signaling from synapse to nucleus - stanford university
TRANSCRIPT

Signaling from synapse to nucleus: the logic behindthe mechanismsKarl Deisseroth�, Paul G Mermelsteiny, Houhui Xiaz§ and Richard W Tsienz#
Signaling from synapse to nucleus is vital for activity-dependent
control of neuronal gene expression and represents a
sophisticated form of neural computation. The nature of specific
signal initiators, nuclear translocators and effectors has become
increasingly clear, and supports the idea that the nucleus is able
to make sense of a surprising amount of fast synaptic information
through intricate biochemical mechanisms. Information transfer
to the nucleus can be conveyed by physical translocation of
messengers at various stages within the multiple signal
transduction cascades that are set in motion by a Ca2þ rise near
the surface membrane. The key role of synapse-to-nucleus
signaling in circadian rhythms, long-term memory, and neuronal
survival sheds light on the logical underpinning of these
signaling mechanisms.
Addresses�Department of Psychiatry and Behavioral Sciences, Stanford University
School of Medicine, Stanford CA 94305, USA
e-mail: [email protected] of Neuroscience, University of Minnesota, 6-145 Jackson
Hall, 321 Church Street South East, Minneapolis, MN 55455, USA
e-mail: [email protected] of Molecular and Cellular Physiology, Stanford University
School of Medicine, Stanford CA 94305, USA§e-mail: [email protected]#e-mail: [email protected]
Current Opinion in Neurobiology 2003, 13:1–12
This review comes from a themed issue on
Signalling mechanisms
Edited by Morgan Sheng and Terrance P Snutch
0959-4388/03/$ – see front matter� 2003 Elsevier Science Ltd. All rights reserved.
DOI 10.1016/S0959-4388(03)00076-X
AbbreviationsBDNF brain-derived neurotrophic factor
CaM calmodulin
CaMK calmodulin-dependent protein kinase
CREB Ca2þ/cAMP responsive element binding protein
DREAM downstream regulatory element antagonistic modulator
NFAT nuclear factor of activated T-cells
NMDAR N-methyl-D-aspartate receptor
NPY neuropeptide-Y
PACAP pituitary adenylyl cyclase activating peptide
PKA protein kinase A
SCN suprachiasmatic nucleus
IntroductionNeuroscientists have long appreciated the computations
that ion channels perform at the surface membranes of
neurons. More recently, it has emerged that the complexity
of this electrical signal processing is rivaled by the intri-
cacies of biochemical signal processing that occur below
the membrane, within elaborate networks of cytoplasmic
signaling proteins. However, only within the past few
years has it become clear that even deeper within the
neuron lies perhaps the most formidable engine of com-
putation of all [1]. The nucleus of the mammalian neuron
creates, shapes, links and responds to these electrical and
biochemical communication streams by actively control-
ling the expression of tens of thousands of genes — a
more diverse repertoire than that presented by any other
known cell type.
Until recently, little was understood about the specific
role of the nucleus, apart from the notion that it could be
involved in the maintenance of long-term changes in
neuronal function [2]; it was not at all obvious how
synapses, as myriad sources of fast electrical signals, could
communicate even a tiny fraction of their rich information
stream to the distant and apparently slow nucleus. It has
now become evident that the nucleus is able to make sense
of a surprising amount of fast synaptic information through
intricate biochemical mechanisms [3–5]. Furthermore, as
we review here, there is now a much clearer understanding
of the crucial role of synapse-to-nucleus signaling in certain
key neuronal processes, namely circadian rhythms, long-
term memory and neuronal survival. This functional and
behavioral level of understanding now anchors our inter-
pretation of the underlying logic of the nuclear signaling
mechanisms employed. An emerging theme as we show
here is that the complexity of neuronal biochemistry may
be necessary to generate signal processing streams (from
the rapid ion channel kinetics that initially shape electrical
information flow to the slower biochemical effectors) that
are specific enough to convey an information-laden signal
to the nucleus.
Many signaling pathways to the nucleusWhat signaling entity physically translocates to the
nucleus to support the transfer of information from the
neuronal cell membrane? In their pioneering paper on
excitation–transcription coupling, Morgan and Curran [6]
put forward a hypothesis for how Ca2þ entry through
L-type Ca2þ channels might activate expression of c-fos,an immediate early gene (IEG; IEGs constitute the first
wave of genes to be turned on, often within minutes, after a
stimulus). They suggested that a calmodulin (CaM)-sen-
sitive kinase would phosphorylate a transcription factor,
initially positioned in the cytoplasm, causing it to move
into the nucleus to activate gene expression. This first
speculation was prescient in broad outline and provocative
1
CONEUR 55
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

in highlighting the following fundamental questions. First,
what are the crucial and specific signal initiators? Second,
what are the nuclear translocators, and why are they
selected for the task? Third and finally, what is the basis
for stimulus specificity and reliable information transfer?
By focusing on the pathways triggered by an intracellular
Ca2þ rise downstream of synaptic activity, we now know
that the signal from the cytoplasm to the nucleus can be
conveyed in several different ways (Figure 1). The trans-
location can take place at various stages within signal
transduction cascades that are set in motion by the Ca2þ
rise. At one extreme, as hypothesized by Morgan and
Curran, the transcription factor itself, the terminal factor
in the biochemical cascades, can move to the nucleus
upon changes in its phosphorylation state. The classic
example is a member of the nuclear factor of activated T-
cells (NFAT) family of transcription factors. Originally
characterized in immune tissue, the NFATc group plays a
crucial role in neuronal plasticity as well as vascular
development and muscular hypertrophy [7]. NFATc4
is expressed within hippocampal neurons, and undergoes
a striking translocation from the cytosol to the nucleus
upon the opening of L-type Ca2þ channels [8]. The trans-
location step is dependent upon calcineurin-mediated
dephosphorylation of NFATc4, which causes the unmask-
ing of multiple nuclear localization signals and the active
transport of the transcription factor through the nuclear
pore complex.
At the other extreme, a mechanism may exist that
transduces directly the free Ca2þ transient that can arise
in the nucleus after neuronal excitation [9]. The tran-
scription factor downstream regulatory element antag-
onistic modulator (DREAM) [10], a Ca2þ binding
protein that contains three active Ca2þ binding motifs
(E-F hands) [11�], is abundant in the nucleus. It has been
proposed that nuclear DREAM remains bound to a
downstream regulatory element (DRE) that acts as a
gene silencer when nuclear Ca2þ is low, but dissociates
upon elevation of Ca2þ causing DRE derepression, acti-
vation of downstream genes such as that which encodes
prodynorphin [10] and attenuation of pain signaling invivo [12�]. It will be interesting to see if DREAM
function can be formally linked to nuclear Ca2þ levels,
and to understand why DREAM is present outside the
nucleus as well as within it.
Are there particular advantages for the signaling mechan-
isms at either one or the other of these extremes? On the
one hand, the nuclear Ca2þ mechanism that is exempli-
fied by DREAM is straightforward, fast and simple, but it
might be a suboptimal choice in general as the nucleus
would suffer from a severe loss of specificity with regards
to the source of Ca2þ and the nature of the initial
membrane signal. On the other hand, using a highly
processed nuclear signal like the transcription factor itself
(as in NFAT translocation) allows for a high level of
signaling specificity, but in placing the bulk of the signal
amplification and processing outside of nucleus more
pressure is placed on nuclear transport mechanisms. As
electrophysiologists know, it is unwise to amplify a signal
before passing it through a channel that can be saturated,
as it could result in serious distortion of the signal.
Activation of DREAM and NFATc4 define the extremes
of information transfer from the plasma membrane to the
nucleus, whereas Ca2þ/cAMP responsive element bind-
ing protein (CREB) signaling mechanisms typically
exemplify the middle ground. In this case, the translo-
cating message is not conveyed by free Ca2þ moving from
the ion channels to the nucleus, or by the transcription
factor CREB, but by intermediate players in the path-
way. In fact, CREB remains constitutively bound to CRE
(Ca2þ/cAMP responsive element) regulatory sites con-
trolling target genes, and becomes activated by phos-
phorylation on its Ser-133 residue (with modulatory
influences exerted by other residues). A fast CaM-depen-
dent kinase cascade and a slow MAP kinase cascade
(likely to be cAMP-modulated) converge on Ser-133
following surface membrane depolarization and Ca2þ
influx, and work together to promote CREB-dependent
gene expression [3,13].
