sistema imune inato dos artrópodes: uma abordagem...
TRANSCRIPT

Sistema imune inato dos artrópodes: uma abordagem comparativa
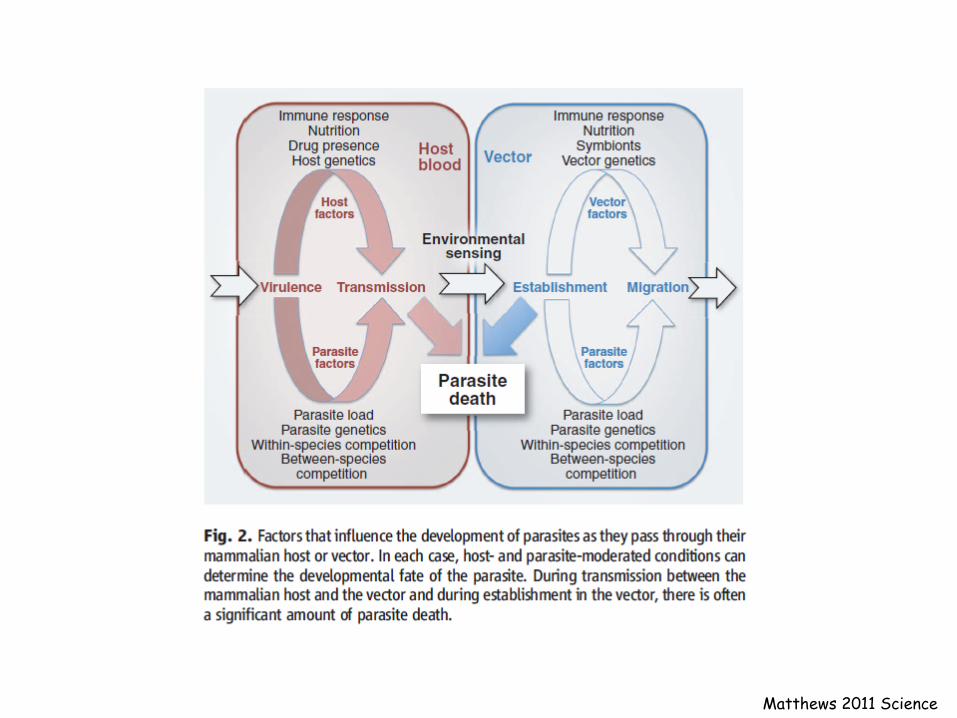
Matthews 2011 Science

The Innate immunity, in addition to be the first line of defense in vertebrates, to stimulate and orient the adaptive immune response
The Innate immunity is the only immune response of invertebrates

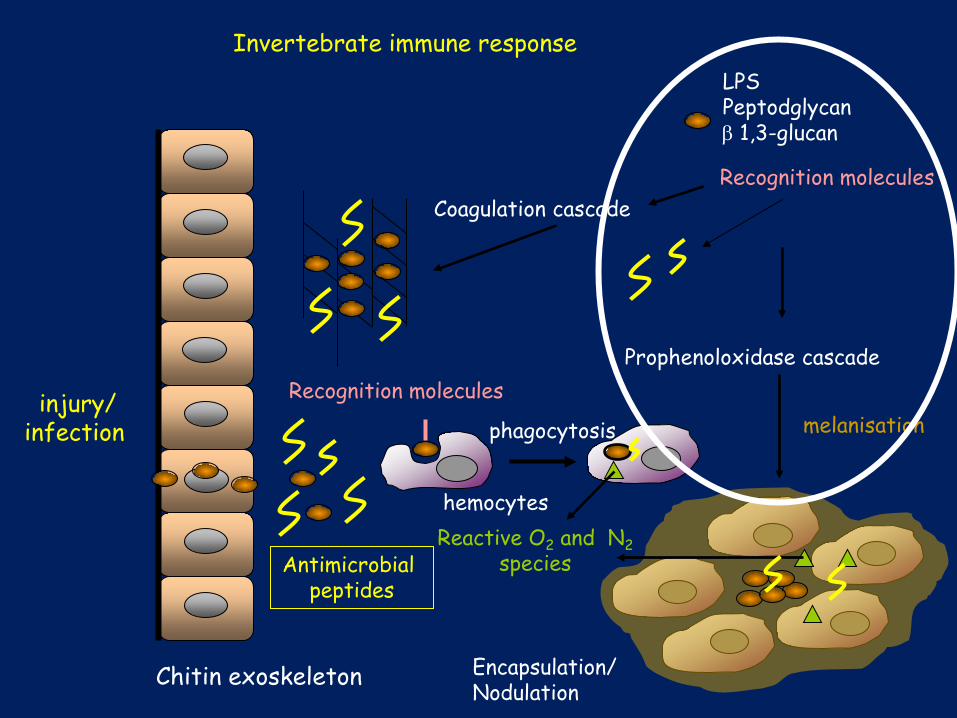
Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response (hemocoel)
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides
protease inhibitors

Innate immunity (Gut)
Oxidative stress
Antimicrobial peptides
protease inhibitors
Gut epithelia

Proteolytic Cascades -PO activation -Coagulation -AMP synthesis
Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides

