serial killing of tumor cells by cytotoxic t cells redirected with a cd19-/cd3-bispecific...
TRANSCRIPT

Serial killing of tumor cells by cytotoxic T cells redirected
with a CD19-/CD3-bispecific single-chain antibody construct
Patrick Hoffmann1, Robert Hofmeister1, Klaus Brischwein1, Christian Brandl1, Sandrine Crommer1, Ralf Bargou2,Christian Itin1, Nadja Prang1 and Patrick A. Baeuerle1*
1Micromet AG, Munich, Germany2Robert-Rossle-Clinic, University Medical Center Charite, Humboldt University of Berlin, and Helios Kliniken, Berlin, Germany
Certain bispecific antibodies exhibit an extraordinary potency andefficacy for target cell lysis by eliciting a polyclonal T-cellresponse. One example is a CD19-/CD3-bispecific single-chainantibody construct (bscCD19xCD3), which at femtomolar concen-trations can redirect cytotoxic T cells to eliminate human B lym-phocytes, B lymphoma cell lines and patient-derived malignant Bcells. Here we have further explored the basis for this highpotency. Using video-assisted microscopy, bscCD19xCD3 wasfound to alter the motility and activity of T cells from a scanningto a killing mode. Individual T cells could eliminate multiple tar-get cells within a 9 hr time period, resulting in nuclear fragmenta-tion and membrane blebbing of target cells. Complete target cellelimination was observed within 24 hr at effector-to-target cellratios as low as 1:5. Under optimal conditions, cell killing startedwithin minutes after addition of bscCD19xCD3, suggesting thatthe rate of serial killing was mostly determined by T-cell move-ment and target cell scanning and lysis. At all times, T cellsremained highly motile, and no clusters of T and target cells wereinduced by the bispecific antibody. Bystanding target-negativecells were not detectably affected. Repeated target cell lysis bybscCD19xCD3-activated T cells increased the proportion ofCD19/CD3 double-positive T cells, which was most likely a conse-quence of transfer of CD19 from B to T cells during cytolytic syn-apse formation. To our knowledge, this is the first study showingthat a bispecific antibody can sustain multiple rounds of target celllysis by T cells.' 2005 Wiley-Liss, Inc.
Key words: cytotoxicity; bispecific antibody; CD19; CD3
Bispecific antibodies recruiting T cells have been under devel-opment for almost 20 years with great ingenuity and in a consider-able variety.1 Such bispecific antibodies hold the promise ofmounting a polyclonal T-cell response against tumor cells byemploying one of the immune system’s most effective killer cellpopulation. Normally, T cells are highly restricted in their specif-icity through a specific T-cell receptor recognizing a tumor-associ-ated peptide antigen presented by tumor cells through MHC classI (HLA) complexes. For activation and differentiation of naive Tcells, T cells require costimulatory signals that are typically absentfrom tumor cells of epithelial origin.2
A hallmark of late-stage tumors is that tumor cells are appa-rently selected for loss of T-cell recognition molecules.3 A fre-quent finding is the downregulation of expression or complete lossof HLA alleles on tumors.4–6 Likewise, tumor cells can becomedeficient in peptide antigen delivery and processing through lossof TAP transporters, which are responsible for loading peptidesonto MHC class I complexes in the ER7 and by exchanging pro-teasome subunits.8 Although certain vaccination approaches withtumor-associated peptides or proteins can induce tumor-specificT-cell responses in patients, the efficacy and response rate of suchtreatments is not very satisfactory and far from clinic routine.9
There is an increasing list of immune escape mechanisms thatinvolve among other strategies the secretion by tumor cells ofT-cell-differentiating cytokines such as TGF-beta and IL-10, theoverexpression of protease inhibitors potentially protecting tumorcells from granzymes and the expression of ligands for pro-apop-totic receptors on T cells.10 The nature and frequency of evasionmechanisms suggests that T cells are indeed the prime enemies oftumor cells.
Bispecific antibodies targeting with one arm a common activat-ing molecule present on all T cells, and with the second arm awidespread tumor-associated antigen, may be able to circumvent avariety of immune escape mechanisms of tumor cells. A well-studied CD19-/CD3-bispecific antibody is bscCD19xCD3.11,12 Asa bispecific single-chain (sc) antibody, bscCD19xCD3 aligns anN-terminal scFv specific for the pan-B-cell antigen CD19 with aC-terminal scFv specific for CD3, the common signaling moleculecomplex associated with the T-cell receptor on all T cells. Becauseof its high potency for T-cell activation, this class of molecules isreferred to as bispecific T-cell engager (BiTE).
BscCD19xCD3 has been analysed in certain detail for efficacyand its mode of action in cell culture models using cell lines,11
with lymphoma patient-derived samples13 and in vivo using SCIDmouse xenotransplant models.14 A common observation in allthese systems was that the BiTE molecule bscCD19xCD3 wasextremely potent in lysis of CD19-positive cells and couldpotently activate unstimulated T cells in the absence of costimuli.Here we have further explored the mechanism of action. To thisend, software and equipment were developed to allow tracking ofthe fate of single T cells and tumor target cells by video-assistedmicroscopy. Essential findings by this method were then supportedwith experiments using larger cell populations in cell culture cyto-toxicity assays. A key finding of our study is that bscCD19xCD3can recruit T cells for multiple rounds of target cell elimination.During this process, T cells appear to acquire and accumulate theCD19 target from their prey. Our new findings are discussed withrespect to other bispecific antibodies and the current understandingof the mode of action underlying BiTE activity.
Material and methods
Reagents
BscCD19xCD3 was provided by Micromet (Munich, Ger-many). Expression and purification of the hexahistidine-taggedprotein were essentially carried out as described.12 Calcein AMwas purchased from Molecular Probes (Eugene, OR). The PKH26Red Fluorescent Cell Linker kit was from Sigma-Aldrich(Taufkirchen, Germany). The anti-CD19-FITC-conjugated anti-body was purchased from DAKO (Hamburg, Germany) and theanti-CD3-PE-conjugated antibody was from Becton-Dickinson(Heidelberg, Germany).