How are these two signals transduced? It is well estab-
lished that in response to physiological synaptic activity in
hippocampal pyramidal neurons, Ca2þ entry through both
N-methyl-D-aspartate receptors (NMDARs) and L-type
Ca2þ channels initiates the nuclear signaling to CREB
[14–16]. Nevertheless, Ca2þ ions can be excluded as the
(Figure 1 Legend) Signaling from the membrane to the nucleus: multiple strategies for information transfer. (a) The transcription factors NFATc4,
CREB and DREAM are all activated following increases in intracellular Ca2þ, yet each relies upon a different mode of information transfer. At rest,
NFATc4 is localized to the cytosol, allowing the transcription factor to be targeted to specific regions of the cell. This would allow for heightened
signaling specificity. Indeed, Ca2þ entry through L-type calcium channels activates this transcription factor preferentially relative to other voltage-
gated Ca2þ channels. Yet, this mode of synapse-to-nucleus signaling is limited in both speed and signal amplification. Conversely, DREAM can be
activated by general rises in intracellular Ca2þ, allowing for rapid information transfer. However, this pathway is limited in regards to signal specificity.CREB activation is typically initiated by nuclear translocation of the calcium sensor CaM or by an activating kinase, providing a combination of the
advantages found for both NFATc4 and DREAM signaling. (b) A snapshot image of the early stages (0–5 min) of activity-dependent gene expression.
Following rises in intracellular Ca2þ, DREAM dissociates from DNA resulting in the lifting of transcriptional repression. Within seconds of Ca2þ entry
through L-type Ca2þ channels and NMDA receptors, CaM translocates to the nucleus, supporting CREB phosphorylation through activation of
CaMKIV. Nearly as rapid, NFATc4 also undergoes translocation to the nucleus following its dephosphorylation by calcineurin (CaN). Other pathways
outlined in (a), including the MAPK/PKA pathways, subsequently begin to exert their influence. Abbreviations, VGCC, voltage-gated calcium channel.
2 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com
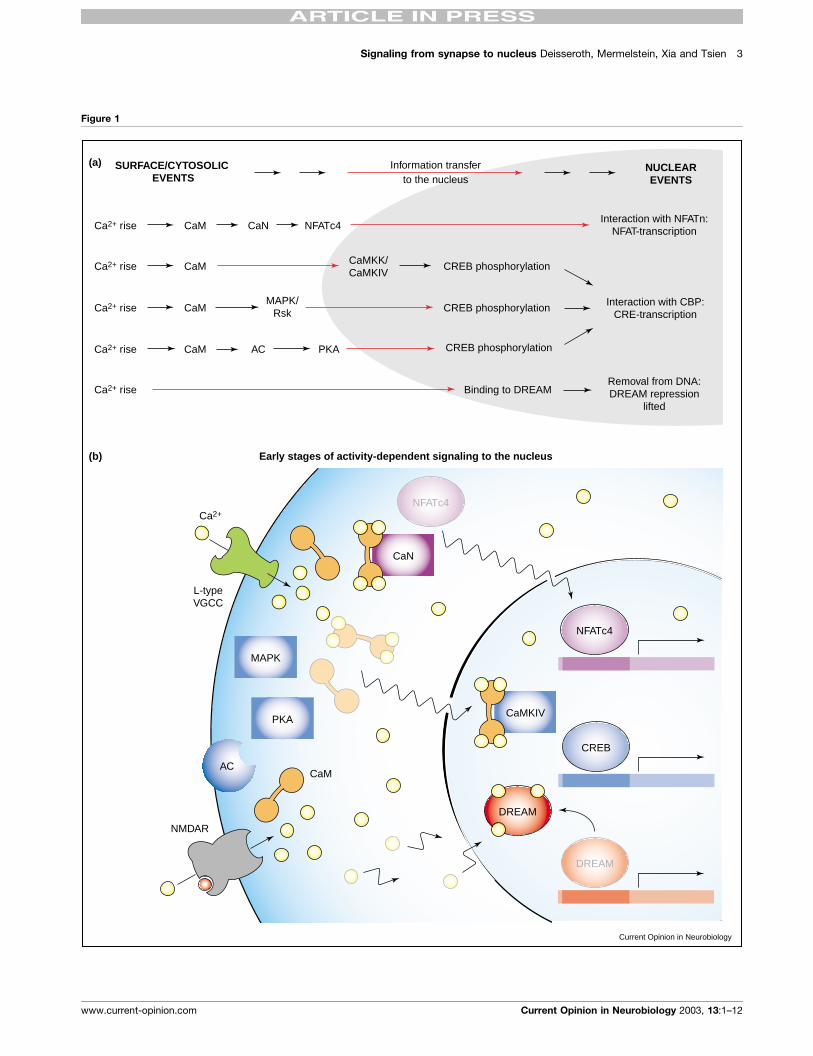
Figure 1
L-typeVGCC
NMDAR
CREB
NFATc4Ca2+
CaM
CaMKIV
CaN
AC
MAPK
PKA
NFATc4
Ca2+ rise
Ca2+ rise
Ca2+ rise
Ca2+ rise
Ca2+ rise
SURFACE/CYTOSOLIC EVENTS
NUCLEAREVENTS
Binding to DREAM
CaM CaMKK/CaMKIV
CREB phosphorylation
CaMMAPK/
Rsk CREB phosphorylation
CaM CaN NFATc4
AC PKA CREB phosphorylationCaM
Removal from DNA:DREAM repression
lifted
Interaction with CBP:CRE-transcription
Interaction with NFATn:NFAT-transcription
Information transferto the nucleus
DREAM
DREAM
(a)
Current Opinion in Neurobiology
(b) Early stages of activity-dependent signaling to the nucleus
Signaling from synapse to nucleus Deisseroth, Mermelstein, Xia and Tsien 3
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

typical nuclear messenger in these cells as CREB activa-
tion at Ser-133 shows marked Ca2þ channel-type speci-
ficity. Working alongside Ca2þ stores, many Ca2þ influx
channels (including L-type, N-type, and P/Q-type Ca2þ
channels as well as NMDA- and AMPA-type glutamate
receptors) can cause a marked elevation in the bulk
cytoplasmic and nuclear Ca2þlevels, and can play impor-
tant roles in the neuron [17]. However, most of these are
ineffective at directly activating CREB, with the excep-
tion of the L-type channel (privileged among the voltage-
gated channels [15,16,18��]) and the NMDA receptor
[14–16]. Therefore, although nuclear Ca2þ elevations
are seen during relatively strong neuronal activity [9]
and may support the signaling in some cases [19,20],
nuclear Ca2þ is not typically sufficient to activate CREB
[15,18��]. Furthermore, it appears that nuclear Ca2þ eleva-
tions are not necessary, as suppression of global cytoplas-
mic and nuclear Ca2þ elevations with intracellular loading
of the Ca2þ chelator EGTA generally does not block Ca2þ-
activated CREB Ser-133 activation [4,14,21��]. These
findings, together with results that demonstrate micro-
meter-scale localization of L-type Ca2þ channels within
the plasma membrane [22��] and a CaM-binding motif on
the L-type channel itself [18��] are both crucial for CREB
phosphorylation, have revealed that Ca2þ typically exerts
its effects on nuclear CREB activation by acting within a
domain <1 mm in radius from its source of entry [14].
