sna3 is an rsp5 adaptor protein that relies on ubiquitination for its mvb sorting
TRANSCRIPT

Traffic 2012; 13: 586–598 © 2011 John Wiley & Sons A/S
doi:10.1111/j.1600-0854.2011.01326.x
Sna3 Is an Rsp5 Adaptor Protein that Relieson Ubiquitination for Its MVB Sorting
Chris MacDonald1, Daniel K. Stringer1,2 and
Robert C. Piper1,∗
1Molecular Physiology and Biophysics, University ofIowa, Iowa City, IA 52246, USA2Present address: Laboratory of Protein Dynamics andSignaling, Center for Cancer Research, National CancerInstitute, Frederick, MD 21702, USA*Corresponding author: Robert C. Piper,[email protected]
The process in which ubiquitin (Ub) conjugation is
required for trafficking of integral membrane proteins
into multivesicular bodies (MVBs) and eventual degra-
dation in the lumen of lysosomes/vacuoles is well
defined. However, Ub-independent pathways into MVBs
are less understood. To better understand this process,
we have further characterized the membrane protein
Sna3, the prototypical Ub-independent cargo protein
sorted through the MVB pathway in yeast. We show
that Sna3 trafficking to the vacuole is critically depen-
dent on Rsp5 ligase activity and ubiquitination. We find
Sna3 undergoes Ub-dependent MVB sorting by either
becoming ubiquitinated itself or associating with other
ubiquitinated membrane protein substrates. In addition,
our functional studies support a role for Sna3 as an
adaptor protein that recruits Rsp5 to cargo such as the
methionine transporter Mup1, resulting in efficient Mup1
delivery to the vacuole.
Key words: endosome, lysosome, multivesicular body,
ubiquitin, vacuole
Received 9 September 2011, revised and accepted for
publication 27 December 2011, uncorrected manuscript
published online 29 December 2011, published online 31
January 2012
The levels of cell surface membrane proteins aredetermined in large part by their trafficking to, anddegradation in, lysosomes. Integral membrane proteinsdestined for degradation are modified by the attachmentof ubiquitin (Ub), which serves as a signal for internalizationand subsequent entry into intralumenal vesicles (ILVs) thataccumulate within late endosomes/multivesicular bodies(MVBs). The actions of Ub ligases and deubiquitinatingenzymes that modify proteins by adding or removingUb, respectively, dictate the fate of proteins as theypass through the endosomal system (1,2). Ubiquitinatedcargo proteins that maintain a Ub signal are transportedto the late endosome where the sequential actionof the Endosomal Sorting Complexes Required forTransport (ESCRT) machinery incorporate ubiquitinatedcargo proteins into MVB ILVs (3). Many of the ESCRT
subunits contain Ub-binding domains, thus forming asystem designed to process and sort ubiquitinated cargo(4). MVBs eventually fuse with lysosomes (or in yeast, thevacuole), thereby delivering the ILV-encapsulated cargofor degradation in the lysosomal lumen.
The major ligase responsible for ubiquitination of integralmembrane proteins at the plasma membrane in yeast isRsp5 (5), which is homologous to mammalian Nedd4(6). The tryptophan–tryptophan (WW) domains withinRsp5 and its homologs have been shown to directlyinteract with a PPXY (PY) motif (7,8) found within someMVB cargo proteins (9,10). However, many membraneproteins lack a PY motif and rely on interaction withadaptor proteins to recruit Rsp5 and facilitate theirubiquitination (11–14), creating a regulatory mechanismfor Rsp5 specificity. Many of these Rsp5 adaptors arethemselves ubiquitinated, and integral membrane Rsp5adaptors, such as Bsd2, also become sorted into the MVBpathway (15).
One cargo protein of particular interest to traffickingstudies is Sna3, a small membrane protein that is sortedefficiently into the MVB pathway and accumulates in thevacuole lumen (16). Many studies have proposed thatSna3 does not use a Ub-dependent sorting mechanism(16–18,20,21). Studies have also revealed that Sna3contains a PY motif that mediates interaction with theWW domains of Rsp5, and mutational studies disruptingthis interaction have shown it to be essential for Sna3sorting to the vacuole (17–19). However, mutationof the cytosolic lysine residues within Sna3 does notperturb its vacuolar sorting, suggesting Sna3 follows aUb-independent route to the vacuole (16). In addition,cells carrying particular mutant alleles of RSP5 that haveattenuated Rsp5 Ub-ligase activity appear to sort Sna3normally through the MVB pathway to the vacuole lumen(17–21). These data support a model whereby Sna3accesses a Ub-independent MVB sorting route via phys-ical association with Rsp5, which in turn associates withthe ESCRT apparatus (17,18,20,21). This model, however,has not been fully verified because other experimentsindicate that Sna3 may require ubiquitination, at least tosome extent, for efficient MVB sorting (18,19).
Parallels between a Ub-independent MVB sortingmechanism for Sna3 and cargoes in other systemshave been drawn, because mutation of the cytoplasmiclysine residues within several mammalian cargo proteinsis not sufficient to fully disrupt their lysosomal entry.These include: the GPCR murine δ opioid receptor, thetransferrin receptor, TfR2, and the CD4 receptor (22–24).Where tested, each lysineless mutant cargo follows anESCRT-dependent route into MVBs (22–25). However,
586 www.traffic.dk
Sna3 Is an Rsp5 Adaptor Protein
the exact nature of an ESCRT-dependent MVB sortingroute that does not use Ub as a sorting signal has notbeen clarified yet.
In this study, we investigate the role ubiquitination playsin Sna3 trafficking and address unresolved questionsregarding this process. We find ubiquitination does controlthe sorting of Sna3 into the MVB pathway. We find thatSna3 behaves as an Rsp5 adaptor protein that contributesto the downregulation of the methionine transporterMup1. In addition, our results indicate that sorting of Sna3into the MVB pathway relies on ubiquitination at somelevel – whether of Sna3 itself or of substrate proteinswith which Sna3 associates.
Results
Sna3 without lysine residues is sorted into the MVB
pathway
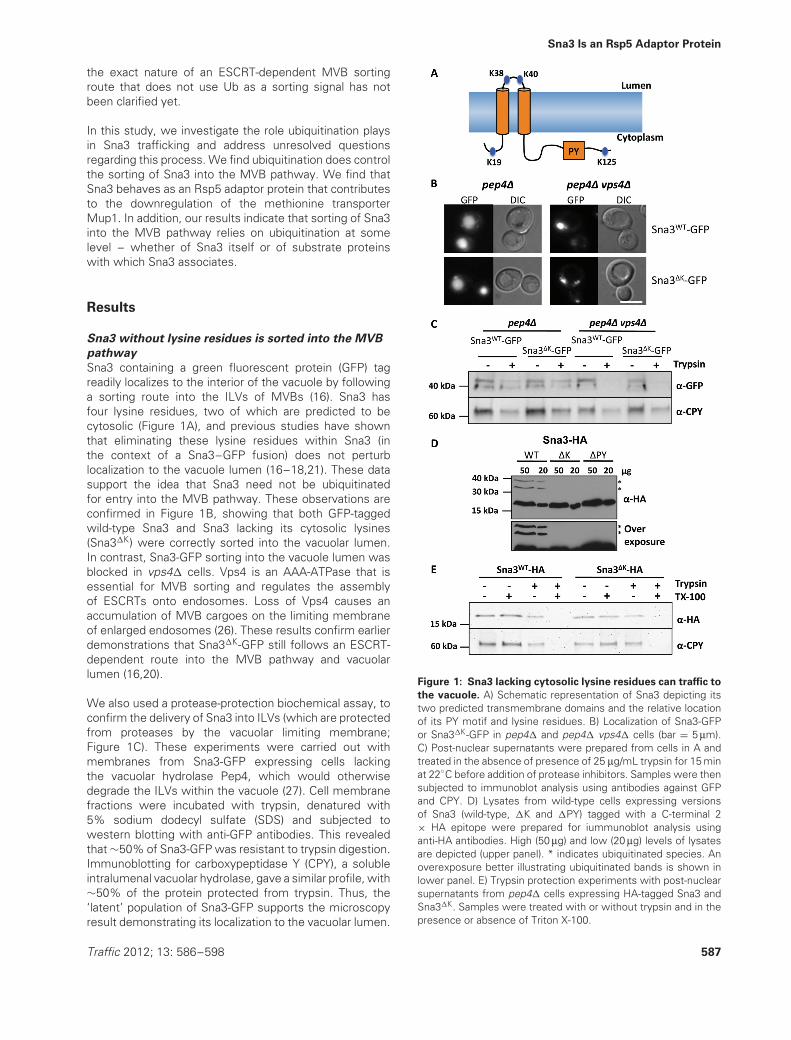
Sna3 containing a green fluorescent protein (GFP) tagreadily localizes to the interior of the vacuole by followinga sorting route into the ILVs of MVBs (16). Sna3 hasfour lysine residues, two of which are predicted to becytosolic (Figure 1A), and previous studies have shownthat eliminating these lysine residues within Sna3 (inthe context of a Sna3–GFP fusion) does not perturblocalization to the vacuole lumen (16–18,21). These datasupport the idea that Sna3 need not be ubiquitinatedfor entry into the MVB pathway. These observations areconfirmed in Figure 1B, showing that both GFP-taggedwild-type Sna3 and Sna3 lacking its cytosolic lysines(Sna3�K) were correctly sorted into the vacuolar lumen.In contrast, Sna3-GFP sorting into the vacuole lumen wasblocked in vps4� cells. Vps4 is an AAA-ATPase that isessential for MVB sorting and regulates the assemblyof ESCRTs onto endosomes. Loss of Vps4 causes anaccumulation of MVB cargoes on the limiting membraneof enlarged endosomes (26). These results confirm earlierdemonstrations that Sna3�K-GFP still follows an ESCRT-dependent route into the MVB pathway and vacuolarlumen (16,20).