Cell culture, cell lines and T-cell isolation
The pre-B-cell lymphoma cells NALM-6 were purchased fromthe Deutsche Sammlung von Mikroorgansimen und Zelllinien(DSMZ, Braunschweig, Germany). TF-1 erythroleukemia cellswere from the European Collection of Cell Cultures (ECACC,Wiltshire, UK). The CD4-positive T-cell clone MC15 was a
The first two authors contributed equally to this work.*Correspondence to: Micromet AG, Staffelseestr. 2, 81477 Munich,
Germany. Fax: þ49-89-89-52-77-505.E-mail: [email protected] 16 July 2004; Accepted after revision 24 November 2004DOI 10.1002/ijc.20908Published online 1 February 2005 in Wiley InterScience (www.interscience.
wiley.com).
Int. J. Cancer: 115, 98–104 (2005)
' 2005 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

subclone of CB15 cells kindly provided by Dr. Fickenscher(University of Heidelberg, Germany). Cells were cultured as re-commended by the supplier.
Peripheral blood mononuclear cells (PBMC) were prepared byFicoll density gradient centrifugation from blood filters obtainedfrom local blood banks. Erythrocytes were removed by incubationin erythrocyte lysis buffer (155 mM NH4Cl, 10 mM KHCO3,100 mM EDTA) for 15 min at room temperature. Cells were cen-trifuged for 5 min at 600g. The supernatant containing the lysederythrocytes was discarded, and cells were resuspended in PBS.Thrombocytes were removed in an additional centrifugation stepfor 15 min at 100g. PBMC were resuspended in culture mediumand typically used from 1 to 4 days after preparation without stim-ulation.
CD8þ T cells were isolated using the Human CD8 Subset Col-umn kit (R&D Systems, Wiesbaden, Germany) according to themanufacturer’s instructions. The CD8þ cytotoxic T-cell clones 11and SuHiE7 were isolated from a mixed lymphocyte reaction.Allogeneic feeder PBMC were prepared by Ficoll density centrifu-gation from heparinized blood. Erythrocytes were removed andthe PBMC washed twice in 15 ml RPMI 1640/1% FCS. After thesecond wash step, the pelleted cells were irradiated at 5000 R. Forthe mixed lymphocyte culture, cell density of the irradiated PBMCwas adjusted to 3 � 106 cells/ml in complete RPMI 1640 contain-ing 5% human serum, 1% glutamine, 1% sodium pyruvate, 1%nonessential amino acids, 1% kanamycin and 5 � 10-5 M 2-mer-captoethanol. In addition, the complete RPMI 1640 medium wassupplemented with 1,000 U recombinant human IL-2. Aliquots(100 ml) of 500,000 cells were transferred to each well of a 96-well round-bottom plate. Cytotoxic T-cell clones were generatedfrom PBMC isolated from heparinized blood. PBMC were pre-pared by Ficoll density centrifugation and erythrocytes removedas described above. CD8þ T cells were enriched by means of theHuman CD8 Subset Column kit (R&D Systems). Subsequently,the CD8þ T cells were resuspended at a density of 3 cells/ml incomplete RPMI 1640 containing 2 mg/ml PHA. Aliquots (100 ml)were transferred to each well of the plates containing the feedercells. The cells were restimulated every 5 to 10 days by removing100 ml of the culture medium and replacing it with 100 ml com-plete RPMI 1640 medium, which contained 106 irradiated PBMC/ml and was supplemented with 500 U recombinant human IL-2and 1 mg/ml PHA.
Video-assisted microscopy
The behaviour of T cells was recorded by means of video-assisted microscopy. Images were acquired using the inversemicroscope Nikon TE 200 equipped with an XYZ axis scanningstage (Marzhauser, Wetzlar, Germany) and a Sony DXC 9100PCCD camera (Sony, Cologne, Germany). For temperature control,the microscope was covered with a plastic hood equipped with aventilated heating system. Prior to the start of the experiment, starttemperature equalization to 378C was performed for 2 hr.
The human T-effector cell clones (1.3 � 104 cells/well) andNALM-6 or TF-1 target cells (4 � 105) were mixed in Nunc 4x2chamber slides (Nunc, Roskilde, Denmark) in 400 ml RPMI 1640/10% FCS containing 25 mM HEPES (Invitrogen, Karlsruhe, Ger-many) at an E:T ratio of 1:30. Cell movement and activity weremonitored in the presence or absence of 50 ng/ml bscCD19xCD3.The movement of cells was recorded every 18 sec. Images wereprocessed using LuciaG 4.7 software (LMI, Prague, Czech Repub-lic). A macro was programmed to mark the mass center of a singlecell by a red dot. The trajectories were visualized by an overlay ofthe captured images.
