somatic motility and hair bundle mechanics, are both ......somatic motility and hair bundle...
TRANSCRIPT

lable at ScienceDirect
Hearing Research 273 (2011) 109e122
Contents lists avai
Hearing Research
journal homepage: www.elsevier .com/locate/heares
Somatic motility and hair bundle mechanics, are both necessary forcochlear amplification?
Anthony W. Peng a, Anthony J. Ricci a,b,*aDepartment of Otolaryngology, Stanford University, Stanford CA 94305, USAbDepartment of Molecular and Cellular Physiology, Stanford University, Stanford CA 94305, USA
a r t i c l e i n f o
Article history:Received 7 January 2010Received in revised form2 March 2010Accepted 8 March 2010Available online 27 April 2010
* Corresponding author at: Department of MoleculaPasteur Dr. Edwards Bldg R145, Stanford University, Sþ650 736 1290; fax: þ650 725 8502.
E-mail address: [email protected] (A.J. Ricci).
0378-5955/$ e see front matter � 2010 Elsevier B.V.doi:10.1016/j.heares.2010.03.094
a b s t r a c t
Hearing organs have evolved to detect sounds across several orders of magnitude of both intensity andfrequency. Detection limits are at the atomic level despite the energy associated with sound beinglimited thermodynamically. Several mechanisms have evolved to account for the remarkable frequencyselectivity, dynamic range, and sensitivity of these various hearing organs, together termed the activeprocess or cochlear amplifier. Similarities between hearing organs of disparate species provides insightinto the factors driving the development of the cochlear amplifier. These properties include: a tonotopicmap, the emergence of a two hair cell system, the separation of efferent and afferent innervations, therole of the tectorial membrane, and the shift from intrinsic tuning and amplification to a more end organdriven process. Two major contributors to the active process are hair bundle mechanics and outer haircell electromotility, the former present in all hair cell organs tested, the latter only present in mammaliancochlear outer hair cells. Both of these processes have advantages and disadvantages, and how theseprocesses interact to generate the active process in the mammalian system is highly disputed. Ahypothesis is put forth suggesting that hair bundle mechanics provides amplification and filtering inmost hair cells, while in mammalian cochlea, outer hair cell motility provides the amplification on a cycleby cycle basis driven by the hair bundle that provides frequency selectivity (in concert with the tectorialmembrane) and compressive nonlinearity. Separating components of the active process may provideadditional sites for regulation of this process.
� 2010 Elsevier B.V. All rights reserved.
1. Introduction
Comparative biology is an excellent tool for identifying selectivepressures shaping the evolution of particular systems. The auditorysystem has conserved properties throughout evolution to over-come common physical obstacles (Fig. 1), while modifying otherproperties for hearing sensitivity, frequency range, and frequencyselectivity (Manley and Koppl, 2008). The evolution and modifi-cation of the system demonstrates how a comparative approachcan identify driving forces behind particular traits. The primaryfunction of the peripheral auditory system is to convert complexairborne vibration into its fundamental frequency components,transmitting information regarding frequency, intensity and timingto the central nervous system. Several independent mechanismshave evolved to accomplish this daunting task and the relative
r and Cellular Physiology, 300tanford CA 94305, USA. Tel.:
All rights reserved.
contribution of each to human hearing remains to be resolved. Thepurpose of the present work is to attempt to gain insight intomechanisms of cochlear amplification and how evolution may havedriven the continued adaptations of the system. Given the abun-dance of excellent reviews on cochlear amplification, somaticmotility, hair bundle mechanics, and comparative hearing(Ashmore, 2008; Dallos, 2008; Fettiplace and Fuchs, 1999;Fettiplace and Hackney, 2006; Hudspeth, 2008; Manley, 2000b,2001), the present work will focus less on the mechanistic andhistorical details associated with each process and more on thestrengths and weaknesses provided by the potential mechanisms.
2. Cochlear amplification
The auditory system differs from other sensory systems in that itis thermodynamically limited; at least for free standing bundles likethose of IHC, the energy associated with auditory detection iscomparable to the energy associated with Brownian motion of thesensory hair bundle (the detector) (Bialek, 1987; Bialek andSchweitzer, 1985; de Vries, 1948a; de Vries, 1948b; Harris, 1968;

Fig. 1. Evolution of hearing organs. An evolutionary tree of the various organismsdiscussed in this review shows that insects and amphibians are furthest frommammals, and stem reptiles (red) are the closest. Adapted from (Manley and Koppl,1998).
A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122110
Hudspeth, 1985). Given the similarity in threshold levels acrossspecies, this energy obstacle was surmounted, perhaps in differentways, by different auditory end organs. Having to detect such lowenergies at threshold may also be responsible for evolving activefrequency selectivity mechanisms, as reducing the frequency rangeof any given detector will limit the thermal noise it must overcome.Post filtering amplification can then be used to further enhance thesignal to noise ratio. The requirement of frequency selectivity priorto amplification is important and may be a driving feature that ledto the conservation of the tonotopic organization across auditoryend organs. Tonotopy is simply how a broad frequency range at thesystem level is produced by the concerted effort of individualsensory cells responding over narrow bands in an ordered manner.Tonotopy is present in species as diverse as crickets and katydids,attesting to its fundamental importance in auditory detection(Romer, 1983; Stumpner and Molina, 2006). Amplification andfiltering alone are not enough to account for the dynamic range ofauditory systems. The human auditory system not only detectsextremely low level sounds, but also operates in a noisy environ-ment and adapts to respond without saturation to sounds thatencompass 12 orders of magnitude in intensity (Moore, 2004). Toaccomplish this feat, auditory systems use a nonlinear amplifierwhose gain varies relative to the input intensity. When sounds areloud the gain is lowered, when sound is low, the gain is raised, thusproviding modulation of input across a broad range of stimulusintensities (Johnstone et al., 1986; Rhode, 1978; Rhode and Robles,1974). This feature, termed compressive nonlinearity, indicates aninput-output function that follows a power law such that theoutput scales as one-third the power of input (Ruggero et al., 1997).Deprivation of metabolic energy to the auditory end organs resultsin a rapid loss of mechanical amplification with thresholdselevating as much as 60 dB, demonstrating that signal processing isnot passive (Davis et al., 1989; Ruggero and Rich, 1991). Concomi-tant with this threshold elevation is a broadening of the frequencyresponse (loss of tuning) and a loss of compressive nonlinearity,illustrating the interaction between these components and thehighly metabolically active nature of the system (Hudspeth, 2008).Together these three properties, compressive nonlinearity,frequency selectivity and amplification, constitute the cochlearamplifier or active process. A fourth property, more a byproductthan a function of the active process, is the ability to providemechanical energy that feeds back through the system to generatesound (Kemp, 1986; Probst et al., 1991). Termed spontaneous
otoacoustic emissions (SOAEs), they are considered a hallmark ofthe active process and demonstrate an active mechanical contri-bution by the cochlear amplifier (Brownell, 1990). Multiple mech-anisms probably underlie the active process, and the relativecontribution of each may vary between species. Thus mechanismsunderlying the active process would be expected to produce each ofthe four associated characteristics (Hudspeth, 2008; Manley andKoppl, 2008).
3. Signal transduction
Several properties of signal transduction are common betweenauditory organs. Airborne vibration is converted, typically via themiddle ear, into fluid vibrations that stimulate the inner ear sensorycells, called hair cells. Fluid vibrations can stimulate directly viafluid induced movement of the sensory hair bundles or indirectlyby causing differential movement of the basilar membrane (BM)(upon which the hair cells sit) and the tectorial membrane (TM) (amembrane in which multiple sensory hair bundles are embedded).The method of stimulation that predominates in a given end organlargely depends on the specific end organ structure.
Hair cells are specialized epithelial cells whose apical pole hasa protrusion of actin filled microvilli (termed stereocilia, thoughstereovilli would be a more appropriate term) of increasing height(Fig. 2) (Tilney and Saunders, 1983). The size and shape of the hairbundle varies between and within end organs, but typically there isa stair-cased array oriented such that deflection toward the talleststereocilia opens mechanosensitive channels located near the topsof these stereocilia, and stimulation away from the tall end, closeschannels (Fig. 2) (Hudspeth, 1982; Hudspeth and Corey, 1977;Hudspeth and Jacobs, 1979; Jaramillo and Hudspeth, 1991;Shotwell et al., 1981; Tilney and Saunders, 1983). Mechanosensi-tivity is posited to be imparted by a thin filamentous link, the tip-link, composed of protocadherin 15 and cadherin 23 (Ahmed et al.,2006; Kazmierczak et al., 2007; Pickles et al., 1984; Siemens et al.,2004). This link connects the top of shorter stereocilia with theside of longer stereocilia, oriented along the staircase patternproviding directional sensitivity (Shotwell et al., 1981). Whereinvestigated, mechanotransduction responses show an adaptivebehavior, a reduction in response (current amplitude) duringa continued stimulation (Fig. 2) (Crawford et al., 1989; Eatock et al.,1987). Adaptation is implicated in a variety of functions thatinclude: increasing the dynamic range of the hair cell, maintainingthe hair bundle in its most sensitive position, providing mechanicalfiltering and amplification, and setting the hair cell resting potential(Eatock, 2000; Eatock et al., 1987; Farris et al., 2006; Fettiplace,2006; Hudspeth, 2008; Ricci et al., 2005). Most of the proteinsinvolved in mechanosensitivity and adaptation remain to be iden-tified, so the specifics of the underlying molecular mechanismshave yet to be elucidated. As more molecules and mechanisms areidentified, better experiments will be possible to discern thespecific role of mechanotransduction and adaptation in establish-ing the cochlear amplifier. The functions associatedwith adaptationwill in part dictate what mechanisms might be plausible; forexample setting the operating point of the bundle is less kineticallydemanding than providing tuning and amplification on a cycle bycycle basis.
4. Comparison of auditory properties
From insects to mammals, hearing evolved separately (Fig. 1)resulting in differences in hearing organ structure, yet manycommonalities exist between organs that may give insight intofactors driving their development (Table 1). These evolutionarydifferences in hearing structure produced distinctive properties in

Fig. 2. Principles of mammalian hair cell mechanisms. (A) Stereocilia at the apex of the hair cell are responsible for mechanotransduction. Positive deflection of stereocilia (black)causes an opening of the mechanotransduction channels leading to an influx of cations into the cell. The calcium component of the current drives an adaptation process, a reductionin current during a constant stimulus that is thought to underlie force generation by the hair bundle. Negative deflection of the stereocilia (red) causes transduction channels toclose, triggering reverse adaptation back to the resting level. Asterisks indicate potential sites of force generation. (B) Somatic motility occurs in the lateral membrane of the hair cell(cross-hatched area). A hyperpolarization of membrane voltage leads to an increase in the lateral membrane surface area, hence an expansion of the hair cell. A depolarization inmembrane voltage leads to a decrease in lateral membrane surface area and a contraction of the hair cell. (C) Cochlear mechanisms at work in the mammalian cochlea include activehair bundle motions coupled through the tectorial membrane (black arrow) and somatic motility (blue arrows) which feedback onto basilar membrane motion (red arrow).
A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 111
frequency selectivity and hearing thresholds. The commonalitiesbetween these diverse species elucidate the underlying principlesof cochlear amplification required for hearing sensitivity andfrequency selectivity.
A comparison of performance across species reveals similaritiesbetween species. Hearing thresholds across species are comparableacross a broad range of frequencies (Fig. 3a). Tuning curves can becompared for different species using center frequencies near thelowest thresholds (Fig. 3b). These curves have a Q10dB associatedwith them, but because center frequencies vary considerablybetween species and Q10dB values vary widely depending onfrequency, the stated values are used for a rough comparison andcannot be strictly interpreted in terms of sensitivity or selectivity.The plot does demonstrate a similarity in tuning between speciesthat is remarkable given the differences in end organ structures andmechanisms. The differences observed in performance appear to bemore correlated to frequency range, for example the bird andmammal curves show sharper high frequency rolloffs; otherdifferences such as thresholds may be associated with middle earproperties rather than end organ detection limits.