We also used a protease-protection biochemical assay, toconfirm the delivery of Sna3 into ILVs (which are protectedfrom proteases by the vacuolar limiting membrane;Figure 1C). These experiments were carried out withmembranes from Sna3-GFP expressing cells lackingthe vacuolar hydrolase Pep4, which would otherwisedegrade the ILVs within the vacuole (27). Cell membranefractions were incubated with trypsin, denatured with5% sodium dodecyl sulfate (SDS) and subjected towestern blotting with anti-GFP antibodies. This revealedthat ∼50% of Sna3-GFP was resistant to trypsin digestion.Immunoblotting for carboxypeptidase Y (CPY), a solubleintralumenal vacuolar hydrolase, gave a similar profile, with∼50% of the protein protected from trypsin. Thus, the‘latent’ population of Sna3-GFP supports the microscopyresult demonstrating its localization to the vacuolar lumen.
Figure 1: Sna3 lacking cytosolic lysine residues can traffic to
the vacuole. A) Schematic representation of Sna3 depicting itstwo predicted transmembrane domains and the relative locationof its PY motif and lysine residues. B) Localization of Sna3-GFPor Sna3�K-GFP in pep4� and pep4� vps4� cells (bar = 5 μm).C) Post-nuclear supernatants were prepared from cells in A andtreated in the absence of presence of 25 μg/mL trypsin for 15 minat 22◦C before addition of protease inhibitors. Samples were thensubjected to immunoblot analysis using antibodies against GFPand CPY. D) Lysates from wild-type cells expressing versionsof Sna3 (wild-type, �K and �PY) tagged with a C-terminal 2× HA epitope were prepared for iummunoblot analysis usinganti-HA antibodies. High (50 μg) and low (20 μg) levels of lysatesare depicted (upper panel). * indicates ubiquitinated species. Anoverexposure better illustrating ubiquitinated bands is shown inlower panel. E) Trypsin protection experiments with post-nuclearsupernatants from pep4� cells expressing HA-tagged Sna3 andSna3�K. Samples were treated with or without trypsin and in thepresence or absence of Triton X-100.
Traffic 2012; 13: 586–598 587

MacDonald et al.
The sensitivity of a portion of Sna3-GFP to trypsin digestionis likely due to rupture of some of the vacuoles duringthe isolation of cell membranes because CPY, a well-established marker for the vacuolar lumen, had the samedigestion profile. Importantly, all of the Sna3-GFP withinmembranes from vps4� mutant cells were sensitive totrypsin treatment, consistent with the accumulation ofSna3-GFP on the limiting membranes of endosomes andvacuoles.
The Sna3�K-GFP protein sorts normally to the vacuolelumen; however, the fused GFP retains surface-exposedlysines (15 in the GFP variant used in this study). Althougheliminating the lysines within Sna3 dramatically reducesubiquitination of Sna3-GFP, the GFP moiety has thecapacity to become ubiquitinated and it is conceivablethat this low level could be driving MVB sorting (19). Tocircumvent this problem, we fused two HA epitopes (YPY-DVPDYA, which do not contain ubiquitinatable lysines)to the C-terminus of Sna3 and Sna3�K. Immunoblotanalysis confirmed that the high-molecular-weight bands,corresponding to ubiquitinated forms of Sna3, were notreadily formed by Sna3�K (Figure 1D). We next assessedthe distribution of Sna3-HA and Sna3�K-HA by trypsinprotection and found that both were protected equallywell (Figure 1E), indicating that lysine ubiquitination is notrequired for entry of Sna3 into the MVB pathway. As afurther control, we performed the protease treatment inthe presence of detergent, thereby disrupting the lipidmembranes and allowing the trypsin to access proteinsin the vacuole. Addition of Triton X-100 had no effecton overall Sna3 levels but led to the complete digestionof both Sna3 and CPY when trypsin is subsequentlyadded. These findings support the model that Sna3 doesnot require its lysines to undergo ubiquitination for MVBsorting (17,18,21). However, they conflict with previousobservations that used immunofluorescence techniquesto demonstrate that HA-tagged Sna3�K is not localizedto the vacuolar lumen (19). One potential explanationfor this discrepancy is that under the conditions used inthe previous studies, the intravacuolar Sna3-HA may nothave been accessible for indirect immunofluorescencedetection; in yeast, this procedure typically requires harshfixation and extraction methods (28).
Sna3 sorting through the MVB pathway is dependent
on both Rsp5 activity and ubiquitination
Although the above studies clearly indicate that ubiquiti-nation of lysine residues within Sna3 are not required forcorrect MVB sorting, there are several caveats to con-cluding that Sna3 strictly uses a different signal than Ubfor its sorting. For instance, proteins can undergo ubiq-uitination of cysteine, serine or threonine residues undersome conditions (29). In addition, proteins can be ubiqui-tinated on their N-terminus (30), and the Sna3-associatedUb-ligase Rsp5, in particular, has been shown to becapable of ubiquitinating the N-terminus of Sna3 at leastin vitro (31).
