specificity of the induction of ferritin synthesis by hemin
TRANSCRIPT

146 Biochimica et Biophysica Acta, 1050 (1990) 146-150 Elsevier
BBAEXP 92101
Specificity of the induction of ferritin synthesis by hemin
J.J. L in 1, S. D a n i e l s - M c Q u e e n 1, L. G a f f i e l d 2, M . M . P a t i n o 2, W.E. W a l d e n 2 an d R.E. T h a c h 1
t Department of Biology, Washington University, St. Louis, MO and 2 Department of Microbiology and Immunology, University of Illinois at Chicago, Chicago, IL (U.S.A.)
(Received 16 May 1990)
Key words: Ferritin synthesis; Hemin; Ferritin repressor protein; (Rabbit)
We have previously reported that hemin derepresses ferritin mRNA translation in vitro. As noted earlier, pre-incuhation of a 90 kDa ferritin repressor protein (FLIP) with hemin prevented subsequent repression of ferritin synthesis in a wheat germ extract. The significance of this observation has been investigated further. Evidence is presented here that this inactivation of FRP is temperature dependent. Neither FeCI3, Fe 3÷ chelated with EDTA, nor protoporphyrin IX caused significant inactivation of FlIP under comparable conditions, whereas Zn z +-protoporphyrin IX produced an intermediate degree of inhibition. The presence of a glutathione redox buffer (GSB), which was previously shown to minimize non-specific side-effeets of hemin, was not necessary for the derepression reaction. Inclusion of mannitol, a free radical scavenger, did not alter the inactivation caused by hemin. Calculation of the expected ratio of hemin monomers to dimers suggests that the active species is the monomer.
Introduction
Ferritins are intracellular proteins that retain iron in a non-toxic form [1-3]. Their synthesis is regulated most dramatically at the translational level in response to the extracellular iron supply [4-9]. Rapid progress has been made in recent years in identifying the ele- ments which comprise the translational regulatory mac- hinery. The first of these elements is a sequence of 28 nucleotides that is found in the 5' untranslated region (5' UTR) of all ferritin mRNAs [10,11], which has been named the 'iron responsive element' (IRE). The pres- ence of an IRE in the 5' UTR of a mRNA confers iron responsiveness upon the translation of any downstream open reading frame (ORF). The second component of the regulatory machinery is a 90 kDa protein that can bind to the IRE when iron availability is low. In doing so it inhibits the translation of adjacent ORFs [12-16]. This protein has been called the 'ferritin repressor pro- tein' (FRP).
Abbreviations: FRP, ferritin repressor protein; GSB, glutathione re- dox buffer; 5' UTR, 5' untranslated region; IRE, iron responsive dement
Correspondence: R.E. Thach, Department of Biology, Washington University, St. Louis, MO 63130, U.S.A.
We have recently shown that hemin has many of the properties expected of a third component of this regu- latory machinery, which may be referred to as the 'ferritin inducer' [17]. In particular, we have shown that hemin interacts with the FRP, either directly or indi- rectly, in such a way as to relieve the repression of ferritin mRNA translation. Further evidence is pre- sented here which describes the requirements for, and specificity of, this reaction.
Materials and Methods
[35S]Methionine ( > 800 Ci/mmol) was obtained from Amersham, GeneScribe-Z TM was from United States Biochemical Corp., mTG(5')ppp(5')G cap analog was from Pharmacia, RNase inhibitor and T7 RNA poly- merase were from Boehringer Mannheim Biochemicals, DNase I and DNA grade Sephadex G-25 from Phar- macia, wheat germ extract and amino acids (minus methionine), were from Promega Corp., and bemin, Co 3+-, Zn 2+- or Sn2+-protoporphyrin IX, and proto- porphyrin IX were from Porphyrin Products. All pro- phyrin solutions were made fresh immediately prior to each experiment according to the manufacturer's in- structions.
Transcripts encoding rabbit L-ferritin and human apolipoprotein A-I were prepared in vitro from plas- mids bearing the corresponding cDNA sequences using T7 RNA polymerases as described previously [17]. An
0167-4781/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

S3PI preparation of FRP was partially purified from rabbit reticulocytes [14], and then fractionated on heparin agarose as described [16]. The peak fractions, which were estimated to contain approx. 2% FRP, were employed in the experiments described here. The FRP preparations were stored in 200 mM KC1, 20 mM Tris-HC1 (pH 7.4), 10% glycerol, 0.1 mM EDTA and 2 mM DTT.
