site-directed mutagenesis analysis of the role of the m5s5 sector of the sarcoplasmic reticulum...
TRANSCRIPT

Site-Directed Mutagenesis Analysis of the Role of the M5S5 Sector of the
Sarcoplasmic Reticulum Ca2+-ATPase“ JENS PETER ANDERSEN,b THOMAS SP)RENSEN,
AND BENTE VILSEN Department of Physiology
University ofAarhus DK-8000 Aarhus C. Denmark
Structure-function relationships of the sarcoplasmic reticulum Ca2+-ATF’ase have been studied intensely by site-directed mutagenesis, and it has become feasible to assign specific functional roles to domains and subdomains as well as to individual amino acid residues on the basis of functional characterizations of the mutant enzymes’@ (FIG. 1). The largest cytoplasmic domain contains residues crucial to ATP binding and phosphoryl transfer. Transmembrane segments M4, M5, and M6 seem to constitute a central region (pore) for occlusion and translocation of Ca2+. Residues Glu309, Glu77 1, Asn796, Thr799, and Asp800 found in these transmembrane seg- ments are essential to Ca2+ occ l~s ion .~-~ Glu908 in transmembrane segment M8 is involved in the optimization of Ca2+ binding but is less crucial than the aforemen- tioned residues in M4, M5, and M6, because the Glu908 - Ala mutant can occlude Ca2+ in the presence of CrATPs and displays Ca2+ uptake activity corresponding to a rate as high as 47% that of the wild type when assayed at a Ca2+ concentration of 200 pM2. Stalk segment S4 connecting transmembrane segment M4 with the phosphoryla- tion domain contains several residues (indicated by open squares in FIG. 1 ) that are important to the conformational rearrangement associated with deocclusion and translocation of Ca2+ and transformation of the phosphoenzyme intermediate from an ADP-sensitive to an ADP-insensitive species (E,P - &P tran~ition).~.~.’ Moreover, many residues in M4, M5, and M6, including Ca2+-binding residues Glu309, Glu77 I , and A ~ n 7 9 6 , ~ are part of a long-distance signaling pathway that relays information from the ion-binding domain in the membrane back to the phosphorylation site to trigger the dephosphorylation of the E2P phosphoenzyme intermediate (residues indicated by diamonds in FIG. I).
Replacement of the highly conserved tyrosine Tyr763 at the M5S5 boundary with glycine uncouples ATPase activity from Ca2+ transport in the sense that the micro- somes containing this mutant display high Ca2+-activated ATPase activity without any net accumulation of Ca2+ in the vesicular space.8 This could mean that Ca2+ is never transported even though ATP is hydrolyzed by the mutant ATPase (true uncoupling) or that following translocation into the microsomal vesicles Ca2+ leaks back through an
“This work was supported by the Danish Medical Research Council and the NOVO Nordisk Foundation.
bAddress for correspondence: Department of Physiology, University of Aarhus, Ole Worms All6 160, DK-8000 Aarhus C, Denmark (tel: 45 8942 2814; fax: 45 8612 9065; e-mail: [email protected]).
333

334 ANNALS NEW YORK ACADEMY OF SCIENCES
\...J M 1 2
- -
3 4 5 6 7 8 9 10
0 = Mutants retaining Ca transport with normal affinity.
0 = Reduced E l P --t E2P. 0 = Reduced E2P- E.
0 = Ca-affinity mutants.
= Phosphorylation-negative mutants.
= Uncoupled mutant. 1:; = Mutant K758i with increased E2P - E2. 0 = ATP-affinity mutants.
FIGURE 1. Ten-helix model of the topology of the sarcoplasmic reticulum CaZ+-ATPase with classification of residues that have been analyzed by site-directed mutagenesis. Filled circles correspond to mutations compatible with normal (or close to normal) function of the expressed SERCAI enzyme. More deleterious mutations are highlighted by showing the wild-type residue in single-letter code inside a symbol indicating the functional class to which the mutant belongs. Double labeling of residues indicates either that two partial reaction steps are affected by the same mutation (e.g., open circles with diamonds) or that two different substituents elicit different effects ( e g , triangles pointing upwards with squares). This classification is based on published papers reviewed previously'-' as well as on unpublished results by the present authors and results personally communicated to us by Dr. David H. MacLennan.
