spindle shape changes as an indicator of …jcs.biologists.org/content/joces/10/1/79.full.pdf ·...
TRANSCRIPT

J. Cell Sci. 10, 79-93 (1972) 79
Printed in Great Britain
SPINDLE SHAPE CHANGES AS AN INDICATOR
OF FORCE PRODUCTION IN CRANE-FLY
SPERMATOCYTES
J. R. L A F O U N T A I N , JR.*
Department of Biological Sciences, State University of Neiv York at Albany, Albany,Nerv York 12203, U.S.A.
SUMMARY
Changes in spindle shape during the first and second meiotic divisions in living Nephrotomasvturalis spermatocytes have been studied and recorded cinematographically with phase-contrast,Nomarski differential interference, and polarization microscopy. With the Nomarski system2 classes of discrete spindle filaments (new terminology) have been observed: continuousfilaments that extend between the poles and appear to form a cage-like framework within whichthe chromosomes move, and kinetochore filaments which are attached to the chromosomes attheir kinetochores and converge toward the poles. Spindle deformation occurs in a mannerconsistent with the hypothesis that whatever tensile forces are transmitted by the kinetochorefilaments are balanced by compressive forces acting on the cage-like framework of continuousfilaments causing them to become splayed. Just when these forces would be expected to reacha maximum, kinetochore filaments undergo a noticeable increase in contrast. It has not beendetermined whether this change represents a crowding of linear elements or addition of drymass to a constant number of linear elements.
INTRODUCTION
The occurrence of dimensional changes (i.e. prometaphase elongation, metaphaseshortening, and anaphase elongation) of mitotic spindles at specific times during thedivision process has been described for several different types of cells (Duncan &Peridsky, 1958; Jacquez & Biesele, 1954; Taylor, i960), but little functional signifi-cance has been ascribed to these changes of shape. Very striking deformations occurin crane-fly spermatocyte spindles, which are clearly demarked by a mitochondrialsheath, and anaphase movement is preceded by a noticeable spindle shortening andwidening (Dietz, 1969). The present study was undertaken to document these dimen-sional changes in spermatocyte spindles of the crane fly, Nephrotoma suturalis, anddetermine their probable cause.
The Zeiss/Nomarski differential interference equipment for transmitted-lightmicroscopy was used extensively in this study to render visible small gradients inoptical path, such as those produced by linear elements of the mitotic spindle (Allen,David & Nomarski, 1969). This study represents the first detailed investigation ofan animal spindle with the Nomarski system under conditions comparable to thoseused in studies on Haemanthus endosperm by Bajer & Allen (1966a, b).
• Present address: Laboratorium fur Elektronenmikroskopie, Institut fiir AllgemeineBotanik, Eidgendssische Technische Hochschule, Zurich, Switzerland.