In most translation reactions, 9.4 /~g of partially purified reticulocyte FRP were pre-incubated for 30 rain at 37°C with varying concentrations of heroin (or analogues thereof), a redox buffer comprising a 5 :1 mixture of glutathione to oxidized glutathione (usually at 2.5 mM and 0.50 mM, respectively; referred to as GSB), 20 mM Hepes-KOH (pH 7.4) and residual salts and buffers carried over from diluted stock solutions (these were approx. 70 mM KC1, 8 mM Hepes-KOH (pH 7.4), 7 mM Tris-HC1 (pH 7.4), 3% glycerol, 0.03 mM EDTA and 0.7 mM DTT in a final volume of 3.2 /~l. After the pre-incubation step the reactions were chilled on ice, and other components necessary for translation were added. Translation reactions (10 /~1 final volume) ordinarily contained 10 fmol each of rabbit L-ferritin and human apolipoprotein A-I tran- scripts, 3 /~1 of wheat germ extract, and 11.5 mM Hepes-KOH (pH 7.4), 102 mM K(OAc), 1.75 mM Mg(OAc)2, 0.8 mM amino acids (minus methionine), and 3 #Ci [3SS]methionine. The Mg 2÷ concentration in the wheat germ extract was reduced by addition of 0.25 mM EDTA. Decreasing the Mg 2÷ concentration re- suited in increased fidelity of initiation at the correct initiation codon [18]. Translation reactions were in- cubated at 24°C for 60 rain. Translation products were analyzed by SDS-PAGE using 12% acrylamide gels, followed by fluorography [19,20]. Where indicated, fluo- rograms were scanned using a densitometer (Joyce Loebl).
For the analysis of restriction endonuclease sensitiv- ity to heroin, XhoI, EcoRI, NciI, HpalI and NarI (15, 16, 15, 5 and 16 units, respectively) were incubated with FRP, GSB, and heroin according to the standard prein- cubation protocol for 30 min at 37 ° C. Then, 1/~1 of the reaction containing XhoI, and 0.1 #1 of the other reac- tions, were combined with appropriate DNA plasmids (1/~g), salts and buffers as recommended by the restric- tion endonuclease suppliers in final volumes of 10/~1. These reactions were then incubated for 30 min at 37 o C, and samples were analyzed by agarose gel elec- trophoresis in the presence of ethidium bromide.
For the calculation of the monomeric hemin con- centration, the equation of Brown et al. [21] was em- ployed, where K = [dimer][H ÷ ]/[monomer] 2 = 4.5.
Results
As described previously [17], FRP was ordinarily pre-incubated with hemin or analogues thereof in the
147
t 2 3 4 S 6 "t 8 9 lOll 12 131415161"t ie
0----
f.-.~
G S B : + + + + + . . . . + + + + + . . . .
F R P : - + + + + + + + + - + + + + + + + - - "
I II I Pre - inc Temp: 3 0 = 3 7 =
Fig. 1. Derepression of ferritin mRNA translation by heroin. All reactions were preceded by a 30 min pre-incubation step at 30 °C or 37 o C, which contained reticulocyte FRP, 200/~M heroin or FeC13 or FeNaEDTA and GSB, where indicated, in a final volume of 3.2 /~l. The mixtures were then chilled and components required for transla- tion were added. In all experiments the subsequent translation reac- tion was carded out at 24°C for 1 h. Ferritin and apolipoprotein products (indicated by 'a' and 'f', respectively) were then resolved by
SDS-PAGE and visualized by fluorography.