Cytotoxicity assay
Two different types of assays were used for the determinationof the cytotoxic activity of bscCD19xCD3. If not otherwise indi-cated, cytotoxicity was determined after 3 hr by means of a calcein
AM release assay. NALM-6 or TF-1 cells (1.5 � 107) werelabeled with 10 mM calcein AM (Molecular Probes) for 30 min at378C in cell culture medium. After 2 washes in cell culturemedium, cell density was adjusted to 3 � 105 cells/ml in RPMI1640/10% FCS and 100 ml aliquots of 30,000 cells were used perassay reaction. In a 96-well round-bottom plate either CD8þ Tcells or the cytotoxic T-cell line MC15 and target cells were co-cultured at the indicated bscCD19xCD3 concentrations in tripli-cate. For the E:T ratios of 10:1, 5:1 and 1:1, the number of targetcells was kept constant at 3 � 104 cells/well. For the E:T ratios of1:5 and 1:10, the number of effector cells was kept constant at 3 �104 cells/well. Target cells were added accordingly to obtain theindicated E:T ratios. BscCD19xCD3 was diluted in RPMI 1640/10% FCS to the required concentration. Total reaction volumewas 200 ml. The reactions were incubated for 3 hr. The release offluorescent dye into the supernatants was measured as marker forcell lysis using a fluorescence reader (Tecan, Crailsheim,Germany). Unspecific release of the fluorescent dye into the super-natant was determined by incubating effector and target cells inthe absence of bscCD19xCD3. To determine total cell lysis, themixture of effector and labelled target cells withoutbscCD19xCD3 was lysed by the addition of 20 ml of 1% saponinfor 10 min. Specific cytotoxicity was calculated according to thefollowing formula:
%spec.lysis
¼ fluorescene of sample� fluorescene of control
fluorescene of total lysis� fluorescene of control
� �� 100
For the analysis of cell lysis after 24 hr, the fluorescent mem-brane dye PKH26 was used to label NALM-6 target cells and todistinguish them from CD8þ effector cells in a flow cytometer-based assay. Briefly, 1 � 107 cells were washed twice with 10 mlPBS before the cells were resuspended in 0.5 ml diluent C pro-vided with the kit. In a second tube, 0.5 ml diluent C were mixedwith 5 ml of PHK26 dye by vortexing. This solution was added tothe cell suspension. Subsequently, the cells were mixed and incu-bated for 2 min at room temperature. Then the staining reactionwas quenched by adding 1 ml FCS. The tube was filled up with15 ml complete cell culture medium and centrifuged for 5 min at300g. After a second wash with 15 ml complete medium, the num-ber of viable cells was determined by Trypan blue exclusion.CD8þ T effector and NALM-6 target cells were co-cultured in96-well round-bottom plates at an E:T ratio of 10:1. Per well,5 � 105 PBMC effector cells were cultured with 5 � 104 PKH26-labeled target cells in the absence or presence of the indicatedMT103 concentrations in a volume of 200 ml. After 24 hr, the cellswere washed twice with 200 ml PBS. Afterward, the cells wereresuspended in 300 ml FACS buffer. Before the cells’ acquisitionusing a FACS Calibur flow cytometer (BD), propidium iodide at afinal concentration of 1 mg/ml was added to the cells to discrimi-nate between alive and dead (PI-positive) cells. Cell lysis was cal-culated according to the following formula:
%spec.lysis
¼ 1� number of alive cells in sample
number of alive cells in control w/o bscCD19xCD3
� ��100
Flow cytometry analysis
The accumulation of CD19 on T cells was analyzed by meansof flow cytometry. T cells were enriched from PBMC using theT Cell Enrichment Column kit (R&D Systems). T cells wereincubated with the target cells in the presence of 1 ng/mlbscCD19xCD3 in 96-well round-bottom plates for 24 hr at 378C.For an E:T ratio of 1:10 2 � 104 isolated T cells were incubatedwith 2 � 105 NALM-6 or Karpas-299 cells, respectively. Toobtain an E:T ratio of 10:1, cell numbers were inverted.
99SERIAL TUMOR CELL LYSIS BY BISPECIFIC ANTIBODY

The mixture of effector T and target cells was stained with anti-CD3-PE and CD19-FITC-conjugated antibodies for 30 min at4–88C. After resuspension of the cell mixture in 200 ml FACS buf-fer (PBS containing 1% FCS and 0.05% Na3N), propidium iodidewas added at a final concentration of 1 mg/ml. Acquisition of thecells was carried out right after staining. Accumulation of CD19on T cells was performed in 2 steps: (i) Live T cells were identi-fied on a CD3-PE/PI dot plot by their CD3 positivity and PI nega-tivity (gate 1); (ii) The shift in CD19 positivity was detected on aCD3-FITC/CD19 dot plot (gate 2). To exclude that the increase ofCD19 on T cells was a consequence potentially formed by cellaggregates, the CD3/CD19 double-positive cells were back-gatedon a FSC/SSC plot.
Results
BsCD19xCD3 alters the behaviour of T cells
To investigate serial killing by bscCD19xCD3-activated T cells,only a few T cells were spiked into a cell culture of CD19-positivetarget cells. The CD19-positive human pre-B lymphoma lineNALM-615 was mixed at an E:T ratio of 1:30 with the humanCD8-positive T-cell clone 11. Clone 11 has an unknown antigenspecificity and cannot mediate lysis of NALM-6 cells in theabsence of bscCD19xCD3. In movies from video-assisted micro-scopy, T cells could be easily distinguished from NALM-6 cellsby their smaller size and high amoeboid motility. Two-dimen-sional T-cell movement was captured by a modified softwaredeveloped to mark the mass center of T cells every 18 sec by a reddot. Under control conditions, resulting T-cell trajectories revealeda relatively even and high pace of T cells allowing an individualcell to cover a relatively large area in the culture dish. In theexample shown (Fig. 1a), the observed T cell scanned an area ofapproximately 6,000 mm2 during the 9 hr recording period, therebybriefly contacting a total of 24 NALM-6 lymphoma cells. Thelymphoma cells showed very little independent movement duringthe observation period.
The behaviour of T cells profoundly changed in the presence of50 ng/ml bscCD19xCD3. T cells now became much more focusedon just a few target cells. In the 3 examples shown in Figure 1b,the area covered by T cells in the same time period as in Figure 1anow ranged from approximately 900–1,200 mm2, and only 5–10target cells were encountered in the 2-dimensional setting duringthe 9 hr incubation. As a consequence of the altered T-cell behav-ior, massive NALM-6 cell lysis was observed. While under con-trol conditions, NALM-6 cells continued dividing and exhibited abright ring in phase contrast microscopy. In the presence ofbscCD19xCD3-activated T cells, a large proportion of NALM-6cells had lost phase contrast brightness and exhibited fragmentedor condensed nuclei and blebbing membranes (Fig. 1c). Previousstudies with bscCD19xCD3 have shown that CD19-positive targetcells undergo apoptosis and cell lysis as determined by eitherchromium-51 release, propidium iodide uptake, calcein AMrelease or annexin-V positivity.11–13
Serial target cell killing by bscCD19xCD3-activated T cells
The activity of single bscCD19xCD3-activated T cells wasmonitored over a 9 hr time period by video-assisted microscopy.In the example shown in Figure 2, one T cell contacted at least 5target cells for a longer period of time. In 4 of 5 contacts, the inter-action of the T cell was very focused on one side of the target cell,whereas with target cell no. 3, the T cell circled the target cell fora prolonged time. The contact time for target cells 1, 2, 4 and 5ranged from 22–97 min; for target cell no. 3, it was 155 min.Whereas the 5 target cells had a normal appearance before contactwith the bscCD19xCD3-activated T cell (Fig. 2b, upper panel), allhad an altered morphology after 9 hr (Fig. 2b, lower panel). Con-densed nuclei and loss of phase contrast brightness indicated thatthe cells had undergone apoptotic changes and lysis. NeighbouringNALM-6 cells, which had not been contacted by the T cell stillappeared normal (Fig. 2b).
