sequential utilization of substrates by pseudomonas putida csv86: signatures of intermediate...
TRANSCRIPT

ARTICLE IN PRESS
Microbiological Research 164 (2009) 429—437
0944-5013/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
1Equal autho
www.elsevier.de/micres
Sequential utilization of substrates byPseudomonas putida CSV86: Signatures ofintermediate metabolites and onlinemeasurements
Aditya Basua,1, Debasish Dasb,1, Prashant Bapatb,Pramod P. Wangikarb, Prashant S. Phalea,�
aBiotechnology Group, School of Biosciences and Bioengineering, Indian Institute of Technology-Bombay, Powai,Mumbai 400 076, IndiabDepartment of Chemical Engineering, Indian Institute of Technology-Bombay, Mumbai 400 076, India
Received 12 October 2006; received in revised form 12 December 2006; accepted 16 February 2007
KEYWORDSPseudomonas putidaCSV86;Preferential utiliza-tion;Aromatic com-pounds;Bioreactor;Online analysis
ee front matter & 2007micres.2007.02.008
ing author. Fax: +91 22 2ess: [email protected] contribution
SummaryPseudomonas putida CSV86 preferentially utilizes aromatics over glucose and co-metabolizes them with organic acids. On aromatics plus glucose, CSV86 utilizedaromatics first with concomitant appearance of transient metabolites such assalicylate, benzaldehyde and benzoate. Citrate was the main extracellularmetabolite observed during glucose uptake. The strain showed simultaneousutilization of organic acids and aromatic compounds. Based on the metaboliteanalysis and growth profiles, we hypothesize that the repression of glucoseutilization could be due to organic acid intermediates generated from aromaticcompound metabolism. The online measurements indicate the instantaneousmetabolic state of the culture. For example, the CO2 evolution and agitation speedshow peak values during the two growth phases in the diauxic growth while dissolvedoxygen values show decrease at the corresponding durations. These measurementscorrelated well with the offline measurements but provided a better time resolutionof the process.& 2007 Elsevier GmbH. All rights reserved.
Elsevier GmbH. All rights rese
572 3486.(P.S. Phale).
Introduction
A large number of pollutants present in theenvironment are aromatic compounds which arerecalcitrant, toxic and carcinogenic (Alexander,
rved.

ARTICLE IN PRESS
A. Basu et al.430
1981). The effective and economical way to removethem is microbial degradation, ‘bioremediation’. Innature, there exists a mixture of simple (e.g.glucose) and complex (e.g. aromatics) carboncompounds which can act as potential growthsubstrates for microorganisms. Under such circum-stances, the simple carbon source is utilized firstfollowed by the complex carbon source, hence thegrowth response is diauxic or biphasic, reflecting asequential rather than simultaneous utilization ofthe carbon sources (Collier et al., 1996). Thisphenomenon is called carbon catabolite repression(CCR) and has been reported in many organisms.The major hindrance to use microorganisms forbioremediation is their preference for simplercarbon sources over complex aromatic compounds.The genus Pseudomonas contains the most versatileorganisms with the ability to mineralize variouscomplex aromatic compounds. However, very littleis known about the sequence of utilization ofsubstrates from a mixture of carbon sources andits regulatory mechanism. It has been reported thatglucose uptake and its utilization in Pseudomonas isrepressed by organic acids such as acetate, citrate,succinate, malate and fumarate (Tiwari and Camp-bell, 1969; Hylemon and Phibbs, 1972; Robertset al., 1973; Midgley and Dawes, 1973; Ng andDawes, 1973; Mukkada et al., 1973). In Pseudomo-nas putida, benzylalcohol-metabolizing enzymeshave been shown to be repressed by acetate(Worsey and Williams, 1975), glucose (Holtelet al., 1994) and succinate (Duetz et al., 1994).Similarly, utilization of phenyl acetic acid inP. putida U and degradation of phenanthrene inP. putida has been reported to be suppressed byglucose (Schleissner et al., 1994; Rentz et al.,2004).
In E. coli, glucose is transported by phosphoe-nolpyruvate (PEP)-dependent transport system(PTS), which drives a protein phosphorylationcascade. This cascade affects c-AMP levels bymodulating the adenylate cyclase activity thusexhibiting CCR (Collier et al., 1996; Stulke andHillen, 1999). Unlike E. coli, very little is knownabout the mechanism of repression of glucose inPseudomonas. Pseudomonads lack PEP-PTS systemfor the glucose transport and c-AMP does not affectthe repression (Siegel et al., 1977; Phillips andMulfinger, 1981). Glucose is transported usingglucose transporter (Stinson et al., 1976; Wylieand Worobec, 1995) by active transport (Framptonand Wood, 1961; Phibbs, and Eagon, 1970; Boos,1974).