Insects have an auditory structure that is different from mostother organisms studied. Insects have two types of hearing organs:an antennal organ and a tympanal organ. The antennal organtransduces vibrations of antennae, and the tympanal organ
transduces vibrations of a tympanic-like membrane (Kossl et al.,2008; Yack, 2004). The organs do not possess the classical struc-ture of a hair cell with an apical sensory hair bundle, but ratherhave a single cilium that acts as a mechanoreceptor. Despite thestark morphological differences, mechanoreceptors may have thesame evolutionary origin dating to early metazoans (Manley andLadher, 2008). Similar to many hearing organs, the tympanalorgan possesses a tonotopic map with tuned receptors along theorgan (Kossl et al., 2008; Oldfield, 1988; Oldfield et al., 1986). Thetympanal organs are tuned to frequencies between 1 kHz and100 kHz being limited only at lower frequencies (Kossl et al., 2008).Receptor cells of the Ancistrura nigrovittata exhibit a characteristicV-shaped tuning curvewith a Q10dB of 1.9 at the lowest threshold of30 dB SPL at 20 kHz (Fig. 3; Table 1) (Stumpner and Molina, 2006).The sensory organelle is also embedded in an apical extracellularstructure (scolopale cap) (Slifer and Sekhon, 1975; Yack, 2004),similar to other hearing organs that have hair bundles embedded ina tectorial membrane. Despite insects evolving independently(Fig. 1) they still possess a similar basis of hearing detection withmechanoreceptor organelles embedded in an apical extracellularmatrix and cells that are frequency tuned and organized ina tonotopic pattern. Whether this is convergence in problemsolving or due to mechanoreceptors having similar origins is opento interpretation.
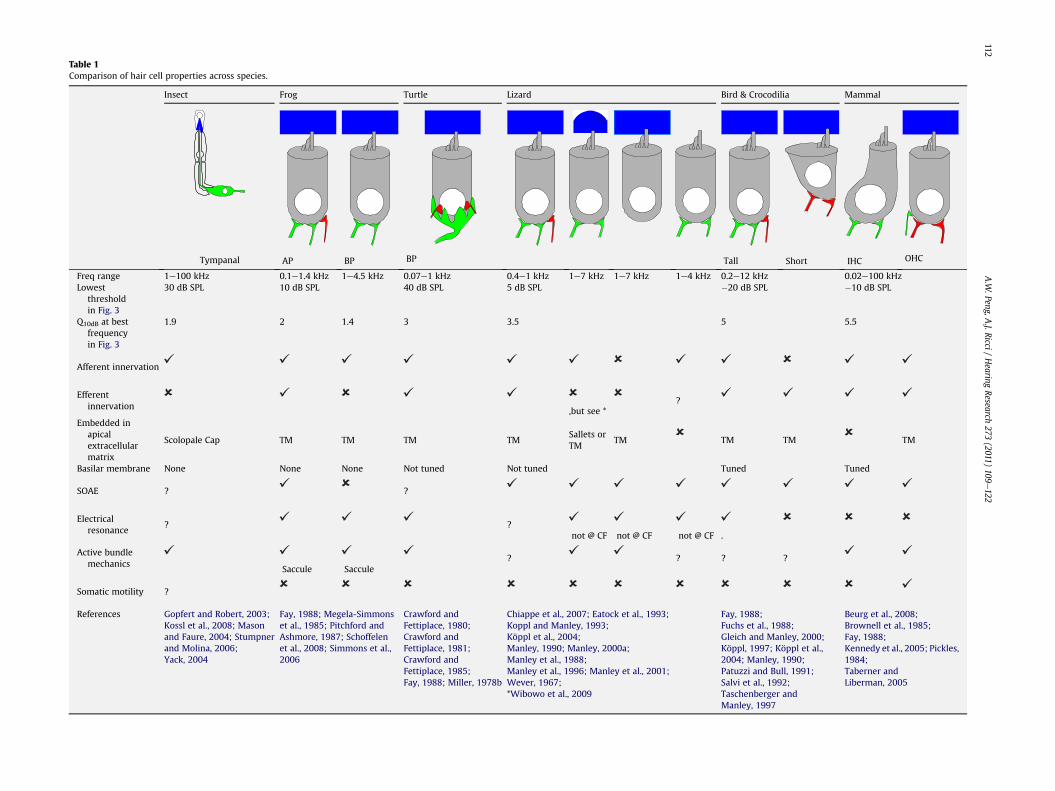
Table 1Comparison of hair cell properties across species.
Insect Frog Turtle Lizard Bird & Crocodilia Mammal
Tympanal AP BP BP Tall Short IHC OHC
Freq range 1e100 kHz 0.1e1.4 kHz 1e4.5 kHz 0.07e1 kHz 0.4e1 kHz 1e7 kHz 1e7 kHz 1e4 kHz 0.2e12 kHz 0.02e100 kHzLowest
thresholdin Fig. 3
30 dB SPL 10 dB SPL 40 dB SPL 5 dB SPL �20 dB SPL �10 dB SPL
Q10dB at bestfrequencyin Fig. 3
1.9 2 1.4 3 3.5 5 5.5
Afferent innervation
Efferentinnervation ,but see *
?
Embedded inapicalextracellularmatrix
Scolopale Cap TM TM TM TMSallets orTM
TM TM TM TM
Basilar membrane None None None Not tuned Not tuned Tuned Tuned
SOAE ? ?
Electricalresonance
? ?not @ CF not @ CF not @ CF .
Active bundlemechanics
Saccule Saccule? ? ? ?
Somatic motility ?
References Gopfert and Robert, 2003;Kossl et al., 2008; Masonand Faure, 2004; Stumpnerand Molina, 2006;Yack, 2004
Fay, 1988; Megela-Simmonset al., 1985; Pitchford andAshmore, 1987; Schoffelenet al., 2008; Simmons et al.,2006
Crawford andFettiplace, 1980;Crawford andFettiplace, 1981;Crawford andFettiplace, 1985;Fay, 1988; Miller, 1978b
Chiappe et al., 2007; Eatock et al., 1993;Koppl and Manley, 1993;Köppl et al., 2004;Manley, 1990; Manley, 2000a;Manley et al., 1988;Manley et al., 1996; Manley et al., 2001;Wever, 1967;*Wibowo et al., 2009
Fay, 1988;Fuchs et al., 1988;Gleich and Manley, 2000;Köppl, 1997; Köppl et al.,2004; Manley, 1990;Patuzzi and Bull, 1991;Salvi et al., 1992;Taschenberger andManley, 1997
Beurg et al., 2008;Brownell et al., 1985;Fay, 1988;Kennedy et al., 2005; Pickles,1984;Taberner andLiberman, 2005
A.W
.Peng,A.J.Ricci
/Hearing
Research273
(2011)109
e122
112

Fig. 3. Comparison of threshold and tuning curves across vertebrate species. (A) Audiograms were obtained from (Fay, 1988) for all species except the bobtail skink which camefrom (Manley, 2000a) and the bush cricket from (Stumpner and Molina, 2006). All curves are behavioral audiograms except for the bobtail skink and the bush cricket which areneural audiograms. (B) Tuning curves for each organism were chosen with center frequencies near their lowest threshold in order to compare the shapes of tuning curves acrossspecies, therefore the sharpness of tuning (Q10dB) cannot be strictly compared here because these values vary depending on center frequencies in different species. Centerfrequencies for the tuning curves and data sources are as follows: chinchilla 8.1 kHz (Ruggero et al., 1990), guinea pig 7 kHz (Pickles, 1984), mouse 10 kHz (Taberner and Liberman,2005), chicken 1.7 kHz (Salvi et al., 1992), barn owl 4.2 kHz (Köppl, 1997), bobtail skink 1.2 kHz (Manley et al., 1988), turtle 330 Hz (Crawford and Fettiplace, 1980), frog 751 Hz(Stiebler and Narins, 1990), bush cricket 20 kHz (Stumpner and Molina, 2006).
A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 113
The amphibian auditory organ, like insects, is also distinct fromthe stem reptiles’ auditory organs because it has two end organs:the amphibian papilla and the basilar papilla. Despite this alter-native organization, common properties of tonotopy and frequencytuned receptor cells are conserved at least in the amphibian papilla(see van Dijk in this special edition). The change in organ structurecorrelates with a shift in the frequency range to 0.1e4.5 kHz withlowered thresholds of 10 dB SPL at 20 kHz and low Q10dB (Table 1,Fig. 3) (Fay, 1988; Stiebler and Narins, 1990). With these changes,a hair cell having a distinct hair bundle composed of stereocilia thatpossess mechanically gated ion channels, graded stereociliaheights, and directional sensitivity emerges (Schoffelen et al.,2008). The similarity of this sensory cell to other chordate groupsis discussed in more detail by Burighel et al. in this issue. The hairbundle for this cell type is also embedded in a tectorial membranestructure on its apical surface (Lewis, 1984).
In a primitive land animal, the turtle, often considered similar tothe stem reptile papilla, but having evolved independently, addi-tional features evolved at the system level. The basilar papilla, theturtle hearing organ, consists of hair cells sitting atop a flexiblebasilar membrane, a membrane found in all vertebrates derivedfrom the stem reptiles. The basilar papilla organization is primitive,consisting of multiple rows of a single hair cell type, orientedsimilarly (Manley, 2000b; Miller, 1978b). The frequency range ofthe organ is narrow (0.07e1 kHz) and thresholds are relativelyhigh, 40 dB SPL at 200 Hz for the Trachemys scripta; however, the
sharpness of tuning is increased to a Q10dB of 3 (Table 1, Fig. 3)(Crawford and Fettiplace, 1980; Fay, 1988). Higher thresholds inturtle have been ascribed to a thicker eardrum as opposed toa difference in detection at the sensory cell (see other papers in thisissue). Auditory papilla hair bundles vary in size, with shorterstereocilia at high frequency regions and longer stereocilia at lowfrequency regions (Hackney et al., 1993), another characteristicfound in all vertebrate auditory organs derived from stem reptiles.
Lizards become more complex with different hair bundleorientations and multiple hair cell types (Table 1). Some lizardbasilar papilla hair bundles are oriented in opposing directions,particularly in the high frequency tuned cells (Manley, 2000a;Miller, 1973, 1978a). The frequency range of lizards stretches from0.4 to 7 kHz. Thresholds lower to 5 dB SPL at 1.2 kHz for the Tiliquarugosa and Q10dB is about the same as the turtle at 3.5 (Fig. 3, Table1) (Manley, 1990; Manley et al., 1988, 1990). The lizard has multipletypes of hair cells. One hair cell type receives afferent and efferentinnervations and is responsible for low frequency hearing (Manley,1990, 2000b), and a second type of hair cell only has afferentinnervations and is responsible for high frequency hearing. Thesehair cells are covered by apical sallets (much like strips of tectorialmembrane) or by continuous tectorial membrane (Manley, 1990,2000b). A third type of cell is not embedded in any apicalmembrane and is referred to as a free standing hair bundle cell(Mulroy and Williams, 1987). A fourth type of hair cell is found inthe tokay gecko, may exist in other lizard species as well, is located

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122114
on the neural high frequency region of the basilar papilla, is coveredby a tectorial membrane, and has no innervation (Chiappe et al.,2007). The functional role of these cells remains unknown but itis not a large leap to hypothesize that these cells will be involved ina mechanical type of tuning and amplification that feeds back ontoinnervated cells.