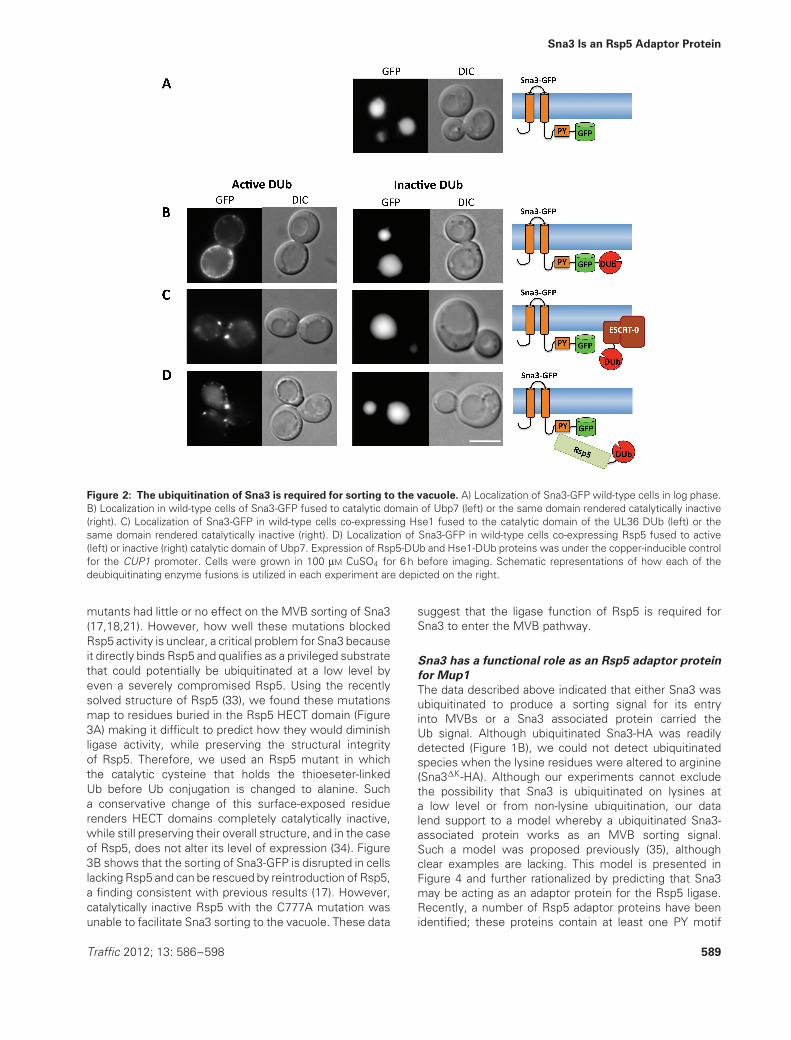
To test whether Sna3 sorting is completely independentof Ub, we used a recently described technique wherebythe catalytic domain of a deubiquitinating enzyme (DUb)can be covalently attached to a protein of interest to makeit resistant to ubiquitination (32). In addition, when theDUb domain is attached to the ESCRT machinery, it canblock the entry into the MVB of cargo that requires cellularubiquitination but not cargo that is fused to Ub so that theC-terminal specific DUb peptidase domain cannot removeit. Finally, when fused to the Ub-ligase Rsp5, the DUbdomain dominantly inhibits Rsp5-mediated ubiquitinationand cargo trafficking by acting as an antagonistic Rsp5substrate-specific DUb. Figure 2 shows that all of theseDUb fusions blocked Sna3-GFP MVB sorting, suggestingthat Sna3 does rely on Ub as a sorting determinant.Sna3-GFP fused to the catalytic domain of yeast Ubp7(Sna3-GFP-Ubp7), failed to localize to the vacuole and wasinstead found largely at the plasma membrane. This effectwas alleviated when the catalytic cysteine residue withinthe Ubp7 DUb domain was mutated to serine. Thus, theblock imposed by the Ubp7 catalytic domain on Sna3-GFPsorting is a consequence of deubiquitination rather thanof steric hindrance or the ability of the Ubp7 domain tointeract with other yeast proteins.
MVB sorting of Sna3-GFP was also dramatically blockedin cells expressing Hse1-DUb indicating that Hse1-DUb iscapable of removing Ub from the Sna3 cargo, effectivelyblocking its MVB sorting. The fact that Hse1 fused tocatalytically inactive DUb had no effect on Sna3 sortingconfirmed that the dominant negative effect of Hse1-DUb on Sna3 sorting is due to deubiquitinating activity,presumably directed toward Sna3 or a Sna3 complex ofproteins. Finally, expressing a dominant-negative Rsp5-DUb also disrupted the sorting of Sna3-GFP. In contrast,an Rsp5-DUb, wherein the DUb domain is catalyticallyinactive, did not perturb Sna3-GFP sorting into the vacuole.Previous studies have shown that blocks in the sortingof ubiquitinated cargo imposed by DUb fusions withboth ESCRT subunits or Rsp5 can be relieved usingcargo translationally fused to ubiquitin, thus demonstratingthat the MVB sorting pathway itself remains operationalunder these conditions (32). If Sna3 were to use a Ub-independent sorting signal, then these proteins shouldnot have blocked its transport to the MVB pathway. Takentogether, our data strongly suggest that Ub serves as asorting signal for the entry of Sna3 into the MVB pathwayand for its accumulation within the vacuole lumen.
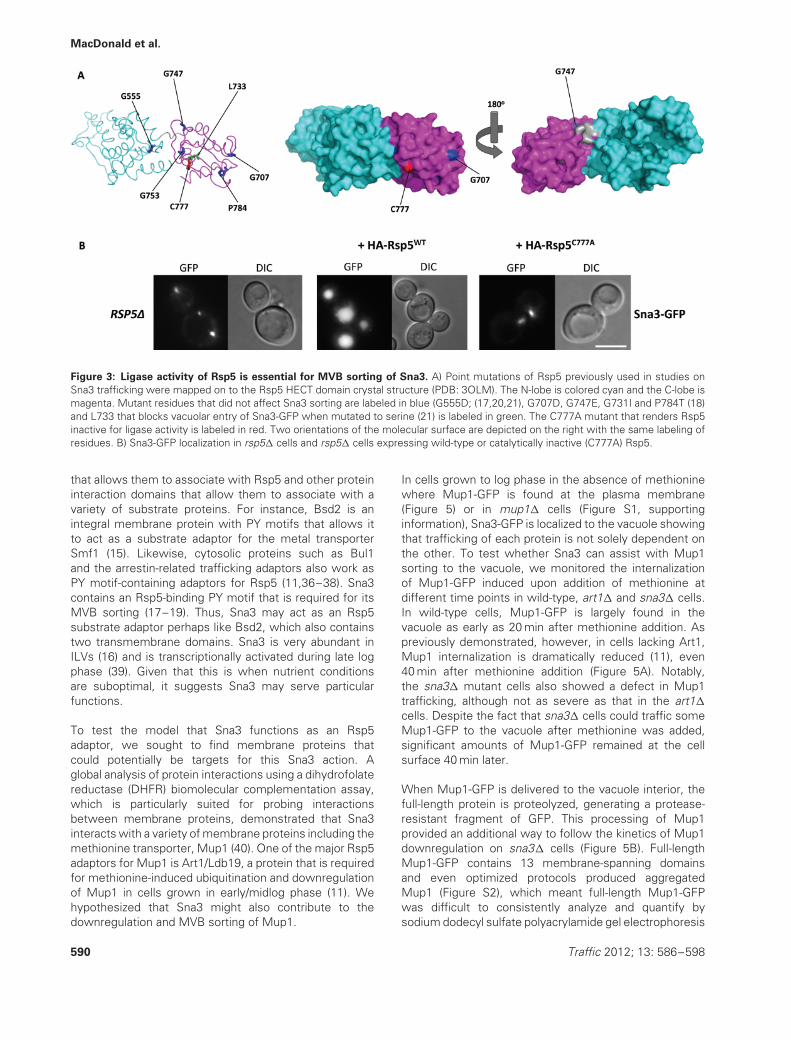
The finding that Sna3 sorting is disrupted by a fusionof Rsp5 with a DUb supports previous studies thatdemonstrate that the interaction between Rsp5 and Sna3facilitates the ubiquitination of Sna3 (17–19,21). However,it is unclear whether the catalytic activity of Rsp5 isrequired or the association with Rsp5 is sufficient forcorrect Sna3 sorting (17,18,21). Sna3 sorting has beenmonitored in the context of various rsp5 mutant allelesthat have attenuated ligase activity due to point mutationsin the HECT domain (Figure 3A). Most of these Rsp5
588 Traffic 2012; 13: 586–598
Sna3 Is an Rsp5 Adaptor Protein
Figure 2: The ubiquitination of Sna3 is required for sorting to the vacuole. A) Localization of Sna3-GFP wild-type cells in log phase.B) Localization in wild-type cells of Sna3-GFP fused to catalytic domain of Ubp7 (left) or the same domain rendered catalytically inactive(right). C) Localization of Sna3-GFP in wild-type cells co-expressing Hse1 fused to the catalytic domain of the UL36 DUb (left) or thesame domain rendered catalytically inactive (right). D) Localization of Sna3-GFP in wild-type cells co-expressing Rsp5 fused to active(left) or inactive (right) catalytic domain of Ubp7. Expression of Rsp5-DUb and Hse1-DUb proteins was under the copper-inducible controlfor the CUP1 promoter. Cells were grown in 100 μM CuSO4 for 6 h before imaging. Schematic representations of how each of thedeubiquitinating enzyme fusions is utilized in each experiment are depicted on the right.
mutants had little or no effect on the MVB sorting of Sna3(17,18,21). However, how well these mutations blockedRsp5 activity is unclear, a critical problem for Sna3 becauseit directly binds Rsp5 and qualifies as a privileged substratethat could potentially be ubiquitinated at a low level byeven a severely compromised Rsp5. Using the recentlysolved structure of Rsp5 (33), we found these mutationsmap to residues buried in the Rsp5 HECT domain (Figure3A) making it difficult to predict how they would diminishligase activity, while preserving the structural integrityof Rsp5. Therefore, we used an Rsp5 mutant in whichthe catalytic cysteine that holds the thioeseter-linkedUb before Ub conjugation is changed to alanine. Sucha conservative change of this surface-exposed residuerenders HECT domains completely catalytically inactive,while still preserving their overall structure, and in the caseof Rsp5, does not alter its level of expression (34). Figure3B shows that the sorting of Sna3-GFP is disrupted in cellslacking Rsp5 and can be rescued by reintroduction of Rsp5,a finding consistent with previous results (17). However,catalytically inactive Rsp5 with the C777A mutation wasunable to facilitate Sna3 sorting to the vacuole. These data
suggest that the ligase function of Rsp5 is required forSna3 to enter the MVB pathway.