The privileged roles of the NMDA receptor and the L-
type channel [14–16] may be attributable to their selec-
tive ability to recruit CaM signaling [15]. While both
channel types can directly bind CaM, both are also
specifically localized at or near synapses in which addi-
tional pools of CaM are present for recruitment. Either or
both mechanisms may underlie the selective ability of
these channels to initiate locally Ca2þ/CaM-activated
MAP kinase signaling to the nucleus [13,18��,21��,23]
and nuclear translocation of CaM [15,20,24,25��]. The
translocation of CaM to the nucleus is dependent upon its
ability to bind to a CaM kinase [20,25��,26]. This binding
would stabilize Ca2þ–CaM, although locally mobilized
CaM could also subsequently make use of Ca2þ from
other sources [19,20], perhaps from nuclear Ca2þstores or
even cytoplasmic Ca2þ waves. Use of ‘supportive’ Ca2þ in
this way would still preserve the specificity of information
carried by the selective synaptic activation of the NMDA
receptor and the L-type channel, as only these channel
types would be able to mobilize the CaM necessary to
utilize efficiently the available Ca2þ. Despite the high
concentrations of detectable CaM in all cellular compart-
ments, free or available CaM is generally limiting in
intracellular biochemical reactions [27]. The resulting
increase in available nuclear CaM results in strong activa-
tion of nuclear CaM-dependent protein kinase kinase
(CaMKK) and its target CaMKIV. These kinases are
ultimately responsible for the early wave of Ser-133
CREB phosphorylation [13,28,29��,30,31]. With stronger
stimulation, the slower MAPK-dependent signaling path-
way comes into play [13]. In this case, Ca2þ/CaM leads to
local activation of the small GTP binding protein Ras,
stimulation and nuclear translocation of mitogen-activated
protein kinase (MAPK) and/or pp90 ribosomal protein S6
kinase (Rsk) [3,18��,32], in some cases with cooperation of
protein kinase A (PKA). In summary, both CaMK and
MAPK pathways to CREB generally use a downstream
Ca2þ effector as the translocating nuclear messenger. This
thereby preserves information about the specific Ca2þ
source, while also leaving ample room for signal amplifica-
tion in the nucleus — a combination of features that have
advantages over the more extreme scenarios.
Clearly then, the transfer of information from the mem-
brane surface of a neuron into the nucleus can be
achieved by physical translocation at various points in
the signal transduction pathway. Understanding the
choice of strategy and the associated information proces-
sing trade-offs can best be attempted in the context of the
functional output of the pathway. Therefore, next we will
review recent work in intact animals describing the dis-
ruption of nuclear signaling pathways and their high-level
behavioral consequences: that is, generation of circadian
rhythms, formation of long-term fear memories, and
survival of brain cells.
Circadian rhythmsThe generation and entrainment of circadian rhythms, of
nearly universal importance in biology, provides an out-
standing example of the functional importance of
synapse-to-nucleus signaling. Many aspects of circadian
rhythm generation are hardwired, in that they occur with-
out specifically requiring neuronal activity, and arise in a
network of transcription factors that generate rhythmicity
from negative feedback signaling. In the suprachiasmatic
nucleus (SCN) of the mammalian hypothalamus, for
example (Figure 2), multiple proteins are involved in
the clockwork mechanism, including dimers involving
the transcription factors of CLOCK and BMAL, and
the period gene products mPer1, mPer2 and mPer3. This
endogenous rhythm is robust but not precise, however,
and numerous other circumstances, including seasonal and
migratory changes in patterns of light and dark, demand
that the cycle be entrainable. In fact, exposure to light
before the end of a subjective night, a signal indicating
that the rhythm needs adjustment, triggers rapid induction
of the genes for mPer1 (within 15–30 min) and mPer2
(within 2 h), leading to an efficient readjustment of the
circadian rhythm.
How are these clock genes activated? In addition to E-box
elements (regulated by the CLOCK/BMAL system),
both mPer1 and mPer2 have activity-sensitive CRE pro-
moter elements that are regulated by the CRE-binding
protein CREB [33]. Evidence is mounting that CREB-
mediated control of these genes referees circadian rhythm
4 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com

cycling and entrainment. First, CREB activation by phos-
phorylation of the Ser-133 residue follows an active
circadian rhythm (peaking near subjective dawn) and
can be directly triggered by entraining stimuli, such as
photic stimulation during darkness [16]. Second, expres-
sion of the CREB-dependent genes c-fos, mPer1 and
mPer2 correlates with CREB activation, and is blocked
when a key modulatory site (Ser-142) on CREB is
mutated [34��]. Third and finally, this CREB mutation
attenuates entrainment of the circadian oscillator with
respect to either phase advances or phase delays [34��].Taken together, these data show that activity-dependent
control of a nuclear transcription factor (CREB) mediates
a very important complex behavior (entrainment of cir-
cadian rhythm).
What are the key design constraints on this activity-
dependent signaling pathway? First, to be useful, the
pathway should be fast by comparison with the circadian
cycle (minutes), although signaling on a scale of seconds
may be unnecessary. It need not resolve closely spaced
stimuli (after all, closely spaced flashes of daylight were
hardly a factor in circadian evolution!). Second, in
responding to electrophysiological events, the pathway
would ideally be sensitive to synaptic input from afferents
that carry sensory information from the retina, but not to
the continually free-running spiking of SCN neurons,
which is intrinsic even in isolated neurons. Third, as
non-photic inputs such as feeding or locomotor activity
modulate entrainment in some cases, the pathway must
allow for the convergence of multiple stimuli, not just
retinal glutamatergic afferents.
Every one of these design requirements seems to be
fulfilled by aspects of synapse-to-nucleus signaling in
SCN neurons. First, phosphorylation of CREB Ser-133
does occur within about 10 min of light flashes or gluta-
matergic stimulation of SCN neurons, followed by mPer1
Figure 2
ZZ
Z Z Z
Outputto SCN
L-typeVGCC
mPer1CREBP P
NMDAR
SCN neuronSynaptic transmissionfrom ganglion cells
S133S142
CLK BMAL
CRE E-Box
mPER1
SCN ACTIVITY
Simple spikes: pacemaker activity
Complex spikes: APs and EPSPs
TIME
CRY
RESULT
Minimal NMDAR,L-type VGCC
activity
Activation of NMDAR and
L-type VGCC:CREB phosph
EFFECT
MinimalmPer1
expression
StrongmPer1
expression
INTERNALCLOCK
Current Opinion in Neurobiology
Input from retina
Activity-dependent regulation of circadian rhythms. Light-induced activation of retinal ganglion cells results in the excitation of SCN neurons. The
increases in synaptic neurotransmission result in the opening of L-type calcium channels and NMDA receptors, directly leading to CREB
phosphorylation and the transcription of several clock genes including mPer1. These changes in gene expression result in the entrainment of the
internal clock to the external environment.
Signaling from synapse to nucleus Deisseroth, Mermelstein, Xia and Tsien 5
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

induction within 15–30 min [34��]. This suggests the
involvement of either the fast CaMKIV signaling pathway
leading to phosphorylation of CREB on Ser-133 or the
slower CaM kinase/MAP kinase-dependent pathway,
both of which can be efficacious at 10 min [13]. In fact,
inhibitors of either CaM kinase or MAP kinase block
CREB activation in SCN neurons [35]; in the future,
circadian rhythmicity and fast signaling to CREB might
also be studied in the SCN of CaMKIV knockout mice.