ANDERSEN et al.: SARCOPLASMIC RETICULUM Ca*+-ATPase 335
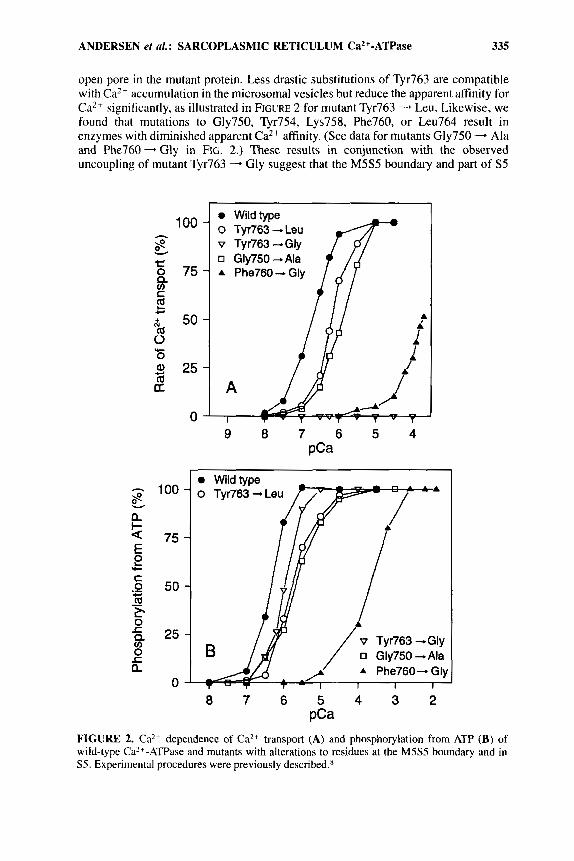
open pore in the mutant protein. Less drastic substitutions of Tyr763 are compatible with CaZt accumulation in the microsomal vesicles but reduce the apparent affinity for Ca2+ significantly, as illustrated in FIGURE 2 for mutant Tyr763 - Leu. Likewise, we found that mutations to Gly750, Tyr754, Lys758, Phe760, or Leu764 result in enzymes with diminished apparent Ca2+ affinity. (See data for mutants Gly750 - Ala and Phe760- Gly in FIG. 2. ) These results in conjunction with the observed uncoupling of mutant Tyr763 - Gly suggest that the M5S5 boundary and part of S5
100 h
8 5 75 E: E Ny 50
u
C
s Y- O
(d +- Q) 25 a
0 9 8 7 6 5 4
PCa
8 7 6 5 4 3 2 PCa
FIGURE 2. Ca2+ dependence of Ca2+ transport (A) and phosphorylation from ATP (B) of wild-type Ca2+-ATPase and mutants with alterations to residues at the M5S5 boundary and in S5. Experimental procedures were previously described.K

336 ANNALS NEW YORK ACADEMY OF SCIENCES
Wildtype + lonophore
Lys758 - Ile
Tyr763 - Gly
Control
I I
0 10 Time (rnin)
FIGURE 3. ATP hydrolysis monitored spectrophotometrically by an NADH-coupled assay8 at 37°C in the presence of 20 mM MOPS (pH 7.0). 100 mM KCl, 2 mM MgCI2, 5 mM MgATP, and 100 pM Ca*+. The bar in the lower lefr corner indicates 0.1 absorbance unit. At the first arrow the microsomes are added to the reaction mixture. At the second arrow the calcium ionophore A23187 is added to relieve back-inhibition by accumulated Ca2+. Finally, EGTA is added to show the background ATPase activity not referable to Ca2+ -ATPase. “Control” indicates microsomes harvested from cells transfected with vector without insert.
are closely associated with the cytoplasmic inlet of the transmembrane transport pathway.
Recently, we found that replacement of Lys7.58 by isoleucine results in a mutant with several unique features which in addition to a reduced apparent Ca2+ affinity include anomalous pH and ATP dependencies of the overall ATPase reaction, as well as a reduced maximum turnover number at neutrai pH and an increased sensitivity to inhibition by an ad ate.^ In the first round of characterization of this mutant, we observed that the ATPase activity measured at a saturating Cazf concentration of 100 pM is slightly inhibited by the addition of Ca2+ ionophore to the sealed right-side-out microsomes (FIG. 3). By contrast, the AWase activity of the wild-type Ca2+-ATPase is enhanced at least twofold by the addition of Ca2+ ionophore due to relief by the ionophore of the “back inhibition” of the rate-limiting EIP - E2P transition imposed by accumulated Ca2+ binding to luminal low-affinity inhibitory sites on E2P. As seen
ANDERSEN et al.: SARCOPLASMIC RETICULUM Ca2+-ATPase 337
100 i
= 75 - 0 R1 .- CI - 6 'p 50 - In 5 a 0
25 -
0
in FIGURE 3, the anomalous ionophore effect observed for mutant Lys758 - Ile resembles that previously reported for the uncoupled mutant Tyr763 - Gly, where the lack of activation by ionophore is caused by the absence of accumulated inhibitory Ca2+ in the vesicular lumen.8 The similar responses of mutants Lys758 - Ile and Tyr763 - Gly to Ca2+ ionophore initially led us to speculate that mutant Lys758 - Ile like mutant Tyr763 - Gly might be uncoupled, but detailed measurements of Ca2+ accumulation over a wide range of Ca2+ concentrations have clearly revealed that mutant Lys758 - Ile is fully coupled. The failure of this mutant to show ionophore- induced activation of the ATPase activity must therefore be ascribed to insensitivity to inhibition by the Ca2+ accumulated in the lumen.