Birds and crocodilia continue to increase the complexity of theirauditory end organ structure. Frequency range is further extended(0.2e12 kHz) and thresholds continue to lower to �20 dB SPL at4 kHz with high Q10dB reaching 5 for the Tyto Alba (Fig. 3, Table 1)(Köppl,1997). The basilar papilla contains hair bundleswith varyingangular orientations and the structure contains two distinct sets ofhair cells based on cell morphology and neural innervation (Fischeret al., 1988; Gleich and Manley, 1988; Manley, 2000b; Tanaka andSmith, 1978; Tilney and Saunders, 1983; Tilney et al., 1987). Tallhair cells, located on the neural side of the basilar papilla, areinnervated by both afferent and efferent nerves; whereas, short haircells, located abneurally, are innervated solely by efferent nervefibers (Fischer, 1994, 1998). Additionally, a tuned basilar membranemotion is suggested (Gummer et al., 1987). Many properties in birdsand crocodilia have parallels in the mammal, attesting to theirsignificance as ascribed by the evolutionary convergence.
Mammals havewhat appear to be themost sophisticated auditoryorganization with the widest range of audible frequencies(0.02e100kHz) and low thresholds (�10dBSPL)with aQ10dB of 4.2 at8 kHz in the Cavia porcellus (Table 1, Fig. 3) (Fay, 1988; Pickles, 1984).The auditory organ consists of a coiled cochleawhere all sensory hairbundlesareorientedaway fromthecenterof thecoil. Themammaliancochlea contains two types of hair cells, the inner hair cells (IHCs) thatreceive the majority of afferent innervations and the outer hair cells(Ehret, 1979; Morrison et al., 1975), (OHCs) that receive efferentinnervation synapsing directly onto the hair cell (Warr, 1992). Thebasilar membrane motion is actively tuned and amplified by OHCs(Rhode, 1971; Rhode and Robles, 1974; Ruggero et al., 1997). Despitewhatatfirst appear tobemajorchanges to theorganizationof theendorgan, mammalian hearing is quite comparable to that of otherspecieswith sensitivities being comparable or in some cases less thanthat found in some birds and lizards.
As the mammalian system incorporates many structural prop-erties found in other hearing organs, insight into functionmay arisefrom a better understanding of the commonalities of these varioushearing organs. For example, the use of both free standing andembedded hair bundles, the divergence in innervation patternsbetween hair cells, and the apparent separation of functions, withone cell type being responsible for amplification and the other forsignal transduction to the brain, are all common features found inspecies aside from themammal. Asmammals have the only hearingorgan with a somatically motile cell type an important questionbecomes: are the preexisting mechanisms of cochlear amplificationin non-mammalian systems lost or obsolete in the face of thedevelopment of electromotility, or are they incorporated andadvanced by the emergence of a new mechanical component?What is the advantage of somatic motility over preexisting ampli-ficationmechanisms? The remaining portions of this article discussthe known mechanisms of amplification and tuning focusing onelectrical resonance, hair bundle mechanics and outer hair cellelectromotility, discussing the strengths, weaknesses and areaswhere more work is needed for each. Finally, a hypothesis isproposed that incorporates the strengths of each mechanism asa possible means of generating a mammalian amplifier.
5. Electrical resonance as an amplification mechanism
Electrical resonance is one of the first tuning and amplificationmechanisms identified in auditory hair cells (Crawford and
Fettiplace, 1978). The interaction of L-type calcium channels andBK-type potassium channels coupled with the membrane capaci-tance creates an inductive circuit oscillating about the cell’s char-acteristic frequency (Crawford and Fettiplace, 1978,1981; Fettiplaceand Crawford, 1978; Fuchs and Evans, 1988; Fuchs et al., 1988;Hudspeth and Lewis, 1988; Pitchford and Ashmore, 1987). Tono-topic variations in the number of calcium and calcium-activatedpotassium channels as well as the activation and deactivationkinetics of potassium channels create a frequency map (Art andFettiplace, 1987; Jones et al., 1999; Wu et al., 1995). Electricalresonance is a major tuning mechanism in turtles, frogs, and birds(Table 1 and 2). A role in lizards remains to be established; electricalresonance has been observed in the lizard, but resonant frequenciesdo not correlate with characteristic frequencies of the cell (Chiappeet al., 2007; Eatock et al., 1993). Theoretical analysis of electricalresonance suggests that it can operate up to about 6 kHz (Wu et al.,1995), though even in higher frequency hearing organs like bird,resonant frequencies have not been directly measured from thehair cell above about 1 kHz; however preferred interval measure-ments in afferent fibers from bird suggest resonant frequencies upto 5 kHz (Köppl, 1997). No evidence for electrical resonance existsin mammalian hair cells. Electrical resonance is mentioned in thecontext of cochlear amplification because it is suggested to providethe tuning and amplification required to completely account forafferent fiber tuning curves; however, it has no mechanical corre-late, and therefore, no spontaneous otoacoustic emissions aregenerated by this mechanism. Thus by definition, electrical reso-nance cannot be the only mechanism responsible for cochlearamplification in these species.
Paradoxically, the strongest data supporting a role for active hairbundle mechanics in cochlear amplification comes from end organsthat exhibit electrical resonance (Fettiplace, 2006; Fettiplace et al.,2001; Hudspeth, 2005, 2008). In fact, the first report of active,tuned hair bundle motions comes from the turtle auditory papillaand may reflect the first example of electromechanical feedback(Crawford and Fettiplace, 1985). Similarly, strong data supportsactive hair bundle oscillations in frog saccule hair bundles, anotherorgan where electrical resonance is quite robust (Bozovic andHudspeth, 2003; Hudspeth and Lewis, 1988; Martin et al., 2003).Yet no data exists investigating the interplay between hair bundleoscillations and electrical resonance. How hair bundle tuningoperates with respect to electrical resonance remains to be eluci-dated and may be important for defining the principles and limi-tations of electromechanical feedback.
Mammalian hair cells show no electrical tuning properties. Ingeneral, both outer and inner hair cells tend to behave as low passfilters, with low input resistances reducing the membrane timeconstant but no obvious frequency selectivity (Dallos, 1985; Russellet al., 1989). Thus electrical resonance represents an example ofa totally cellular mechanism being responsible for tuning andamplification, yet it is completely absent in mammalian hair cellsand replaced by what appears to be a more systems based tuningmechanism.
6. Hair bundles as the cochlear amplifier
That hearing thresholds and frequency selectivity are compa-rable across species attests to the argument that an active processexists in hearing organs other than mammals (Table 1; Fig. 3). Theability to measure otoacoustic emissions from non-mammalianspecies further supports a mechanical mechanism of amplificationwith conserved properties (Koppl and Manley, 1993; Manley et al.,1987, 1996, 2001; Taschenberger and Manley, 1997; van Dijk et al.,1989). Pharmacological sensitivities of these same emissionsbetween mammalian and non-mammalian species argue for some

Table
2Prop
erties
ofco
chlear
amplifi
cation
.
Amplifi
cation
Freq
uen
cyTu
ning
Com
pressiveNon
-linea
rity
Electrical
Reson
ance
Pro
:-Amplifi
essign
alswhen
stim
ulatedat
resonan
tfreq
uen
cyCo
n:-
Reson
antfreq
uen
cies
only
uptoe6kH
zPro
:-Haircellelectrical
tuningch
aracteristicsab
leto
completely
acco
untforafferenttuningproperties
Con:-
Upper
freq
uen
cylim
itofe6kH
z
Pro
:-Presen
tdueto
saturation
with
grea
terstim
ulusleve
lsCo
n:-
Nomechan
ical
correlate
HairBundle
Mechan
ics
Pro
:-Fo
rcege
nerationallowsam
plifi
cation
ofinco
mingsign
alCo
n:-
Limited
forcege
neration,force
perpen
dicular
toba
silarmem
bran
emotion
Pro
:-Can
theo
retically
tuneupto
100kH
z-TM
allowsco
ncerted
effort
ofOHCsan
dresu
ltsin
sharper
tuning.
Con:-
Highfreq
uen
cyoscilla
tion
shav
enot
been
show
ndirectly
Pro
:-Directob
servationshav
ebe
enmad
eCo
n:-
Limited
operatingrange
Somatic
Motility
Pro
:-Amplifysign
alsupto
atleast79
kHzas
motion
isseen
atthesehighfreq
uen
cies
-Largeforces
canbe
generated
-Fo
rcein
series
withba
silarmem
bran
emotion
Con:-
Amplifi
cation
follo
wsmem
bran
epoten
tial,thereforeis
limited
toab
out1kH
zby
themem
bran
etimeco
nstan
t
Pro
:-Th
eoretically
canop
erateat
highfreq
uen
cyCo
n:-
Nokn
ownfreq
uen
cytuningor
tonotop
icva
riations
-Limited
bymem
bran
etimeco
nstan
t
Pro
:Co
n:-
Nomechan
ism
forge
nerationof
compressivenon
linea
rity
know
n
A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 115
convergence in mechanism (Stewart and Hudspeth, 2000). To date,no somatically motile cells have been identified in non-mammalianhearing organs, leaving active hair bundle mechanics as the likelysource of amplification (He et al., 2003). Alone, a negative dataargument is weak; fortunately several elegant experiments per-formed in lizard clearly localize the site of the active process, themechanism underlying otoacoustic emissions, to the stereociliaryhair bundles and further demonstrate that the mechanism iscalcium dependent (Manley and Koppl, 2008; Manley et al., 2001).By utilizing a lizard organ where hair bundles are oriented 180�
from each other, investigators were able to modulate electricallyinduced emissions with low frequency sound in a manner consis-tent with generation being localized to the sensory hair bundle.Additionally, modeling data suggests that the hair bundles playa larger role in cochlear amplification in birds thanmammals due todifferences in the morphology of the basilar papilla compared tothe mammalian organ of Corti (Sul and Iwasa, 2009). Theseprovocative data clearly point to the hair bundle as a potential siteof cochlear amplification.
Data suggest the hair bundle has the attributes required forcochlear amplification. Perhaps the simplest evidence, albeitcircumstantial, for the hair bundle involvement in cochlearamplification is data demonstrating tonotopic variations inmechanotransduction properties. In most species with tectorialmembranes, hair bundles vary in height and number of stereociliasuch that high frequency bundles are shorter and have more ster-eocilia than low frequency hair bundles, thus passively preparingthem to respond best to their frequency range (Hackney et al., 1993;Koppl and Authier, 1995; Tilney and Saunders, 1983). Whereinvestigated, the size of the mechanotransducer current variestonotopically, being larger in high frequency cells (He et al., 2004;Kennedy et al., 2003; Ricci and Fettiplace, 1997; Ricci et al., 2005;Waguespack et al., 2007). The increase in current amplitude is inpart due to a larger single channel conductance but also due to theincreased number of channels associated with the larger number ofstereocilia (Beurg et al., 2006; Ricci et al., 2003). Activation kinetics,where measurements are technically feasible, also show a tono-topic gradient as do adaptation kinetics, both correlating well withcharacteristic frequency of the afferent fiber (Ricci et al., 2005).Minimally, it may be argued that activation and adaptation canprovide tuning to the hair bundle. Based on measured properties,the tuning provided by a single sensory hair bundle is relativelybroad and inconsistent with the sharp tuning observed in afferentfibers (Ricci et al., 2005). One problemwith direct interpretation ofthese measurements is that they are made under non-physiologicalconditions in terms of temperature, endolymph composition, andendocochlear potential, so specific features related to tuning maynot be directly interpretable. A second problem with direct inter-pretation of these measurements is that the bundles are unloaded,since the tectorial membrane is removed. From modeling data, themass loaded onto the hair bundle can create sharper tuning of thehair bundle resonance (Dierkes et al., 2008; Nam and Fettiplace,2008). Data from the lizard also support the argument that tecto-rial membrane attachments can enhance hair bundle tuningproperties (Authier and Manley, 1995; Manley et al., 1988). Addi-tional measurements of hair bundle interactions with the tectorialmembrane are needed to separate hair bundle mechanics fromtectorial membrane resonance properties in order to quantify therole of the hair bundle in frequency selectivity.