To further support serial target cell killing by T cells in the pres-ence of bscCD19xCD3, cytotoxicity assays were performed at var-ious E:T ratios by determining the release of calcein AM fromNALM-6 target cells preloaded with the fluorescent dye. Althoughafter 4 hr appreciable cell lysis was only seen with E:T ratios of10:1 and 5:1 (Fig. 3a), close to complete cell lysis was achievedafter 24 hr at E:T ratios ranging from 10:1 to 1:5 (Fig. 3b). After24 hr, even an E:T ratio of 1:10 yielded at 1 ng/ml approximately50% cell lysis. The efficient cell lysis at very low E:T ratios isconsistent with a serial target cell killing, and the kinetics suggestthat time is the limiting factor for low T-cell numbers to efficientlyeliminate larger numbers of target cells. Half maximal doses ofbscCD19xCD3 for cell lysis were in the range of 20 (0.36 pM atE:T ¼ 1:1) to 200 pg/ml (3.6 pM at E:T ¼ 1:10).
To investigate how rapidly T cells can eliminate target cellsafter addition of bscCD19xCD3, a kinetic analysis was performedat an excess of T cells over target cells. The high E:T ratio ensuredthat T cells had immediate access to their target cells without theneed for much movement and scanning. The amount of calceinreleased into the cell culture supernatant was determined after theindicated periods of time (Fig. 4) and expressed as percent of the
FIGURE 1 – The effect of bscCD19xCD3 on the motility of T cellsand viability of NALM-6 B lymphoma cells monitored over 9 hr. (a)Trajectory of 1 representative T cell in the absence of bscCD19xCD3.(b) Trajectories of 3 different T cells in the presence of bscCD19xCD3(50 ng/ml). (c) Morphology of the NALM-6 target cells at the begin-ning (left) and after 9 hr bscCD19xCD3 incubation (right). HumanCD8-positive T-cell clone 11 and NALM-6 human pre-B lymphomacells were mixed at an E:T ratio of 1:30.
100 HOFFMANN ET AL.

calcein AM released after complete cell lysis by detergent treat-ment. After as little time as 10 min, ca. 40% target cell lysis wasreached (Figs. 4a,b). Half maximal lysis was achieved after20 min, and lysis plateaued after 100 min at close to 100%. Thisshowed that cell lysis by bscCD19xCD3-activated T cells couldstart without a significant lag phase and reach completion.
Bystanding target-negative cells are not harmedby bscCD19xCD3-activated T cells
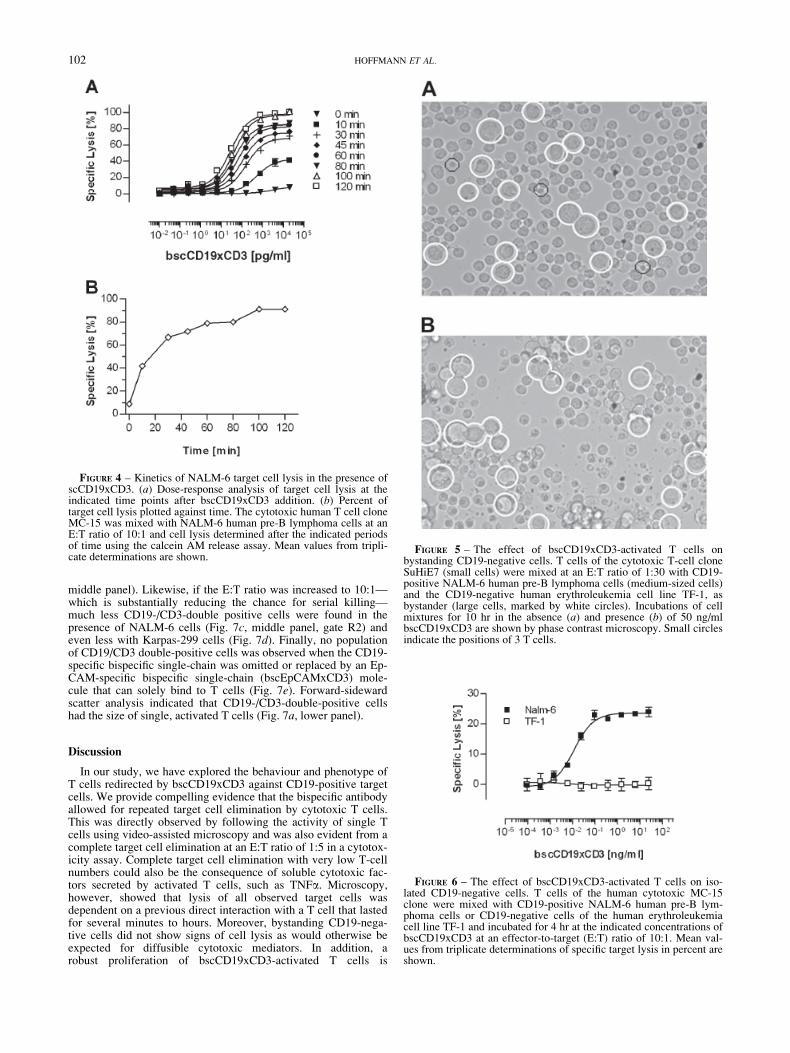
The specificity of target cell lysis was tested by mixing CD19-positive NALM-6 cells with CD19-negative TF-1 cells, a humanerythroleukemia line. TF-1 cells were also chosen because theycan be distinguished under the microscope by their larger sizefrom NALM-6 cells. After 9 hr in the presence of bscCD19xCD3-activated T cells at an E:T ratio of 1:30, a large proportion of themid-sized NALM-6 cells showed fragmented or condensed nuclei,and the number of cell fragments was substantially increased. Theappearance of the large CD19-negative TF-1 cells was, however,unchanged, and a bright ring was still seen upon phase contrastmicroscopy (Fig. 5).