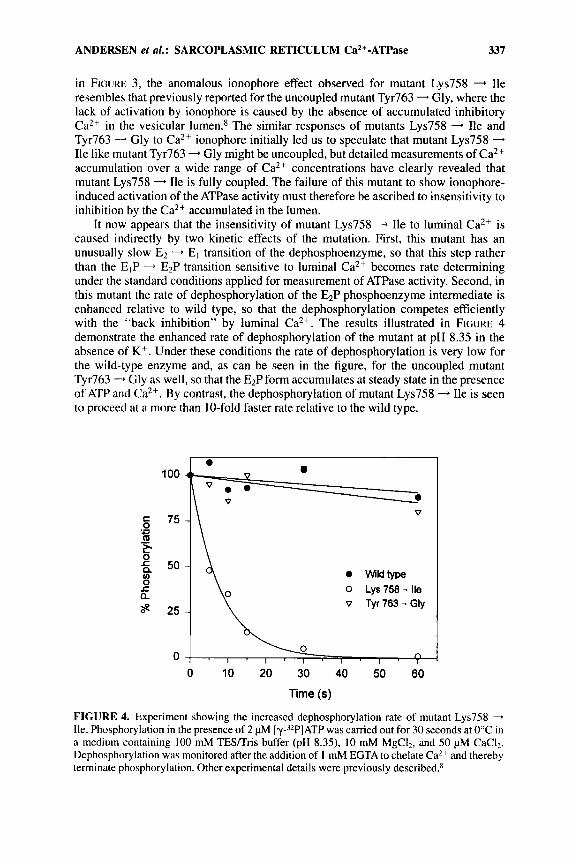
It now appears that the insensitivity of mutant Lys758 - Ile to luminal Ca2+ is caused indirectly by two kinetic effects of the mutation. First, this mutant has an unusually slow E2 - El transition of the dephosphoenzyme, so that this step rather than the EIP - E2P transition sensitive to luminal Ca2+ becomes rate determining under the standard conditions applied for measurement of ATPase activity. Second, in this mutant the rate of dephosphorylation of the E2P phosphoenzyme intermediate is enhanced relative to wild type, so that the dephosphorylation competes efficiently with the "back inhibition" by luminal Ca2+. The results illustrated in FIGURE 4 demonstrate the enhanced rate of dephosphorylation of the mutant at pH 8.35 in the absence of K + . Under these conditions the rate of dephosphorylation is very low for the wild-type enzyme and, as can be seen in the figure, for the uncoupled mutant Tyr763 - Gly as well, so that the E2P form accumulates at steady state in the presence of ATP and Ca2+. By contrast, the dephosphorylation of mutant Lys758 - Ile is seen to proceed at a more than 10-fold faster rate relative to the wild type.
0
-0
V
0 Wildtype 0 Lys758 - Ile v Tyr763- Gly
n l ' l ' l ' l ~ l ~ l
FIGURE 4. Experiment showing the increased dephosphorylation rate of mutant Lys758 - Ile. Phosphorylation in the presence of 2 @I [Y-'~P]ATP was carried out for 30 seconds at 0°C in a medium containing 100 mM TES/Tris buffer (pH 8.35), 10 mM MgCI2, and 50 pM CaCI2. Dephosphorylation was monitored after the addition of I mM EGTA to chelate Ca2+ and thereby terminate phosphorylation. Other experimental details were previously described.R

338 ANNALS NEW YORK ACADEMY OF SCIENCES
In conclusion, the M5S5 sector seems to play an important role in the coupling of ATP hydrolysis to Ca2+ translocation (gate control) and in the optimization of Ca2+ binding as well as in conformational changes involved in dephosphorylation of the E2P phosphoenzyme intermediate. The Lys758 - Ile mutant is the first Ca2+-ATPase mutant for which an increased rate of dephosphorylation of E2P relative to the wild type has been reported.
REFERENCES
1. ANDERSEN, J. P. & B. VILSEN. 1994. In The Sodium Pump: Structure, Mechanism, Hormonal Control and its Role in Disease. E. Bamberg & W. Schoner, Eds.: 98-109. Steinkopff. Darmstadt.
2. ANDERSEN, J. P. 1995. Biosci. Rep. 15: 243-261. 3. VTLSEN, B. 1995. Acta Physiol. Scand. 154 (Suppl. 624): 1-146. 4. VILSEN, B. & J. P. ANDERSEN. 1992. J. Biol. Chem. 267: 25739-25743. 5 . ANDERSEN, J. P. & B. VILSEN. 1994. J. Biol. Chem. 269: 15931-15936. 6. VILSEN, B., J. P. ANDERSEN, D. M. CLARKE & D. H. MACLENNAN. 1989. J. Biol. Chem.
7. VILSEN, B., J. P. ANDERSEN & D. H. MACLENNAN. 1991. J. Biol. Chem. 266: 18839-18845. 8. ANDERSEN, J. P. 1995. J. Biol. Chem. 270: 908-914. 9. S0RENSEN, T., B. VILSEN & J. P. ANDERSEN. Manuscript in preparation.
264: 21024-21030.