Schematic representation of the prophenolxidase cascade activation (Royet et al. 2004, Mol Immunol, 41: 1063)
PHENOL
QUINONE

Plasmodium berghei ookinetes crossing the midgut epithelim
Christophides 2004 Immunol Review 198:127

Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides

Coagulation cascade
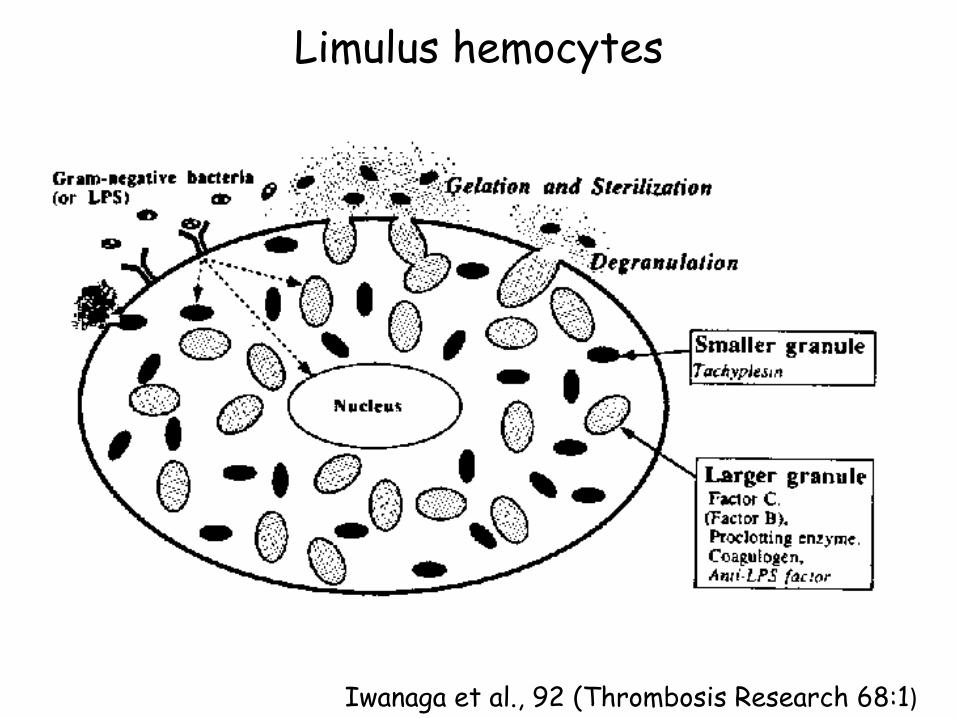
Limulus hemocytes
Iwanaga et al., 92 (Thrombosis Research 68:1)

Factor B
Factor C'
Factor B'
Clotting Enzyme
Factor G
β 1,3 glucan
Factor G'
Coagulogen Coagulin
Proclotting Enzyme
LICI-2
LICI-3
Coagulation cascade (Iwanaga, 2002. Curr Opin Immunol, 14:87 LPS
Factor C
LICI-1
Limulus

Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides

Recognition molecules
TEP (complement-like) PGRP-S PGRP-L
GNBP/β GRP Lectins

TEP1 (Blandin, 2004 Cell)

Science, 2004, 303:2030
Osta et al, 2004 Science

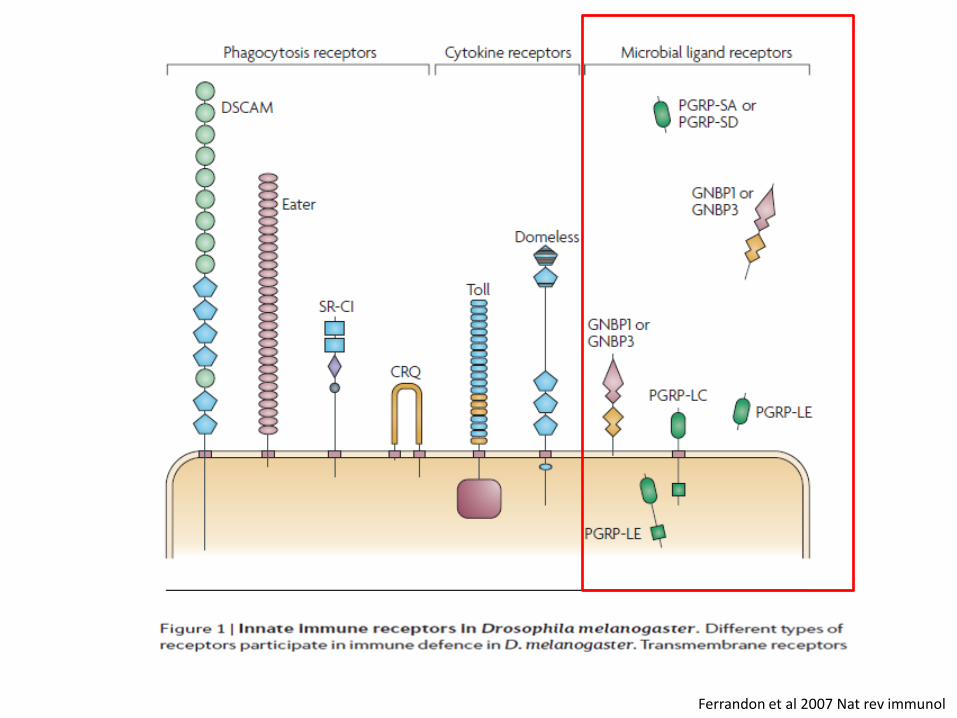
Ferrandon et al 2007 Nat rev immunol

Ferrandon et al 2007 Nat rev immunol
metchnikowin
Antifungal AMP

Ferrandon et al 2007 Nat rev immunol
Cecropins, attacins, defensin, drosocin
Anti-Gram negative AMP, except defensin (gram +)