P. putida CSV86 utilizes naphthalene, methyl-naphthalene, benzylalcohol, salicylate and benzo-ate as the sole source of carbon and energy.
Naphthalene metabolism occurs via salicylate andcatechol, while benzyl alcohol degradation occursthrough benzoate and catechol (Mahajan et al.,1994; Basu et al., 2003). This strain utilizes glucoseby intracellular direct phosphorylative pathway,does not produce or utilize gluconate or 2-ketogluconate and has inducible, active glucosetransport mechanism (Basu and Phale, 2006). Whengrown on dual carbon source, the strain prefersaromatic compounds over glucose, described asreverse CCR (Basu et al., 2006). Strain CSV86 co-metabolizes aromatic compound plus organic acidand prefers organic acids over glucose. It was ofinterest to monitor the uptake of extracellularsubstrates, and secretion and subsequent uptake ofmetabolites. We present a detailed analysis ofextracellular metabolites, diauxic growth patternsand their qualitative correlation with various onlinemeasurements such as pH, dissolved oxygen, stirrerspeed and CO2 evolution rate. Further insight in theinstantaneous metabolic state of the culture wasobtained by measuring the CO2 evolution rate andthe oxygen demand as measured via the stirrerspeed and the dissolved oxygen levels. Resultsobtained from batch fermentation strengthen ourearlier unique observation that P. putida CSV86preferentially utilizes aromatic compounds overglucose and co-metabolizes aromatics and organicacids. The trends in the online measurementscorrelate well with the offline measurements.
Materials and methods
Organism, culture medium and growthconditions
P. putida CSV86 was grown on 150ml minimal saltmedium [MSM (Basu et al., 2003)] in 500mlcapacity baffled Erlenmeyer flasks at 30 1C on arotary shaker (200 rpm). The medium was supple-mented aseptically with appropriate aromaticcompound (0.1%) or glucose or organic acid(0.25%) either alone or in combination as indicated.
Bioreactor and cultivation condition
Batch cultivations were performed in a Biostat B5bioreactor (B. Brown, Germany) with a workingvolume of 3 L. The temperature was maintained at30 1C and dissolved oxygen was maintained atminimum 40% via cascade control with agitator.The aeration rate was kept at 3 Lmin�1 and wascontrolled by a mass flow controller (B Brown,Germany). The concentration of O2 and CO2 ðqCO2
Þ
in the exit gas was measured by infrared spectroscopy

ARTICLE IN PRESS
Sequential utilization of substrates by Pseudomonas putida CSV86 431
and paramagnetic analysis, respectively (analyzerBINOS1002M with sample-conditioning unit, Rose-mount analytical, Germany). Growth was moni-tored at 540 nm using spectrophotometer (JascoV750, Japan).
The batch duration varied from 25 to 40 hdepending on the substrates. The batch recordingwas terminated when the carbon substrates andtransient metabolites were exhausted from themedia. Of the main substrates used, glucose,aromatic compounds and organic acids were fullysubstitutable as growth can occur on any one as thesole source of carbon.
A total of five process variables were measuredonline and recorded every 5min. These includevent CO2 (%), vent O2 (%), pH, dissolved O2 (%) andstirring rate (rpm). Note that the measurementsindicate some physical phenomenon related to themetabolic state of the culture. For example, thevalues of vent CO2 (%) were used to compute theinstantaneous rates of CO2 production ðqCO2
mmolL�1 h�1). The peaks in the qCO2
profile maycorrespond to an increased metabolic activity ofthe culture compared to the rest of the batch. Thedissolved O2 concentration was maintained at 40%of saturation and hence its value typically variesbetween 40–100% of saturation. The stirring rate isa control variable, which is varied between120–1200 rpm in order to maintain dissolved O2 at40%. A higher oxygen demand by the fermentationculture usually leads to a higher stirring rate. Theactual stirring rate is dependent on the rate ofoxygen consumption and the hydrodynamic para-meters such as oxygen partial pressure, viscosity ofthe broth, etc. Similarly, the trends of pH valuesmay be qualitative indicators of phenomena such asorganic acid production or consumption. The pHwas maintained at a set point of 7.2 by addition ofalkali. Thus, pH excursions below the set pointwere not permitted as the organism was found tobe sensitive to such pH changes (data not shown).Thus, the online measurements contain informationon the overall state of the culture. Developmentof a quantitative model to incorporate thesemeasurements would be beyond the scope ofthis study. However, we point out some quali-tative correlations between the offline and onlinemeasurements.