Second, with regards to selective coupling to particular
electrical events at the membrane, certain pathways of
nuclear signaling in hypothalamic neurons have been
found to respond relatively selectively to membrane
potentials generated by synaptic input in preference to
action potentials alone [36], as in various ganglionic
neurons [37,38] and hippocampal pyramidal neurons
[14,39]. It has become increasingly clear how gene
expression avoids indiscriminate activation by the con-
tinuous tonic spiking of these cells. CREB signaling in
hypothalamic SCN neurons responds to retinal glutama-
tergic afferents through a pathway initiated by NMDA-
type glutamate receptors and voltage-gated L-type Ca2þ
channels. Activation of the NMDARs requires synaptic
glutamate, and therefore needs more than just spikes to
occur. Among the many voltage-gated Ca2þ channels,
only the L-type channel has been found to couple to
mPER1 [40]. If this remains true when the contributions
of the other channels are rigorously examined [15], it
suggests that there is a possibility that L-type channels
help to ‘filter out’ brief single action potentials by virtue
of their voltage- and time-dependent kinetics [39]. This
selectivity would probably not be possible if the nuclear
message were conveyed by Ca2þ ions for example, as
strongly spike-responsive Ca2þ channels (N-type or P/Q-
type) also generate large increases in bulk cytoplasmic
and nuclear Ca2þ. The use of CaM as a synapse-to-
nucleus messenger [15] would avoid this difficulty, a
possibility that needs to be tested in SCN neurons. Third,
signaling in the SCN appears well equipped to integrate
modulatory input from non-photic CNS systems, includ-
ing that of neuropeptide-Y (NPY; provided by afferents
from the thalamus), serotonin, and pituitary adenylyl
cyclase activating peptide (PACAP). All three systems
appear to mediate behavioral influences on circadian
rhythmicity, and are known to use G protein signaling
to the cAMP/PKA pathway as a means of exerting control
over intracellular events (PACAP positively, NPY nega-
tively, and serotonin being capable of both). Indeed,
application of forskolin, a direct activator of the cAMP/
PKA pathway, promotes mPer1 expression in SCN neurons
[41,42], and expression of dominant negative PKA inhibits
CRE-dependent gene expression in SCN neurons [41].
PKA could influence nuclear signaling in several different
ways, for example, by direct phosphorylation of CREB
residue Ser-133, by stimulating a CREB transcriptional
coactivator such as CBP or p300 [43], or by potentiating
MAPK signaling [23].
Equally intriguing is the possibility that PKA-mediated
signaling could act through direct modulation of ion
channels, thereby regulating the ability of L-type Ca2þ
channels to respond to the tonic electrical activity of SCN
neurons. This would support the convergence of diverse
inputs and allow a non-photic input to recruit exactly the
same signaling pathway as a light stimulus, even without
retinally driven synaptic input. Indeed, it is known that
the ability of L-type channels to filter out action poten-
tials is conditional, in that direct potentiation of ion flux
restores some responsiveness to brief depolarizations [39].
It is also likely that spike width broadening or high-
frequency bursts of action potentials would be able to
conditionally overcome this filter [44]. To test the validity
of this idea in SCN circadian rhythmicity, it would be
important to conduct electrophysiological studies of SCN
action potentials and the resulting L-type currents at
378C [45]. Testing at this temperature would avoid the
opposing effects on spike width and channel gating
kinetics that are present under even slightly lower tem-
peratures, and under a range of waveform and modulation
conditions. The potential modulation of action potential
filtering could also be tested in vivo, by using L-type
antagonists and potentiators to see if direct manipulation
of L-type channels alters the behavioral responses to non-
photic inputs through NPY, serotonin or PACAP.
Learning and memory exemplified byfear conditioningMemory formation is another vital high-level behavioral
function that depends on nuclear events [2,46,47]. Again,
knowledge of the behavioral output of the nuclear event
can greatly illuminate our understanding of the logic that
underlies the nuclear signaling. Much attention has been
directed towards the hippocampus, which not only sup-
ports explicit memory formation in tasks like spatial
learning but also contributes crucially to contextual fear
conditioning, in which the animal learns to associate a
complex, generally benign environment with an aversive
stimulus (Figure 3). The amygdala mediates cued fear
memories, formed when an aversive stimulus is asso-
ciated with a single stimulus, such as an auditory tone.
In both cases, subsequent presentation of the harmless
stimulus alone will trigger fear behavior such as freezing.
Neurons in both the amygdala and the hippocampus
display robust activation of CREB during these learning
tasks [25��,48], and in both structures, genetic interfer-
ence with CREB function prevents storage of long-term
fear memories [49,50�]. Additionally, in either one or the
other region, memory deficits associated with reduced
CREB function can be partially overcome by spaced
repeated training, whereas high-level CREB function
can allow long-term memories to form with single training
sessions [50�,51,52�].
Given these similarities, several predictions can be made.
First, that the hippocampus and amygdala would employ
6 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com

a common molecular mechanism of signaling to CREB.
Second, that CREB should respond strongly to synaptic
input that bears incoming processed sensory information
in both cases. Third, that activity-dependent signaling
events that lead to CREB activation should display con-
siderable speed and temporal resolution, as temporal
relationships among events are important in elucidating
their causal relationship. Fourth and finally, that stimuli
that are spaced and repeated, those most appropriate for
associations by the brain should be particularly advan-
taged in activating CREB by comparison with single
stimulus bouts.
In the hippocampus, CREB signaling triggered by phy-
siological electrical activity is indeed selectively depen-
dent on Ca2þ entry through L-type calcium channels and
Figure 3
AMYGDALAHIPPOCAMPUS
Contextual information Cue information
+ Pain/fear
One exposure underintense conditions
Multiple spaced exposures
Strong CaM/CaMKIVCREB phosphorylation
Support from MAPK/PKA
Strong CaM/CaMKIVCREB phosphorylation
Support from MAPK/PKAInactivation of CaN
Facilitation of CaM signaling
Possibly spurious or rarebut too important to ignore
Clearly related andassociation should be
learned
Result
Stimulus
Response
Properconclusion
One or massedexposure
Mild CaM/CaMKIV-mediatedCREB phosphorylation
Spurious or rare, possibly anunimportant association
Current Opinion in Neurobiology
No CREB-mediatedlong-term memory
CREB-mediatedlong-term memory
genetic orpharmacological
alteration in CREB signaling
Activity-dependent fear conditioning within the hippocampus and the amygdala. The pairing of contextual and cue information with an adverse
stimulus results in heightened CREB activation within the hippocampus and the amygdala. Dependent upon the stimulus train, one or more CREB
signaling pathways are recruited to mediate long-term memory. With either genetic or pharmacological manipulation of the CREB signaling pathways,
changes in learning and memory can be achieved.
Signaling from synapse to nucleus Deisseroth, Mermelstein, Xia and Tsien 7
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

NMDA receptors [14–16]. This pattern of source-speci-
ficity confers selective responsiveness to synaptic activity
for the reasons discussed above. The role of these path-
ways for Ca2þ entry in amygdalar CREB activation has
not been tested yet, but L-type Ca2þ channels and
NMDA receptors are both required for amygdalar forma-
tion of long-term fear memories [53,54], which are
CREB-dependent. In both the hippocampus and the
amygdala neuronal activity causes rapid nuclear translo-
cation of CaM and CaMKIV-dependent CREB phosphor-
ylation [15,25��,28,29��,30], further reinforcing the
similarities in CREB signaling. Furthermore, it has been
demonstrated in both structures that the CaM/CaMKIV
pathway is required for establishing CREB-dependent
fear memories [25��,29��]. As noted above, the employ-
ment of a downstream effector of Ca2þ as a nuclear
messenger, rather than free Ca2þ itself, facilitates Ca2þ
source discrimination by the nucleus [2,5,15,18��]. The
use of CaM as the messenger, which is capable of sig-
nificant nuclear translocation within 15 s [20], sacrifices
little in the way of speed.
Interestingly, the CaM nuclear translocation pathway,
widely employed by non-neuronal cell types as well as
neurons [15,24,26,27,55–58], is now also known to be
almost ubiquitous throughout the mammalian brain,
including hippocampus, amygdala, somatosensory cor-
tex, insular cortex, anterior cingulate cortex and cere-
bellum [15,20,25��]. However, an exception has been
found, three groups have determined that there is little
CaM translocation in a substructure of the hippocampal
formation called the dentate gyrus [19,20,25��]. Although
the dentate gyrus is anatomically positioned as the ‘gate-
way’ to the hippocampus proper, its precise role is
unknown. Dentate granule neurons differ from hippo-
campal pyramidal cells in many ways, including their
characteristics of development, regeneration, morphol-
ogy and synaptic plasticity. It would be interesting to find
out if the role of these neurons at the systems level were
somehow linked to unusual features of their subcellular
signaling.