Park & Lee. 2012 Insect Immuno (Insect Mol Biol and Biochem ed Gilbert)

Park & Lee. 2012 Insect Immuno (Insect Mol Biol and Biochem ed Gilbert)

SCIENCE VOL 319 8 FEBRUARY 2008

Bae et al. 2010 Trends Immunol



Medzhitov, Preston-Hurlburt and Janeway (1997). The successful cloning of a human homologue of Drosophila Toll was reported, and this Toll-like receptor (TLR) was shown to activate NF-kB17. The immune relevance of this TLR was dramatically highlighted when Poltorak, Beutler et al 1998 and Quereshi et al. 1999 discovered that the lps mutation in mice, which abolishes the response to bacterial LPS,corresponded to a loss-of-function mutation in this receptor
Lemaitre, Hoffmann et al (1996). A striking result was that loss-of-function mutations in the Toll receptor compromised the survival of flies faced with fungal infection and the challenge-dependent transcription of the antifungal peptide Drosomycin.
(Hoffmann 2003 Nature).
Nusslein-Volhard et al. (1980). The gene dorsal (member of the NF-kB family) from Drosophila had been genetically identified as a regulator of dorsoventral patterning in the early embryo. It was also known that in the embryo, dissociation of the Dorsal protein from its inhibitor, the Cactus protein, was dependent on activation of the transmembrane receptor Toll by a proteolytically cleaved form of a cytokine-like protein, Spaetzle
Engstrom, Hultmark et al. (1993) and Kappler, Hoffmann et al (1993). The promoter regions of the antimicrobial peptide genes from Drosophila contain nucleotide motifs similar to mammalian binding sites for NF-kB/Rel proteins. Mutations of these sequences in the early 1990s abolished the immune-inducibility of the corresponding genes in reporter fly lines9,10.
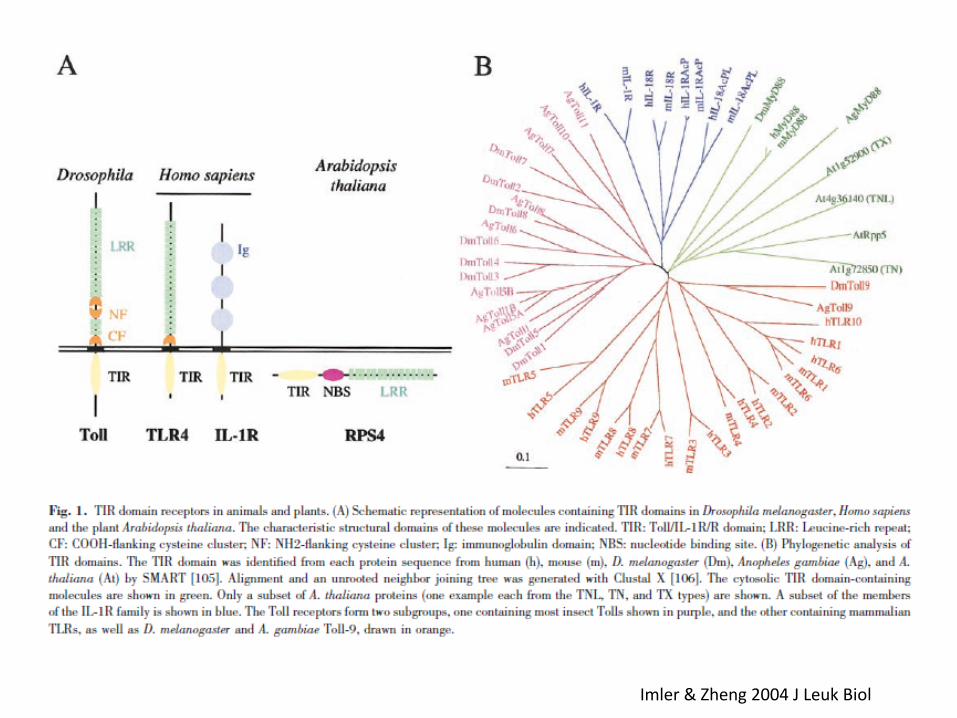
Imler & Zheng 2004 J Leuk Biol
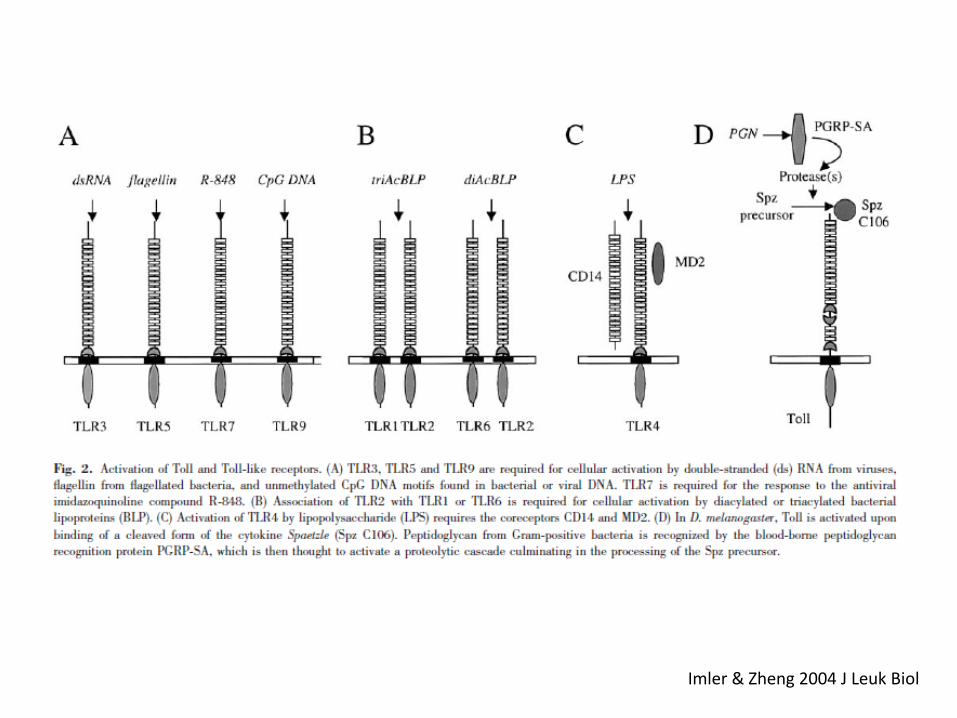
Imler & Zheng 2004 J Leuk Biol

Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides

Reactive oxygen species
Phagocytosis
Melanisation
NADPH oxidase complexe
O2 •O2
- NADPH NADP+
pH 7.0
H2O2 •O2- 2H+ + 2O2
+
Cl- ClO-
OH• Fe3+
hypochloride
hidroxyl superoxide
hydrogen peroxyde

ERO production by hemocytes from B. microplus
Pereira et al. (2001) Experimental Parasitology 99: 66–72
•O2-
H2O2
M. luteus zimozan
E. coli LPS

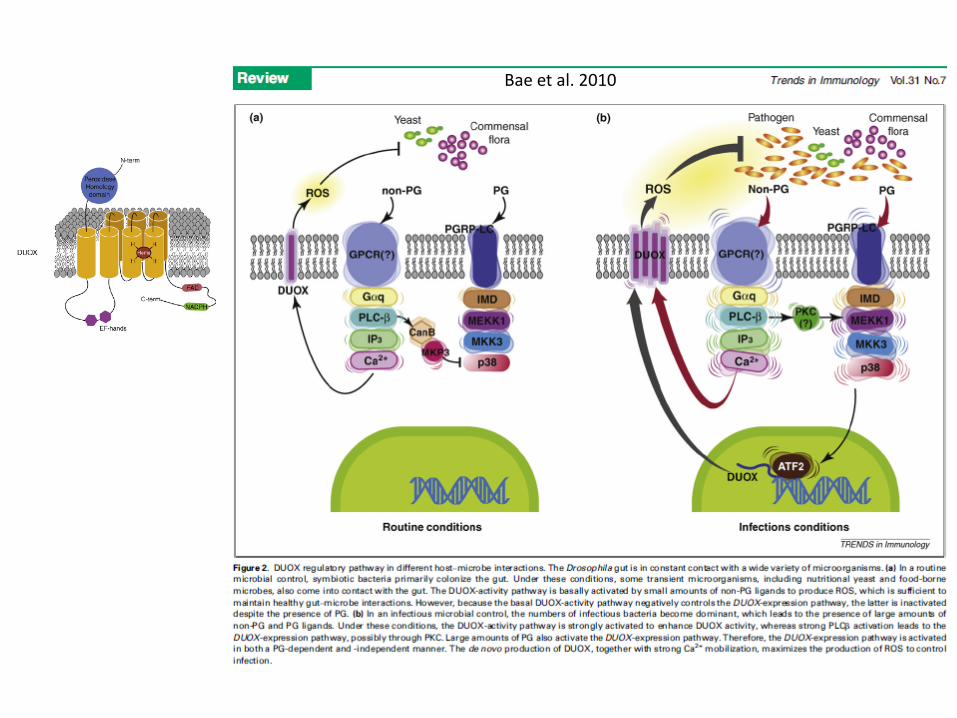
Bae et al. 2010

Christophides 2004 Immunol Review 198:127
Mosquito immune responses
Christophides 2004 Immunol Review 198:127

Science 2010

Science 2010

Science 2011


In this study we assessed the reciprocal interactions between the mosquito's midgut microbiota and dengue virus infection that are, to a large extent, mediated by the mosquito's innate immune system. We observed a marked decrease in susceptibility to dengue virus infection when mosquitoes harbored certain field-derived bacterial isolates in their midgut. Transcript abundance analysis of selected antimicrobial peptide genes suggested that the mosquito's microbiota elicits a basal immune activity that appears to act against dengue virus infection. Conversely, the elicitation of the mosquito immune response by dengue virus infection itself influences the microbial load of the mosquito midgut. In sum, we show that the mosquito's microbiota influences dengue virus infection of the mosquito, which in turn activates its antibacterial responses.