presence or absence of a redox buffer comprised of glutathione plus oxidized glutathione (termed GSB) for various times and temperatures. Wheat germ lysate, L-ferritin and apolipoprotein A-I transcripts, [35S]- methionine and other components required for transla- tion were then added, and the mixture was further incubated for 1 h at 24 ° C. The labelled ferritin and apolipoprotein products synthesized during the second incubation were analyzed by SDS-PAGE and fiuorogra- phy. (Details of all procedures are described in Materi- als and Methods). A typical result is shown in Fig. 1. Here it is evident that derepression by hemin is temper- ature dependent (Fig. 1; compare lane 5 with lane 14, or lane 8 with lane 17). The presence of GSB is not required for this reaction (compare lane 5 with lane 8, or lane 14 with lane 17). Higher temperatures (e.g., 42°C; not shown) tended to inactivate FRP in the presence or absence of heroin or GSB. Trivalent iron in other forms, such as FeC13 (lanes 3, 6, 12 and 15) or Fe3+-EDTA (lanes 4, 7, 13 and 16) did not mimic the effect of heroin.
A variety of heroin analogues have been tested for relief activity. This was done in the presence or absence of 10 mM mannitol, which effectively scavenges the hydroxyl radical [22]. Mannitol did not alter the relief activity of hemin (Fig. 2; compare lanes 7 and 8), suggesting that the hemin effect is not due to the generation of hydroxyl radicals. Several hemin ana- logues, such as protoporphyrin IX (lanes 15 and 16), exhibited little or no relief activity. Zn2+-proto- porphyrin IX showed an intermediate level of activity (Fig. 2, Lanes 11 and 12). SnE+-protoporphyrin IX inhibited synthesis of apolipoprotein at concentrations above 10 /~M so its efficacy as an inducer of ferritin

148
CI--~
f_,.
I 2 3 4 5 6 7 8 9 I0 II 12 1 3 1 4 1 5 1 6
F R P : - - * + - - + + - - + + - - + +
Monnitol:- + - + - + - + - + - + - + - + I I I I I I
P°rphyrin:t none Heroin Z n ~ ' - P P ~ P P I I E
Fig. 2. Derepression of ferritin mRNA translation by hemin ana- logues. FRP was preincubated with 200 #M heroin or protoprophyrin IX, or 100 #N Zn2+-protoporphyrin IX and 10 mM mannitol, where indicated, and GSB for 30 rain at 37°C. Subsequent translation reactions and product analyses were conducted as described in Fig. 1.
synthesis could not be evaluated. In contrast, Co 3 +-pro- toporphyrin IX was found to be slightly more effective than hemin in relieving repression [17]. The fact that Co 3+ but not Zn 2+ can effectively replace Fe 3+ in the porphyrin complex suggests that a specific binding site for hernin exists on the FRP molecule. Indeed, competi- tion studies revealed that neither protoporphyrin IX nor Zn2+-protoporphyrin IX at high concentrations signifi- cantly reduced the derepressive effect of hemin (not shown). This is in apparent contrast with the enzyme heine oxygenase, for example, where the inhibitory con- stants (K i) of Co 3 +- and Zn 2 +-protoporphyrin IX differ by only a factor of 4 [23].
Inasmuch as heroin and other metalloporphyrins are known to be a highly reactive [24-26], the possibility that the inactivation of FRP may be artifactual must be considered. The data presented here and previously [17] render this unlikely, since effects of hemin on transla- tion of the internal standard message (coding for apo- lipoprotein) were minimal, especially in the presence of GSB. However, to address this question further we studied the effects of heroin on a variety of restriction endonucleases. The concentrations of these enzymes
were first adjusted to be non-limiting over the time course studied, so that even very small inhibitory effects could be detected. These enzymes were then included in standard preincubation reactions (as in lane 7 of Fig. 2, for example), in duplicate, containing either 200 ~tM hemin or no hemin. Subsequently, a 0.1 ~tl or 1 /L1 aliquot was withdrawn and added to 9/zl of DNA plus appropriate buffers and salts. Digestion of the DNA was monitored at various times by agarose gel electro- phoresis. A typical result is shown in Fig. 3, where the cleavage of DNA after 30 min is shown. Here it is evident that EcoR I and NarI, for example, have both cleaved about one third of the M2 DNA (which has a single cleavage site for each enzyme) after 30 min. Preincubation of these enzymes with hemin clearly did not affect their ability to cleave DNA. A similar result was obtained with XhoI, although due to the generally poor stability of this enzyme the duplicate reactions were not identical. While the reactions with XhoI and HpalI had almost reached completion, studies with lower amounts of enzyme showed little if any effect of heroin. Similar results were obtained with SalI and HindlII (not shown). From this we conclude that hemin is not generally toxic to enzymes that recognize nucleic acids under the conditions used here.