What are the advantages of this pathway? Use of the
CaM/CaMKIV pathway to CREB lends itself well not
just to speed but also to implementing repetitive spaced
stimulus responsiveness. Brief mild stimulation of hip-
pocampal neurons that selectively activates the calmo-
dulin/CaMKIV pathway to CREB [13], also creates a
primed state in which subsequent similar stimuli are
more potent and even faster in their effects on nuclear
CREB [20]. The increased speed of CREB formation
suggests that the priming effect might be attributable to
the nuclear use of persistently translocated CaM. After
nuclear translocation, return of CaM to the cytoplasm
proceeds gradually over about 1 hour [15]. This extended
time course fits reasonably well with behavioral data on
spaced stimuli [51,52�].
As noted earlier, CREB responds to multiple converging
signaling pathways in hippocampal neurons, including a
slower but more prolonged MAPK-mediated activation
pathway that overlaps with the calmodulin/CaMKIV
pathway [3–5,13], and a calcineurin/protein phospha-
tase-1-mediated CREB deactivation pathway [31,59].
Flexible association of aversive stimuli with a wide range
of environmental situations of different importance and
temporal structure may require such complexity of sig-
naling. The MAPK pathway to CREB has poorer tem-
poral resolution than the CAMK pathway (slower in
onset, and more persistent), but it has several interesting
properties, including further prolongation by spaced
repeated stimulation [23] or PKA activation [23,32,60],
and selective responsiveness to stronger stimulation [13].
Indeed, activation of the MAPK pathway can be involved
in forming both amygdalar [61] and hippocampal [62]
fear memories. Similarly, the calcineurin/PP1 pathway
responds to repeated stimuli by potentiating CREB func-
tion — in this case through activity-dependent inactiva-
tion of calcineurin [31,63]. Notably, direct calcineurin
inhibition potentiates memory formation in the amygdala
[64�] and the hippocampus [65�].
Taken together, these data collected in the amygdala and
the hippocampus reveal a clear pattern — stimuli of
greater behavioral significance are endowed with a more
potent means of activating CREB. In this manner, the
nuclear computer capitalizes on the diverse characteris-
tics of its many incoming signaling pathways, including
sensitivity and temporal resolution, to calculate the sta-
tistical significance of candidate associations for storage in
long-term memory.
Control of neuronal survival and deathNeurons receive specific instructions to survive or to die,
the correct choice being crucial for development and
mature functioning of the brain. The decision to die is
particularly interesting, as neurons reach this all-or-none
choice after weighing a wide array of external inputs,
including synaptic events. This is reminiscent of faster
electrophysiological choices made between quiescence
and firing an action potential. The nucleus plays a key
role in the survival/death computation, largely through
activity-dependent regulation of transcription factors
such as CREB, MEF2 and NF-kB. An emerging theme
is the highly conditional influence of these transcription
factors, in that they promote survival in some circum-
stances but not in others.
It has been known for years that stimulation of electrical
activity at mild to rapid rates (for example, 5Hz to 50Hz
electrical stimuli) works through both NMDA receptors
and L-type Ca2þ channels to activate nuclear CREB
[14,15]. Accordingly, activation of CREB would not be
expected to promote cell death in the normal brain. Indeed,
CREB generally favors neuronal survival. Postnatal
8 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com

genetic inactivation of CREB (along with cAMP response
element modulator [CREM]) causes widespread apopto-
sis of CNS neurons [66]. Other studies have shown that a
low level of activation of NMDA receptors or L-type Ca2þ
channels promotes survival of cerebellar granule neurons,
presumably through the activity of CaMKIV and CREB
[67]. CREB also mediates the survival-promoting effects
of brain-derived neurotrophic factor (BDNF) [68,69] and
other trophic factors.
On the other hand, excitatory stimuli that are known to be
highly toxic to mature neurons, such as steady prolonged
application of glutamate, promote general cellular dys-
function and death [70–72]. Furthermore, it has been
suggested that under conditions of very strong and long
excitation, CREB can actually promote apoptosis by
acting through a nuclear p53 cascade [73]. Perhaps this
action of CREB is aimed at eliminating neurons whose
function is deranged by excessive stimulation, thus main-
taining the integrity of CNS function. This may be
reminiscent of the other effects of p53, ‘guardian of the
genome’, which prevents mitosis and potential tumori-
genesis in the setting of DNA damage.
Transcription factors from the MEF2 family [74], espe-
cially MEF2A, -C and -D, also appear to promote survival
of cerebellar granule cells and cerebral cortical neurons in
response to NMDA application or mild depolarization
[75–77]. MEF2 function can be elevated directly by
calcineurin and indirectly by CaMKIV [74], once again
underscoring the importance of CaM signaling to the
nucleus [15,31]. Additional activity-dependent signals
provided by p38 MAP kinases and NFAT converge to
promote MEF2 function [74]. However, a pro-apoptotic
role for modified MEF2 proteins has also been identified
[75,77]. Cleavage of these transcription factors follows
activation of caspases, generating MEF2 fragments cap-
able of DNA binding but lacking transactivation poten-
tial. These dominant-negative factors then strongly
promote apoptosis. Thus, as in the situation with CREB,
the influence of the transcription factor in favoring or
disfavoring survival depends on the cellular context.
The NF-kB family of transcription factors is strongly
expressed in the CNS, especially in the hippocampus
and the cerebellum. They are activated by growth factors
and cytokines as well as by depolarization or glutamate
application, which leads to nuclear translocation and in
general, promotion of neuronal survival [78]. However,
several studies have identified a pro-apoptotic role for
NF-kB. Though considerable controversy remains, the
net effect of NF-kB appears to depend on the type of
stimulation, the degree of NF-kB activation, and the
particular NF-kB subunits that are expressed. Interest-
ingly, beyond simple nuclear translocation [6] of NF-kB
[79] (Figure 1), the p65 subunit of NF-kB can be further
activated by phosphorylation at Ser-276, a residue whose
amino acid sequence environment is nearly identical to
that of CREB Ser-133. It has been proposed [79] that the
site can support modulation of NF-kB function by CaM-
KIV or Rsk kinases, leading to disparate effects on
survival or death that depend on the profile of kinase
activation. Here again, as with CREB and MEF2, we see
an emerging role of nuclear computation in the integra-
tion of rich streams of incoming information to reach a
crucial decision between cell survival and cell death.
Concluding remarks and future prospectsWe have deliberately focused on mechanisms that convey
an initial message from the neuronal surface to a nuclear
transcription factor, but in which the final outcome in
gene expression also depends on multiple additional
signaling events, as illustrated below for the cases of
CREB- and NFAT-dependent transcription. In the case
of CREB, attainment of the Ser-133 activated state is
further modulated by additional activity-dependent
events, including calcineurin-regulated dephosphoryla-
tion of Ser-133 as well as the phosphorylation of Ser-
142 discussed above. Stabilization of the CREB transcrip-
tional complex is likely to be further affected by activity-
dependent regulation of co-activators, including the
CREB binding protein (CBP) and possibly the closely
related p300 [43]. With respect to activation of NFAT-
dependent transcription, required co-activators include
activator protein-1 (AP1) (composed partly by the CREB-
regulated factor c-fos) and MEF2 [80], which is CaM-
regulated in its own right as noted above. Neuronal
activity not only triggers nuclear translocation of NFATc4
but it additionally regulates NFATc4-dependent tran-
scription by delaying nuclear export of NFAT through L-
type channel mediated inhibition of glycogen synthase
kinase 3b [8]. The result is prolonged NFATc4 activation
and increased NFAT-dependent gene expression [8]; this
is analogous to CRE-mediated gene expression when
CREB activity is extended by activity-dependent inhibi-
tion of CREB dephosphorylation [31,63]. Judging by
these examples of signaling complexity, we have only
seen the tip of the iceberg with respect to the conver-
gence or apposition of multiple mechanisms.