Encapsulation/ Nodulation
Chitin exoskeleton
Coagulation cascade
injury/ infection
Invertebrate immune response
melanisation
Prophenoloxidase cascade
LPS Peptodglycan β 1,3-glucan
phagocytosis
Reactive O2 and N2 species
Recognition molecules
hemocytes
Recognition molecules
Antimicrobial peptides

Antimicrobial peptides (AMPs) are a key element for innate immunity

1942: Balls & Hale identificaram o primeiro PAM em plantas (tionina) 1947: Matrick & Hirsch identificaram nisina, um PAM produzido por Lactococcus lactis 1980: Grupo do Hans Boman identificaram o primeiro PAM em insetos: cecropina 1983: Lehrer & Selsted identificaram defensinas em macrófagos de coelhos
PAMs - Histórico
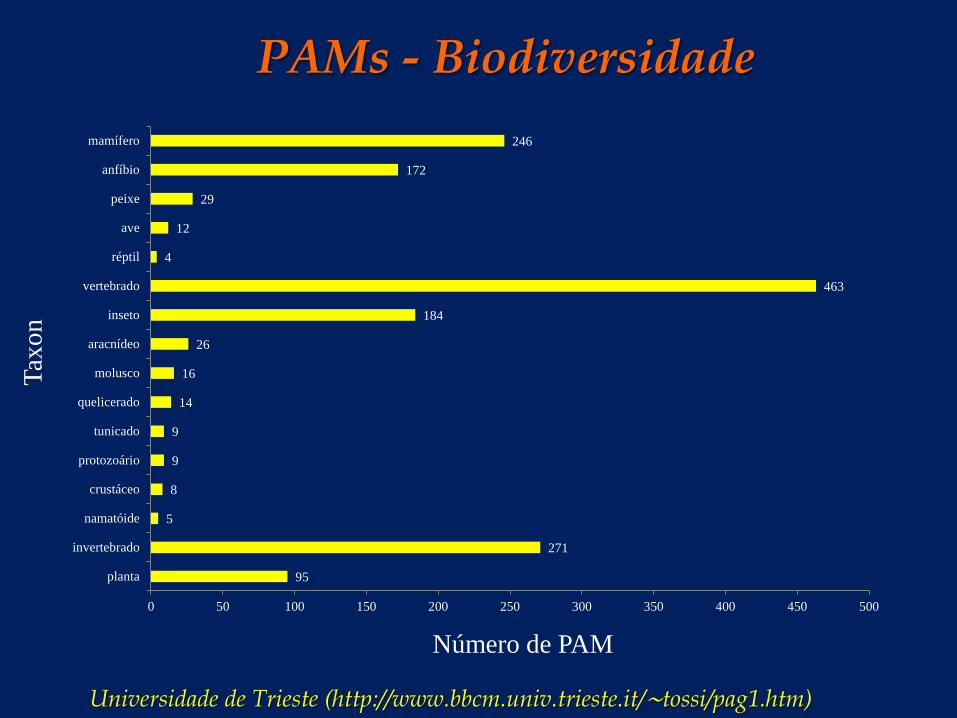
PAMs - Biodiversidade
Universidade de Trieste (http://www.bbcm.univ.trieste.it/∼tossi/pag1.htm)
95
271
5
8
9
9
14
16
26
184
463
4
12
29
172
246
planta
invertebrado
namatóide
crustáceo
protozoário
tunicado
quelicerado
molusco
aracnídeo
inseto
vertebrado
réptil
ave
peixe
anfíbio
mamífero
0 50 100 150 200 250 300 350 400 450 500
Número de PAM
Taxo
n

Saliva histatina, β - defensina
Rins β – defensina, hepicidina
Linfócitos α - defensina
Intestino α - defensina
Pele β - defensina
Trato urogenital α - & β - defensinas
Vias aéreas β - defensina, catelicidina
Distribuição tecidual dos PAMs humano