Analytical techniques
Quantitation of substrates and various intermedi-ate metabolites from the cell-free culture mediumwas performed by HPLC using HP-Aminex-87-Hcolumn (Biorad). The column temperature was
maintained at 65 1C. A mobile phase of sulfuricacid (5mM) with flow rate of 0.6mlmin�1 was usedto resolve glucose and organic acids. Glucose(retention time, rt, 10.67min) was detected by RIdetector while pyruvate (rt, 10.35min), citrate (rt,9.02min), succinate (rt, 12.43min) and salicylate(rt, 15.90min) by UV detector at 210 nm. Benzylalcohol (rt, 7.71min), benzaldehyde (rt, 12.43min)and benzoate (rt, 5.60min) were detected by UVdetector (210 nm) using Symmetrys C18 5 mmcolumn (WATERSTM) in HPLC (WATERSTM 1525). Themobile phase used was, water: acetonitrile: phos-phoric acid (6.99:3:0.01, v/v/v) with a flow rate of0.7mlmin�1 (Buhler et al., 2001).
Results
Here, we present analysis of substrate uptake,extracellular metabolites and growth for severalbatch runs involving P. putida CSV86. The batchesdiffer in their initial substrate combination andin turn in the profiles of substrate utilizationand production of biomass and extracellular meta-bolites.
Growth profiles on aromatic compound plusglucose
When grown on benzyl alcohol plus glucose,P. putida CSV86 showed a diauxic growth profilewith the first growth phase of 8 h followed by adistinct lag phase of �7 h and second log phasestarting at �17 h (Fig. 1, Panel A). Quantitativeanalysis of various transient metabolites from theculture medium was carried out via HPLC. In thefirst growth phase, benzyl alcohol was consumedwith concomitant appearance of benzaldehyde andbenzoate, which were subsequently utilized beforecells entered into the second log phase. Glucoseconcentration remained unchanged till �16 h anddeclined rapidly thereafter as the culture enteredinto the second log phase. During the second logphase, citrate accumulated in the medium (Fig. 1,Panel A). The first peak value of CO2 in the exit gasstream was observed between 7 and 9 h followed bythe second peak between 17 and 20 h, withconcomitant drop in DO, increase in stirrer speedas well as fall in the pH (Fig. 1, Panel B). Thesepeaks indicate a higher metabolic activity com-pared to the rest of the batch. Interestingly, thepeaks correspond well with the two exponentialgrowth phases in the diauxic growth. Note that theresults shown in Fig. 1 correspond to a batch runwhere the inoculum was grown on benzyl alcohol.However, the pattern of substrate uptake remained

ARTICLE IN PRESS
Bio
mass (
g.L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
Glu
cose (
g.L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
Benzyl alc
ohol (g
.L-1
); B
enzoic
acid
(g.L
-1)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Benzald
ehyde (
g.L
-1)
0.00
0.04
0.08
0.12
0.16
0.20
Citra
te (
g.L
-1)
0.0
0.1
0.2
0.3
0.4
Age (h)
0 5 10 15 20 25 30
qC
O2 (
mm
ole
L-1
h-1
)
0
2
4
6
8
10
12
14
16
18
DO
(%
)
40
60
80
100
120
140
Agitato
r x 1
00 (
RP
M)
1.2
2.2
3.2
4.2
5.2
pH
7.0
7.2
7.4
7.6
A
B
Figure 1. Growth profile of Pseudomonas putida CSV86 on benzyl alcohol (0.1%) plus glucose (0.25%). Benzyl alcohol-grown cells were used as inoculum. Panel A: Represents time-dependent changes in the biomass (K), glucose (J),benzyl alcohol (m), benzaldehyde (’), benzoate (n) and citrate (E). Panel B: Represents the online data, vent CO2 (—),DO (– –), pH ( � � � ) and agitator speed (– � � –).
A. Basu et al.432
unchanged with glucose grown inoculum (data notshown).