Sna3 has a functional role as an Rsp5 adaptor protein
for Mup1
The data described above indicated that either Sna3 wasubiquitinated to produce a sorting signal for its entryinto MVBs or a Sna3 associated protein carried theUb signal. Although ubiquitinated Sna3-HA was readilydetected (Figure 1B), we could not detect ubiquitinatedspecies when the lysine residues were altered to arginine(Sna3�K-HA). Although our experiments cannot excludethe possibility that Sna3 is ubiquitinated on lysines ata low level or from non-lysine ubiquitination, our datalend support to a model whereby a ubiquitinated Sna3-associated protein works as an MVB sorting signal.Such a model was proposed previously (35), althoughclear examples are lacking. This model is presented inFigure 4 and further rationalized by predicting that Sna3may be acting as an adaptor protein for the Rsp5 ligase.Recently, a number of Rsp5 adaptor proteins have beenidentified; these proteins contain at least one PY motif
Traffic 2012; 13: 586–598 589
MacDonald et al.
Figure 3: Ligase activity of Rsp5 is essential for MVB sorting of Sna3. A) Point mutations of Rsp5 previously used in studies onSna3 trafficking were mapped on to the Rsp5 HECT domain crystal structure (PDB: 3OLM). The N-lobe is colored cyan and the C-lobe ismagenta. Mutant residues that did not affect Sna3 sorting are labeled in blue (G555D; (17,20,21), G707D, G747E, G731I and P784T (18)and L733 that blocks vacuolar entry of Sna3-GFP when mutated to serine (21) is labeled in green. The C777A mutant that renders Rsp5inactive for ligase activity is labeled in red. Two orientations of the molecular surface are depicted on the right with the same labeling ofresidues. B) Sna3-GFP localization in rsp5� cells and rsp5� cells expressing wild-type or catalytically inactive (C777A) Rsp5.
that allows them to associate with Rsp5 and other proteininteraction domains that allow them to associate with avariety of substrate proteins. For instance, Bsd2 is anintegral membrane protein with PY motifs that allows itto act as a substrate adaptor for the metal transporterSmf1 (15). Likewise, cytosolic proteins such as Bul1and the arrestin-related trafficking adaptors also work asPY motif-containing adaptors for Rsp5 (11,36–38). Sna3contains an Rsp5-binding PY motif that is required for itsMVB sorting (17–19). Thus, Sna3 may act as an Rsp5substrate adaptor perhaps like Bsd2, which also containstwo transmembrane domains. Sna3 is very abundant inILVs (16) and is transcriptionally activated during late logphase (39). Given that this is when nutrient conditionsare suboptimal, it suggests Sna3 may serve particularfunctions.
To test the model that Sna3 functions as an Rsp5adaptor, we sought to find membrane proteins thatcould potentially be targets for this Sna3 action. Aglobal analysis of protein interactions using a dihydrofolatereductase (DHFR) biomolecular complementation assay,which is particularly suited for probing interactionsbetween membrane proteins, demonstrated that Sna3interacts with a variety of membrane proteins including themethionine transporter, Mup1 (40). One of the major Rsp5adaptors for Mup1 is Art1/Ldb19, a protein that is requiredfor methionine-induced ubiquitination and downregulationof Mup1 in cells grown in early/midlog phase (11). Wehypothesized that Sna3 might also contribute to thedownregulation and MVB sorting of Mup1.
In cells grown to log phase in the absence of methioninewhere Mup1-GFP is found at the plasma membrane(Figure 5) or in mup1� cells (Figure S1, supportinginformation), Sna3-GFP is localized to the vacuole showingthat trafficking of each protein is not solely dependent onthe other. To test whether Sna3 can assist with Mup1sorting to the vacuole, we monitored the internalizationof Mup1-GFP induced upon addition of methionine atdifferent time points in wild-type, art1� and sna3� cells.In wild-type cells, Mup1-GFP is largely found in thevacuole as early as 20 min after methionine addition. Aspreviously demonstrated, however, in cells lacking Art1,Mup1 internalization is dramatically reduced (11), even40 min after methionine addition (Figure 5A). Notably,the sna3� mutant cells also showed a defect in Mup1trafficking, although not as severe as that in the art1�
cells. Despite the fact that sna3� cells could traffic someMup1-GFP to the vacuole after methionine was added,significant amounts of Mup1-GFP remained at the cellsurface 40 min later.
When Mup1-GFP is delivered to the vacuole interior, thefull-length protein is proteolyzed, generating a protease-resistant fragment of GFP. This processing of Mup1provided an additional way to follow the kinetics of Mup1downregulation on sna3� cells (Figure 5B). Full-lengthMup1-GFP contains 13 membrane-spanning domainsand even optimized protocols produced aggregatedMup1 (Figure S2), which meant full-length Mup1-GFPwas difficult to consistently analyze and quantify bysodium dodecyl sulfate polyacrylamide gel electrophoresis
590 Traffic 2012; 13: 586–598

Sna3 Is an Rsp5 Adaptor Protein
Figure 4: Model for Sna3 function and sorting. A) Rsp5 is recruited to Mup1 in response to high levels of methionine throughinteraction with the Art1 adaptor protein. This association with Rsp5 results in the ubiquitination of Art1. B) Mup1 is also internalizedin response to nutrient stress, where Rsp5 can be recruited to Mup1 via Sna3, which can act as a complementary adaptor protein.There is partial redundancy between Art1 and Sna3, each of which contributes to MVB sorting. Sna3 can also be ubiquitinated by Rsp5.C) A lysineless version of Sna3 (cytosolic lysine to arginine mutations), which can still associate with both Rsp5 and Mup1, fostersubiquitination of Mup1 and uses Mup1-Ub to enter the MVB pathway.
(SDS-PAGE). However, extraction and detection ofthe vacuole-processed soluble GFP (vpGFP) allowed ameasure of how much Mup1-GFP was being targetedto the vacuole. In wild-type cells, methionine additionresulted in a dramatic increase in Mup1-GFP derivedvpGFP, whereas, no increase was observed in art1�
cells. The sna3� cells were also defective, producingvpGFP less rapidly than wild-type, an effect mostobvious from comparison at the 20-min time point.These data demonstrate that Sna3 can contribute to thedownregulation and MVB sorting of Mup1.
Delivery of plasma membrane proteins to the vac-uole/lysosome can be induced by various mechanisms,such as ligand binding to pheromone receptors (41),substrate interaction with transporters (11,42), nutrientstarvation (43) and toxic stress (44). In an effort to induceMup1 downregulation through a different mechanism, westressed cells by growing them just past a late log phase,at which nutrients would be limiting. Microarray exper-iments have shown that these growth conditions alsoinduce expression of SNA3 (39). In the absence of methio-nine, Mup1 was localized to the vacuole under theseconditions in wild-type cells (Figure 6A). Interestingly,Mup1-GFP was also localized to the vacuole in art1� cells,demonstrating that Art1 is not required for all modes ofMup1 downregulation. In contrast, Mup1-GFP targeting to
the vacuole was defective in cells lacking Sna3. The sortingdefect in sna3� cells was corrected by reintroduction ofplasmid expressed Sna3-HA or Sna3�K-HA without lysineresidues. However, sorting was not restored by Sna3�PY-HA, lacking the ability of binding Rsp5 (Figure 6B). Thesefindings support the hypothesis that Sna3 not only acts asan Rsp5 adaptor protein for Mup1 but also that its role isdistinct from that of Art1.