Distribuição tecidual dos PAMs em insetos
Aparelho bucal drosomicina metchnikowina defensina
Glândula salivar drosomicina
cálice & oviduto drosocina cecropina
Receptáculo seminal & espermateca drosomicina defensina cecropina
Traquéia drosomicina Instestino médio
diptericina atacina
GFP
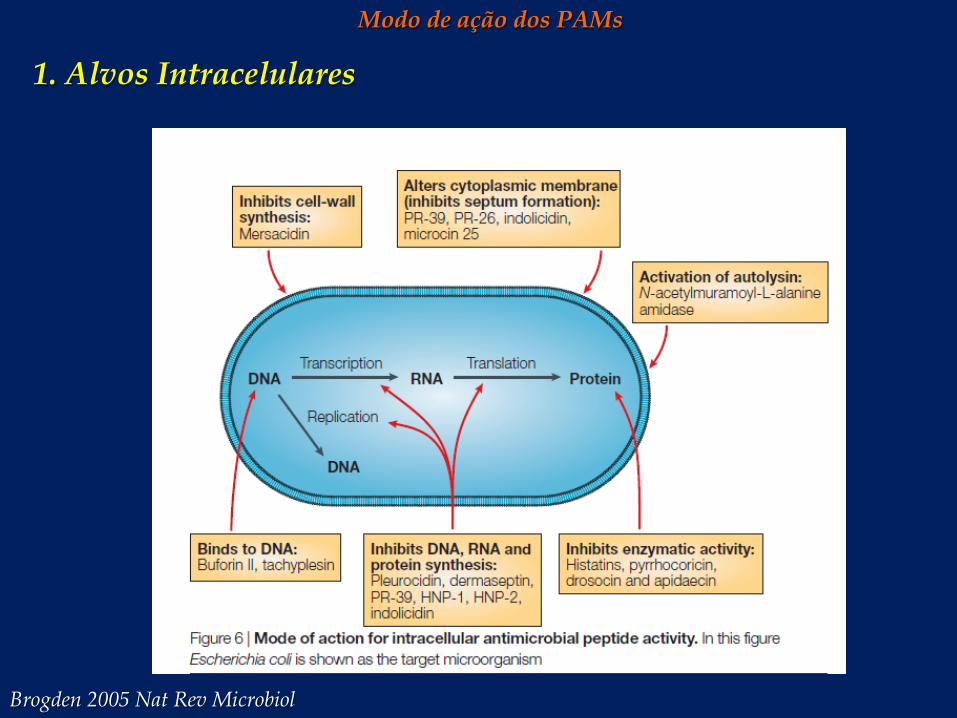
Modo de ação dos PAMs
1. Alvos Intracelulares
Brogden 2005 Nat Rev Microbiol

A: Atração eletrostática estruturação em α-hélice
B: Aumento na [PAM]
C: Agregação pontual desestabilização, permeabilização
e efluxo do soluto
Shai 1999 BBActa
Modo de ação dos PAMs 2a. Permeabilização de membrana (carpet-like)
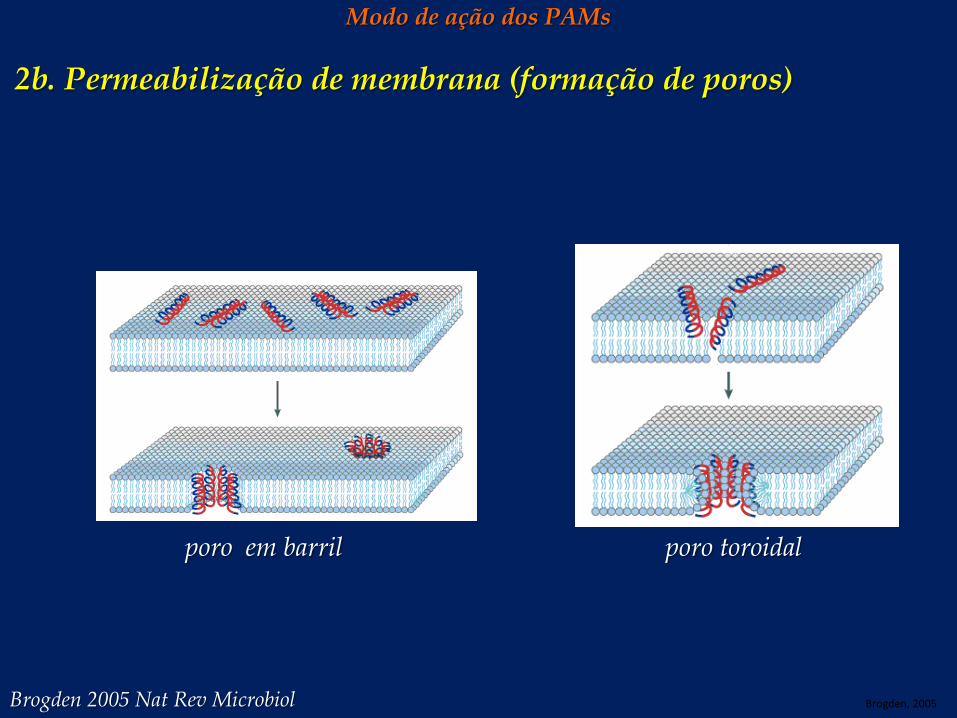
Modo de ação dos PAMs 2b. Permeabilização de membrana (formação de poros)
poro em barril poro toroidal
Brogden, 2005 Brogden 2005 Nat Rev Microbiol