Intricate networks of downstream genes represent the
output of nuclear processing. A large number of genes
show activity modulation in neurons, and many of these
have plausible behavioral roles. Within hippocampal
neurons, activation of NFATc4 leads to an upregulation
of inositol 1,4,5-triphosphate receptor type 1 (IP3R1) [8]
and BDNF [81]; both proteins are known to influence
learning and memory [82–84]. CREB likewise drives
BDNF expression [69] as well as c-fos and other factors
implicated in neuronal survival and memory formation. In
circadian rhythm generation, the CREB-regulated mPergenes are well-characterized targets of retinal activity and
have clear behavioral roles. But exactly how the wide
array of activity-regulated genes generate meaningful
Signaling from synapse to nucleus Deisseroth, Mermelstein, Xia and Tsien 9
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

changes in the brain’s neural networks is still not under-
stood. Though many aspects of nuclear computation
remain shrouded in mystery, it is clear that a combination
of in vitro analysis and behavioral investigation can begin
to illuminate the rationale for the unique biochemical
complexity of neurons, and the elegant logic behind the
design of nuclear signaling pathways.
AcknowledgementsRW Tsien is supported by grants from the National Institute of GeneralMedical Sciences (58234), National Institute of Neurological Disorders andStroke (24067) and National Institute of Mental Health (48108). PGMermelstein is supported by a grant from the National Institute ofNeurological Disorders and Stroke (41302), and a grant from the WhitehallFoundation. K Deisseroth and PG Mermelstein contributed equally to themanuscript.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest��of outstanding interest
1. Bito H, Deisseroth K, Tsien RW: Ca2þ-dependent regulationin neuronal gene expression. Curr Opin Neurobiol 1997,7:419-429.
2. Bailey CH, Bartsch D, Kandel ER: Toward a molecular definitionof long-term memory storage. Proc Natl Acad Sci USA 1996,93:13445-13452.
3. West AE, Griffith EC, Greenberg ME: Regulation of transcriptionfactors by neuronal activity. Nat Rev Neurosci 2002, 3:921-931.
4. Bradley J, Finkbeiner S: An evaluation of specificity in activity-dependent gene expression in neurons. Prog Neurobiol 2002,67:469-477.
5. Lonze BE, Ginty DD: Function and regulation of CREB familytranscription factors in the nervous system. Neuron 2002,35:605-623.
6. Morgan JI, Curran T: Role of ion flux in the control of c-fosexpression. Nature 1986, 322:552-555.
7. Crabtree GR, Olson EN: NFAT signaling: choreographing thesocial lives of cells. Cell 2002, 109:S67-S79.
8. Graef IA, Mermelstein PG, Stankunas K, Neilson JR, Deisseroth K,Tsien RW, Crabtree GR: L-type calcium channels and GSK-3regulate the activity of NF-ATc4 in hippocampal neurons.Nature 1999, 401:703-708.
9. Lipscombe D, Madison DV, Poenie M, Reuter H, Tsien RW,Tsien RY: Imaging of cytosolic Ca2þ transients arising fromCa2þ stores and Ca2þ channels in sympathetic neurons.Neuron 1988, 1:355-365.
10. Carrion AM, Link WA, Ledo F, Mellstrom B, Naranjo JR: DREAM isa Ca2þ-regulated transcriptional repressor. Nature 1999,398:80-84.
11.�
Osawa M, Tong KI, Lilliehook C, Wasco W, Buxbaum JD,Cheng HY, Penninger JM, Ikura M, Ames JB: Calcium-regulatedDNA binding and oligomerization of the neuronal calcium-sensing protein, calsenilin/DREAM/KChIP3. J Biol Chem 2001,276:41005-41013.
Osawa et al. [11�] and Cheng et al. [12�] characterized key aspects ofDREAM [10], a nuclear and cytoplasmic protein that can bind Ca2þ ions(Kd � 14 mM). Though nuclear Ca2þ levels probably do not reach highmicromolar concentrations, one of the functions of DREAM is to act as atranscription factor, and it has also been hypothesized to act as a directtarget of nuclear Ca2þ ions. Early demonstrations of the other two mainmechanistic paradigms in Ca2þ-dependent nuclear signaling in neuronswere obtained for translocation of the transcription factor itself [8] andtranslocation of an intermediate messenger [15].
12.�
Cheng HY, Pitcher GM, Laviolette SR, Whishaw IQ, Tong KI,Kockeritz LK, Wada T, Joza NA, Crackower M, Goncalves J
et al.: DREAM is a critical transcriptional repressor for painmodulation. Cell 2002, 108:31-43.
See annotation to Osawa et al. [11�].
13. Wu GY, Deisseroth K, Tsien RW: Activity-dependent CREBphosphorylation: convergence of a fast, sensitive calmodulinkinase pathway and a slow, less sensitive mitogen- activatedprotein kinase pathway. Proc Natl Acad Sci USA 2001,98:2808-2813.
14. Deisseroth K,Bito H,Tsien RW:Signaling from synapse to nucleus:postsynaptic CREB phosphorylation during multiple forms ofhippocampal synaptic plasticity. Neuron 1996, 16:89-101.
15. Deisseroth K, Heist EK, Tsien RW: Translocation of calmodulin tothe nucleus supports CREB phosphorylation in hippocampalneurons. Nature 1998, 392:198-202.
16. Ginty DD, Kornhauser JM, Thompson MA, Bading H, Mayo KE,Takahashi JS, Greenberg ME: Regulation of CREBphosphorylation in the suprachiasmatic nucleus by light and acircadian clock. Science 1993, 260:238-241.
17. Snutch TP, Sutton KG, Zamponi GW: Voltage-dependent calciumchannels — beyond dihydropyridine antagonists. Curr OpinPharmacol 2001, 1:11-16.
18.��
Dolmetsch RE, Pajvani U, Fife K, Spotts JM, Greenberg ME:Signaling to the nucleus by an L-type calcium channel-calmodulin complex through the MAP kinase pathway.Science 2001, 294:333-339.
The studies by Deisseroth et al., Dolmetsch et al., Hardingham et al. andWeick et al. [14,18��,21��,22��] all demonstrate that Ca2þ typically actslocally, near the channels in the surface membrane, to initiate nuclearsignaling. This result was first demonstrated [14] using a combination offast and slow intracellular Ca2þ buffers to show that Ca2þ can act withinabout 1 mm of the channel source to activate nuclear CREB. Similarresults were later obtained focusing on the slow pathway to CREB[21��]. It subsequently became clear that a CaM-binding site directlyon the L-type channel helps to confer the privileged ability of the L-typechannel to signal to nuclear CREB [18��], and that micron-scale localiza-tion of the L-type channel, and therefore micron-scale local signaling, islikewise crucial for signaling to CREB [22��].
19. Hardingham GE, Arnold FJ, Bading H: Nuclear calcium signalingcontrols CREB-mediated gene expression triggered bysynaptic activity. Nat Neurosci 2001, 4:261-267.
20. Mermelstein PG, Deisseroth K, Dasgupta N, Isaksen AL, Tsien RW:Calmodulin priming: nuclear translocation of a calmodulincomplex and the memory of prior neuronal activity. Proc NatlAcad Sci USA 2001, 98:15342-15347.
21.��
Hardingham GE, Arnold FJ, Bading H: A calcium microdomainnear NMDA receptors: on switch for ERK-dependent synapse-to-nucleus communication. Nat Neurosci 2001, 4:565-566.
See annotation for Dolmetsch et al. [18��].
22.��
Weick JP, Groth RD, Isaksen AL, Mermelstein PG: Interactionswith PDZ proteins are required for L-type calcium channels toactivate cAMP response element binding protein-dependentgene expression. J Neurosci 2003, 23:3446-3456.
See annotation for Dolmetsch et al. [18��].
23. Wu GY, Deisseroth K, Tsien RW: Spaced stimuli stabilize MAPKpathway activation and its effects on dendritic morphology.Nat Neurosci 2001, 4:151-158.