As noted previously [17], the concentrations of hemin and Co3+-protoporphyrin IX that produced half-maxi- mal derepression of reticulocyte FRP were relatively high, on the order of 50/~M. This compares to the value of 5-20/~M heroin that is required to inhibit the elF-2 kinase, known as the hemin regulated inhibitor (HRI), in reticulocytes [27]. Similarly, 50-200 /JM hemin has been employed to induce ferritin synthesis in cells grow- ing in culture [10,11]. These values contrast strikingly with the concentration of free or loosely bound heroin that exists in vivo, which is thought to be in the order of 0.1/zM in the absence of exogenous iron [28,29]. How- ever, because of the well-known tendency of metallo-
lin MI.
lineor M2 DNA
]-]er~irl~ | "l" ÷ ~ I i m ~ '{P ' t ~ ~ m 't1" 't" I ~ ~ 't" "11" ~ ~ "1" 41" I I I I I
Xho I uncut Eco RI Nci ! uncut Hpan Nor T M! M2
DNA DNA Fig. 3. Absence of inhibition of restriction endonaclease activity by heroin. Duplicate samples of XhoI, EcoRI, NciI, HpalI, and NarI were each preincubated with FRP and GSB for 30 min at 37 ° C, in the presence or absence of 200 pM hemin. Reactions were then chilled, and aliquots were withdrawn and added to reaction mixtures containing M1 DNA [14] for XhoI, or M2 DNA [14] for the other enzymes, plus salts and buffers optimal for each enzyme. These second reactions were then incubated at 37 o C. Samples were withdrawn after 30 rain and analyzed for cleavage of
DNA by agarose gel electrophoresis.
Z lOOnM
-I- lOaM • • _o n.- bJ
InM
OJ nM
o~ O.OInM O,OInM OJnM InM IOnM IOOnM IpM IOpM I l~d IO00~JM
TOTAL HI:'MIN
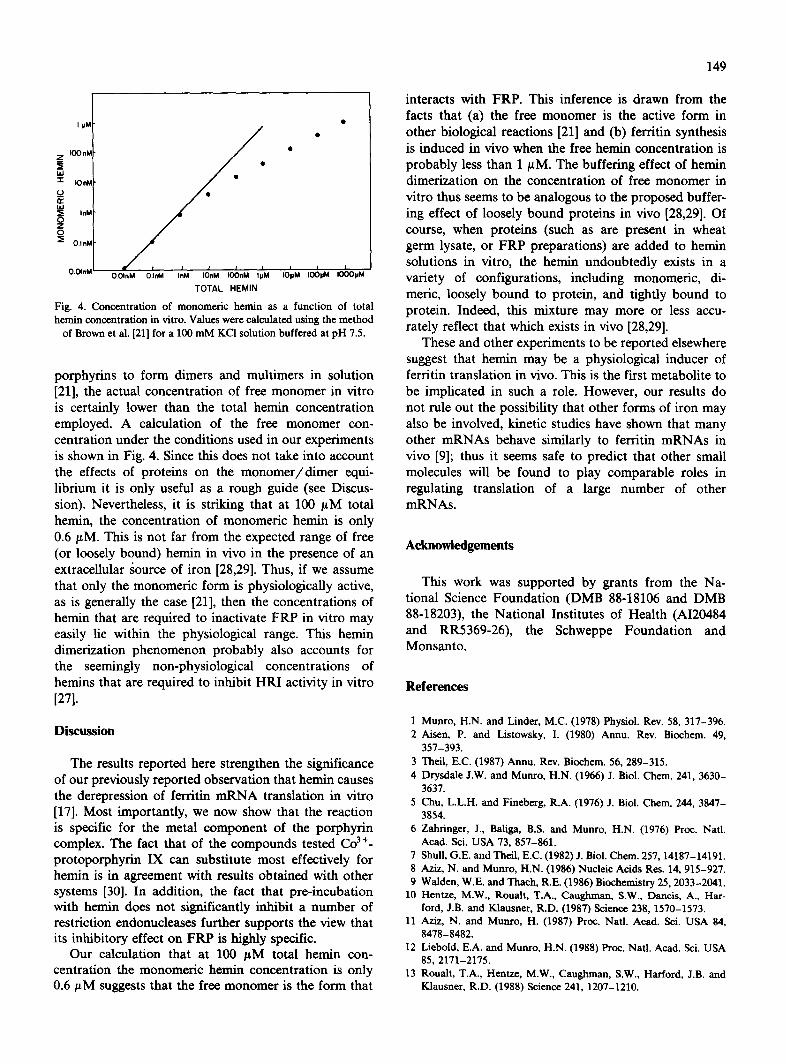
Fig. 4. Concentration of monomeric heroin as a function of total heroin concentration in vitro. Values were calculated using the method
of Brown et al. [21] for a 100 mM KC1 solution buffered at pH 7.5.