Hair bundles may provide considerably more to cochlearamplification than only frequency selectivity. Distortion productotoacoustic emissions (DPOAEs) are produced by the cochlea inresponse to two primary frequency inputs that generate additionalfrequency outputs often of the form (2f1ef2) or (f2ef1) and areindicative of a functioning active process. DPOAEs were identified

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122116
in individual hair bundles of frog sacculus and are thought toemerge from nonlinearities in the mechanotransduction process(Jaramillo and Hudspeth, 1993). SOAEs are present in a variety ofnon-mammalian species and argue for the existence of a mechan-ical amplifier (Koppl and Manley, 1993; Manley and Gallo, 1997;Manley et al., 1996; Taschenberger and Manley, 1997). Sponta-neous hair bundle oscillations, a possible correlate to SOAEs albeitat much lower frequencies, have been identified in frog sacculusand turtle auditory hair bundles, and both the hair bundle oscilla-tions and the SOAEs show similar sensitivities to pharmacologicalmanipulations (Ricci et al., 1998; Stewart and Hudspeth, 2000). Hairbundles can amplify mechanical stimulations (Martin andHudspeth, 1999) and can use mechanical noise to enhance sensi-tivity (Indresano et al., 2003; Jaramillo and Wiesenfeld, 1998). Hairbundles show compressive nonlinearity about the hair bundlesoptimal stimulation frequency (Martin and Hudspeth, 2001). Hairbundles can generate force that may feedback into the basilarmembrane (Benser et al., 1996; Beurg et al., 2008; Howard andHudspeth, 1988; Jaramillo and Hudspeth, 1993; Kennedy et al.,2006; Ricci et al., 2000, 2002). However the amount of forcea hair bundle can generate is typically small, considerably smallerthan that generated by outer hair cell somatic motility. The forcegeneration is limited, partly due to the sparse number of mecha-notransducer channels present. More recent work in rat outer haircells, however, indicates that these bundles can generate consid-erably more force than previously measured in the frog or turtle(Kennedy et al., 2005). Questions however have arisen aboutwhether somatic motility played a role in themeasured hair bundlemovements (Beurg et al., 2008; Jia and He, 2005; Kennedy et al.,2006). Thus issues remain as to whether hair bundles cangenerate enough force to drive basilar membrane movementswhen mechanically loaded by the tectorial membrane. Proof ofprinciple experiments in an in vitro gerbil preparation demonstratecalcium dependent movements, which are probably driven by theconcerted efforts of active hair bundles (Chan and Hudspeth, 2005).An interesting experiment would be to probe hair bundlemechanics in the prestin mutant that no longer is motile as ameansof quantifying the contribution of the hair bundle in the mamma-lian system (Dallos et al., 2008).
An additional issue regarding hair bundle mechanics underlyingthe active process is that the proposed mechanism for hair bundleforce generation may not be fast enough. The general idea is thata region of mechanical instability is created (a Hopf bifurcation)that causes hair bundles to oscillate between energy minima(Hudspeth, 2008; Martin et al., 2000, 2003). Data supporting thishypothesis are strong. The factors suggested to be involved increating the bifurcation are mechanotransduction channel gatingcompliance interacting with adaptation mechanisms (Hudspeth,2005; Hudspeth et al., 2000). At present myosin 1c is implicatedas underlying the mechanisms of slow and possibly fast adaptation(Gillespie and Cyr, 2004; Holt et al., 2002; Stauffer et al., 2005).Given the kinetic properties of myosin 1c, it seems unlikely thathigh frequency responses could be generated. Similarly, a calcium-driven mechanism might not be expected to work up to 100 kHz,particularly if the calcium needs to diffuse away from the mecha-notransduction channels to a distant binding site 10e100 s of nmaway (Beurg et al., 2009; Ricci et al., 1998). Recent arguments thatmyosin 1c underlies fast adaptation, a mechanism thought toinvolve myosin’s interaction with calcium causing a tilting but notuncoupling from the actin seems in opposition to a role in ampli-fication because the predicted movement and force generation arequite small (probably perpendicular to the basilar membranemotion) and unlikely to feedback to basilar membrane motion, sospeed is gained at the cost of power and dynamic range (Staufferet al., 2005). A better understanding of the molecular
mechanisms of adaptation will give insight into the limitations andadvantages of its application to cochlear amplification.
Despite the underlying issues with mechanism, directmeasurements of hair bundle and mechanotransducer propertiessupport the argument that hair bundles can operate at highfrequencies. Activation kinetics of mechanotransduction are quitefast, currently faster than can be stimulated (rise times in the tensof microseconds), therefore, quite possibly capable of operatingcycle by cycle at very high frequencies (Ricci et al., 2005). Fastertechnology is needed to clearly determine the kinetic limits ofgating compliance. Adaptation rates in mammal are also quite fast,tens of microseconds for fast adaptation and milliseconds for theslow component, and these values are probably underestimates ofthe upper limits because adaptation rates are slowed by stimulusrise times (Kennedy et al., 2003; Ricci et al., 2005; Wu et al., 1999).Thus it appears that the kinetics of the measured processes arefaster than the hypothesized molecular mechanism might support,but certainly in the appropriate range to serve as a component ofthe cochlear amplifier.
Earlier arguments demonstrated that limited thermal energy atthreshold requires signal filtering and amplification. This argumentholds regardless of the final function of the hair cell or thefrequency range over which the hair cell responds. Hair bundleresponse properties are remarkably similar across hair cell types(see caveat regarding outer hair cells below) and end organs andcan provide the amplification and filtering needed for signaldetection. We hypothesize that this intrinsic mechanism is main-tained throughout evolution and that further specializations haveoccurred in some species permitting higher frequency hearing anda separation of function into hair cells that transduce informationcentrally and those that amplify and filter signals at the periphery.
7. Two hair cell system
The simplest auditory end organs, like those in turtles and frogshave single hair cell types; hair cells receive both efferent andafferent innervation and hair cell properties vary in a tonotopicallydefined manner (Crawford and Fettiplace, 1980; Sneary, 1988a,1988b). In general, the hair cells are all doing the same thing, justat different frequencies. Birds, although still in a papilla, have twohair cells types based on both morphology and innervations. Shorthair cells have broad apical surfaces and cuticular plate regions andreceive efferent innervation whereas the tall hair cells receive bothafferent and efferent innervations (Fischer, 1994). Recently a similarpattern was identified in a lizard (Chiappe et al., 2007). The func-tional significance of these two hair cell systems remains to beidentified as no clear experimental paradigm has been able toseparate functions between cell types. In the mammalian cochleathere is also a two hair cell system. A variety of differences havebeen identified between outer hair cells (OHCs) and inner hair cells(IHCs): (a) OHCs vary tonotopically in their mechanotransductionproperties while IHCs do not (Beurg et al., 2006), (b) OHCs have hairbundles embedded in the tectorial membrane, IHC bundles are freestanding (Lim,1980), (c) OHCs showa tonotopic gradient in calciumbuffering while IHCs do not (Hackney et al., 2005), (d) OHCs havehigh levels of Ca-ATPases in the stereocilia while IHCs do not (Gratiet al., 2006) (e) OHCs receive the majority of efferent innervations(Warr, 1992; Warr et al., 1997) while IHCs receive the majority ofafferent innervations (Ehret, 1979; Morrison et al., 1975), and (f)OHCs show electromotility while IHCs do not (Ashmore, 2008). Inaddition, in mammalian systems, significant data supports theargument that OHCs are required for cochlear amplification.Selective damage to OHCs either by noise or toxin exposureelevates thresholds and reduces frequency selectivity (Ashmore,2008). Although not conclusive, given that changes in basilar

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 117
membrane mechanics are inevitable with OHC loss, OHCs certainlyare strongly implicated as being involved in the generation of thecochlear amplifier. It is unclear at this point whether other two haircell systems also assign cochlear amplification to one hair cell typethough it seems plausible given the innervation patterns observed.
8. Somatic motility as the cochlear amplifier
Somatic motility was first described in the guinea pig (Brownellet al., 1985), and extended to all other mammalian species testedincluding human (Oghalai et al., 1998). In response to membranevoltage changes, OHCs change length (Brownell et al., 1985).Evidence suggest the movement is voltage, not current driven(Ashmore, 1987; Santos-Sacchi and Dilger, 1988). A novel mecha-nism of force generation was identified involving a membraneprotein that changes conformation in response to voltage fluctua-tions (Zheng et al., 2000). The conformational change results in analtered membrane surface area leading to OHC length changes(Dallos et al., 1993; Iwasa, 1994). OHC length changes are in serieswith the basilar membrane and can augment or diminish soundevoked movements of this membrane depending on phase. Aconcomitant change in membrane capacitance was identified,similar to gating charge for ion channels and is a signature for themotility mechanism (Santos-Sacchi, 1991). The observation ofsomatic motility in OHCs served to focus the field on the potentialof this process to underlie cochlear amplification. The identificationof prestin, a member of the SLC26 family of anion-bicarbonatetransporter proteins has allowed for a detailed analysis of themechanisms of prestin action and also for a more direct investi-gation of the role of OHC motility in generating the active process.In this light, investigations of the functional significance of prestinare far ahead of those regarding hair bundle mechanics, where solittle is known regarding molecular mechanisms.
Does somatic motility have the requisite properties of frequencyselectivity, compressive nonlinearity and amplification that havebeen ascribed to cochlear amplification? To date, there is no datasupporting the conclusion that OHC motility is tuned. There are noknown tonotopic gradients in prestin levels or intrinsic propertiesof somatic motility that would make motility frequency selective.OHC motility can operate at very high frequencies under non-physiological conditions (Frank et al., 1999); however, given that itis voltage dependent and limited by the membrane time constantunder physiological conditions, motility will not operate on a cycleby cycle basis much beyond 1 kHz. That it can physically work atmuch higher frequencies has led to several suggested means bywhich the membrane time constant can be circumvented (seeAshmore, 2008; Hudspeth, 2008, and Dallos, 2008). Possibilitiessuch as chloride modulation via a stretch activated channel(Rybalchenko and Santos-Sacchi, 2003), extracellular current flowmodulating prestin (Dallos and Evans, 1995) or gradients inintrinsic electrical properties of OHC all have merit, but none havebeen adequately tested in order to be accepted or refuted.Regardless of whether these clever possibilities hold water, theywould simply allow the motility process to operate at highfrequencies but would not provide frequency selectivity. Given thatmotility is a voltage-dependent phenomenon, one might posit thattuning would be provided by the electrical properties of the haircell basolateral membrane, but as mentioned earlier no evidenceexists for such a tuning mechanism. Compressive nonlinearity isa hallmark of the active process, yet here too, data does not supportthe argument that compressive nonlinearity is a property of OHCmotility. The voltage dependence of motility folIows a Boltzmannfunction. It might be possible that variations in basolateral elec-trical or mechanotransducer channel properties of the OHCs mightimpose a compressive nonlinearity onto the OHC receptor potential
and that this could feedback onto motility, but even here data islacking. Finally, amplification is a critical component of the activeprocess. Here OHCs excel, generating large forces and movementsin series with basilar membrane motion, which in turn directlyalters hair bundle motion. Forces are considerably larger than thosemeasured thus far from hair bundle active mechanisms.