To corroborate these findings TF-1 cells were also separatelyincubated with increasing concentrations of bscCD19xCD3 at anE:T ratio of 10:1 for 3 hr in a cytotoxicity assay. Even at a 100-fold higher E:T ratio as used in Figure 5 and at bscCD19xCD3concentrations up to 150 ng/ml no lysis of TF-1 was observed,whereas NALM-6 cells were effectively killed under identicalassay conditions (Fig. 6).
Serial killing leads to CD19 target accumulation on T cells
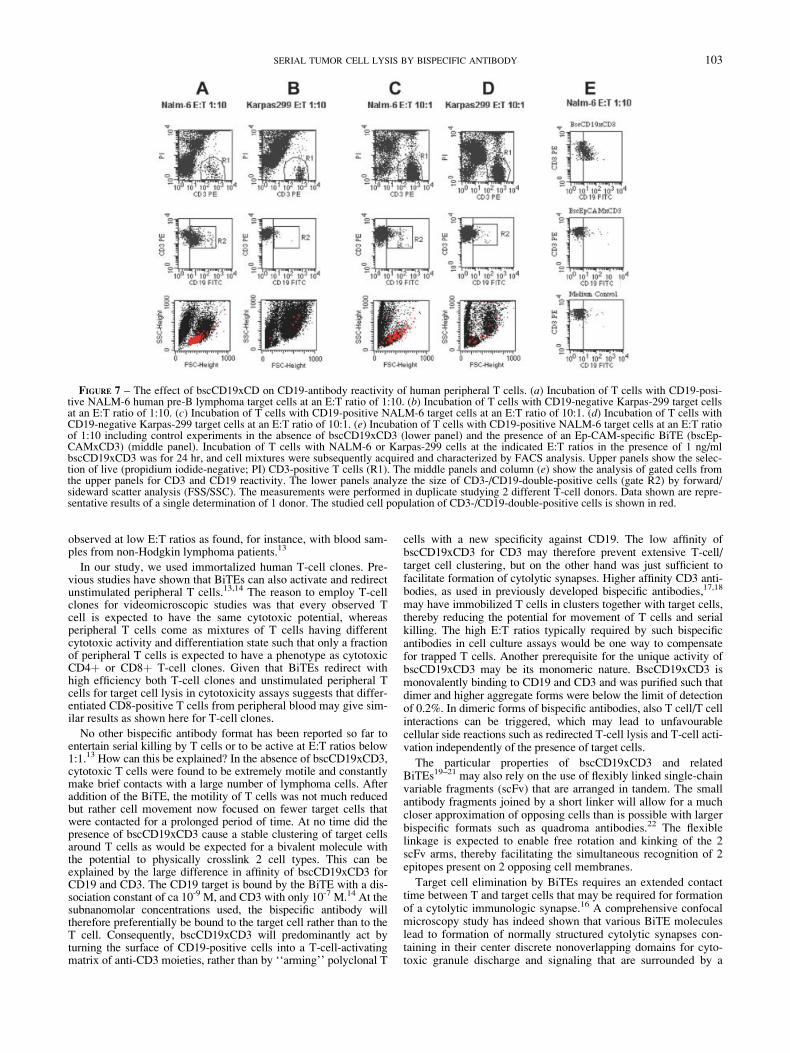
Formation of cytolytic immunologic synapses is required fortarget cell lysis by cytotoxic T cells.16 It has been reported that insuch synapses membrane fusions can occur. Here we testedwhether T-cell killing by bscCD19xCD3 at very low E:T ratiosleads to accumulation of the CD19 antigen on T cells, resulting ina T-cell population that is double-positive for CD3 and CD19. Asshown in Figure 7, we indeed observed that a substantial portionof CD3-positive T cells (50–60%) became CD19-positive after24 hr at an E:T ratio of 1:10 (Fig. 7a, middle panel, gate R2). IfCD19-positive NALM-6 were substituted by CD19-negativeKarpas-299 cells, no such cell population was observed (Fig. 7b,
FIGURE 2 – Serial killing oftarget cells by bscCD19xCD3.(a) Prolonged contacts of 1 individ-ual T cell with 5 different NALM-6B lymphoma cells during a 9 hrincubation. Pictures were taken atthe following time points afterBiTE addition: 1:62 min, 2:100min; 3:256 min; 4:310 min; 5:356min. (b) Morphologic changes ofNALM-6 cells being contacted by1 T cell after 9 hr. The human cyto-toxic T-cell clone 11 was mixed ata 1:30 ratio with NALM-6 pre-Bhuman lymphoma cells in the pres-ence of 50 ng/ml bscCD19xCD3.The 2-dimensional movement ofcells was recorded by video-assisted phase contrast microscopy.The red dots indicate the positionsof the mass center of individualT cells. The positions of T cells areshown every 18 sec. Those targetcells that were in longest contactwith the T cell, as indicated bytrajectories, are marked by a circle.
FIGURE 3 – The effect of effector-to-target ratio and incubationtime on the lysis of NALM-6 target cells by a human T-cell clone inthe presence of variable concentrations of bscCD19xCD3. (a) NALM-6 target cell lysis after 4 hr. (b) NALM-6 target cell lysis after 24 hr.CD8þ T cells isolated from a healthy donor were incubated withNALM-6 target cells at the indicated E:T ratios and bscCD19xCD3concentrations for 4 and 24 hr, respectively. Cell lysis was determinedby a calcein AM release assay after 4 hr and a flow cytometry-basedassay using the fluorescence dye PKH26 after 24 hr. Mean valuesfrom triplicate values are shown.