Jenssen & Hancock 2010 Methods Mol Biol
Propriedades imuno-modulatórias dos PAMs
A e B. Modulam a resposta induzida por TLR; C. Atuam na diferenciação de células dendríticas;
D. e E. Atuam na maturação e diferenciação dos monócitos a macrófagos; G. Promovem a migração de leucócitos para o local da infecção; H. Promovem a expressão de moléculas co-estimulatórias pelas células dendríticas; I e J. Atuam sobre a cascata de sinalização - transcriçao gênica de moléculas efetoras;
K. Promovem a desgranulação celular

Peptídeos antimicrobianos de aracnídeos
Laboratório de Imunologia e Bioquímica de Artrópodes Depto Parasitologia – ICB - USP
Rhipicephalus (Boophilus) microplus
Acanthoscurria gomesiana

Moléculas antimicrobianas dos hemócitos da Acanthocurria gomesiana
Migalina (Pereira et al. 2007 BBRC)
DVYKGGGGGRYGGGRYGGGGGYGGGLGGGGLGGGGLGGGKGLGGGGLGG GGLGGGGLGGGGLGGLGGGGLGGGGLGGGGLGGGGLGGGKGLGGGGLGG GGLGGGRGGGYGGGGGYGGGYGGGYGGGKYK
Acantoscurrinas (Lorenzini et al. 2003 Dev Comp Immunol)
Gomesina (Silva et al. 2000 JBC )
ZCRRLCYKQRCVTYCRGR

Peptídeos antimicrobianos - R. (Boophilus) microplus
hemolinfa
intestino
ovos
GFGCPFNQGACHRHCRSIRRRGGYCAGLIKQTCTCYRN
Defensina (Fogaça et al. 2004 Dev Comp Immunol)
HHQELCTKGDDALVTELECIRLRISPETNAAFD NAVQQLNCLNRACAYRKMCATNNLEQAMSV YFTNEQIKEIHDAAT CCDPEAHHEHDH
Microplusina (Fogaça et al. 2004 Dev Comp Immunol Silva et al 2009, JBC)
Ixodidina (Fogaça et al. 2006 Peptides)
ZRGSRGQRCGPGEVFNQCGSACPRVCGRPPAQ ACTLQCVSGCFCRRGYIRTQRGGCIPERQCHQR
FLSFPTTKTYFPHFDLSHGSAQVKGHGAK Hb 33-61 (Fogaça et al 1999 JBC)
FKLLSHSLLVTLASHLP Hb 98-114 (Belmonte 2010 )

1 µm
Esteves et al, 2008
20 µm
Phagocytosis ESTs library : expression of immune genes (AMPs)
Interaction Anaplasma marginale with embryonic cell line from R. B. microplus : BME26

Esteves et al., 2009
10 µm
8 DPI 14 DPI
20 DPI 20 DPI
BME26 infected with Anaplasma marginale
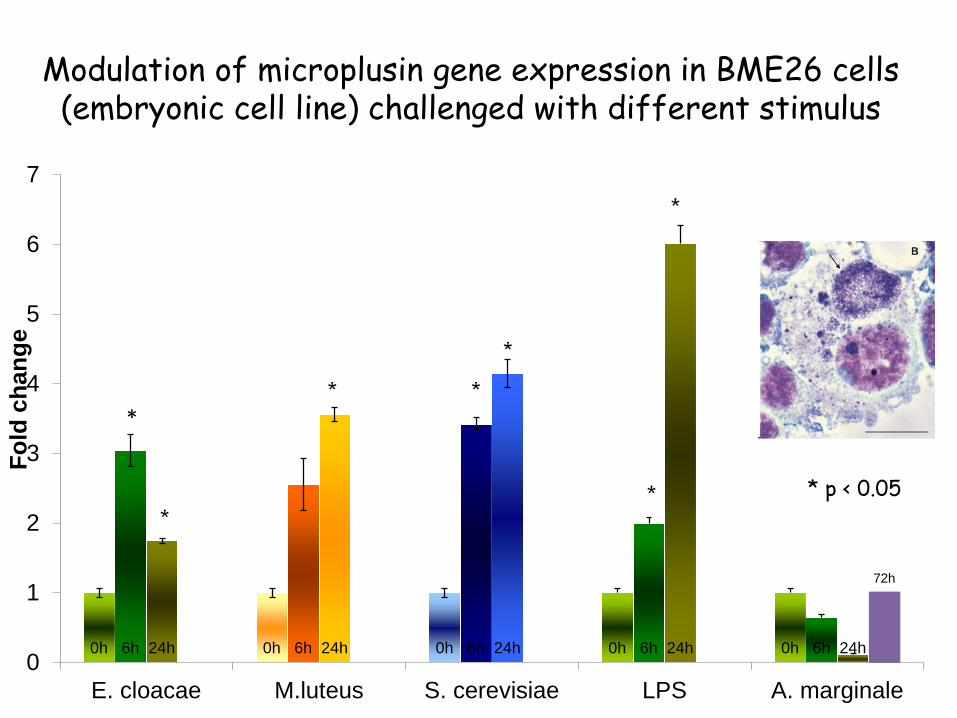
Modulation of microplusin gene expression in BME26 cells (embryonic cell line) challenged with different stimulus
0h 0h 0h 0h 0h 6h 6h 6h 6h 6h 24h 24h 24h 24h 24h
72h
0
1
2
3
4
5
6
7
E. cloacae M.luteus S. cerevisiae LPS A. marginale
Fold
cha
nge
*
* p < 0.05