9. Unifying theory
The above arguments might imply that OHC motility is ill suitedfor providing cochlear amplification; however it seems unlikelythat these mammalian specializations are for naught. Perhaps thedifficulty is more with the assumption that a single cell or a singleprocess need account for the entire system’s abilities. Thisassumption stems from early work where tuning and amplificationappear to be inherent to the sensory cell (see electrical resonancesection). However as the complexity of the inner ear organincreased, processing appears to be less inherent to individual haircells and more delegated to the organ. This seems to be the caseparticularly with the two hair cell system. Cells with no afferentinnervations must be serving some type of feedback role, therebymodulating the output of the primary sensory cell. When consid-ering the mammalian cochlea it seems more reasonable to assumethat both hair bundle mechanics and somatic motility are impor-tant for the active process but perhaps in somewhat different ways.Consider that if amplification properties are not present in the freestanding mammalian IHC bundles, the work required of OHCs tolower threshold and provide frequency selectivity would be evengreater. Similarly though, IHC hair bundle mechanics play little rolein generating cochlear amplification as they are free standingbundles and the preponderance of data support the argument thatthe cochlear amplifier resides in the OHC (Ashmore, 2008). Onepossibility is that hair bundle mechanics can serve a specificintrinsic amplification process for a given bundle while OHCmotility serves as a gain enhancement to the system. A simplescenario might be that hair bundle mechanics provide the tuningand compressive nonlinearity associated with the active processand that these features are directly mechanically amplified by OHCmotility. In this sense OHC motility can provide a cycle by cycle(assuming the membrane time constant issue is circumvented)amplification to a hair bundle process that has the required tuningand compressive properties but is more limited in its force gener-ating abilities, particularly at high frequencies (Fig. 4). It also allowsfor all hair cells, not just OHCs, to possess an amplification mech-anism. Indirect support of this possibility is the observation thatOHC mechanotransduction has a limited slow component ofadaptation. The slow component of adaptation is thought toprovide the motor for hair bundle amplification, so perhaps theslower adaptationmechanism is replaced by somaticmotility as theengine powering the process, alleviating limitations like calciumclearance while operating at high frequencies. This division of laborwould provide an excellent means of limiting thermal noiseamplification associated with Brownian motion of the hair bundle.If this possibility were true, it would be extremely difficult toseparate between these two potential mechanisms becauseperturbation of one feeds back onto the other. Data from prestinknockout animals indirectly supports this hypothesis by demon-strating that distortion product otoacoustic emissions are presentdespite a loss of OHC motility (Liberman et al., 2004). Theremaining DPOAEs were reduced as would be expected if thecomponent providing gain were removed but also demonstratingthe involvement of another mechanism, aside from OHCmotility inDPOAE generation.
One problem, quite difficult to surmount, is identifying causallinks between possible mechanisms of cochlear amplification.

Fig. 4. Schematic representation of the unifying theory of cochlear amplification. (A) In some vertebrates, like turtle basilar papilla shown here, intrinsic mechanisms within thehair cell are enough to produce sharp tuning and amplification without basilar membrane tuning. Iso-intensity responses of the hair cell (blue) and basilar membrane (red) areschematized for 3 different intensity levels. Red arrows indicate direction of sound stimulation. (B) Hair cells of the mammalian cochlea alone are not able to feedback onto thebasilar membrane, however the hair bundle has some intrinsic tuning properties providing it with compressive non-linearily and some tuning as compared to corresponding basilarmembrane motion. (C) The introduction of a tectorial membrane further sharpens tuning with some amplification provided by the concerted effort of multiple hair bundles that canfeedback onto basilar membrane. (D) The addition of somatic motility provides further gain to the system, which is fed back onto the basilar membrane. The gain is necessary tobring the motion above the level of the noise (cross-hatched area) in the system to make the stimulus detectable.
A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122118
Given a system where many signal processing events occur inseries, alteration of any given step can lead to loss of function of thewhole system, making it unclear whether the alteration left theother signal processing events intact. For example several cleverprestin mutations, along with prestin knockouts have been used toargue that somatic motility can completely account for cochlearamplification. The knockout data are difficult to interpret based onchanges in basilar membrane mechanics and on the cell deathinduced by the knockout (Cheatham et al., 2004; Liberman et al.,2002; Wu et al., 2004). However mutations that alter or removethe voltage dependent motility also lead to profound hearing lossimplicating a critical role for somatic motility (Dallos et al., 2008).These provocative data certainly further implicate somatic motilityas being an important component of the active process, but they donot causally demonstrate that motility alone is responsible foramplification. The above hypothesis, incorporating both hair
bundle and somatic motility would result in similar findings tothose reported because the hair bundle alone would not generateenough force to drive basilar membrane motion or the hair bundleposition may be moved away from its most sensitive locationreducing force generation by the bundle. It is important to recog-nize that the ability of the hair bundle to amplify is criticallydependent on the resting position of the hair bundle; it must bepoised at the Hopf bifurcation to generate requisite tuning and gain.Any perturbation of this position either directly or indirectly willreduce the ability of the hair bundle to provide filtering andamplification.
Tecta mice where the OHC bundles are no longer tethered to thetectorial membrane are deaf (Legan et al., 2000). It has been arguedthat the existence of electrically evoked emissions in these animalsdemonstrate that somatic motility is completely responsible for theactive process because hair bundle movements would no longer be

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 119
capable of moving the basilar membrane (Mellado Lagarde et al.,2008). Are electrically evoked emissions a complete representa-tion of the active process or perhaps simply a means of probing anelectrical component of mechanical amplification? The resultsobtained in these experiments could be accounted for by the abovehypothesis that incorporates both mechanisms of amplification,losing the hair bundle contribution by uncoupling the tectorialmembrane results in loss of mechanically induced cochlearamplification by bypassing the hair bundle while electrical stimu-lation activates somatic motility directly, bypassing the need formechanosensitivity. So again the likely interaction between themodes of amplification make interpretation of data meant toimpact only one mechanism more elusive.
It would seem that both hair bundle mechanics and somaticmotility are important in establishing the cochlear amplifier in themammalian cochlea. Each has advantages and disadvantages (Table2). The significance of either mechanism is not diminished by thecontribution of the other, and many important questions remain tobe addressed. For example how do these two processes interact andfeedback on each other? Being able to manipulate hair bundleproperties at the molecular level will provide critical tools neededto address this question. How is the output of the OHC translated tothe IHC, and more generally, in a two hair cell system, how do thecells interact?
In non-mammalian vertebrates, like birds and some lizards,where the cochlear amplifier appears to reside in the hair bundle,the most likely source for interaction is the tectorial membrane.Tectorial membrane mechanical properties have been investigatedacross a variety of species, and in all cases including lizards, birds,and mammals, gradients in mechanical properties were identifiedand indicative of a role in frequency tuning (Koppl and Authier,1995; Richter et al., 2007; Weaver and Schweitzer, 1994). Geneti-cally modified tectorial membranes can increase frequency tuning(Russell et al., 2007). Tectorial membranes are also reported ashaving resonant properties (Gummer et al., 1996). Theoreticalanalysis support the contention that loading the hair bundle cansharpen frequency tuning and also that the properties of the tec-torial membrane can serve to coordinate hair bundle motion andforce generation (Dierkes et al., 2008; Nam and Fettiplace, 2008).Further investigations into the tuning and coupling properties ofthe tectorial membrane are needed as these properties are prob-ably critically important to our understanding of amplificationmechanisms. The properties of the tectorial membrane are possiblycritical for the establishment of the cochlear amplifier by sharp-ening hair bundle tuning. Together, hair bundle mechanics, tecto-rial membrane properties, and outer hair cell motility probablycombine to generate the mature mammalian cochlear amplifier.
10. Advantages of a two hair cell system
It has been argued that OHCs serve to extend the frequencyrange of the cochlea, but many examples of high frequency hearingexist in systems without OHCs, for example, ultrasound can bedetected in a variety of frogs and many insects have high frequencyhearing (Arch et al., 2008, 2009; Farris and Hoy, 2000; Faure andHoy, 2000; Feng et al., 2006; Shen et al., 2008). You might arguethat the cochlea is over-engineered for human hearing since ourmost utilized frequency range (for speech and localization) iscomparably sensed bymany species that do not have OHCs. Despitethe numerous differences in end organ structures, frequencyselectivity and thresholds are remarkably similar between species.Perhaps the differences employed in the mammal are not simply toovercome physical limitations but to serve some higher orderfunction. The increase in parameter numbers, for example, two haircells, motility, hair bundle mechanics, and a separation of
innervations patterns, may provide more sites for regulation.Perhaps finer central control allows better selectivity of incomingsignals.
The two hair cell systemmay allow for the functional separationof amplification and information transfer, which may result froma conflict arising at high frequencies between maximizing bothmechanical amplification and voltage response (Chiappe et al.,2007). One population of cells may maintain a low resting proba-bility of opening, 0.1e0.2, in order to maximize the voltage changeduring high frequency sine wave stimulation (like IHCs); whereasa second population of hair cells responsible for amplificationmaintain a much higher resting open probability, closer to 0.5 inorder tomaximize sensitivity (like OHCs). This is an interesting ideathat is perhaps supported by the high concentrations of Ca-ATPasesin OHC bundles as compared to IHC hair bundles (Dumont et al.,2001; Grati et al., 2006). This hypothesis is intriguing in that it istestable across all two hair cell end organs. It would be interesting,for example, to see if short avian or the noninnervated lizard haircells have higher levels of Ca-ATPases in their stereocilia whencompared to tall hair cells or innervated lizard hair cells. A corre-sponding change with the advent of the two hair cell system is thatone population receives efferent innervations, probably theamplifying cells as in mammals, but not yet confirmed in othervertebrates.
In non-mammalian two hair cell systems, the efferent systemmay serve to detune and attenuate selective frequency rangeswithout altering spontaneous activity driven by the sensory cell. Insingle hair cell systems, efferent stimulation will reduce sponta-neous activity in the nerve fiber while also detuning and attenu-ating. In the mammal it is possible that an additional level ofregulation is available, where not only can spontaneous activity bemaintained, but also amplification may be separately regulatedfrom tuning, as hair bundle mechanics and somatic motility may beregulated separately. Separate regulation may be possible forexample if second messenger levels of cyclic AMP are altered viaefferent stimulation. Cyclic AMP can modulate the set point of thehair bundle with no known effects on somatic motility (Ricci andFettiplace, 1997). The fact that IHC bundles are not directlycoupled to the tectorial membrane supports the hypothesis that theresting position of the IHC bundle is unaffected by efferent stimu-lation to OHCs.
11. Conclusions
The physical limitations associated with audition have beenovercome throughout evolution in a variety if ways. Severalmechanisms have arisen to establish the sensitivity and frequencyranges observed. The presence of frequency discrimination,compressive nonlinearity, amplification, and otoacoustic emissionsacross these disparate hearing organs support the requirement ofa common cochlear amplification mechanism to establish lowthresholds and sharp tuning. Two mechanisms are proposed to beinvolved in generating cochlear amplification. Hair bundle activemechanics driven by channel gating and adaptation propertiesappear to be common across species, while in mammalian haircells, somatic motility is thought to play a critical role. A newhypothesis is presented that suggests somatic motility interactswith hair bundle motility to provide cycle by cycle cochlearamplification. Hair bundle mechanics can provide frequencyselectivity and compressive nonlinearity while somatic motility canprovide amplification. Interactions with the tectorial membrane areposited to shape frequency response. It is proposed that the sepa-ration of amplification and tuning within hair cells and betweenhair cells provides multiple sites of regulation and control of inputs.Given the similarity in response properties across diverse species it

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122120
might be that the specializations inmammalian hearing serve somehigher order functions.
Acknowledgements
Thanks to Ham Farris for discussions on insect hearing andhigher order functions. Thanks to Geoff Manley for his expertise inthe details of the comparative aspects of hearing. This work wassupported by NIDCD funding to AJR, RO1DC003896.
References
Ahmed, Z.M., Goodyear, R., Riazuddin, S., Lagziel, A., Legan, P.K., Behra, M.,Burgess, S.M., Lilley, K.S., Wilcox, E.R., Griffith, A.J., Frolenkov, G.I.,Belyantseva, I.A., Richardson, G.P., Friedman, T.B., 2006. The tip-link antigen,a protein associated with the transduction complex of sensory hair cells, isprotocadherin-15. J. Neurosci. 26, 7022e7034.