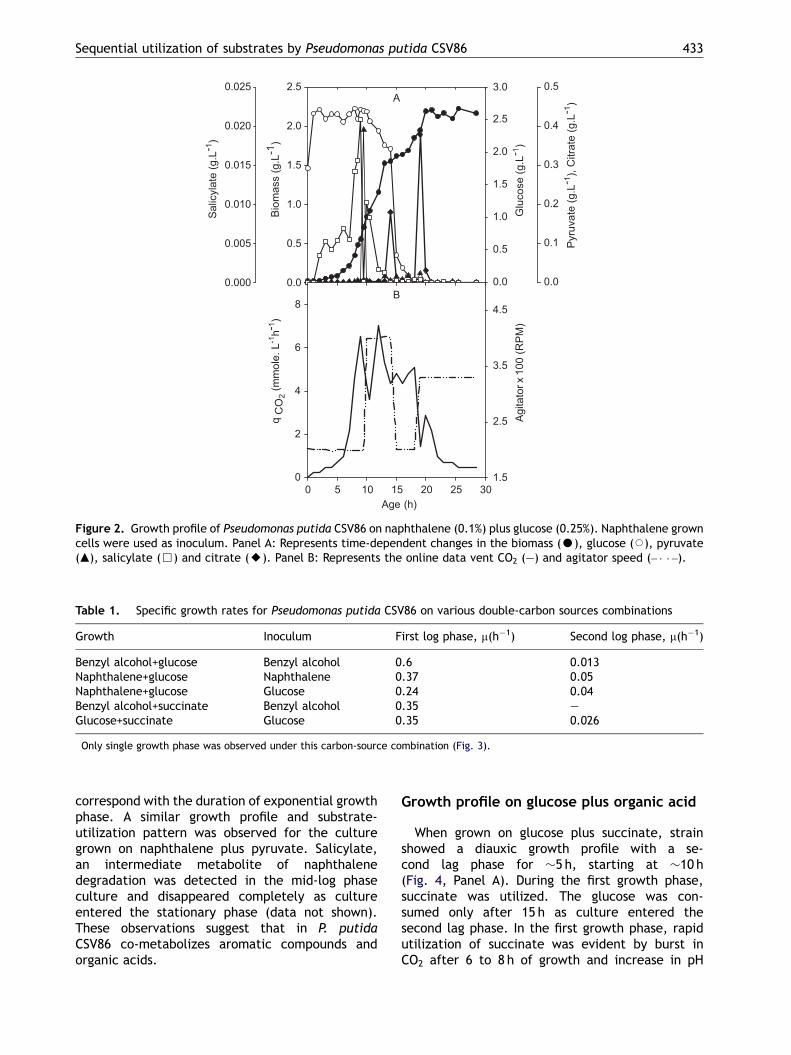
When grown on naphthalene plus glucose, CSV86showed a similar biphasic growth profile withcorresponding substrate utilization profile (Fig. 2,Panel A). Culture showed distinct first growthphase up to �11 h followed by a very short (�1 h)second lag phase and second log phase startingaround �13 h. During the first log phase, spentmedium showed appearance of salicylate, whichwas consumed before the culture entered into thesecond log phase. Glucose concentration remainedunchanged in the first growth phase and thendeclined rapidly as cells entered the second logphase (Fig. 2, Panel A). In the first phase, the burstin CO2 evolution could be attributed to the highrespiration rates as well as to the CO2 evolutiondue to the formation of catechol from salicylate(Fig. 2, Panel B). The sequence of substrateutilization remained unchanged regardless ofwhether the inoculum was grown on glucose orthe respective aromatic compound (data notshown). The specific growth rates under various
conditions are shown in Table 1. The specific growthrate of CSV86 when grown on benzyl alcohol,naphthalene or succinate (first log phase) wassignificantly higher than that on glucose (secondlog phase). A similar trend was observed forbatches where the inoculum was grown on glucose(Table 1).
Growth profiles on aromatic compound plusorganic acid
On benzyl alcohol plus succinate, strain CSV86showed a single growth phase (Fig. 3, Panel A).Both succinate and benzyl alcohol were con-sumed simultaneously. The intermediate me-tabolites, benzaldehyde and benzoate producedand consumed before the culture entered thestationary phase (�16 h). A significant burst inCO2 was observed between 9 and 13 h concomi-tant with an increase in pH (7.4–8.6), fall inDO and increase in stirrer speed (Fig. 3, Panel B).These peaks (and dips) in the online measurements
ARTICLE IN PRESS
Bio
mass (
g.L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
Glu
cose (
g.L
- 1)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Pyru
vate
(g.L
-1),
Citra
te (
g.L
-1)
0.0
0.1
0.2
0.3
0.4
0.5
Salic
yla
te (
g.L
-1)
0.000
0.005
0.010
0.015
0.020
0.025
Age (h)
0 5 10 15 20 25 30
q C
O2 (
mm
ole
. L
-1h-1
)
0
2
4
6
8
Agitato
rx
100 (
RP
M)
1.5
2.5
3.5
4.5
A
B
Figure 2. Growth profile of Pseudomonas putida CSV86 on naphthalene (0.1%) plus glucose (0.25%). Naphthalene growncells were used as inoculum. Panel A: Represents time-dependent changes in the biomass (K), glucose (J), pyruvate(m), salicylate (&) and citrate (E). Panel B: Represents the online data vent CO2 (—) and agitator speed (– � � –).