Microplusin Defensin Ixodidin
A. marginale no change upregulated upregulated
Modulation of AMP gene expression
Different signaling pathway
M. luteus up regulated no change no change
S. cerevisiae upregulated upregulated downregulated
E. cloacae upregulated upregulated upregulated
Does AMPs gene silencing affect the Anaplasma marginale infection?
No difference in bacteria number was detected in silenced microplusin and defensin genes

Janaina Peixoto
Signalling pathways in tick immunity (Toll, IMD and Jak/Stat)
Anaplasma marginale Other microorganisms non-pathogenics


Tick genes modulated in response to A. marginale
infection using BME 26 cells
Transcriptome: Suppression-subtractive
hybridization (SSH) libraries
Proteome: 2D LC-MS/MS

DIFFERENTIAL PROTEOMIC ANALYSIS OF BME26 CELLS INFECTED WITH Anaplasma marginale
Carlos Eduardo Cruz Cláudia Angeli

[NaCl]
Protein extraction at 24h post-infection
Protein quantitation (BCA) / trypsin digestion / desalting
uHPLC
• 0 mM• 25 mM• 50 mM• 100 mM• 200 mM• 500 mM
SCX C18
Infection of culture cells with A .marginale
LC-MS/MS
Solvent B: 80% ACN / 0.1% AF
• 5% - 40% B: 100 min• 40% - 95% B: 1 min• 95% B: 10 min• 95% - 5% B: 1 min

Relative quantification (Qspec)
Databases • Rhipicephalus sp • Anaplasma sp • Bos taurus • Human keratin • Trypsin • R. microplus ESTs (BmiGI database - http://www.tigr.org) translated with OrfPredictor (http://proteomics.ysu.edu/tools/OrfPredictor.html)
Spectral analyses: Sequest
215 Sequences : 81 134

Aminoacid catabolism (4.5%)
Vacuolar ATPase activity (3.0%)
Cell cycle regulation (1.5%)
Nucleic acid metabolism (6.0%)
Transcriptional/post-transcriptional
regulation (6.7%)
Mitochondrial metabolism (3.7%)
Translational/post-translational
regulation/protein folding (25.4%)Vesicular trafficking
(9.7%)
Proteolysis (9.0%)
Redox homeostasis/detox./reductase activity
(9.0%)
Ion transport (4.5%)
Others (17.2%)
Downregulated
Aminoacid metabolism (4.9%)
Aminoacid catabolism (1.2%)
Cell cycle regulation (3.7%)Nucleic acid
metabolism (2.5%)
Transcriptional/post-transcriptional
regulation (8.6%)
Mitochondrial metabolism (16.0%)
Translational/post-translational
regulation/protein folding (21.0%)
Vesicular trafficking (8.6%)
Proteolysis (4.9%)
Redox homeostasis/detox./reductase activity
(6.2%)
ROS generation (2.5%)
Others (19.8%)
Upregulated

Host cell proteins involved in redox homeostasis modulated by infection (24h)
Accession number Sequence name Fold-change
ISCW023288 aldehyde dehydrogenase 2 2.5gi222376748 glutaredoxin 3 2.7gi156025971 glutathione S-transferase 2.6gi82845164 thiol-disulfide isomerase and thioredoxin 3.4TC15651 SH3 domain-binding glutamic acid-rich protein 2.4gi222393859 dual oxidase 3.9gi82848299 ER membrane-associated oxidoreductin 4.5
gi82845200 aldehyde dehydrogenase 4.0TC23737 aldehyde dehydrogenase 2.1gi82843598 peroxiredoxin 2.6gi49571790 glutathione S-transferase 2.6TC18741 glutathione S-transferase 2.8gi82848235 Mn superoxide dismutase 9.5gi49561935 thioredoxin-dependent peroxide reductase 2.8gi67083289 thioredoxin peroxidase 3.1ISCW010532 alkyl hydroperoxide reductase 8.5
Upregulated
Downregulated
Sandra Kalil
Stress oxidative characterization during the Anaplasma marginale’s infection in BME26

Carlos Cruz Pós-doc
Sandra Kalil Doutoranda
Janaína Peixoto Doutoranda
Eliane Esteves Pós-doc
Thaís Bifano Pós doc
Caracaterização global de genes e proteínas
diferencialmente expressas pela BME26 frente infecção
por A. marginale
Efeito da temperatura na transmissão transovariana de A. marginale em fêmeas do
carrapato bovino
Paula Pohl Pós-doc
Cláudia Angeli Técnica especialista
Metabolismo energético
Estresse oxidativo
Sistema Imune
Vias de sinalização
Mário Balanco Técnico especialista
Rafael Rosa Pós-doc

Andréa C. Fogaça – USP Gloria Braz - UFRJ Igor Almeida, Ernesto Nakayasu - UTEP, USA José Pires – UFRJ Pedro L. Oliveira - UFRJ Renato Mortara – UNIFESP Ueti Massaro – WSU, USA
Financial support: FAPESP, CNPq, Capes, INCT-EM
Collaborators