porphyrins to form dimers and multimers in solution [21], the actual concentration of free monomer in vitro is certainly lower than the total hemin concentration employed. A calculation of the free monomer con- centration under the conditions used in our experiments is shown in Fig. 4. Since this does not take into account the effects of proteins on the monomer /d imer equi- librium it is only useful as a rough guide (see Discus- sion). Nevertheless, it is striking that at 100 /tM total hemin, the concentration of monomeric hemin is only 0.6/ tM. This is not far from the expected range of free (or loosely bound) hemin in vivo in the presence of an extracellular ~ource of iron [28,29]. Thus, if we assume that only the monomeric form is physiologically active, as is generally the case [21], then the concentrations of hemin that are required to inactivate FRP in vitro may easily lie within the physiological range. This hemin dimerization phenomenon probably also accounts for the seemingly non-physiological concentrations of hemins that are required to inhibit HRI activity in vitro [271.
Discussion
The results reported here strengthen the significance of our previously reported observation that hemin causes the derepression of ferritin mRNA translation in vitro [17]. Most importantly, we now show that the reaction is specific for the metal component of the porphyrin complex. The fact that of the compounds tested Co 3 ÷- protoporphyrin IX can substitute most effectively for hemin is in agreement with results obtained with other systems [30]. In addition, the fact that pre-incubation with hemin does not significantly inhibit a number of restriction endonucleases further supports the view that its inhibitory effect on FRP is highly specific.
Our calculation that at 100 /tM total hernin con- centration the monomeric heroin concentration is only 0.6 #M suggests that the free monomer is the form that
149
interacts with FRP. This inference is drawn from the facts that (a) the free monomer is the active form in other biological reactions [21] and (b) ferritin synthesis is induced in vivo when the free hemin concentration is probably less than 1/~M. The buffeting effect of heroin dimerization on the concentration of free monomer in vitro thus seems to be analogous to the proposed buffer- ing effect of loosely bound proteins in vivo [28,29]. Of course, when proteins (such as are present in wheat germ lysate, or FRP preparations) are added to hemin solutions in vitro, the heroin undoubtedly exists in a variety of configurations, including monomeric, di- meric, loosely bound to protein, and tightly bound to protein. Indeed, this mixture may more or less accu- rately reflect that which exists in vivo [28,29].
These and other experiments to be reported elsewhere suggest that hemin may be a physiological inducer of ferritin translation in vivo. This is the first metabolite to be implicated in such a role. However, our results do not rule out the possibility that other forms of iron may also be involved, kinetic studies have shown that many other mRNAs behave similarly to ferritin mRNAs in vivo [9]; thus it seems safe to predict that other small molecules will be found to play comparable roles in regulating translation of a large number of other mRNAs.
Acknowledgements
This work was supported by grants from the Na- tional Science Foundation (DMB 88-18106 and DMB 88-18203), the National Institutes of Health (AI20484 and RR5369-26), the Schweppe Foundat ion and Monsanto.