Table 1. Specific growth rates for Pseudomonas putida CSV86 on various double-carbon sources combinations
Growth Inoculum First log phase, m(h�1) Second log phase, m(h�1)
Benzyl alcohol+glucose Benzyl alcohol 0.6 0.013Naphthalene+glucose Naphthalene 0.37 0.05Naphthalene+glucose Glucose 0.24 0.04Benzyl alcohol+succinate Benzyl alcohol 0.35 —�
Glucose+succinate Glucose 0.35 0.026
�Only single growth phase was observed under this carbon-source combination (Fig. 3).
Sequential utilization of substrates by Pseudomonas putida CSV86 433
correspond with the duration of exponential growthphase. A similar growth profile and substrate-utilization pattern was observed for the culturegrown on naphthalene plus pyruvate. Salicylate,an intermediate metabolite of naphthalenedegradation was detected in the mid-log phaseculture and disappeared completely as cultureentered the stationary phase (data not shown).These observations suggest that in P. putidaCSV86 co-metabolizes aromatic compounds andorganic acids.
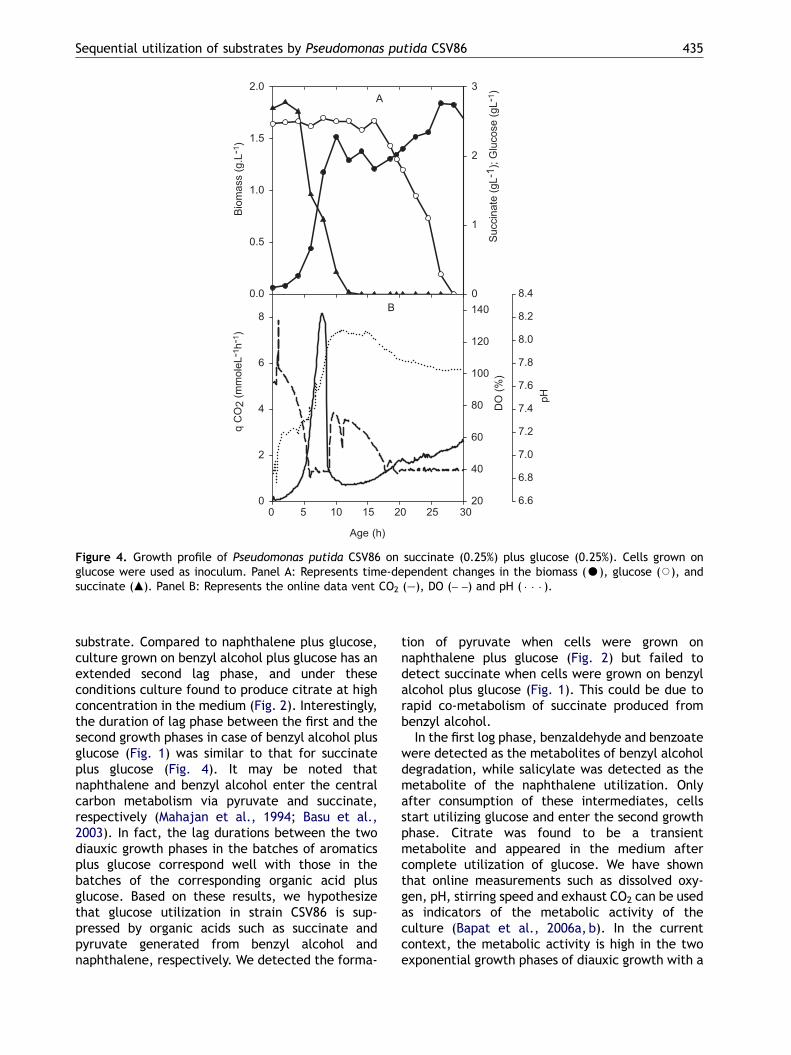
Growth profile on glucose plus organic acid
When grown on glucose plus succinate, strainshowed a diauxic growth profile with a se-cond lag phase for �5 h, starting at �10 h(Fig. 4, Panel A). During the first growth phase,succinate was utilized. The glucose was con-sumed only after 15 h as culture entered thesecond lag phase. In the first growth phase, rapidutilization of succinate was evident by burst inCO2 after 6 to 8 h of growth and increase in pH

ARTICLE IN PRESS
Bio
mass (
g.L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
Succin
ate
(g.L
-1)
0.0
0.5
1.0
1.5
2.0
2.5
Benzyl alc
ohol(g.L
-1)
0.0
0.2
0.4
0.6
0.8
Benzald
ehyde (
g.L
-1)
0.00
0.02
0.04
0.06
0.08
Benzoic
acid
(g.L
-1)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Age (h)
0 5 10 15 20 25
q C
O2 (
mm
ole
L-1
h-1
)
0
5
10
15
20
25
DO
(%
)
20
40
60
80
100
120
pH
7.0
7.6
8.2
8.8
Agitato
r x100 (
RP
M)
1.1
2.1
3.1
4.1
5.1
A
B
Figure 3. Growth profile of Pseudomonas putida CSV86 on benzyl alcohol (0.1%) plus succinate (0.25%). Cells grown onbenzyl alcohol were used as inoculum. Panel A: Represents time-dependent changes in the biomass (K), succinate (J),benzyl alcohol (m), benzaldehyde (’) and benzoate (n). Panel B: Represents the online data vent CO2 (—), DO (– –), pH( � � � ) and agitator speed (– � � –).
