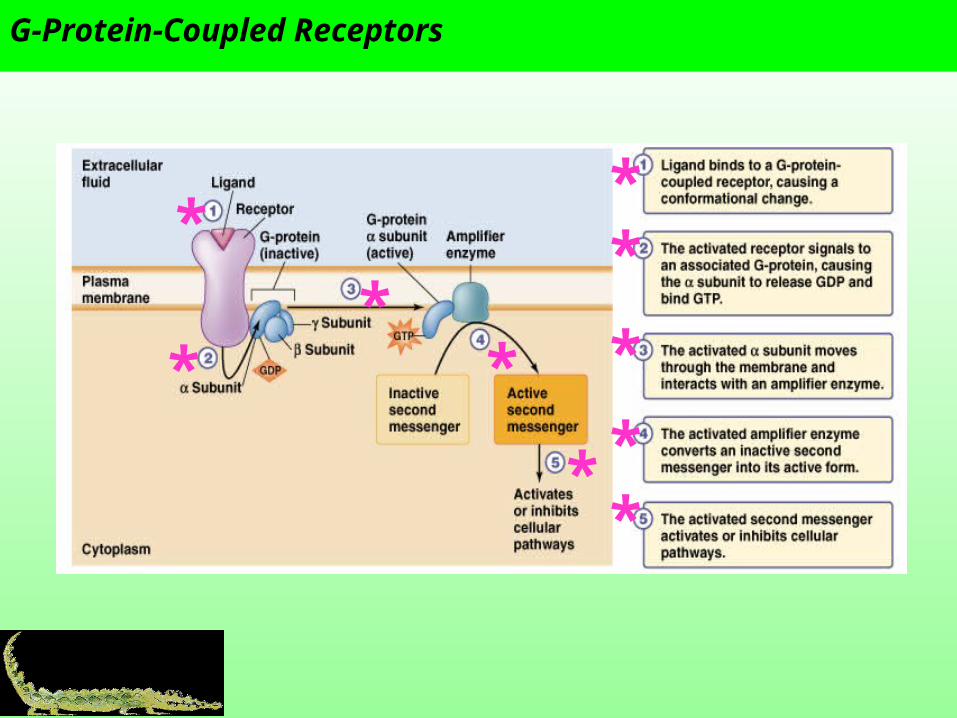
G-Protein-Coupled Receptors
*
**
*
*
**
*
**

G-protein and adenylate cyclase
* *
**
*
*
* ****
**
*

The inositol-phospholipid signaling pathway
You don’t have to memorize this weeeeeeebut be aware of it and know which taste is transmitted using this pathway ie bitter

Salt taste
The Na+ enters into the cell through the passive amiloride-sensitive Na+ channel
These proteins are found in frog skin and kidney Amiloride will block Na+ salt taste reception Entry of Na+ into the cell of course causes the
cell to depolarize Need a large concentration of Na+ to trigger a
sufficient depolarization to signal to the post-synaptic sensory neuron

Salt taste
*
*
*

Sour taste
Taste response produced by acids, excess protons (H+). These positive ions enter the cell through a H+, cation specific ion channel and in turn depolarize the cell to threshold for an action potential.

Sour taste
***
*

Sweet taste There are specific membrane receptors for different sweeteners and sugars These receptors are not ligand gated ion channels but rather are
metabotropic receptors These receptors belong to the family of seven transmembrane domain
proteins that are linked to signaling cascades through G proteins. In mammals a combination of the T1R2/T1R3 receptors have a response to sugars and sweeteners
These receptors stimulate a G protein (Gp) which in this case activates phosopholipase C (PLC)
PLC breaks down PIP2 (phosphatidylinositol 4,5-bisphosphate) into IP3 (inositol triphosphosphate) and DAG
IP3 will bind to and activate a ion channel (TRP channel called TRPM5) which allows Ca2+ to influx into the cell
This pathway leads to a depolarization and threshold is reached to trigger an action potential
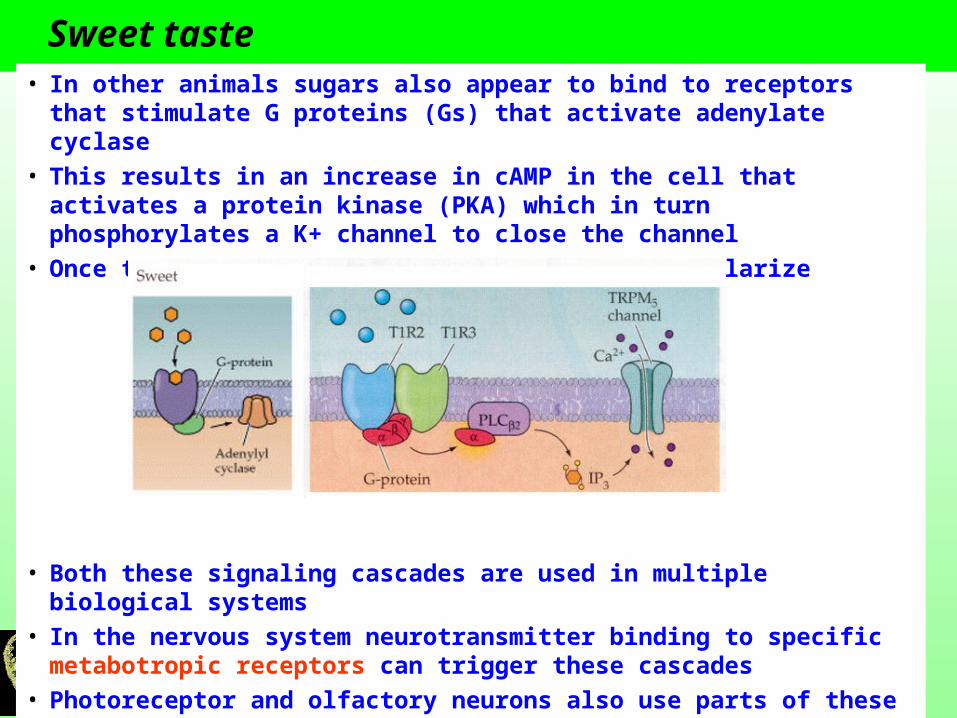
Sweet taste • In other animals sugars also appear to bind to receptors that stimulate G
proteins (Gs) that activate adenylate cyclase
• This results in an increase in cAMP in the cell that activates a protein kinase (PKA) which in turn phosphorylates a K+ channel to close the channel
• Once the K+ channel is close the cell will depolarize
• Both these signaling cascades are used in multiple biological systems
• In the nervous system neurotransmitter binding to specific metabotropic receptors can trigger these cascades
• Photoreceptor and olfactory neurons also use parts of these cascades for their sensory transduction