As an additional test, we exploited our observation thata Sna3-DUb fusion protein no longer localizes to thevacuole (Figure 2). We reasoned that if Sna3 is a bona fideRsp5 Ub-ligase adaptor protein for Mup1, the expressionof Sna3-DUb should disrupt the Ub-dependent sortingof target proteins such as Mup1 into the MVB pathway.We co-expressed either Sna3-mCherry or Sna3-mCherry-Ubp7 with Mup1-GFP, and induced downregulationof cell surface Mup1-GFP with a 40-min treatment ofmethionine. As expected, Sna3-mCherry and Mup1-GFPwere found exclusively in the vacuolar interior. However,expressing Sna3-mCherry-Ubp7 dramatically reduced theamount of Mup1-GFP delivered into the vacuole (Figure7A). Mup1 labeling at the plasma membrane after methio-nine addition was more pronounced in cells expressingSna3-GFP-Ubp7 than in sna3� cells (Figure 5), presumablybecause the DUb fusion dominantly acts to deubiquitinateMup1 regardless of whether Sna3 or another mechanism,
Traffic 2012; 13: 586–598 591

MacDonald et al.
Figure 5: Sna3 is an adaptor protein for the methionine transporter Mup1. A) Wild-type, sna3� and art1� cells transformed with aplasmid expressing Mup1-GFP were grown to an OD600 = 1.0 in minimal media. Mup1-GFP internalization was then induced by additionof 20 μg/mL methionine. B) Cells were harvested at given time points after methionine addition, lysed and subjected to SDS-PAGE andimmunoblot analysis using anti-GFP (upper panels) and anti-PGK (lower panels) antibodies.
such as Art1 recruitment of Rsp5, was responsible forits ubiquitination. We found that in addition to methionineinducing downregulation of Mup1, Sna3-mCherry-Ubp7also blocked MVB sorting of Mup1-GFP in response tohigh cell density (Figure S3). Collectively, these datasupport a role for Sna3 as an Rsp5 adaptor for Mup1.
Sna3 can be sorted into the MVB pathway by its
substrate
According to our model (Figure 4), Sna3 associates withsubstrates such as Mup1 and uses its PY motif tobind Rsp5, which in turn ubiquitinates either Sna3 orMup1 or both proteins. Previous studies have shownthat loss of the PY motif within Sna3 (Sna3�PY) blocksthe ubiquitination and MVB sorting of Sna3 and insteadcauses it to localize to puncta (2–3 larger puncta as wellas numerous smaller puncta (17–19). We also found thatSna3�PY-GFP did not localize to the vacuole interior (Figure7B) and that ubiquitination of Sna3�PY-HA is blocked(Figure 1D). Thus, failure of Sna3�PY to sort to the MVB
is explained in our model (Figure 4) as a consequenceof loss of Ub from either Sna3 or its substrate protein.This model predicts that restoring ubiquitination of thesubstrate will also restore MVB sorting of Sna3�PY. To testthis, we expressed Mup1-mCherry-Ub under control of theCUP1 promoter, with the addition of copper allowing highlevels of protein expression. In addition to the in-framefusion of Ub, Mup1 internalization was further favored byadding methionine to the growth media. Mup1-mCherry-Ub was exclusively found inside the vacuole. Moreover,expression of Mup1-mCherry-Ub was sufficient to alsoinduce MVB sorting of Sna3�PY-GFP (Figure 7B). Thesedata support the idea of a ‘piggyback’ mechanism forsorting of Sna3 and its possible substrates.
Discussion
Previous studies provided evidence both for and againstthe hypothesis that Sna3 requires lysine ubiquitination
592 Traffic 2012; 13: 586–598

Sna3 Is an Rsp5 Adaptor Protein
Figure 6: Mup1 internalization is induced at high cell density. Wild-type cells expressing Mup1-GFP were grown in minimal mediaand GFP localization was assessed at OD600 = 1.0 (upper panels) and then at OD600 = 2.0 (lower panels).
for its MVB sorting (16–19,21). To date, this discrepancycould be explained by constructs expressing Sna3 taggedwith lysine-containing GFP that could act as acceptors forubiquitination (17–19). By using a biochemical approachwith versions of Sna3 tagged either with or without lysineresidues, we have confirmed that the lysine residueswithin Sna3 are not essential for its correct sorting throughthe MVB pathway (Figure 1).
However, in contrast to previous studies that findubiquitination superfluous for Sna3 MVB sorting, our datasupport the model that Ub does serve a critical role in Sna3trafficking. This was most effectively shown by followingSna3 that was tagged with a DUb catalytic domain thatconfers the ability to deubiquitinate proteins to whichit is attached (32). Additionally, expressing the ESCRT-0subunit Hse1 or the Ub ligase Rsp5 as a fusion with aDUb enzyme also disrupted Sna3 trafficking, implicatingUb as the sorting signal for ESCRT mediated sorting ofSna3 into the vacuole (Figure 2). Previous data in supportof a Ub-independent sorting mechanism for Sna3 alsoincluded the observation that sorting was not perturbedin doa4� cells, which have reduced levels of Ub andubiquitination (16). However, a more in-depth investigationusing different strains and growing cells to different opticaldensities found that Sna3-GFP could not localize to thevacuole in some doa4� cells (19), supporting the view thatubiquitination was somehow important. Another indicationthat ubiquitination might play a role in this process wasa kinetic delay observed in vacuolar entry of Sna3 lackinglysine residues compared withwild-type (18).
A Ub-independent MVB sorting mechanism was alsosupported by the observation that mutations thatcompromised Rsp5 activity did not block Sna3 sorting
(17–20). However, Sna3 binds directly to Rsp5, probablygiving Sna3 a privileged status as a substrate that mightbe effective even when Rsp5 activity is compromised.Thus, even the use of Rsp5 mutants with residual ligaseactivity might be enough to support Sna3 sorting. Theonly previously used rsp5 HECT domain mutant that didnot support Sna3 sorting was L733A (21), which eithercould have less ligase activity or might instead havecompromised structure. We find that direct inactivationof Rsp5 via mutation of its catalytic cysteine, while stillpreserving the overall structure of Rsp5, blocks Sna3sorting and phenocopies the rsp5� phenotype. Thisresult correlates with our observation that Rsp5 fused toa DUb causes missorting of Sna3 (Figure 2). Collectively,these findings are not consistent with the view that directprotein–protein interactions via Rsp5 are what drivean apparent Ub-independent sorting route into MVBs.In addition, these results agree with previous findingsshowing that proper MVB sorting of Sna3�K-GFP in cellswith reduced Rsp5 activity is severely perturbed (18).
Our data (Figure 7) also agree with previous observationsthat mutation of the Rsp5-binding PY motif within Sna3ablates vacuolar localization (17–19). Yet, we were ableto force Sna3�PY-GFP into the lumen of the vacuole byoverexpressing one of its proposed substrate partners,Mup1, fused to Ub. This finding demonstrates thatassociation with ubiquitinated proteins is sufficient forSna3�PY to be processed by the ESCRT machinery andincorporated into ILVs. The idea that Sna3 can use eitherits own lysine ubiquitination or piggyback with otherubiquitinated proteins for MVB sorting explains previousresults showing Sna3 lacking its lysine residues fails tosort correctly in mutants with reduced Rsp5 activity, eventhough each of those mutant conditions alone does not
Traffic 2012; 13: 586–598 593

MacDonald et al.
Figure 7: The interdependence between Mup1 and Sna3 for MVB sorting. A) Mup1-GFP (green) was co-expressed in wild-typecells with either Sna3-mCherry (red) or Sna3-mCherry fused to the catalytic domain of Ubp7 (Sna3-mCherry-Ubp7). B) Localization inwild-type cells of Sna3-GFP mutant lacking its Rsp5-binding PY motif (Sna3�PY-GFP: upper left) or Mup1-mCherry-Ub (upper right).Lower panels show wild-type cells co-expressing Sna3�PY-GFP and Mup1-mCherry-Ub. Mup1-mCherry-Ub was induced under the CUP1promoter by addition of 100 μM CuSO4 to the growth media.
block Sna3 sorting (18). Moreover, our data underscorethe possibility that a wide variety of proteins may boosttheir MVB sorting efficiency by associating with otherubiquitinated proteins.