101SERIAL TUMOR CELL LYSIS BY BISPECIFIC ANTIBODY
middle panel). Likewise, if the E:T ratio was increased to 10:1—which is substantially reducing the chance for serial killing—much less CD19-/CD3-double positive cells were found in thepresence of NALM-6 cells (Fig. 7c, middle panel, gate R2) andeven less with Karpas-299 cells (Fig. 7d). Finally, no populationof CD19/CD3 double-positive cells was observed when the CD19-specific bispecific single-chain was omitted or replaced by an Ep-CAM-specific bispecific single-chain (bscEpCAMxCD3) mole-cule that can solely bind to T cells (Fig. 7e). Forward-sidewardscatter analysis indicated that CD19-/CD3-double-positive cellshad the size of single, activated T cells (Fig. 7a, lower panel).
Discussion
In our study, we have explored the behaviour and phenotype ofT cells redirected by bscCD19xCD3 against CD19-positive targetcells. We provide compelling evidence that the bispecific antibodyallowed for repeated target cell elimination by cytotoxic T cells.This was directly observed by following the activity of single Tcells using video-assisted microscopy and was also evident from acomplete target cell elimination at an E:T ratio of 1:5 in a cytotox-icity assay. Complete target cell elimination with very low T-cellnumbers could also be the consequence of soluble cytotoxic fac-tors secreted by activated T cells, such as TNFa. Microscopy,however, showed that lysis of all observed target cells wasdependent on a previous direct interaction with a T cell that lastedfor several minutes to hours. Moreover, bystanding CD19-nega-tive cells did not show signs of cell lysis as would otherwise beexpected for diffusible cytotoxic mediators. In addition, arobust proliferation of bscCD19xCD3-activated T cells is
FIGURE 4 – Kinetics of NALM-6 target cell lysis in the presence ofscCD19xCD3. (a) Dose-response analysis of target cell lysis at theindicated time points after bscCD19xCD3 addition. (b) Percent oftarget cell lysis plotted against time. The cytotoxic human T cell cloneMC-15 was mixed with NALM-6 human pre-B lymphoma cells at anE:T ratio of 10:1 and cell lysis determined after the indicated periodsof time using the calcein AM release assay. Mean values from tripli-cate determinations are shown.
FIGURE 5 – The effect of bscCD19xCD3-activated T cells onbystanding CD19-negative cells. T cells of the cytotoxic T-cell cloneSuHiE7 (small cells) were mixed at an E:T ratio of 1:30 with CD19-positive NALM-6 human pre-B lymphoma cells (medium-sized cells)and the CD19-negative human erythroleukemia cell line TF-1, asbystander (large cells, marked by white circles). Incubations of cellmixtures for 10 hr in the absence (a) and presence (b) of 50 ng/mlbscCD19xCD3 are shown by phase contrast microscopy. Small circlesindicate the positions of 3 T cells.
FIGURE 6 – The effect of bscCD19xCD3-activated T cells on iso-lated CD19-negative cells. T cells of the human cytotoxic MC-15clone were mixed with CD19-positive NALM-6 human pre-B lym-phoma cells or CD19-negative cells of the human erythroleukemiacell line TF-1 and incubated for 4 hr at the indicated concentrations ofbscCD19xCD3 at an effector-to-target (E:T) ratio of 10:1. Mean val-ues from triplicate determinations of specific target lysis in percent areshown.
102 HOFFMANN ET AL.
observed at low E:T ratios as found, for instance, with blood sam-ples from non-Hodgkin lymphoma patients.13
In our study, we used immortalized human T-cell clones. Pre-vious studies have shown that BiTEs can also activate and redirectunstimulated peripheral T cells.13,14 The reason to employ T-cellclones for videomicroscopic studies was that every observed Tcell is expected to have the same cytotoxic potential, whereasperipheral T cells come as mixtures of T cells having differentcytotoxic activity and differentiation state such that only a fractionof peripheral T cells is expected to have a phenotype as cytotoxicCD4þ or CD8þ T-cell clones. Given that BiTEs redirect withhigh efficiency both T-cell clones and unstimulated peripheral Tcells for target cell lysis in cytotoxicity assays suggests that differ-entiated CD8-positive T cells from peripheral blood may give sim-ilar results as shown here for T-cell clones.
No other bispecific antibody format has been reported so far toentertain serial killing by T cells or to be active at E:T ratios below1:1.13 How can this be explained? In the absence of bscCD19xCD3,cytotoxic T cells were found to be extremely motile and constantlymake brief contacts with a large number of lymphoma cells. Afteraddition of the BiTE, the motility of T cells was not much reducedbut rather cell movement now focused on fewer target cells thatwere contacted for a prolonged period of time. At no time did thepresence of bscCD19xCD3 cause a stable clustering of target cellsaround T cells as would be expected for a bivalent molecule withthe potential to physically crosslink 2 cell types. This can beexplained by the large difference in affinity of bscCD19xCD3 forCD19 and CD3. The CD19 target is bound by the BiTE with a dis-sociation constant of ca 10-9 M, and CD3 with only 10-7 M.14 At thesubnanomolar concentrations used, the bispecific antibody willtherefore preferentially be bound to the target cell rather than to theT cell. Consequently, bscCD19xCD3 will predominantly act byturning the surface of CD19-positive cells into a T-cell-activatingmatrix of anti-CD3 moieties, rather than by ‘‘arming’’ polyclonal T
cells with a new specificity against CD19. The low affinity ofbscCD19xCD3 for CD3 may therefore prevent extensive T-cell/target cell clustering, but on the other hand was just sufficient tofacilitate formation of cytolytic synapses. Higher affinity CD3 anti-bodies, as used in previously developed bispecific antibodies,17,18
may have immobilized T cells in clusters together with target cells,thereby reducing the potential for movement of T cells and serialkilling. The high E:T ratios typically required by such bispecificantibodies in cell culture assays would be one way to compensatefor trapped T cells. Another prerequisite for the unique activity ofbscCD19xCD3 may be its monomeric nature. BscCD19xCD3 ismonovalently binding to CD19 and CD3 and was purified such thatdimer and higher aggregate forms were below the limit of detectionof 0.2%. In dimeric forms of bispecific antibodies, also T cell/T cellinteractions can be triggered, which may lead to unfavourablecellular side reactions such as redirected T-cell lysis and T-cell acti-vation independently of the presence of target cells.