A. Basu et al.434
from 7.2 to 8.1 with concomitant fall in DO (Fig. 4,Panel B).
Discussion
P. putida CSV86 utilizes naphthalene via salicy-late and catechol ‘meta-cleavage’ pathway (Maha-jan et al., 1994) and benzyl alcohol viabenzaldehyde, benzoate and catechol ‘ortho-clea-vage’ pathway (Basu et al., 2003). The strainutilizes organic acids and aromatic compoundsmore efficiently and faster than glucose (Basu andPhale, 2006). The enzyme-induction studies andgrowth profiles at the shake flask level suggestedthat CSV86 preferentially utilizes aromatic com-pounds over glucose. Even at low concentration ofnaphthalene (0.025% naphthalene+0.25% glucose),aromatic compound was used preferentially overglucose (Basu et al., 2006). In the present study wemonitored growth profiles with detailed analysis ofsubstrate utilization, appearance of transient ex-tracellular metabolites and online parameters such
as DO, CO2, pH and stirrer speed profiles for batchfermentation runs. We argued that the offlineanalyses of concentrations in the liquid phaseprovided direct information on the substrate-uptake pattern, while the online analyses providedinformation on the metabolic activity of the cultureat a time resolution that is far better than that ofoffline analyses.
On double-carbon source such as aromaticcompound plus glucose, CSV86 showed diauxicgrowth pattern with utilization of aromatic com-pounds in the first growth phase and a distinctsecond lag phase before the uptake of glucosecommenced (Figs. 1 and 2). Irrespective of thearomatic carbon source, glucose was used in thesecond growth phase with the specific growth rateon glucose being lower than that on aromatics. Thiswas true regardless of whether the inoculum wasgrown on glucose or respective aromatic compound(data not shown for glucose-grown inoculum),implying that there may be a genetic regulationinvolved in determining the sequence of substrateuptake rather than a simple adaptation to the
ARTICLE IN PRESS
Bio
ma
ss (
g.L
- 1)
0.0
0.5
1.0
1.5
2.0
Su
ccin
ate
(g
L-1
); G
luco
se
(g
L- 1
)
0
1
2
3
Age (h)
0 5 10 15 20 25 30
q C
O2
(m
mo
leL
- 1h
- 1)
0
2
4
6
8
DO
(%
)20
40
60
80
100
120
140
pH
6.6
6.8
7.0
7.2
7.4
7.6
7.8
8.0
8.2
8.4
A
B
Figure 4. Growth profile of Pseudomonas putida CSV86 on succinate (0.25%) plus glucose (0.25%). Cells grown onglucose were used as inoculum. Panel A: Represents time-dependent changes in the biomass (K), glucose (J), andsuccinate (m). Panel B: Represents the online data vent CO2 (—), DO (– –) and pH ( � � � ).
Sequential utilization of substrates by Pseudomonas putida CSV86 435
substrate. Compared to naphthalene plus glucose,culture grown on benzyl alcohol plus glucose has anextended second lag phase, and under theseconditions culture found to produce citrate at highconcentration in the medium (Fig. 2). Interestingly,the duration of lag phase between the first and thesecond growth phases in case of benzyl alcohol plusglucose (Fig. 1) was similar to that for succinateplus glucose (Fig. 4). It may be noted thatnaphthalene and benzyl alcohol enter the centralcarbon metabolism via pyruvate and succinate,respectively (Mahajan et al., 1994; Basu et al.,2003). In fact, the lag durations between the twodiauxic growth phases in the batches of aromaticsplus glucose correspond well with those in thebatches of the corresponding organic acid plusglucose. Based on these results, we hypothesizethat glucose utilization in strain CSV86 is sup-pressed by organic acids such as succinate andpyruvate generated from benzyl alcohol andnaphthalene, respectively. We detected the forma-
tion of pyruvate when cells were grown onnaphthalene plus glucose (Fig. 2) but failed todetect succinate when cells were grown on benzylalcohol plus glucose (Fig. 1). This could be due torapid co-metabolism of succinate produced frombenzyl alcohol.