Similar Ub-independent routes into MVBs have beenproposed in mammalian cells (22,23,35,45,46). Althoughit is important to remember that there may be otherESCRT-independent ILV pathways such as generation of
594 Traffic 2012; 13: 586–598

Sna3 Is an Rsp5 Adaptor Protein
exosomes, the relevant issue for our studies is whethersorting along an ESCRT-dependent lysosomal degradativepathway into MVB ILVs occurs without Ub as a sortingsignal. Our data suggest that the basis of using Sna3 asan example of Ub-independent MVB sorting and applyingthat analogy to pathways in higher eukaryotes may beinappropriate. Likewise, the idea that a Ub-independentmechanism for Sna3 sorting fulfills a unique requirementfor formation of ILVs themselves, as proposed (47), nowseems unlikely. Studies on Sna3 have thus far beenfocused on its ability to serve as an MVB cargo. In thisstudy, we define a role for Sna3 as an Rsp5 adaptor forat least one substrate, the methionine transporter Mup1.This function overlaps with other Rsp5 adaptors such asArt1, which are required together for optimal methionine-induced delivery of Mup1 to the vacuole. A further test ofthis adaptor model might be to analyze the ubiquitinationstatus of Mup1 in sna3� cells. However, in log phase cells,Sna3 only contributes to the overall efficiency of Mup1trafficking, in contrast to the necessary role Art1 plays (11).Indeed, we speculate that Sna3 may play its major roleat later steps in the endocytic pathway after Art1 initiatesUb-dependent internalization from the plasma membrane.Regardless of exactly when Sna3 acts, it is likely to have amodest effect on the overall levels of ubiquitinated Mup1.Mup1 is known to be ubiquitinated by other proteins suchas Art1, which could operate in a variety of compartmentsother than where Sna3 may act. Experimentally, we foundit impractical to measure such subtle differences becauseMup1 readily oligomerizes once solubilized (Figure S2).Nonetheless, the idea that Sna3 is an Rsp5 adaptor isfurther supported by the block of Mup1 trafficking tothe vacuole caused by expression of Sna3-fused to aDUb as well as the ability of ‘pre-ubiquitinated’ Mup1(Mup1-mCherry-Ub) to pull Sna3�PY into the MVB pathway(Figure 7).
Collectively, our work indicates that Sna3 has a function asan Rsp5 adaptor protein that facilitates the ubiquitinationof Mup1 as well as perhaps a variety of other membraneproteins. Our data demonstrate that the quondamexample of Sna3 conforms to the canonical mode wherebyUb serves as the sorting signal for ESCRT-dependentMVB sorting. Moreover, it is the first to indicate that Sna3works as an Rsp5 adaptor. Sna3 itself has been shown tobe a very abundant protein within MVB ILVs suggestingthat it may facilitate ubiquitination and sorting of a widevariety of proteins besides Mup1 and the few othermembrane proteins identified in interaction screens. Wespeculate that the abundance of Sna3, its possible wideclient range, and its status as a privileged substrate forubiquitination by Rsp5 may help trap other ubiquitinatedproteins into a membrane subdomain where they can belargely deubiquitinated by Doa4 (thus recycling some Ubfrom vacuolar degradation) while collectively remaining aubiquitinated complex of proteins sufficient for completionof sorting into MVB ILVs.
Materials and Methods
ReagentsGeneral chemicals and antibodies against GFP, CPY, PGK and thehemagglutinin (HA) epitope sequence YPYDVPDYA were used aspreviously described (32). Yeast strains and plasmids used are listedin Tables 1 and 2, respectively.
Cell cultureYeast cultures were grown in standard minimal media supplemented withappropriate amino acids. Cells were generally grown overnight and thenresuspended in fresh media and grown to midlog phase. If a specific celldensity was required, optical density (OD) at 600 nm was monitored beforecells harvesting (OD600 = 1.0, or 2.0) and further processing. Expressionof proteins under the CUP1 promoter was induced by the addition of100 μM CuSO4 to the media. Mup1 internalization was induced by theaseptic addition of 20 μg/mL methionine to the media. To obtain the rsp5�
strain used in Figure 3, we circumvented the lethal requirement of Rsp5by genetic modification of MGA2, which encodes one of the functionallyredundant transcription factors required for the essential activation of theOLE1 gene (48). We integrated the myc-HIS3 sequence at basepair 2196 ofMGA2 to produce a truncated version of Mga2 lacking its trans-membranedomain, which can function without proteasome processing (49,50). Theplasmid encoding wild-type Rsp5 was then expelled from this strain using5-Fluoroorotic acid.
Fluorescence microscopyCells were grown to a midlog phase in minimal media before harvestingand resuspension in 100 mM Tris–HCl (pH = 8.0) buffer containing0.2% (wt/v) NaN3 and 0.2% (wt/v) NaF. Cells expressing GFP ormCherry fluorescent fusion proteins were then spotted onto a glassplate and viewed with an epifluorescence microscope (BX60; Olympus)with a 100× objective lens with numerical apperture (NA) 1.4. Cellswere viewed through either differential interference contrast (DIC) opticsor appropriate fluorescence filters at room temperature, and imageswere captured with a cooled charge-coupled device camera (Orca R2;Hamamatsu Photonics) using iVision-Mac software (Biovision Technology).Digital processing of images was performed in PhotoShop (version CS4;Adobe).
Trypsin protection assayOvernight cell cultures were used to inoculate 100 mL minimal media,which was grown over night to an OD600 = 1.0. Cells were then harvestedand incubated in 100 mM Tris pH 9.0 buffer, containing 50 mM dithiothreitol(DTT) for 5 min at room temperature. Cells were harvested again andresuspended in 1.2 M sorbitol, 50 mM KPO4 pH 7.5, 0.05% (wt/v) NaN3,0.05% NaFl, before addition of 100 (μL zymolase and incubation at 30◦Cfor 90 min. Spheroplasts were then layered onto a 5-mL solution of 1.2 M
sucrose, 50 mM KPO4 pH 7.5, 0.05% (wt/v) sodium azide, 0.05% sodiumfluoride in a 15-mL conical tube. Samples were centrifuged at 1000 × g
for 10 min, and supernatant discarded. Samples were then resuspendedin 1 mL ice cold intracellular lysis buffer (ILB; 100 mM KAc, 50 mM KCl,200 mM sorbitol, 20 mM PIPES pH 6.8) containing a protease inhibitorcocktail tablet (Roche) and stored on ice for 10 min. Sample was thencentrifuged at 400 × g on a tabletop centrifuge at 4◦C for 10 min. Thesupernatant was then transferred to a fresh Beckman ultrafuge tubeand subjected to high-speed centrifugation at 72 000 × g for 30 min.The pellet was resuspended in ILB lacking protease inhibitors, and therelative protein concentration was determined using a Bicinchoninic acid(BCA) assay. Samples were normalized to equal protein concentrationsand then separated into separated tubes and treated as indicated withILB alone, 25 (g/mL trypsin, and Triton X-100 at a final concentrationof 0.2% (v/v). Samples were incubated at room temperature for 30 minbefore an equal volume of 8 M urea, 5% SDS, 50 mM Tris pH 6.8 containingprotease inhibitors was added to each tube, and the samples were resolved
Traffic 2012; 13: 586–598 595

MacDonald et al.