The particular properties of bscCD19xCD3 and relatedBiTEs19–21 may also rely on the use of flexibly linked single-chainvariable fragments (scFv) that are arranged in tandem. The smallantibody fragments joined by a short linker will allow for a muchcloser approximation of opposing cells than is possible with largerbispecific formats such as quadroma antibodies.22 The flexiblelinkage is expected to enable free rotation and kinking of the 2scFv arms, thereby facilitating the simultaneous recognition of 2epitopes present on 2 opposing cell membranes.
Target cell elimination by BiTEs requires an extended contacttime between T and target cells that may be required for formationof a cytolytic immunologic synapse.16 A comprehensive confocalmicroscopy study has indeed shown that various BiTE moleculeslead to formation of normally structured cytolytic synapses con-taining in their center discrete nonoverlapping domains for cyto-toxic granule discharge and signaling that are surrounded by a
FIGURE 7 – The effect of bscCD19xCD on CD19-antibody reactivity of human peripheral T cells. (a) Incubation of T cells with CD19-posi-tive NALM-6 human pre-B lymphoma target cells at an E:T ratio of 1:10. (b) Incubation of T cells with CD19-negative Karpas-299 target cellsat an E:T ratio of 1:10. (c) Incubation of T cells with CD19-positive NALM-6 target cells at an E:T ratio of 10:1. (d) Incubation of T cells withCD19-negative Karpas-299 target cells at an E:T ratio of 10:1. (e) Incubation of T cells with CD19-positive NALM-6 target cells at an E:T ratioof 1:10 including control experiments in the absence of bscCD19xCD3 (lower panel) and the presence of an Ep-CAM-specific BiTE (bscEp-CAMxCD3) (middle panel). Incubation of T cells with NALM-6 or Karpas-299 cells at the indicated E:T ratios in the presence of 1 ng/mlbscCD19xCD3 was for 24 hr, and cell mixtures were subsequently acquired and characterized by FACS analysis. Upper panels show the selec-tion of live (propidium iodide-negative; PI) CD3-positive T cells (R1). The middle panels and column (e) show the analysis of gated cells fromthe upper panels for CD3 and CD19 reactivity. The lower panels analyze the size of CD3-/CD19-double-positive cells (gate R2) by forward/sideward scatter analysis (FSS/SSC). The measurements were performed in duplicate studying 2 different T-cell donors. Data shown are repre-sentative results of a single determination of 1 donor. The studied cell population of CD3-/CD19-double-positive cells is shown in red.
103SERIAL TUMOR CELL LYSIS BY BISPECIFIC ANTIBODY

zone of cell adhesion (Offner et al., data submitted). Here we canfurther support these findings by the observation that under certainconditions T cells can acquire and accumulate CD19 antigen fromtheir target cells. This apparent antigen transfer was only observedat very low E:T ratios that enable multiple rounds of target cellelimination by T cells and did not occur in the absence of theCD19-specific BiTE or the presence of a BiTE solely recognizingT cells. We consider it unlikely that we studied stable conjugatesof T and B cells. The low affinity of the BiTE for CD3, theabsence of cell clusters in video microscopy and the small size ofCD19/CD3-double-positive cells in forward/sideward scatter anal-ysis speak in favor of single CD19-positive T cell. The small sizeof CD19/CD3-double-positive cells also makes it unlikely thatCD3 antigen was transferred from T cells to the larger NALM-6 Blymphoma cells. Another less likely explanation is that weinduced an aberrant neoexpression of CD19 in T cells. De novoCD19 expression was observed in a low percentage of myeloidleukemia cases,23 where it is thought to be the consequence ofgene translocation events.
Electron microscopy studies revealed that membrane contactsin cytolytic immunologic synapses become so close that fusions ofopposing plasma membranes can occur.16 As a consequence, Tcells can acquire the antigen specificity of the target cell uponrepeated target cell lysis. It was discussed that the acquisition ofloaded MHC class I complexes by this reaction can be instrumen-tal to downmodulate a specific T-cell response through mutualkilling of specific T cells, a process referred to as fratricide.24 Fornow, we could not obtain evidence that CD19-positive T cells aremore susceptible to lysis than CD19-negative T cells, but this willrequire further investigations.
The kinetic analysis of cell lysis performed in our study indi-cated that synapse formation and cell lysis apparently started with-out a lag phase and that target cell lysis in the presence of anexcess of cytotoxic T cells reached completion after 100 min. Thistime frame covers well the variable duration of T cell/target cellcontacts observed in video microscopy in the presence ofbscCD19xCD3. Our data suggest that a therapeutic use ofbscCD19xCD3 must consider that drug action entirely relies onT-cell activity and, consequently, on the time necessary for T cellsto reach and scan potential target cells, to eventually form a cyto-lytic synapse with a target cell, disengage from the killed target,move on and detect the next target cell for elimination. In ourexperiments, T cells could eliminate 4–6 target cells in 9 hr. How-ever, this was in a 2-dimensional setting. The rate of serial killingmay be higher in a 3-dimensional tumor tissue because of highertarget density and 3-dimensional accessibility. On the other hand,T cells in a tumor may have to cover larger distances and needtime to penetrate nontumor tissue in particular if only few targetcells are left in tissue. Effector-to-target ratio, tumor-target cellload, tumor penetration by bispecific antibody and T cells, T-celltrafficking, T-cell number, composition and activity, and escapemechanisms acquired by tumor cells will all factor into the in vivoefficacy of bscCD19xCD3. With our finding that bscCD19xCD3can sustain repeated target cell lysis by T cells, a further prerequi-site for in vivo efficacy of bscCD19xCD3 would be fulfilled.
Acknowledgements
We thank Dr. S. Zeman and Dr. J. Suzich for critical reading ofthe manuscript.