In the first log phase, benzaldehyde and benzoatewere detected as the metabolites of benzyl alcoholdegradation, while salicylate was detected as themetabolite of the naphthalene utilization. Onlyafter consumption of these intermediates, cellsstart utilizing glucose and enter the second growthphase. Citrate was found to be a transientmetabolite and appeared in the medium aftercomplete utilization of glucose. We have shownthat online measurements such as dissolved oxy-gen, pH, stirring speed and exhaust CO2 can be usedas indicators of the metabolic activity of theculture (Bapat et al., 2006a, b). In the currentcontext, the metabolic activity is high in the twoexponential growth phases of diauxic growth with a

ARTICLE IN PRESS
A. Basu et al.436
basal metabolic activity during the rest of thebatch. This is reflected nicely in terms of peaks anddips in the profiles of the online measurements.Increase in the concentration of CO2, drop in DOand increase in the stirrer speed during the first andsecond log phase of the culture correspond wellwith the respective growth phases. Results ob-tained with shake flask (Basu et al., 2006) andbioreactor studies strongly indicate that strainCSV86 prefers aromatic compounds and theirintermediate metabolites over glucose, and thataromatics and organic acids are co-metabolized.Repression of degradation of aromatic compoundby glucose in different strains of P. putida has beenreported for – benzyl alcohol and phenol at thetranscription level (Holtel et al., 1994; Mulleret al., 1996) and phenylacetic acid at transportlevel (Schleissner et al., 1994). Similarly, organicacids such as succinate, lactate, fumarate andacetate have been reported to repress phenol,chlorocatechol, toluene and benzyl alcohol degra-dation at the transcription level (Duetz et al.,1994, 1996; Muller et al., 1996; McFall et al.,1997). However, the utilization of aromatics wasrepressed neither by glucose nor by organic acidsfor strain CSV86. On the contrary, the utilization ofglucose was repressed by both aromatics andorganic acids. Repression of glucose uptake byorganic acids has been reported for other Pseudo-monas sp. (Tiwari and Campbell, 1969; Hylemonand Phibbs, 1972; Ng and Dawes, 1973). Theobserved patterns of substrate utilization areunique and may have implications in industrialapplications as well as for studies on regulatorymechanism. Understanding the mechanism of re-pression of glucose by aromatic compounds at themolecular and kinetic level will help to develop agenetically structured kinetic model. Such modelscan be used to predict the sequence and kinetics ofsubstrate uptake under different growth condi-tions. Such detailed understanding will facilitatethe use of versatile microbes like P. putida CSV86for the bioremediation of important pollutants.
Acknowledgment
AB thanks the University Grants Commission, Indiafor the award of senior research fellowship. PP andPW would like to thank the Department of Biotech-nology, Govt. of India for the research grant.
References
Alexander M. Biodegradation of chemicals of environ-mental concern. Science 1981;211:132–8.
Bapat PM, Das D, Dave NN, Wangikar PP. Phase shifts inthe stoichiometry of rifamycin B fermentation andcorrelation with the trends in the parameters mea-sured online. J Biotechnol 2006a;127:115–28.
Bapat PM, Sohoni SV, Moses TA, Wangikar PP. A cyberneticmodel to predict the effect of freely available nitrogensubstrate on rifamycin B production in complex media.Appl Microbiol Biotechnol 2006b;72:662–70.
Basu A, Phale PS. Inducible uptake and metabolism ofglucose by the phosphorylative pathway in Pseudomonasputida CSV86. FEMS Microbiol Lett 2006;259:311–6.
Basu A, Dixit SS, Phale PS. Metabolism of benzyl alcoholvia catechol ortho-pathway in methylnaphthalene-degrading Pseudomonas putida CSV86. Appl MicrobiolBiotechnol 2003;62:579–85.
Basu A, Apte SK, Phale PS. Preferential utilization ofaromatic compounds over glucose by Pseudomonasputida CSV86. Appl Environ Microbiol 2006;72:2226–30.
Boos W. Bacterial transport. Annu Rev Biochem 1974;43:123–46.
Buhler BBW, Bernhard H, Schmid A. Characterization andapplication of xylene monooxygenase for multistepbiocatalysis. Appl Environ Microbiol 2001;68:560–8.
Collier DN, Hager PW, Phibbs Jr. PV. Catabolite repressioncontrol in the Pseudomonads. Res Microbiol 1996;147:551–61.
Duetz WA, Marques S, de JC, Ramos JL, van Andel JG.Inducibility of the TOL catabolic pathway in Pseudo-monas putida (pWW0) growing on succinate incontinuous culture: evidence of carbon cataboliterepression control. J Bacteriol 1994;176:2354–61.