Table 1: Yeast strains used in this study
Strain Genotype Reference
BY4742 MATα his3�0 leu2�0 lys2�0 ura3�0 (51)BY4742 YOR322C MATα art1� :: Kanr his3�0 leu2�0 lys2�0 ura3�0 (52)yPL3661 MATα pep4� ::LEU2 his3�0 lys2�0 ura3�0 This studyBY4742 YJL151C MATα sna3� :: Kanr his3�0 leu2�0 lys2�0 ura3�0 (52)yPL1781 MATα vps4� :: NEO, pep4-�1137, ura3-52, his-�200, leu2-3,112,
lys2-801, trp1-�901, suc2-�9This study
yPL4111 MATα ura3-52, leu2-3,112, ade2, his3-11, trp1-1, can1-100 rsp5�::LEU2,MGA2(1-732) :: 13myc-HIS3MX
This study
Table 2: Plasmids used in this study
Plasmid Description Reference
pRS313 Low copy yeast shuttle plasmid containing HIS3 marker (53)pRS315 Low copy yeast shuttle plasmid containing LEU2 marker (53)pRS316 Low copy yeast shuttle plasmid containing URA3 marker (53)Figure 1
pPL3667 pRS316 expressing Sna3-GFP from SNA3 promoter This studypPL2279 pRS316 expressing Sna3�K-GFP from SNA3 promoter. Sna3 (K19R K125R) This studypPL4618 pRS316 expressing Sna3-2 × HA from SNA3 promoter This studypPL4619 pRS316 expressing Sna3�K-2 × HA from SNA3 promoter. Sna3 (K19R K125R) This study
Figure 2pPL2089 pRS315 expressing Sna3-GFP from SNA3 promoter This studypPL3886 pRS316 expressing Sna3-GFP-Ubp7 from SNA3 promoter. Ubp7 residues 561–1071 This studypPL3904 pRS316 expressing Sna3-GFP-ubp7* from SNA3 promoter. pPL3904 with Upb7 C618S mutation This studypPL3874 pRS313 expressing Hse1-Ubp7 from CUP1 promoter. Ubp7 residues 561–1071 This studypPL4141 pRS315 expressing Hse1-ul36* from CUP1 promoter. Inactive UL36 catalytic domain (32)pPL3742 pRS316 expressing Rsp5-Ubp7-3 × HA from CUP1 promoter. Active Ubp7 catalytic domain (32)pPL4502 pRS316 expressing Rsp5-ubp7*-3 × HA from CUP1 promoter. Inactive Ubp7 catalytic domain (32)
Figure 3pPL4708 pRS315 expressing Sna3-GFP (pPL2089) from SNA3 promoter converted to ADE2 This studypPL4772 pPL4708 expressing HA-Rsp5C777A from RSP5 promoter This studypRS416-RSP5 pRS416 expressing Rsp5 from RSP5 promoter (54)
Figures 5 and 6pCHL642 pRS416 expressing Mup1-GFP from MUP1 promoter (11)pPL4621 pRS316 expressing Sna3�PY-2 × HA from SNA3 promoter. Sna3 P106PAY mutated to G106GAA This study
Figure 7pPL4146 pRS315 expressing Mup1-GFP from CUP1 promoter (32)pPL4064 pRS316 expressing Sna3-mCherry from SNA3 promoter This studypPL4066 pRS316 expressing Sna3-mCherry-Ubp7 from SNA3 promoter. Ubp7 residues 561–1071 This studypPL4566 pRS316 expressing Sna3�PY-GFP from SNA3 promoter. P106PAY mutated to G106GAA This studypPL4568 pRS316 expressing Mup1-mCherry-Ub from CUP1 promoter This study
by SDS-PAGE. SDS-PAGE and immunoblot analyses were performed asdescribed previously (32).
Acknowledgments
This work was supported by NIH R01GM58202 to R. C. P. We thank DavidKatzmann for helpful discussion.
Supporting Information
Additional Supporting Information may be found in the online version ofthis article:
Figure S1: GFP versions of Mup1 and Sna3 (including mutants: �K and�PY) were expressed in mup1� cells and grown to log phase beforeimaging localization of tagged proteins.
Figure S2: Wild-type cells were grown to midlog phase in minimal mediabefore addition of 20 μg/mL methionine. Samples were extracted fromcultures at 0, 20 and 40 min after methionine addition, harvested andresuspended in freshly made SDS/urea sample buffer. Samples wereresolved by SDS-PAGE followed by a 1-h incubation of the gel in transferbuffer containing 0.05% (wt/v) SDS. Gels were then subjected to a 3-htransfer in buffer containing SDS, before immunoblot analysis as described.Upper panel shows α-GFP immunoblot, where the insoluble proteinsaggregates that were unable to migrate through the gel can be observedat the top. The membrane was then stripped and reprobed with α-PGKantibodies as a loading control (lower panel).
Figure S3: Wild-type cells expressing Mup1-GFP were grown beyond logphase to an OD600 = 2.0 before imaging. Mup1-GFP containing cells wereeither co-expressing Sna3-mCherry (upper panels) or Sna3-mCherry-Ubp7(lower panels). A merged image depicts the co-localization of these twofluorescently labeled proteins.
596 Traffic 2012; 13: 586–598

Sna3 Is an Rsp5 Adaptor Protein
Please note: Wiley-Blackwell are not responsible for the content orfunctionality of any supporting materials supplied by the authors.Any queries (other than missing material) should be directed to thecorresponding author for the article.
References
1. d’Azzo A, Bongiovanni A, Nastasi T. E3 ubiquitin ligases asregulators of membrane protein trafficking and degradation. Traffic2005;6:429–441.
2. Komander D, Clague MJ, Urbe S. Breaking the chains: structureand function of the deubiquitinases. Nat Rev Mol Cell Biol2009;10:550–563.
3. Raiborg C, Stenmark H. The ESCRT machinery in endosomal sortingof ubiquitylated membrane proteins. Nature 2009;458:445–452.
4. Shields SB, Piper RC. How ubiquitin functions with ESCRTs. Traffic2011;12:1306–1317.
5. Lauwers E, Erpapazoglou Z, Haguenauer-Tsapis R, Andre B. Theubiquitin code of yeast permease trafficking. Trends Cell Biol2010;20:196–204.
6. Rotin D, Kumar S. Physiological functions of the HECT family ofubiquitin ligases. Nat Rev Mol Cell Biol 2009;10:398–409.
7. Chen HI, Einbond A, Kwak SJ, Linn H, Koepf E, Peterson S, KellyJW, Sudol M. Characterization of the WW domain of human yes-associated protein and its polyproline-containing ligands. J Biol Chem1997;272:17070–17077.
8. Sudol M, Chen HI, Bougeret C, Einbond A, Bork P. Characterizationof a novel protein-binding module--the WW domain. FEBS Lett1995;369:67–71.
9. Schild L, Lu Y, Gautschi I, Schneeberger E, Lifton RP, Rossier BC.Identification of a PY motif in the epithelial Na channel subunits asa target sequence for mutations causing channel activation found inLiddle syndrome. EMBO J 1996;15:2381–2387.
10. Staub O, Rotin D. Role of ubiquitylation in cellular membrane transport.Physiol Rev 2006;86:669–707.
11. Lin CH, MacGurn JA, Chu T, Stefan CJ, Emr SD. Arrestin-relatedubiquitin-ligase adaptors regulate endocytosis and protein turnover atthe cell surface. Cell 2008;135:714–725.
12. Nikko E, Pelham HR. Arrestin-mediated endocytosis of yeast plasmamembrane transporters. Traffic 2009;10:1856–1867.
13. Nikko E, Sullivan JA, Pelham HR. Arrestin-like proteins mediateubiquitination and endocytosis of the yeast metal transporter Smf1.EMBO Rep 2008;9:1216–1221.
14. Sullivan JA, Lewis MJ, Nikko E, Pelham HR. Multiple interactionsdrive adaptor-mediated recruitment of the ubiquitin ligase rsp5to membrane proteins in vivo and in vitro. Mol Biol Cell2007;18:2429–2440.
15. Hettema EH, Valdez-Taubas J, Pelham HR. Bsd2 binds the ubiquitinligase Rsp5 and mediates the ubiquitination of transmembraneproteins. EMBO J 2004;23:1279–1288.
16. Reggiori F, Pelham HR. Sorting of proteins into multivesicularbodies: ubiquitin-dependent and -independent targeting. EMBO J2001;20:5176–5186.
17. McNatt MW, McKittrick I, West M, Odorizzi G. Direct binding to Rsp5mediates ubiquitin-independent sorting of Sna3 via the multivesicularbody pathway. Mol Biol Cell 2007;18:697–706.
18. Oestreich AJ, Aboian M, Lee J, Azmi I, Payne J, Issaka R, Davies BA,Katzmann DJ. Characterization of multiple multivesicular body sortingdeterminants within Sna3: a role for the ubiquitin ligase Rsp5. MolBiol Cell 2007;18:707–720.