Arch, V.S., Grafe, T.U., Narins, P.M., 2008. Ultrasonic signalling by a Bornean frog.Biol. Lett. 4, 19e22.
Arch, V.S., Grafe, T.U., Gridi-Papp, M., Narins, P.M., 2009. Pure ultrasonic commu-nication in an endemic Bornean frog. PLoS One 4, e5413.
Art, J.J., Fettiplace, R., 1987. Variation of membrane properties in hair cells isolatedfrom the turtle cochlea. J. Physiol. (London) 385, 207e242.
Ashmore, J.F., 1987. A fast motile response in guinea-pig outer hair cells: the cellularbasis of the cochlear amplifier. J. Physiol. (London) 388, 323e347.
Ashmore, J., 2008. Cochlear outer hair cell motility. Physiol. Rev. 88, 173e210.Authier, S., Manley, G.A., 1995. A model of frequency tuning in the basilar papilla of
the Tokay gecko, Gekko gecko. Hear. Res. 82, 1e13.Benser, M.E., Marquis, R.E., Hudspeth, A.J., 1996. Rapid, active hair bundle move-
ments in hair cells from the bullfrog’s sacculus. J. Neurosci. 16, 5629e5643.Beurg, M., Evans, M.G., Hackney, C.M., Fettiplace, R., 2006. A large-conductance
calcium-selective mechanotransducer channel in mammalian cochlear haircells. J. Neurosci. 26, 10992e11000.
Beurg, M., Nam, J.H., Crawford, A., Fettiplace, R., 2008. The actions of calcium on hairbundle mechanics in mammalian cochlear hair cells. Biophys. J. 94, 2639e2653.
Beurg, M., Fettiplace, R., Nam, J.H., Ricci, A.J., 2009. Localization of inner hair cellmechanotransducer channels using high-speed calcium imaging. Nat. Neurosci.12, 553e558.
Bialek, W., 1987. Physical limits to sensation and perception. Annu. Rev. Biophys.Biophys. Chem. 16, 455e478.
Bialek, W., Schweitzer, A., 1985. Quantum noise and the threshold of hearing. Phys.Rev. Lett. 54, 725e728.
Bozovic, D., Hudspeth, A.J., 2003. Hair-bundle movements elicited by transepithelialelectrical stimulation of hair cells in the sacculus of the bullfrog. Proc. Natl.Acad. Sci. U.S.A. 100, 958e963.
Brownell, W.E., 1990. Outer hair cell electromotility and otoacoustic emissions. Ear.Hear. 11, 82e92.
Brownell, W.E., Bader, C.R., Bertrand, D., de Ribaupierre, Y., 1985. Evoked mechanicalresponses of isolated cochlear outer hair cells. Science 227, 194e196.
Chan, D.K., Hudspeth, A.J., 2005. Ca2þ current-driven nonlinear amplification bythe mammalian cochlea in vitro. Nat. Neurosci. 8, 149e155.
Cheatham, M.A., Huynh, K.H., Gao, J., Zuo, J., Dallos, P., 2004. Cochlear function inPrestin knockout mice. J. Physiol. 560, 821e830.
Chiappe, M.E., Kozlov, A.S., Hudspeth, A.J., 2007. The structural and functionaldifferentiation of hair cells in a lizard’s basilar papilla suggests an operationalprinciple of amniote cochleas. J. Neurosci. 27, 11978e11985.
Crawford, A.C., Evans, M.G., Fettiplace, R., 1989. Activation and adaptation oftransducer currents in turtle hair cells. J. Physiol. (London) 419, 405e434.
Crawford, A.C., Fettiplace, R., 1978. Ringing responses in cochlear hair cells of theturtle [proceedings]. J. Physiol. (London) 284, 120Pe122P.
Crawford, A.C., Fettiplace, R., 1980. The frequency selectivity of auditory nerve fibresand hair cells in the cochlea of the turtle. J. Physiol. (London) 306, 79e125.
Crawford, A.C., Fettiplace, R., 1981. An electrical tuning mechanism in turtlecochlear hair cells. J. Physiol. (London) 312, 377e412.
Crawford, A.C., Fettiplace, R., 1985. The mechanical properties of ciliary bundles ofturtle cochlear hair cells. J. Physiol. (London) 364, 359e379.
Dallos, P., 1985. Response characteristics of mammalian cochlear hair cells. J. Neu-rosci. 5, 1591e1608.
Dallos, P., 2008. Cochlear amplification, outer hair cells and prestin. Curr. Opin.Neurobiol. 18, 370e376.
Dallos, P., Hallworth, R., Evans, B.N., 1993. Theory of electrically driven shapechanges of cochlear outer hair cells. J. Neurophysiol. 70, 299e323.
Dallos, P., Wu, X., Cheatham, M.A., Gao, J., Zheng, J., Anderson, C.T., Jia, S., Wang, X.,Cheng, W.H., Sengupta, S., He, D.Z., Zuo, J., 2008. Prestin-based outer hair cellmotility is necessary for mammalian cochlear amplification. Neuron 58,333e339.
Dallos, P., Evans, B.N., 1995. High-frequency motility of outer hair cells and thecochlear amplifier. Science 267, 2006e2009.
Davis, R.I., Ahroon, W.A., Hamernik, R.P., 1989. The relation among hearing loss,sensory cell loss and tuning characteristics in the chinchilla. Hear. Res. 41, 1e14.
Dierkes, K., Lindner, B., Julicher, F., 2008. Enhancement of sensitivity gain andfrequency tuning by coupling of active hair bundles. Proc. Natl. Acad. Sci. U.S.A.105, 18669e18674.
van Dijk, P., Wit, H.P., Segenhout, J.M., 1989. Spontaneous otoacoustic emissions inthe European edible frog (Rana esculenta): spectral details and temperaturedependence. Hear. Res. 42, 273e282.
Dumont, R.A., Lins, U., Filoteo, A.G., Penniston, J.T., Kachar, B., Gillespie, P.G., 2001.Plasma membrane Ca2þ-ATPase isoform 2a is the PMCA of hair bundles. J.Neurosci. 21, 5066e5078.
Eatock, R.A., 2000. Adaptation in hair cells. Annu. Rev. Neurosci. 23, 285e314.Eatock, R.A., Corey, D.P., Hudspeth, A.J., 1987. Adaptation of mechanoelectrical
transduction in hair cells of the bullfrog’s sacculus. J. Neurosci. 7, 2821e2836.Eatock, R.A., Saeki, M., Hutzler, M.J., 1993. Electrical resonance of isolated hair cells
does not account for acoustic tuning in the free-standing region of the alligatorlizard’s cochlea. J. Neurosci. 13, 1767e1783.
Ehret, G., 1979. Quantitative analysis of nerve fibre densities in the cochlea of thehouse mouse (Mus musculus). J. Comp. Neurol. 183, 73e88.
Farris, H.E., Wells, G.B., Ricci, A.J., 2006. Steady-state adaptation of mechano-transduction modulates the resting potential of auditory hair cells, providing anassay for endolymph [Ca2þ]. J. Neurosci. 26, 12526e12536.
Farris, H.E., Hoy, R.R., 2000. Ultrasound sensitivity in the cricket, Eunemobius car-olinus (Gryllidae, Nemobiinae). J. Acoust. Soc. Am. 107, 1727e1736.
Faure, P.A., Hoy, R.R., 2000. Neuroethology of the katydid T-cell. I. Tuning andresponses to pure tones. J. Exp. Biol. 203, 3225e3242.
Fay, R.R., 1988. Hearing in Vertebrates: a Psychophysics Databook. Hill-Fay Associ-ates, Winnetka, Ill.
Feng, A.S., Narins, P.M., Xu, C.H., Lin, W.Y., Yu, Z.L., Qiu, Q., Xu, Z.M., Shen, J.X., 2006.Ultrasonic communication in frogs. Nature 440, 333e336.
Fettiplace, R., 2006. Active hair bundle movements in auditory hair cells. J. Physiol..Fettiplace, R., Crawford, A.C., 1978. The coding of sound pressure and frequency in
cochlear hair cells of the terrapin. Proc. R. Soc. Lond. B. Biol. Sci. 203, 209e218.Fettiplace, R., Ricci, A.J., Hackney, C.M., 2001. Clues to the cochlear amplifier from
the turtle ear. Trends. Neurosci. 24, 169e175.Fettiplace, R., Fuchs, P.A., 1999. Mechanisms of hair cell tuning. Annu. Rev. Physiol.
61, 809e834.Fettiplace, R., Hackney, C.M., 2006. The sensory and motor roles of auditory hair
cells. Nat. Rev. Neurosci. 7, 19e29.Fischer, F.P., 1994. Quantitative TEM analysis of the barn owl basilar papilla. Hear.
Res. 73, 1e15.Fischer, F.P., 1998. Hair cell morphology and innervation in the basilar papilla of the
emu (Dromaius novaehollandiae). Hear. Res. 121, 112e124.Fischer, F.P., Koppl, C., Manley, G.A., 1988. The basilar papilla of the barn owl Tyto
alba: a quantitative morphological SEM analysis. Hear. Res. 34, 87e101.Frank, G., Hemmert, W., Gummer, A.W., 1999. Limiting dynamics of high-frequency
electromechanical transduction of outer hair cells. Proc. Natl. Acad. Sci. U.S.A.96, 4420e4425.
Fuchs, P.A., Nagai, T., Evans, M.G., 1988. Electrical tuning in hair cells isolated fromthe chick cochlea. J. Neurosci. 8, 2460e2467.
Fuchs, P.A., Evans, M.G., 1988. Voltage oscillations and ionic conductances in haircells isolated from the alligator cochlea. J. Comp. Physiol. A 164, 151e163.
Gillespie, P.G., Cyr, J.L., 2004. Myosin-1c, the hair cell’s adaptation motor. Annu. Rev.Physiol. 66, 521e545.
Gleich, O., Manley, G.A., 1988. Quantitative morphological analysis of the sensoryepithelium of the starling and pigeon basilar papilla. Hear. Res. 34, 69e85.
Gleich, O., Manley, G.A., 2000. The hearing organs of birds and crocodilia. In:Dooling, R.J., Fay, R.R., Popper, A.N. (Eds.), Comparative Hearing. Birds andReptiles. Springer, New York, pp. 70e138.
Gopfert, M.C., Robert, D., 2003. Motion generation by Drosophila mechanosensoryneurons. Proc. Natl. Acad. Sci. U.S.A. 100, 5514e5519.
Grati, M., Schneider, M.E., Lipkow, K., Strehler, E.E., Wenthold, R.J., Kachar, B., 2006.Rapid turnover of stereocilia membrane proteins: evidence from the traffickingand mobility of plasma membrane Ca(2þ)-ATPase 2. J. Neurosci. 26,6386e6395.
Gummer, a.W., Smolders, J.W., Klinke, R., 1987. Basilar membrane motion in thepigeon measured with the Mössbauer technique. Hear. Res. 29, 63e92.
Gummer, A.W., Hemmert, W., Zenner, H.P., 1996. Resonant tectorial membranemotion in the inner ear: its crucial role in frequency tuning. Proc. Natl. Acad. Sci.U.S.A. 93, 8727e8732.
Hackney, C.M., Fettiplace, R., Furness, D.N., 1993. The functional morphology ofstereociliary bundles on turtle cochlear hair cells. Hear. Rea. 69, 163e175.
Hackney, C.M., Mahendrasingam, S., Penn, A., Fettiplace, R., 2005. The concentra-tions of calcium buffering proteins in mammalian cochlear hair cells. J. Neu-rosci. 25, 7867e7875.
Harris, G.G., 1968. Brownian motion in the cochlear partition. J. Acoust. Soc. Am. 44,176e186.