References
1. Segal DM, Weiner GJ, Weiner LM. Bispecific antibodies in cancertherapy. Curr Opin Immunol 1999;11:558–62.
2. McAdam AJ, Schweitzer AN, Sharpe AH. The role of B7 co-stimula-tion in activation and differentiation of CD4þ and CD8þ T cells.Immunol Rev 1998;165:231–47.
3. Gabrilovich D, Pisarev V. Tumor escape from immune response:mechanisms and targets of activity. Curr Drug Targets 2003;4:525–36.
4. Hicklin DJ, Marincola FM, Ferrone S. HLA class I antigen downregu-lation in human cancers: T-cell immunotherapy revives an old story.Mol Med Today 1999;5:178–86.
5. Kageshita T, Hirai S, Ono T, Hicklin DJ, Ferrone S. Down-regulationof HLA class I antigen-processing molecules in malignant melanoma:association with disease progression. Am J Pathol 1999;154:745–54.
6. Rees RC, Mian S. Selective MHC expression in tumours modulatesadaptive and innate antitumour responses. Cancer Immunol Immun-other 1999;48:374–81.
7. Johnsen AK, Templeton DJ, Sy M, Harding CV. Deficiency of trans-porter for antigen presentation (TAP) in tumor cells allows evasion ofimmune surveillance and increases tumorigenesis. J Immunol 1999;163:4224–31.
8. Miyagi T, Tatsumi T, Takehara T, Kanto T, Kuzushita N, SugimotoY, Jinushi M, Kasahara A, Sasaki Y, Hori M, Hayashi N. Impairedexpression of proteasome subunits and human leukocyte antigensclass I in human colon cancer cells. J Gastroenterol Hepatol 2003;18:32–40.
9. Spadaro M, Lanzardo S, Curcio C, Forni G, Cavallo F. Immunologi-cal inhibition of carcinogenesis. Cancer Immunol Immunother 2004;53:204–16.
10. Baeuerle PA, Kufer P, Lutterbuse R. Bispecific antibodies for polyclo-nal T-cell engagement. Curr Opin Mol Ther 2003;5:413–9.
11. Dreier T, Lorenczewski G, Brandl C, Hoffmann P, Syring U, Hana-kam F, Kufer P, Riethmuller G, Bargou R, Baeuerle PA. Extremelypotent, rapid and costimulation-independent cytotoxic T-cell responseagainst lymphoma cells catalyzed by a single-chain bispecific anti-body. Int J Cancer 2002;100:690–7.
12. Loffler A, Kufer P, Lutterbuse R, Zettl F, Daniel PT, Schwenken-becher JM, Riethmuller G, Dorken B, Bargou RC. A recombinantbispecific single-chain antibody, CD19xCD3, induces rapid and highlymphoma-directed cytotoxicity by unstimulated T lymphocytes.Blood 2000;95:2098–103.
13. Loffler A, Gruen M, Wuchter C, Schriever F, Kufer P, Dreier T,Hanakam F, Baeuerle PA, Bommert K, Karawajew L, Dorken B,
Bargou RC. Efficient elimination of chronic lymphocytic leukaemia Bcells by autologous T cells with a bispecific anti-CD19/anti-CD3 sin-gle-chain antibody construct. Leukemia 2003;17:900–9.
14. Dreier T, Baeuerle PA, Fichtner I, Grun M, Schlereth B, Lorenczew-ski G, Kufer P, Lutterbuse R, Riethmuller G, Gjorstrup P, Bargou RC.T cell costimulus-independent and very efficacious inhibition oftumor growth in mice bearing subcutaneous or leukemic human B celllymphoma xenografts by a CD19-/CD3- bispecific single-chain anti-body construct. J Immunol 2003;170:4397–402.
15. Hurwitz R, Hozier J, LeBien T, Minowada J, Gajl-Peczalska K,Kubonishi I, Kersey J. Characterization of a leukemic cell line of thepre-B phenotype. Int J Cancer 1979;23:174–80.
16. Stinchcombe JC, Bossi G, Booth S, Griffiths GM. The immunologicalsynapse of CTL contains a secretory domain and membrane bridges.Immunity 2001;15:751–61.
17. Zhu Z, Carter P. Identification of heavy chain residues in a humanizedanti-CD3 antibody important for efficient antigen binding and T cellactivation. J Immunol 1995;155:1903–10.
18. Zhu Z, Lewis GD, Carter P. Engineering high affinity humanizedanti-p185HER2/anti-CD3 bispecific F(ab0)2 for efficient lysis ofp185HER2 overexpressing tumor cells. Int J Cancer 1995;62:319–24.
19. Wimberger P, Xiang W, Mayr D, Diebold J, Dreier T, Baeuerle PA,Kimmig R. Efficient tumor cell lysis by autologous, tumor-resident Tlymphocytes in primary ovarian cancer samples by an EP-CAM-/CD3-bispecific antibody. Int J Cancer 2003;105:241–8.
20. Honemann D, Kufer P, Rimpler MM, Chatterjee M, Friedl S, RiecherF, Bommert K, Dorken B, Bargou RC. A novel recombinant bispecificsingle-chain antibody, bscWue-1 x CD3, induces T-cell-mediatedcytotoxicity towards human multiple myeloma cells. Leukemia 2004;18:636–44.
21. Kufer P, Mack M, Gruber R, Lutterbuse R, Zettl F, Riethmuller G.Construction and biological activity of a recombinant bispecific sin-gle-chain antibody designed for therapy of minimal residual colorectalcancer. Cancer Immunol Immunother 1997;45:193–7.
22. Kufer P, Lutterbuse R, Baeuerle PA. A revival of bispecific antibod-ies. Trends Biotechnol 2004;22:238–44.
23. Chang H, Salma F, Yi QL, Patterson B, Brien B, Minden MD.Prognostic relevance of immunophenotyping in 379 patients withacute myeloid leukemia. Leuk Res 2004;28:43–8.
24. Huang JF, Yang Y, Sepulveda H, Shi W, Hwang I, Peterson PA,Jackson MR, Sprent J, Cai Z. TCR-mediated internalization of pep-tide-MHC complexes acquired by T cells. Science 1999;286:952–4.
104 HOFFMANN ET AL.