24. Wang D, Tolbert LM, Carlson KW, Sadee W: Nuclear Ca2þ/calmodulin translocation activated by mu-opioid (OP3)receptor. J Neurochem 2000, 74:1418-1425.
25.��
Wei F, Qiu CS, Liauw J, Robinson DA, Ho N, Chatila T, Zhuo M:Calcium calmodulin-dependent protein kinase IV is requiredfor fear memory. Nat Neurosci 2002, 5:573-579.
The studies by Wei et al. and Kang et al. [25��,29��] demonstrated thecrucial importance of the calmodulin–CaMKIV pathway to CREB inmemory formation in the amygdala and hippocampus.
26. Liao B, Paschal BM, Luby-Phelps K: Mechanism of Ca2þ-dependent nuclear accumulation of calmodulin. Proc Natl AcadSci USA 1999, 96:6217-6222.
27. Teruel MN, Chen W, Persechini A, Meyer T: Differential codes forfree Ca2þ-calmodulin signals in nucleus and cytosol. Curr Biol2000, 10:86-94.
10 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com

28. Ribar TJ, Rodriguiz RM, Khiroug L, Wetsel WC, Augustine GJ,Means AR: Cerebellar defects in Ca2þ/calmodulin kinase IV-deficient mice. J Neurosci 2000, 20:RC107:1-5.
29.��
Kang H, Sun LD, Atkins CM, Soderling TR, Wilson MA, Tonegawa S:An important role of neural activity-dependent CaMKIVsignaling in the consolidation of long-term memory. Cell 2001,106:771-783.
See annotation for Wei et al. [25��].
30. Ho N, Liauw JA, Blaeser F, Wei F, Hanissian S, Muglia LM,Wozniak DF, Nardi A, Arvin KL, Holtzman DM et al.: Impairedsynaptic plasticity and cAMP response element-bindingprotein activation in Ca2þ/calmodulin-dependent proteinkinase type IV/Gr- deficient mice. J Neurosci 2000,20:6459-6472.
31. Bito H, Deisseroth K, Tsien RW: CREB phosphorylation anddephosphorylation: a Ca2þ- and stimulus duration-dependentswitch for hippocampal gene expression. Cell 1996,87:1203-1214.
32. Impey S, Obrietan K, Wong ST, Poser S, Yano S, Wayman G,Deloulme JC, Chan G, Storm DR: Cross talk between ERK andPKA is required for Ca2þ stimulation of CREB- dependenttranscription and ERK nuclear translocation. Neuron 1998,21:869-883.
33. King DP, Takahashi JS: Molecular genetics of circadian rhythmsin mammals. Annu Rev Neurosci 2000, 23:713-742.
34.��
Gau D, Lemberger T, von Gall C, Kretz O, Le Minh N, Gass P,Schmid W, Schibler U, Korf HW, Schutz G: Phosphorylation ofCREB Ser142 regulates light-induced phase shifts of thecircadian clock. Neuron 2002, 34:245-253.
This paper provided the first direct causal evidence linking CREB mod-ulation to the important behavioral process of circadian rhythm entrain-ment. A crucial aspect of this link may lie in channel biophysics. Luckmanet al. [36], Mermelstein et al. [39] and Deisseroth et al. [14] demonstratedthat in hypothalamic and hippocampal neurons, nuclear signaling canfilter out action potentials and respond selectively to synaptic potentials.A biophysical explanation was provided [39] linked to the activation gatingkinetics of L-type calcium channels, a property that may be crucial in thebehavioral roles of CREB (Figure 2).
35. Schurov IL, Hepworth TJ, Hastings MH: Dopaminergicsignalling in the rodent neonatal suprachiasmatic nucleusidentifies a role for protein kinase A and mitogen-activatedprotein kinase in circadian entrainment. Eur J Neurosci 2002,15:223-232.
36. Luckman SM, Dyball RE, Leng G: Induction of c-fos expression inhypothalamic magnocellular neurons requires synapticactivation and not simply increased spike activity. J Neurosci1994, 14:4825-4830.
37. Chalazonitis A, Zigmond RE: Effects of synaptic and antidromicstimulation on tyrosine hydroxylase activity in the rat superiorcervical ganglion. J Physiol 1980, 300:525-538.
38. Brosenitsch TA, Salgado-Commissariat D, Kunze DL, Katz DM: Arole for L-type calcium channels in developmental regulation oftransmitter phenotype in primary sensory neurons. J Neurosci1998, 18:1047-1055.
39. Mermelstein PG, Bito H, Deisseroth K, Tsien RW: Criticaldependence of cAMP response element-binding proteinphosphorylation on L-type calcium channels supports aselective response to EPSPs in preference to action potentials.J Neurosci 2000, 20:266-273.
40. Akiyama M, Minami Y, Nakajima T, Moriya T, Shibata S:Calcium and pituitary adenylate cyclase-activating polypeptideinduced expression of circadian clock gene mPer1 in themouse cerebellar granule cell culture. J Neurochem 2001,78:499-508.
41. Obrietan K, Impey S, Smith D, Athos J, Storm DR: Circadianregulation of cAMP response element-mediated geneexpression in the suprachiasmatic nuclei. J Biol Chem 1999,274:17748-17756.
42. Travnickova-Bendova Z, Cermakian N, Reppert SM, Sassone-Corsi P: Bimodal regulation of mPeriod promoters by CREB-dependent signaling and CLOCK/BMAL1 activity. Proc NatlAcad Sci USA 2002, 99:7728-7733.
43. Deisseroth K, Tsien RW: Dynamic multiphosphorylationpasswords for activity-dependent gene expression.Neuron 2002, 34:179-182.
44. Dudek SM, Fields RD: Somatic action potentials are sufficientfor late-phase LTP-related cell signaling. Proc Natl Acad SciUSA 2002, 99:3962-3967.
45. Liu Z, Otsu Y, Murphy TH: Analysis of the preference of theL-type VSCCs for different firing patterns in HEK cells andhippocampal neurons. Soc Neurosci Abstr 1999, 707:17.
46. Schafe GE, Nader K, Blair HT, LeDoux JE: Memory consolidationof Pavlovian fear conditioning: a cellular and molecularperspective. Trends Neurosci 2001, 24:540-546.
47. Milner B, Squire LR, Kandel ER: Cognitive neuroscience and thestudy of memory. Neuron 1998, 20:445-468.
48. Impey S, Smith DM, Obrietan K, Donahue R, Wade C, Storm DR:Stimulation of cAMP response element (CRE)-mediatedtranscription during contextual learning. Nat Neurosci 1998,1:595-601.
49. Bourtchuladze R, Frenguelli B, Blendy J, Cioffi D, Schutz G,Silva AJ: Deficient long-term memory in mice with a targetedmutation of the cAMP-responsive element-binding protein.Cell 1994, 79:59-68.
50.�
Kida S, Josselyn SA, de Ortiz SP, Kogan JH, Chevere I, MasushigeS, Silva AJ: CREB required for the stability of new andreactivated fear memories. Nat Neurosci 2002, 5:348-355.
This study, along with that of Kogan et al. and Josselyn et al. [51,52�],demonstrates the importance of CREB in fear memory formation in theamygdala and the hippocampus, as well as the use of spaced trainingprotocols to overcome insufficient CREB activation and lead to robustmemory formation in both structures.
51. Kogan JH, Frankland PW, Blendy JA, Coblentz J, Marowitz Z,Schutz G, Silva AJ: Spaced training induces normal long-termmemory in CREB mutant mice. Curr Biol 1997, 7:1-11.
52.�
Josselyn SA, Shi C, Carlezon WA Jr, Neve RL, Nestler EJ, Davis M:Long-term memory is facilitated by cAMP response element-binding protein overexpression in the amygdala. J Neurosci2001, 21:2404-2412.
See annotation to Kida et al. [50�].
53. Bauer EP, Schafe GE, LeDoux JE: NMDA receptors and L-typevoltage-gated calcium channels contribute to long-termpotentiation and different components of fear memoryformation in the lateral amygdala. J Neurosci 2002,22:5239-5249.