He, D.Z., Beisel, K.W., Chen, L., Ding, D.L., Jia, S., Fritzsch, B., Salvi, R., 2003. Chick haircells do not exhibit voltage-dependent somatic motility. J. Physiol. 546, 511e520.
He, D.Z., Jia, S., Dallos, P., 2004. Mechanoelectrical transduction of adult outer haircells studied in a gerbil hemicochlea. Nature 429, 766e770.
Holt, J.R., Gillespie, S.K., Provance, D.W., Shah, K., Shokat, K.M., Corey, D.P.,Mercer, J.A., Gillespie, P.G., 2002. A chemical-genetic strategy implicatesmyosin-1c in adaptation by hair cells. Cell 108, 371e381.
Howard, J., Hudspeth, A.J., 1988. Compliance of the hair bundle associated withgating of mechanoelectrical transduction channels in the bullfrog’s saccularhair cell. Neuron 1, 189e199.

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122 121
Hudspeth, A.J., 1982. Extracellular current flow and the site of transduction byvertebrate hair cells. J. Neurosci. 2, 1e10.
Hudspeth, A.J., 1985. The cellular basis of hearing: the biophysics of hair cells.Science 230, 745e752.
Hudspeth, A.J., 2005. How the ear’s works work: mechanoelectrical transductionand amplification by hair cells. C. R. Biol. 328, 155e162.
Hudspeth, A.J., 2008. Making an effort to listen: mechanical amplification in the ear.Neuron 59, 530e545.
Hudspeth, A.J., Corey, D.P., 1977. Sensitivity, polarity, and conductance change in theresponse of vertebrate hair cells to controlled mechanical stimuli. Proc. Natl.Acad. Sci. U.S.A. 74, 2407e2411.
Hudspeth, A.J., Choe, Y., Mehta, A.D., Martin, P., 2000. Putting ion channels to work:mechanoelectrical transduction, adaptation, and amplification by hair cells [InProcess Citation]. Proc. Natl. Acad. Sci. U.S.A. 97, 11765e11772.
Hudspeth, A.J., Jacobs, R., 1979. Stereocilia mediate transduction in vertebrate haircells (auditory system/cilium/vestibular system). Proc. Natl. Acad. Sci. U.S.A. 76,1506e1509.
Hudspeth, A.J., Lewis, R.S., 1988. A model for electrical resonance and frequencytuning in saccular hair cells of the bull-frog, Rana catesbeiana. J. Physiol.(London) 400, 275e297.
Indresano, A.A., Frank, J.E., Middleton, P., Jaramillo, F., 2003. Mechanical noiseenhances signal transmission in the bullfrog sacculus. J. Assoc. Res. Otolaryngol..
Iwasa, K.H., 1994. A membrane motor model for the fast motility of the outer haircell. J. Acoust. Soc. Am. 96, 2216e2224.
Jaramillo, F., Hudspeth, A.J., 1991. Localization of the hair cell’s transduction chan-nels at the hair bundle’s top by iontophoretic application of a channel blocker.Neuron 7, 409e420.
Jaramillo, F., Hudspeth, A.J., 1993. Displacement-clamp measurement of the forcesexerted by gating springs in the hair bundle. Proc. Natl. Acad. Sci. U.S.A. 90,1330e1334.
Jaramillo, F., Wiesenfeld, K., 1998. Mechanoelectrical transduction assisted byBrownian motion: a role for noise in the auditory system. Nat. Neurosci. 1,384e388.
Jia, S., He, D.Z., 2005. Motility-associated hair-bundle motion in mammalian outerhair cells. Nat. Neurosci. 8, 1028e1034.
Johnstone, B.M., Patuzzi, R., Yates, G.K., 1986. Basilar membrane measurements andthe travelling wave. Hear. Res. 22, 147e153.
Jones, E.M., Gray-Keller, M., Fettiplace, R., 1999. The role of Ca2þ-activated Kþchannel spliced variants in the tonotopic organization of the turtle cochlea [seecomments]. J. Physiol. (London) 518, 653e665.
Kazmierczak, P., Sakaguchi, H., Tokita, J., Wilson-Kubalek, E.M., Milligan, R.A.,Muller, U., Kachar, B., 2007. Cadherin 23 and protocadherin 15 interact to formtip-link filaments in sensory hair cells. Nature 449, 87e91.
Kemp, D.T., 1986. Otoacoustic emissions, travelling waves and cochlear mecha-nisms. Hear. Res. 22, 95e104.
Kennedy, H.J., Evans, M.G., Crawford, A.C., Fettiplace, R., 2003. Fast adaptation ofmechanoelectrical transducer channels in mammalian cochlear hair cells. Nat.Neurosci. 6, 832e836.
Kennedy, H.J., Crawford, A.C., Fettiplace, R., 2005. Force generation by mammalianhair bundles supports a role in cochlear amplification. Nature 433, 880e883.
Kennedy, H.J., Evans, M.G., Crawford, A.C., Fettiplace, R., 2006. Depolarization ofcochlear outer hair cells evokes active hair bundle motion by two mechanisms.J. Neurosci. 26, 2757e2766.
Köppl, C., 1997. Frequency tuning and spontaneous activity in the auditory nerveand cochlear nucleus magnocellularis of the barn owl Tyto alba. J. Neurophysiol.77, 364e377.
Koppl, C., Authier, S., 1995. Quantitative anatomical basis for a model of micro-mechanical frequency tuning in theTokaygecko,Gekkogecko.Hear. Res. 82,14e25.
Köppl, C., Forge, A., Manley, G.A., 2004. Low density of membrane particles inauditory hair cells of lizards and birds suggests an absence of somatic motility. J.Comp. Neurol. 479, 149e155.
Koppl, C., Manley, G.A., 1993. Spontaneous otoacoustic emissions in the bobtaillizard. I: general characteristics. Hear. Res. 71, 157e169.
Kossl, M., Mockel, D., Weber, M., Seyfarth, E.A., 2008. Otoacoustic emissions frominsect ears: evidence of active hearing? J. Comp. Physiol. A. Neuroethol. Sens.Neural Behav. Physiol. 194, 597e609.
Legan, P.K., Lukashkina, V.A., Goodyear, R.J., Kossi, M., Russell, I.J., Richardson, G.P.,2000. A targeted deletion in alpha-tectorin reveals that the tectorial membraneis required for the gain and timing of cochlear feedback. Neuron 28, 273e285.
Lewis, E.R., 1984. On the frog amphibian papilla. Scan Electron Microsc., 1899e1913.Liberman, M.C., Gao, J., He, D.Z., Wu, X., Jia, S., Zuo, J., 2002. Prestin is required for
electromotility of the outer hair cell and for the cochlear amplifier. Nature 419,300e304.
Liberman, M.C., Zuo, J., Guinan Jr., J.J., 2004. Otoacoustic emissions without somaticmotility: can stereocilia mechanics drive the mammalian cochlea? J. Acoust.Soc. Am. 116, 1649e1655.
Lim, D.J., 1980. Cochlear anatomy related to cochlear micromechanics. A review. J.Acoust. Soc. Am. 67, 1686e1695.
Manley, G.A., 1990. Peripheral Hearing Mechanisms in Reptiles and Birds. Springer-Verlag, Berlin, New York.
Manley, G.A., 2000a. The hearing organs of lizards. In: Dooling, R.J., Fay, R.R.,Popper, A.N. (Eds.), Comparative Hearing. Birds and Reptiles. Springer, NewYork, pp. 139e196.
Manley, G.A., 2000b. Cochlear mechanisms from a phylogenetic viewpoint. Proc.Natl. Acad. Sci. U.S.A. 97, 11736e11743.
Manley, G.A., 2001. Evidence for an active process and a cochlear amplifier innonmammals. J. Neurophysiol. 86, 541e549.
Manley, G.A., Schulze, M., Oeckinghaus, H., 1987. Otoacoustic emissions in a songbird. Hear. Res. 26, 257e266.
Manley, G.A., Yates, G.K., Koppl, C., 1988. Auditory peripheral tuning: evidence fora simple resonance phenomenon in the lizard Tiliqua. Hear. Res. 33, 181e189.
Manley, G.A., Köppl, C., Johnstone, B.M., 1990. Peripheral auditory processing in thebobtail lizard Tiliqua rugosa. J.Comp. Physiol. A 167, 89e99.
Manley, G.A., Gallo, L., Koppl, C., 1996. Spontaneous otoacoustic emissions in twogecko species, Gekko gecko and Eublepharis macularius. J. Acoust. Soc. Am. 99,1588e1603.
Manley, G.A., Kirk, D.L., Koppl, C., Yates, G.K., 2001. In vivo evidence for a cochlearamplifier in thehair-cellbundleof lizards.Proc.Natl.Acad. Sci.U.S.A.98,2826e2831.
Manley, G.A., Gallo, L., 1997. Otoacoustic emissions, hair cells, and myosin motors. J.Acoust. Soc. Am. 102, 1049e1055.
Manley, G.A., Koppl, C., 1998. Phylogenetic development of the cochlea and itsinnervation. Curr. Opin. Neurobiol. 8, 468e474.
Manley, G.A., Koppl, C., 2008. What have lizard ears taught us about auditoryphysiology? Hear. Res. 238, 3e11.
Manley, G.A., Ladher, R., 2008. Phylogeny and evolution of ciliated mechanoreceptorcells. In: Allan, I.B., Akimichi, K., Gordon, M.S., Gerald, W., Thomas, D.A.,Richard, H.M., Peter, D., Donata, O., Stuart, F., Gary, K.B., Bushnell, M.C., Jon, H.K.,Esther, G. (Eds.), The Senses: a Comprehensive Reference, vol. 3. AcademicPress, New York, pp. 1e34.
Martin, P., Mehta, A.D., Hudspeth, A.J., 2000. Negative hair-bundle stiffness betraysa mechanism for mechanical amplification by the hair cell [In Process Citation].Proc. Natl. Acad. Sci. U.S.A. 97, 12026e12031.
Martin, P., Bozovic, D., Choe, Y., Hudspeth, A.J., 2003. Spontaneous oscillation by hairbundles of the bullfrog’s sacculus. J. Neurosci. 23, 4533e4548.
Martin, P., Hudspeth, A.J., 1999. Active hair-bundle movements can amplify a haircell’s response to oscillatory mechanical stimuli. Proc. Natl. Acad. Sci. U.S.A. 96,14306e14311.
Martin, P., Hudspeth, A.J., 2001. Compressive nonlinearity in the hair bundle’s activeresponse tomechanical stimulation. Proc. Natl. Acad. Sci. U.S.A. 98,14386e14391.
Mason, A.C., Faure, P.A., 2004. The physiology of insect auditory afferents. Microsc.Res. Tech. 63, 338e350.
Megela-Simmons, a., Moss, C.F., Daniel, K.M., 1985. Behavioral audiograms of thebullfrog (Rana catesbeiana) and the green tree frog (Hyla cinerea). J. Acoust. Soc.Am. 78, 1236e1244.
Mellado Lagarde, M.M., Drexl, M., Lukashkina, V.A., Lukashkin, A.N., Russell, I.J.,2008. Outer hair cell somatic, not hair bundle, motility is the basis of thecochlear amplifier. Nat. Neurosci. 11, 746e748.
Miller, M.R., 1973. Scanning electron microscope studies of some lizard basilarpapillae. Am. J. Anat. 138, 301e329.
Miller, M.R., 1978a. Further scanning electron microscope studies of lizard auditorypapillae. J. Morphol. 156, 381e417.
Miller, M.R., 1978b. Scanning electron microscope studies of the papilla basilaris ofsome turtles and snakes. Am. J. Anat. 151, 409e435.
Moore, B.C.J., 2004. An Introduction to the Psychology of Hearing, fifth ed..Academic Press, Amsterdam, Boston.