Duetz WA, Marques S, Wind B, Ramos JL, van Andel JG.Catabolite repression of the toluene degradation path-way in Pseudomonas putida harboring pWW0 undervarious conditions of nutrient limitation in chemostatculture. Appl Environ Microbiol 1996;62:601–6.
Frampton EW, Wood WA. Carbohydrate oxidation byPseudomonas fluorescens VI. Conversion of 2-keto-6-phosphogluconate to pyruvate. J Biol Chem 1961;236:2571–7.
Holtel A, Marques S, Mohler I, Jakubzik U, Timmis KN.Carbon source-dependent inhibition of xyl operonexpression of the Pseudomonas putida TOL plasmid.J Bacteriol 1994;176:1773–6.
Hylemon PB, Phibbs Jr. PV. Independent regulation ofhexose-catabolizing enzymes and glucose transportactivity in Pseudomonas aeruginosa. Biochem BiophysRes Commun 1972;48:1041–8.
Mahajan MC, Phale PS, Vaidyanathan CS. Evidence forthe involvement of multiple pathways in the biode-gradation of 1- and 2-methylnaphthalene by Pseudo-monas putida CSV86. Arch Microbiol 1994;161:425–33.
McFall SM, Abraham B, Narsolis CG, Chakrabarty AM. Atricarboxylic acid cycle intermediate regulating tran-scription of a chloroaromatic biodegradative pathway:fumarate-mediated repression of the clcABD operon.J Bacteriol 1997;179:6729–35.
Midgley M, Dawes EA. The regulation of transport ofglucose and methyl alpha-glucoside in Pseudomonasaeruginosa. Biochem J 1973;132:141–54.

ARTICLE IN PRESS
Sequential utilization of substrates by Pseudomonas putida CSV86 437
Mukkada AJ, Long GL, Romano AH. The uptake of 2-deoxy-D-glucose by Pseudomonas aeruginosa and itsregulation. Biochem J 1973;132:155–62.
Muller C, Petruschka L, Cuypers H, Burchhardt G, HerrmannH. Carbon catabolite repression of phenol degradation inPseudomonas putida is mediated by the inhibition of theactivator protein PhlR. J Bacteriol 1996;178:2030–6.
Ng FM, Dawes EA. Chemostat studies on the regulation ofglucose metabolism in Pseudomonas aeruginosa bycitrate. Biochem J 1973;132:129–40.
Phibbs Jr. PV, Eagon RG. Transport and phosphorylation ofglucose, fructose, and mannitol by Pseudomonasaeruginosa. Arch Biochem Biophys 1970;138:470–82.
Phillips AT, Mulfinger LM. Cyclic adenosine 30,50-monopho-sphate levels in Pseudomonas putida and Pseudomonasaeruginosa during induction and carbon catabolite repre-ssion of histidase synthesis. J Bacteriol 1981;145:1286–92.
Rentz JA, Alvarez PJ, Schnoor JL. Repression of Pseudo-monas putida phenanthrene-degrading activity byplant root extracts and exudates. Environ Microbiol2004;6:574–83.
Roberts BK, Midgley M, Dawes EA. The metabolism of 2-oxogluconate by Pseudomonas aeruginosa. J GenMicrobiol 1973;78:319–29.
Schleissner C, Olivera ER, Fernandez-Valverde M, LuengoJM. Aerobic catabolism of phenylacetic acid in
Pseudomonas putida U: biochemical characterizationof a specific phenylacetic acid transport system andformal demonstration that phenylacetyl-coenzyme Ais a catabolic intermediate. J Bacteriol 1994;176:7667–76.
Siegel LS, Hylemon PB, Phibbs Jr. PV. Cyclic adenosine30,50-monophosphate levels and activities of adenylatecyclase and cyclic adenosine 30,50-monophosphatephosphodiesterase in Pseudomonas and Bacteroides.J Bacteriol 1977;129:87–96.
Stinson MW, Cohen MA, Merrick JM. Isolation of dicar-boxylic acid- and glucose-binding proteins fromPseudomonas aeruginosa. J Bacteriol 1976;128:573–9.
Stulke J, Hillen W. Carbon catabolite repression inbacteria. Curr Opin Microbiol 1999;2:195–201.
Tiwari NP, Campbell JJ. Enzymatic control of themetabolic activity of Pseudomonas aeruginosa grownin glucose or succinate media. Biochim Biophys Acta1969;192:395–401.
Worsey MJ, Williams PA. Metabolism of toluene andxylenes by Pseudomonas (putida (arvilla) mt-2:evidence for a new function of the TOL plasmid. JBacteriol 1975;124:7–13.
Wylie JL, Worobec EA. The OprB porin plays a central rolein carbohydrate uptake in Pseudomonas aeruginosa.J Bacteriol 1995;177:3021–6.