54. Weisskopf MG, Bauer EP, LeDoux JE: L-type voltage-gatedcalcium channels mediate NMDA-independent associativelong-term potentiation at thalamic input synapses to theamygdala. J Neurosci 1999, 19:10512-10519.
55. Vendrell M, Pujol MJ, Tusell JM, Serratosa J: Effect of differentconvulsants on calmodulin levels and proto-oncogene c-fosexpression in the central nervous system. Brain Res Mol BrainRes 1992, 14:285-292.
56. Milikan JM, Bolsover SR: Use of fluorescently labelledcalmodulins as tools to measure subcellular calmodulinactivation in living dorsal root ganglion cells. Pflugers Arch 2000,439:394-400.
57. Luby-Phelps K, Hori M, Phelps JM, Won D: Ca2þ-regulateddynamic compartmentalization of calmodulin in living smoothmuscle cells. J Biol Chem 1995, 270:21532-21538.
58. Craske M, Takeo T, Gerasimenko O, Vaillant C, Torok K,Petersen OH, Tepikin AV: Hormone-induced secretory andnuclear translocation of calmodulin: oscillations of calmodulinconcentration with the nucleus as an integrator. Proc Natl AcadSci USA 1999, 96:4426-4431.
59. Sala C, Rudolph-Correia S, Sheng M: Developmentally regulatedNMDA receptor-dependent dephosphorylation of cAMPresponse element-binding protein (CREB) in hippocampalneurons. J Neurosci 2000, 20:3529-3536.
60. Martin KC, Michael D, Rose JC, Barad M, Casadio A, Zhu H, KandelER: MAP kinase translocates into the nucleus of the
Signaling from synapse to nucleus Deisseroth, Mermelstein, Xia and Tsien 11
www.current-opinion.com Current Opinion in Neurobiology 2003, 13:1–12

presynaptic cell and is required for long-term facilitation inAplysia. Neuron 1997, 18:899-912.
61. Schafe GE, Atkins CM, Swank MW, Bauer EP, Sweatt JD, LeDouxJE: Activation of ERK/MAP kinase in the amygdala is requiredfor memory consolidation of Pavlovian fear conditioning.J Neurosci 2000, 20:8177-8187.
62. Adams JP, Sweatt JD: Molecular psychology: roles for the ERKMAP kinase cascade in memory. Annu Rev Pharmacol Toxicol2002, 42:135-163.
63. Liu F, Graybiel AM: Activity-regulated phosphorylation ofcAMP response element binding protein in the developingstriatum: implications for patterning the neurochemicalphenotypes of striatal compartments. Dev Neurosci 1998,20:229-236.
64.�
Lin CH, Lee CC, Gean PW: Involvement of a calcineurin cascadein amygdala depotentiation and quenching of fear memory.Mol Pharmacol 2003, 63:44-52.
This study, and that of Malleret et al. shows that calcineurin inhibition canpromote memory formation in the amygdala [64�] and the hippocampus[65�], a behavioral outcome that had been predicted earlier based on thepotentiating effects of calcineurin inhibition on CREB function [31,63].
65.�
Malleret G, Haditsch U, Genoux D, Jones MW, Bliss TV, VanhooseAM, Weitlauf C, Kandel ER, Winder DG, Mansuy IM: Inducible andreversible enhancement of learning, memory, and long-termpotentiation by genetic inhibition of calcineurin. Cell 2001,104:675-686.
See annotation for Lin et al. [64�].
66. Mantamadiotis T, Lemberger T, Bleckmann SC, Kern H, Kretz O,Martin Villalba A, Tronche F, Kellendonk C, Gau D, Kapfhammer Jet al.: Disruption of CREB function in brain leads toneurodegeneration. Nat Genet 2002, 31:47-54.
67. See V, Boutillier AL, Bito H, Loeffler JP: Calcium/calmodulin-dependent protein kinase type IV (CaMKIV) inhibits apoptosisinduced by potassium deprivation in cerebellar granuleneurons. FASEB J 2001, 15:134-144.
68. Bonni A, Brunet A, West AE, Datta SR, Takasu MA, Greenberg ME:Cell survival promoted by the Ras-MAPK signaling pathway bytranscription-dependent and -independent mechanisms.Science 1999, 286:1358-1362.
69. Shieh PB, Hu SC, Bobb K, Timmusk T, Ghosh A: Identification of asignaling pathway involved in calcium regulation of BDNFexpression. Neuron 1998, 20:727-740.
70. Hardingham GE, Bading H: Coupling of extrasynaptic NMDAreceptors to a CREB shut-off pathway is developmentallyregulated. Biochim Biophys Acta 2002, 1600:148-153.
71. Hardingham GE, Fukunaga Y, Bading H: Extrasynaptic NMDARsoppose synaptic NMDARs by triggering CREB shut-off and celldeath pathways. Nat Neurosci 2002, 5:405-414.
72. Yu SP, Yeh C, Strasser U, Tian M, Choi DW: NMDA receptor-mediated Kþ efflux and neuronal apoptosis. Science 1999,284:336-339.
73. Vyas S, Biguet NF, Michel PP, Monaco L, Foulkes NS, Evan GI,Sassone-Corsi P, Agid Y: Molecular mechanisms of neuronalcell death: implications for nuclear factors responding to cAMPand phorbol esters. Mol Cell Neurosci 2002, 21:1-14.
74. McKinsey TA, Zhang CL, Olson EN: MEF2: a calcium-dependentregulator of cell division, differentiation and death.Trends Biochem Sci 2002, 27:40-47.
75. Okamoto S, Li Z, Ju C, Scholzke MN, Mathews E, Cui J,Salvesen GS, Bossy-Wetzel E, Lipton SA: Dominant-interferingforms of MEF2 generated by caspase cleavage contribute toNMDA-induced neuronal apoptosis. Proc Natl Acad Sci USA2002, 99:3974-3979.
76. Mao Z, Bonni A, Xia F, Nadal-Vicens M, Greenberg ME: Neuronalactivity-dependent cell survival mediated by transcriptionfactor MEF2. Science 1999, 286:785-790.
77. Li M, Linseman DA, Allen MP, Meintzer MK, Wang X, Laessig T,Wierman ME, Heidenreich KA: Myocyte enhancer factor 2A and2D undergo phosphorylation and caspase- mediateddegradation during apoptosis of rat cerebellar granuleneurons. J Neurosci 2001, 21:6544-6552.
78. Mattson MP, Culmsee C, Yu Z, Camandola S: Roles of nuclearfactor kappaB in neuronal survival and plasticity. J Neurochem2000, 74:443-456.
79. Vermeulen L, De Wilde G, Notebaert S, Vanden Berghe W,Haegeman G: Regulation of the transcriptional activity of thenuclear factor-kappaB p65 subunit. Biochem Pharmacol 2002,64:963-970.
80. Blaeser F, Ho N, Prywes R, Chatila TA: Ca2þ-dependent geneexpression mediated by MEF2 transcription factors.J Biol Chem 2000, 275:197-209.
81. Groth RD, Mermelstein PG: Induction of NFAT-dependenttranscription underlies BDNF-induced synaptic plasticity inhippocampal neurons. Soc Neurosci Abstr 2002, 151:15.
82. Inoue T, Kato K, Kohda K, Mikoshiba K: Type 1 inositol 1,4,5-trisphosphate receptor is required for induction of long-termdepression in cerebellar Purkinje neurons. J Neurosci 1998,18:5366-5373.
83. Poo MM: Neurotrophins as synaptic modulators. Nat RevNeurosci 2001, 2:24-32.
84. Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS,Bertolino A, Zaitsev E, Gold B, Goldman D, Dean M et al.: TheBDNF val66met polymorphism affects activity-dependentsecretion of BDNF and human memory and hippocampalfunction. Cell 2003, 112:257-269.
12 Signalling mechanisms
Current Opinion in Neurobiology 2003, 13:1–12 www.current-opinion.com