Morrison, D., Schindler, R.A., Wersall, J., 1975. A quantitative analysis of the afferentinnervation of the organ of corti in guinea pig. Acta. Otolaryngol. 79, 11e23.
Mulroy, M.J., Williams, R.S., 1987. Auditory stereocilia in the alligator lizard. Hear.Res. 25, 11e21.
Nam, J.H., Fettiplace, R., 2008. Theoretical conditions for high-frequency hair bundleoscillations in auditory hair cells. Biophys. J. 95, 4948e4962.
Oghalai, J.S., Holt, J.R., Nakagawa, T., Jung, T.M., Coker, N.J., Jenkins, H.A., Eatock, R.A.,Brownell, W.E., 1998. Ionic currents and electromotility in inner ear hair cellsfrom humans. J. Neurophysiol. 79, 2235e2239.
Oldfield, B.P., 1988. Tonotopic organization of the insect auditory pathway. Trends.Neurosci. 11, 267e270.
Oldfield, B.P., Kleindienst, H.U., Huber, F., 1986. Physiology and tonotopic organi-zation of auditory receptors in the cricket Gryllus bimaculatus DeGeer. J. Comp.Physiol. A 159, 457e464.
Patuzzi, R.B., Bull, C.L., 1991. Electrical responses from the chicken basilar papilla.Hear. Res. 53, 57e77.
Pickles, J.O., 1984. Frequency threshold curves and simultaneous masking functionsin single fibres of the guinea pig auditory nerve. Hear. Res. 14, 245e256.
Pickles, J.O., Comis, S.D., Osborne, M.P., 1984. Cross-links between stereocilia in theguinea pig organ of Corti, and their possible relation to sensory transduction.Hear. Res. 15, 103e112.
Pitchford, S., Ashmore, J.F., 1987. An electrical resonance in hair cells of theamphibian papilla of the frog Rana temporaria. Hear. Res. 27, 75e83.
Probst, R., Lonsbury-Martin, B.L., Martin, G.K., 1991. A review of otoacoustic emis-sions. J. Acoust. Soc. Am. 89, 2027e2067.
Rhode, W.S., 1971. Observations of the vibration of the basilar membrane in squirrelmonkeys using the Mossbauer technique. J. Acoust. Soc. Am. 49 (Suppl. 2),1218þ.
Rhode, W.S., 1978. Some observations on cochlear mechanics. J. Acoust. Soc. Am. 64,158e176.
Rhode, W.S., Robles, L., 1974. Evidence from Mossbauer experiments for nonlinearvibration in the cochlea. J. Acoust. Soc. Am. 55, 588e596.
Ricci, A.J., Wu, Y.C., Fettiplace, R., 1998. The endogenous calcium buffer and the timecourse of transducer adaptation in auditory hair cells. J. Neurosci. 18,8261e8277.

A.W. Peng, A.J. Ricci / Hearing Research 273 (2011) 109e122122
Ricci, A.J., Crawford, A.C., Fettiplace, R., 2000. Active hair bundle motion linked tofast transducer adaptation in auditory hair cells [In Process Citation]. J. Neu-rosci. 20, 7131e7142.
Ricci, A.J., Crawford, A.C., Fettiplace, R., 2002. Mechanisms of active hair bundlemotion in auditory hair cells. J. Neurosci. 22, 44e52.
Ricci, A.J., Crawford, A.C., Fettiplace, R., 2003. Tonotopic variation in the conduc-tance of the hair cell mechanotransducer channel. Neuron 40, 983e990.
Ricci, A.J., Kennedy, H.J., Crawford, A.C., Fettiplace, R., 2005. The transductionchannel filter in auditory hair cells. J. Neurosci. 25, 7831e7839.
Ricci,A.J., Fettiplace,R.,1997.TheeffectsofcalciumbufferingandcyclicAMPonmechano-electrical transduction in turtle auditory hair cells. J. Physiol. (London) 501,111e124.
Richter, C.P., Emadi, G., Getnick, G., Quesnel, A., Dallos, P., 2007. Tectorial membranestiffness gradients. Biophys. J. 93, 2265e2276.
Romer, H., 1983. Tonotopic organization of the auditory neuropile in the bushcricketTettigonia viridissima. Nature 306, 60e62.
Ruggero, M.A., Rich, N.C., Robles, L., Shivapuja, B.G., 1990. Middle-ear response inthe chinchilla and its relationship to mechanics at the base of the cochlea. J.Acoust. Soc. Am. 87, 1612.
Ruggero, M.A., Rich, N.C., Recio, A., Narayan, S.S., Robles, L., 1997. Basilar-membraneresponses to tones at the base of the chinchilla cochlea. J. Acoust. Soc. Am. 101,2151e2163.
Ruggero, M.A., Rich, N.C., 1991. Application of a commercially-manufacturedDoppler-shift laser velocimeter to the measurement of basilar-membranevibration. Hear. Res. 51, 215e230.
Russell, I.J., Richardson, G.P., Kossl, M., 1989. The responses of cochlear hair cells totonic displacements of the sensory hair bundle. Hear. Res. 43, 55e69.
Russell, I.J., Legan, P.K., Lukashkina, V.A., Lukashkin, A.N., Goodyear, R.J.,Richardson, G.P., 2007. Sharpened cochlear tuning in a mouse with a geneticallymodified tectorial membrane. Nat. Neurosci. 10, 215e223.
Rybalchenko, V., Santos-Sacchi, J., 2003. Cl- flux through a non-selective, stretch-sensitive conductance influences the outer hair cell motor of the guinea-pig. J.Physiol. 547, 873e891.
Salvi, R., Saunders, S., Powers, N., Boettcher, F., 1992. Discharge patterns of cochlearganglion neurons in the chicken. J. Comp. Physiol. A Neuroethol. Sens. NeuralBehav. Physiol. 170, 227e241.
Santos-Sacchi, J., 1991. Reversible inhibition of voltage-dependent outer hair cellmotility and capacitance. J. Neurosci. 11, 3096e3110.
Santos-Sacchi, J., Dilger, J.P., 1988. Whole cell currents and mechanical responses ofisolated outer hair cells. Hear. Res. 35, 143e150.
Schoffelen, R.L., Segenhout, J.M., van Dijk, P., 2008. Mechanics of the exceptionalanuran ear. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 194,417e428.
Shen, J.X., Feng, A.S., Xu, Z.M., Yu, Z.L., Arch, V.S., Yu, X.J., Narins, P.M., 2008.Ultrasonic frogs show hyperacute phonotaxis to female courtship calls. Nature453, 914e916.
Shotwell, S.L., Jacobs, R., Hudspeth, A.J., 1981. Directional sensitivity of individualvertebrate hair cells to controlled deflection of their hair bundles. Ann. N. Y.Acad. Sci. 374, 1e10.
Siemens, J., Lillo, C., Dumont, R.A., Reynolds, A., Williams, D.S., Gillespie, P.G.,Muller, U., 2004. Cadherin 23 is a component of the tip link in hair-cell ster-eocilia. Nature 428, 950e955.
Simmons, D., Meenderink, S., Vassilakis, P., 2006. Anatomy, physiology, and functionof auditory end-organs in the frog inner ear. In: Hearing and Sound Commu-nication in Amphibians, pp. 184e220.
Slifer, E.H., Sekhon, S.S., 1975. The femoral chordotonal organs of a grasshopper,Orthoptera, Acrididae. J. Neurocytol. 4, 419e438.
Sneary, M.G., 1988a. Auditory receptor of the red-eared turtle: I. General ultra-structure. J. Comp. Neurol. 276, 573e587.
Sneary, M.G., 1988b. Auditory receptor of the red-eared turtle: II. Afferent andefferent synapses and innervation patterns. J. Comp. Neurol. 276,588e606.
Stauffer, E.A., Scarborough, J.D., Hirono, M., Miller, E.D., Shah, K., Mercer, J.A.,Holt, J.R., Gillespie, P.G., 2005. Fast adaptation in vestibular hair cells requiresMyosin-1c activity. Neuron 47, 541e553.
Stewart, C.E., Hudspeth, A.J., 2000. Effects of salicylates and aminoglycosides onspontaneous otoacoustic emissions in the Tokay gecko. Proc. Natl. Acad. Sci.U.S.A. 97, 454e459.
Stiebler, I.B., Narins, P.M., 1990. Temperature-dependence of auditory nerveresponse properties in the frog. Hear. Res. 46, 63e81.
Stumpner, A., Molina, J., 2006. Diversity of intersegmental auditory neurons ina bush cricket. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 192,1359e1376.
Sul, B., Iwasa, K.H., 2009. Effectiveness of hair bundle motility as the cochlearamplifier. Biophys. J. 97, 2653e2663.
Taberner, A.M., Liberman, M.C., 2005. Response properties of single auditory nervefibers in the mouse. J. Neurophysiol. 93, 557e569.
Tanaka, K., Smith, C.A., 1978. Structure of the chicken’s inner ear: SEM and TEMstudy. Am. J. Anat. 153, 251e271.
Taschenberger, G., Manley, G.A., 1997. Spontaneous otoacoustic emissions in thebarn owl. Hear. Res. 110, 61e76.
Tilney, M.S., Tilney, L.G., DeRosier, D.J., 1987. The distribution of hair cell bundlelengths and orientations suggests an unexpected pattern of hair cell stimulationin the chick cochlea. Hear. Res. 25, 141e151.
Tilney, L.G., Saunders, J.C., 1983. Actin filaments, stereocilia, and hair cells of the birdcochlea. I. Length, number, width, and distribution of stereocilia of each hair cellare related to the position of the hair cell on the cochlea. J. Cell Biol. 96,807e821.
de Vries, H., 1948a. Brownian movement and hearing. Physica 14, 48e60.de Vries, H., 1948b. The minimum audible energy. Acta Oto-Laryngologica 36,
230e235.Waguespack, J., Salles, F.T., Kachar, B., Ricci, A.J., 2007. Stepwise morphological and
functional maturation of mechanotransduction in rat outer hair cells. J. Neu-rosci. 27, 13890e13902.
Warr, W.B., 1992. Organization of olivocochlear efferent systems in mammals. In:Webster, D.B., Popper, A.N., Fay, R.R. (Eds.), The Mammalian Auditory Pathway:Neuroanatomy. Springer-Verlag, New York, pp. 410e448.
Warr, W.B., Boche, J.B., Neely, S.T., 1997. Efferent innervation of the inner hair cellregion: origins and terminations of two lateral olivocochlear systems. Hear. Res.108, 89e111.
Weaver, S.P., Schweitzer, L., 1994. A radial gradient of fibril density in the gerbiltectorial membrane. Hear. Res. 76, 1e6.
Wever, E.G., 1967. The tectorial membrane of the lizard ear: types of structure. J.Morphol. 122, 307e319.
Wibowo, E., Brockhausen, J., Koppl, C., 2009. Efferent innervation to the auditorybasilar papilla of scincid lizards. J. Comp. Neurol. 516, 74e85.
Wu, Y.C., Art, J.J., Goodman, M.B., Fettiplace, R., 1995. A kinetic description of thecalcium-activated potassium channel and its application to electrical tuning ofhair cells. Prog. Biophys. Mol. Biol. 63, 131e158.
Wu, Y.-C., Ricci, A.J., Fettiplace, R., 1999. Two components of transducer adaptationin auditory hair cells. J. Neurophysiol. 82, 2171e2181.
Wu, X., Gao, J., Guo, Y., Zuo, J., 2004. Hearing threshold elevation precedes hair-cellloss in prestin knockout mice. Brain Res. Mol. Brain. Res. 126, 30e37.
Yack, J.E., 2004. The structure and function of auditory chordotonal organs ininsects. Microsc. Res. Tech. 63, 315e337.
Zheng, J., Shen, W., He, D.Z., Long, K.B., Madison, L.D., Dallos, P., 2000. Prestin is themotor protein of cochlear outer hair cells. Nature 405, 149e155.