19. Stawiecka-Mirota M, Pokrzywa W, Morvan J, Zoladek T, Haguenauer-Tsapis R, Urban-Grimal D, Morsomme P. Targeting of Sna3pto the endosomal pathway depends on its interaction withRsp5p and multivesicular body sorting on its ubiquitylation. Traffic2007;8:1280–1296.
20. Katzmann DJ, Sarkar S, Chu T, Audhya A, Emr SD. Multivesicularbody sorting: ubiquitin ligase Rsp5 is required for the modification andsorting of carboxypeptidase S. Mol Biol Cell 2004;15:468–480.
21. Watson H, Bonifacino JS. Direct binding to Rsp5p regulatesubiquitination-independent vacuolar transport of Sna3p. Mol Biol Cell2007;18:1781–1789.
22. Chen J, Wang J, Meyers KR, Enns CA. Transferrin-directed internaliza-tion and cycling of transferrin receptor 2. Traffic 2009;10:1488–1501.
23. daSilva LL, Sougrat R, Burgos PV, Janvier K, Mattera R, BonifacinoJS. Human immunodeficiency virus type 1 Nef protein targets CD4 tothe multivesicular body pathway. J Virol 2009;83:6578–6590.
24. Tanowitz M, Von Zastrow M. Ubiquitination-independent traffickingof G protein-coupled receptors to lysosomes. J Biol Chem2002;277:50219–50222.
25. Hislop JN, Marley A, Von Zastrow M. Role of mammalianvacuolar protein-sorting proteins in endocytic trafficking of a non-ubiquitinated G protein-coupled receptor to lysosomes. J Biol Chem2004;279:22522–22531.
26. Babst M, Wendland B, Estepa EJ, Emr SD. The Vps4p AAA ATPaseregulates membrane association of a Vps protein complex requiredfor normal endosome function. EMBO J 1998;17:2982–2993.
27. Baba M, Osumi M, Scott SV, Klionsky DJ, Ohsumi Y. Twodistinct pathways for targeting proteins from the cytoplasm to thevacuole/lysosome. J Cell Biol 1997;139:1687–1695.
28. Silver P. Indirect immunofluorescence labeling in the yeastSaccharomyces cerevisiae. Cold Spring Harb Protoc 2009;2009:pdb.prot5317.
29. Wang X, Herr RA, Hansen TH. Ubiquitination of substrates byesterification. Traffic 2011.
30. Ciechanover A, Ben-Saadon R. N-terminal ubiquitination: more proteinsubstrates join in. Trends Cell Biol 2004;14:103–106.
31. Kim HC, Huibregtse JM. Polyubiquitination by HECT E3s and the deter-minants of chain type specificity. Mol Cell Biol 2009;29:3307–3318.
32. Stringer DK, Piper RC. A single ubiquitin is sufficient for cargo proteinentry into MVBs in the absence of ESCRT ubiquitination. J Cell Biol2011;192:229–242.
33. Kim HC, Steffen AM, Oldham ML, Chen J, Huibregtse JM. Structureand function of a HECT domain ubiquitin-binding site. EMBO Rep2011;12:334–341.
34. Kamadurai HB, Souphron J, Scott DC, Duda DM, Miller DJ, StringerD, Piper RC, Schulman BA. Insights into ubiquitin transfer cascadesfrom a structure of a UbcH5B approximately ubiquitin-HECT(NEDD4L)complex. Mol Cell 2009;36:1095–1102.
35. Hanyaloglu AC, von Zastrow M. A novel sorting sequence in thebeta2-adrenergic receptor switches recycling from default to theHrs-dependent mechanism. J Biol Chem 2007;282:3095–3104.
36. Helliwell SB, Losko S, Kaiser CA. Components of a ubiquitin ligasecomplex specify polyubiquitination and intracellular trafficking of thegeneral amino acid permease. J Cell Biol 2001;153:649–662.
37. Soetens O, De Craene JO, Andre B. Ubiquitin is required for sortingto the vacuole of the yeast general amino acid permease, Gap1. J BiolChem 2001;276:43949–43957.
38. Yashiroda H, Kaida D, Toh-e A, Kikuchi Y. The PY-motif of Bul1 proteinis essential for growth of Saccharomyces cerevisiae under variousstress conditions. Gene 1998;225:39–46.
39. Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, StorzG, Botstein D, Brown PO. Genomic expression programs in theresponse of yeast cells to environmental changes. Mol Biol Cell2000;11:4241–4257.
40. Tarassov K, Messier V, Landry CR, Radinovic S, Serna Molina MM,Shames I, Malitskaya Y, Vogel J, Bussey H, Michnick SW. An in vivomap of the yeast protein interactome. Science 2008;320:1465–1470.
41. Hicke L, Dunn R. Regulation of membrane protein transport byubiquitin and ubiquitin-binding proteins. Annu Rev Cell Dev Biol2003;19:141–172.
42. Paiva S, Vieira N, Nondier I, Haguenauer-Tsapis R, Casal M, Urban-Grimal D. Glucose-induced ubiquitylation and endocytosis of the yeastJen1 transporter: role of lysine 63-linked ubiquitin chains. J Biol Chem2009;284:19228–19236.
43. Beck T, Schmidt A, Hall MN. Starvation induces vacuolar targetingand degradation of the tryptophan permease in yeast. J Cell Biol1999;146:1227–1238.
44. Galan JM, Haguenauer-Tsapis R. Ubiquitin lys63 is involved inubiquitination of a yeast plasma membrane protein. EMBO J1997;16:5847–5854.
45. Henry AG, White IJ, Marsh M, von Zastrow M, Hislop JN. The role ofubiquitination in lysosomal trafficking of delta-opioid receptors. Traffic2010;12:170–184.
Traffic 2012; 13: 586–598 597

MacDonald et al.
46. Yamashita Y, Kojima K, Tsukahara T, Agawa H, Yamada K, Amano Y,Kurotori N, Tanaka N, Sugamura K, Takeshita T. Ubiquitin-independentbinding of Hrs mediates endosomal sorting of the interleukin-2receptor beta-chain. J Cell Sci 2008;121:1727–1738.
47. Nickerson DP, Russell MR, Odorizzi G. A concentric circle model ofmultivesicular body cargo sorting. EMBO Rep 2007;8:644–650.
48. Zhang F, Chen Y, Kweon DH, Kim CS, Shin YK. The four-helix bundle ofthe neuronal target membrane SNARE complex is neither disorderedin the middle nor uncoiled at the C-terminal region. J Biol Chem2002;277:24294–24298.
49. Chellappa R, Kandasamy P, Oh CS, Jiang Y, Vemula M, Martin CE.The membrane proteins, Spt23p and Mga2p, play distinct roles inthe activation of Saccharomyces cerevisiae OLE1 gene expression.Fatty acid-mediated regulation of Mga2p activity is independent ofits proteolytic processing into a soluble transcription activator. J BiolChem 2001;276:43548–43556.
50. Hoppe T, Matuschewski K, Rape M, Schlenker S, UlrichHD, Jentsch S. Activation of a membrane-bound transcription
factor by regulated ubiquitin/proteasome-dependent processing. Cell2000;102:577–586.
51. Brachmann CB, Davies A, Cost GJ, Caputo E, Li J, Hieter P, BoekeJD. Designer deletion strains derived from Saccharomyces cerevisiaeS288C: a useful set of strains and plasmids for PCR-mediated genedisruption and other applications. Yeast 1998;14:115–132.
52. Winzeler EA, Shoemaker DD, Astromoff A, Liang H, AndersonK, Andre B, Bangham R, Benito R, Boeke JD, Bussey H, ChuAM, Connelly C, Davis K, Dietrich F, Dow SW, et al. Functionalcharacterization of the S. cerevisiae genome by gene deletion andparallel analysis. Science 1999;285:901–906.
53. Sikorski RS, Hieter P. A system of shuttle vectors and yeast hoststrains designed for efficient manipulation of DNA in Saccharomycescerevisiae. Genetics 1989;122:19–27.
54. Wang G, Yang J, Huibregtse JM. Functional domains of the Rsp5ubiquitin-protein ligase. Mol Cell Biol 1999;19:342–352.
598 Traffic 2012; 13: 586–598