References
1 Munro, H.N. and Linder, M.C. (1978) Physiol. Rev. 58, 317-396. 2 Aisen, P. and Listowsky, I. (1980) Annu. Rev. Biochem. 49,
357-393. 3 Theil, E,C. (1987) Armu. Rev. Biochem. 56, 289-315. 4 Drysdale J.W. and Munro, H.N. (1966) J. Biol. Chem. 241, 3630-
3637. 5 Chu, L.L.H. and Fineberg, R.A. (1976) J. Biol. Chem. 244, 3847-
3854. 6 Zahringer, J., Baliga, B.S. and Munro, H.N. (1976) Proc. Natl.
Acad. SCi. USA 73, 857-861. 7 Shull, G.E. and Theft, E.C. (1982) J. Biol. Chem. 257, 14187-14191. 8 Aziz, N. and Munro, H.N. (1986) Nucleic Acids Res. 14, 915-927. 9 Walden, W.E. and Thach, R.E. (1986) Biochemistry 25, 2033-2041.
10 Hentze, M.W., Rouah, T.A., Caughman, S.W., Daneis, A., Har- ford, J.B. and Klansner, R.D. (1987) Science 238, 1570-1573.
11 Aziz, N. and Munro, H. (1987) Proc. Natl. Acad. Sci. USA 84, 8478-8482.
12 Liebold, E.A. and Munro, H.N. (1988) Proc. Natl. Acad. SCi. USA 85, 2171-2175.
13 Roualt, T.A., Hentze, M.W., Caughrnan, S.W., Harford, J.B. and Klausner, R.D. (1988) Science 241, 1207-1210.

150
14 Walden, W.E., Daniels-McQueen, S., Brown, P.H., Garfield, L., Russell, D.A., Bielser, D., Bailey, L.C. and Thach, R.E. (1988) Proc. Natl. Acad. Sci. USA 85, 9503-9507.
15 Brown, P.H., Daniels-McQueen, S., Walden, W.E., Patino, M.M., Gaffield, L., Bielser, D. and Thach, R.E. (1989) J. Biol. Chem. 264, 13383-13386.
16 Walden, W.E., Patino, M.M. and Gaffield, L. (1989) J. Biol. Chem. 264, 13765-13769.
17 Lin, J.-J., Daniels-McQueen, S,, Patino, M.M., Gaffield, L., Walden, W.E. and Thach, R.E. (1990) Science 247, 74-77.
18 Dasso, M.C. and Jackson, R.J. (1987) Translational Control, Cold Spring Harbor Laboratory, Cold Spring Harbor.
19 Walden, W.E., Daniels-McQueen, S., Smith, L.L. and Thach, R.E. (1987) Gene 61,314-327.
20 Giulian, G.G., Shanahan, M.F., Graham, J.M. and Moss, R.L. (1985) Fed. Proc. 44, 686.
21 Brown, S.B., Hatzikonstantinou, H. and Herries, D. (1980) Int. J. Biochem. 12, 701-707.
22 Floyd, R.A., West, M.S., Eneff, K.L., Hogsett, W.E. and Tingey, D.T. (1988) Arch. Biochem. Biophys. 262, 266-272.
23 Abraham, N.G., Mitrione, S.M., Hodgson, W.J., Levere, R.D. and Shibihara, S. (1988) Adv. Exp. Med. Biol. 241, 97-116.
24 Vincent, S.H. and Muller-Eberhard, U. (1987) J. Lab. Clin. Med. 110, 475-482.
25 Vincent, S.H., Grady, R.W., Shaklai, N., Snider, J.M. and Muller- Eberhard, U. (1988) Arch. Biochem. Biophys. 265, 539-550.
26 Aft, R.L. and Mueller, G.C. (1985) J. Biol. Chem. 259, 301-305. 27 London, I.M., Levin, D.H., Matts, R.L., Thomas, N.S.B.,
Petryshyn, R. and Chen, J.-J. (1987) in The Enzymes (Boyer, P.D. and Krebs, E.G. eds) Vol. XVIII, Part B, pp. 360-377, Academic Press, Orlando.
28 Granick, S., Sinclair, P., Sassa, S. and Grieninger, G. (1975) J. Biol. Chem. 250, 9215-9225.
29 Grandchamps, B., Bissell, D.M., Licko, V. and Schmid, R. (1981) J. Biol. Chem. 256, 11677-11683.
30 Chang, C.S. and Sassa, S. (1982) J. Biol. Chem. 257, 3650-3654.