Sweet taste
**
**
*
*
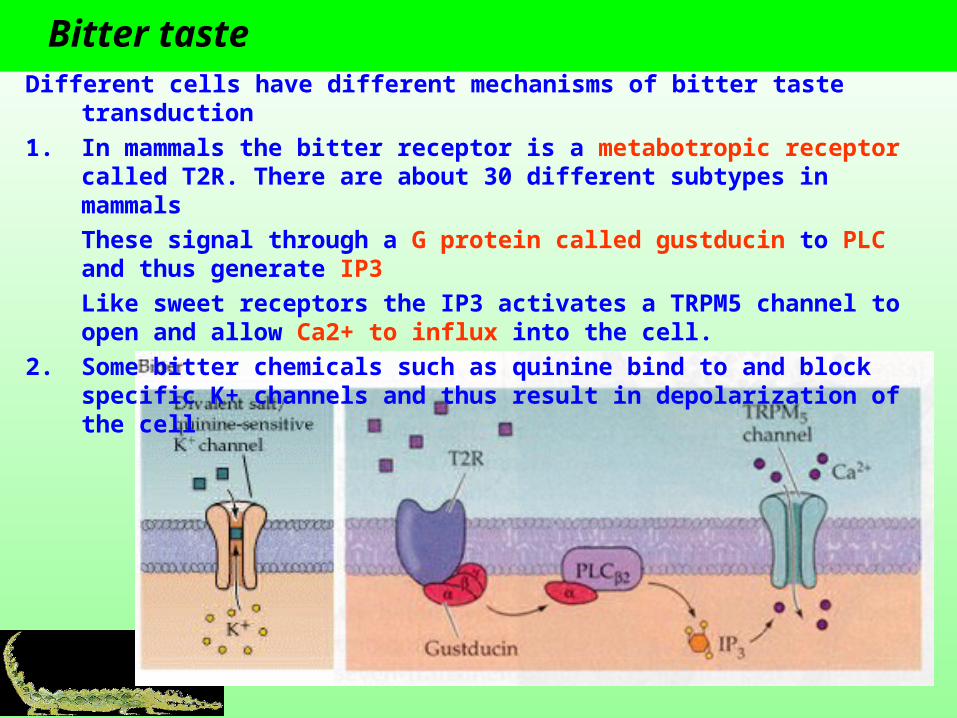
Bitter taste Different cells have different mechanisms of bitter taste transduction
1. In mammals the bitter receptor is a metabotropic receptor called T2R. There are about 30 different subtypes in mammals
These signal through a G protein called gustducin to PLC and thus generate IP3
Like sweet receptors the IP3 activates a TRPM5 channel to open and allow Ca2+ to influx into the cell.
2. Some bitter chemicals such as quinine bind to and block specific K+ channels and thus result in depolarization of the cell

Bitter taste
**
**
*

Amino acid taste cellsIn some animals (catfish) there are a high number of amino acid taste cells
There appears to be multiple ways that animals respond to amino aicds
1. In fish and other amphibians, amino acids such as L-arginine and L-proline bind to specific receptors which are ligand gated ion channels
2. In mammals there are taste cells that respond to L-glutamate. In these cells L-glutamate activates a metabotropic receptor glutamate receptor linked to a G protein. Glutamate binds to many different metabotropic receptors and in taste cells it is the mGluR4 that is responsible for the taste transduction
3. In mammals there are also two metabotropic receptors T1R1/T1R3 that combine to respond to the standard 20 amino acids. This combination signals through G protein activation of PLC and the generation of IP3 and the activation of the TRPM5 channel.