80 J. R. LaFountain, Jr.
MATERIALS AND METHODS
Larvae of Nephrotoma suturalis were cultured in the laboratory on macerated tissue paperand ground nettle leaves according to the method of Forer (1964). Spermatocytes from testesof fourth instar larvae were spread as a monolayer under KEL-F 10 oil on to a large (24 x50 mm) coverglass, which was inverted on to a smaller (22 x 40 mm) coverglass to make asandwich and sealed with Valap (a mixture of Vaseline, lanoline and paraffin). A ring of Vaselinewas applied to the smaller coverglass before making the sandwich, and it served as a spacer toreduce cell flattening. The coverglass sandwich was then mounted on to a brass holder that fitson to a microscope stage. Spermatocytes prepared in this way could live for 4-5 h at roomtemperature (20—23 °C), and both meiotic divisions were observed in many cells.
Microscopical observations were made with a Zeiss Photomicroscope I equipped with selectedhigh-extinction Nomarski differential interference and polarization optics and positive phase-contrast optics. A neofluar (40/0-75 N.A.) objective was used for phase contrast; strain-free(40/085 N.A.) and achromatic (40/085 N.A. oil immersion) objectives were used with polariza-tion optics; and an achromat (40/0-85 N.A. oil immersion) and a planachromat (100/125 N.A.oil immersion) were used with the Nomarski system. Tungsten and mercury arc (with a green(546 run) filter) lamps were used with phase and Nomarski optics, and a Spectra-Physics argon-ion laser (LaFountain, Muckenthaler, & Allen, 1968) was used as a light source for the polariza-tion microscope. Operational extinction factors for the Nomarski equipment ranged between200 and 250, while extinction factors for the polarization optics were between 5 x io3 and8x io3.
Cin6 records were made on Eastman Recordak Micro-file AHU, Plus X Negative, and Ekta-chrome Commercial films with an Arriflex 16-mm camera driven by a Sage series 500 Cin6-photomicrographic apparatus; 3 5-mm records were made with the Photomicroscope I automaticcamera on Adox KB 14 film. Black and white films were processed in Diafine and colour filmswere processed commercially.
RESULTS
Nephrotoma spermatocytes are cultured so that the spindles are not flattened buthave a circular cross-section in a plane perpendicular to the pole-to-pole axis. Spindlelength is defined as the distance between the 2 poles of the spindle, and the accuracyof measurements of spindle length depends on the accuracy with which the poles canbe determined. Since the spindle poles are defined in these cells as isotropic regionsat the polar ends of the highly birefrigent chromosomal fibres, comparisons of inter-polar distances at different stages of division can be made with the polarization micro-scope (Fig. 1). Spindle width (at the spindle equator) can be determined accuratelybecause the spindle border is defined at its periphery by a dense mitochondrialsheath, which completely encases the spindle (Figs. 2, 3).
In order to translate spindle shape changes as viewed in the plane of the pole-to-pole axis into shape changes of the entire spindle, it is necessary to know if the shapeof the spindle is constant about the pole-to-pole axis. If spindles are axially symmetri-cal, then the shape of the entire spindle is defined by its longitudinal median section.As a test of spindle symmetry, ocular micrometer measurements of spindle equatorialwidth (perpendicular to the optical axis) were compared with measurements of spindlewidth parallel to the optical axis made with the calibrated fine focus of the Photo-microscope I. For these measurements Nomarski optics were used to 'optically sec-tion' (Allen et al. 1969) cells and thereby determine the vertical distance from theupper boundary of the spindle to the lower boundary as defined by the mitochondrial

Spindle shape changes and force production 81
sheath. Over 20 measurements were made of cells during different stages of divisionin several preparations and were restricted to spindles in which both poles were clearlyin the same focal plane. In all cases studied, spindle equatorial width along both axeswas the same (the 2 values differed from one another by no more than 1 /tm), indicatingthat the spindles were not flattened by the preparation method used and that anassumption of axial symmetry along the pole-to-pole axis is valid.
During meiosis I in Nephrotoma spermatocytes, a spindle begins to form betweenthe 2 asters immediately after the nuclear membrane breaks down, and the chromo-somes generally begin to undergo random metakinetic movements and chromosomestretching. (A full description of these and other chromosome movements will not beincluded here because they closely resemble phenomena observed in other species ofcrane flies, which have been described in great detail by Dietz, 1956.) As the spindleforms, the asters move in opposite directions and establish the spindle poles (Fig. 2 A,B). Some structural rigidity on the part of the spindle can be inferred from the factthat cells elongate parallel to the spindle axis, and also, neighbouring cells may bepushed aside to allow for this elongation. Within 15-20 min after nuclear membranebreakdown, the spindle appears ellipsoidal and has the shape of a prolate spheroid(Fig. 2B).
As the chromosomes become stabilized into a metaphase plate configuration, thespindle itself shortens, while its equatorial diameter increases. Micrographs madewith either phase-contrast or Nomarski optics illustrate this change (Fig. 2 c). More-over, time-lapse cine records made with the polarization microscope permit anaccurate determination of the extent of spindle shortening (Fig. 1). In the mostextreme cases, interpolar distance is reduced as much as 20% during metaphase ascompared to early prometaphase. Just before the onset of anaphase, spindles appearnearly diamond-shaped in optical section, and their shape approximates that of abicone (Fig. 2 c). After separation of sister dyads and during anaphase, spindle shapegradually changes once more. The diamond shape of metaphase slowly changes intoan ellipsoid, and the shape of the spindle returns to a prolate spheroid (Fig. 2E).Spindle width decreases, and spindle length slowly increases (Fig. 1), so that in manyinstances, spindle shape at the moment when dyads reach the poles is almost identicalto the prolate spheroid observed during prometaphase. The main observable differencebetween the 2 stages is the position of the chromosomes within the spindle. Althoughnot all spindles studied showed anaphase elongation back to prometaphase length,none showed interpolar distances greater than prometaphase values until after cyto-kinesis begins (Fig. 2F).
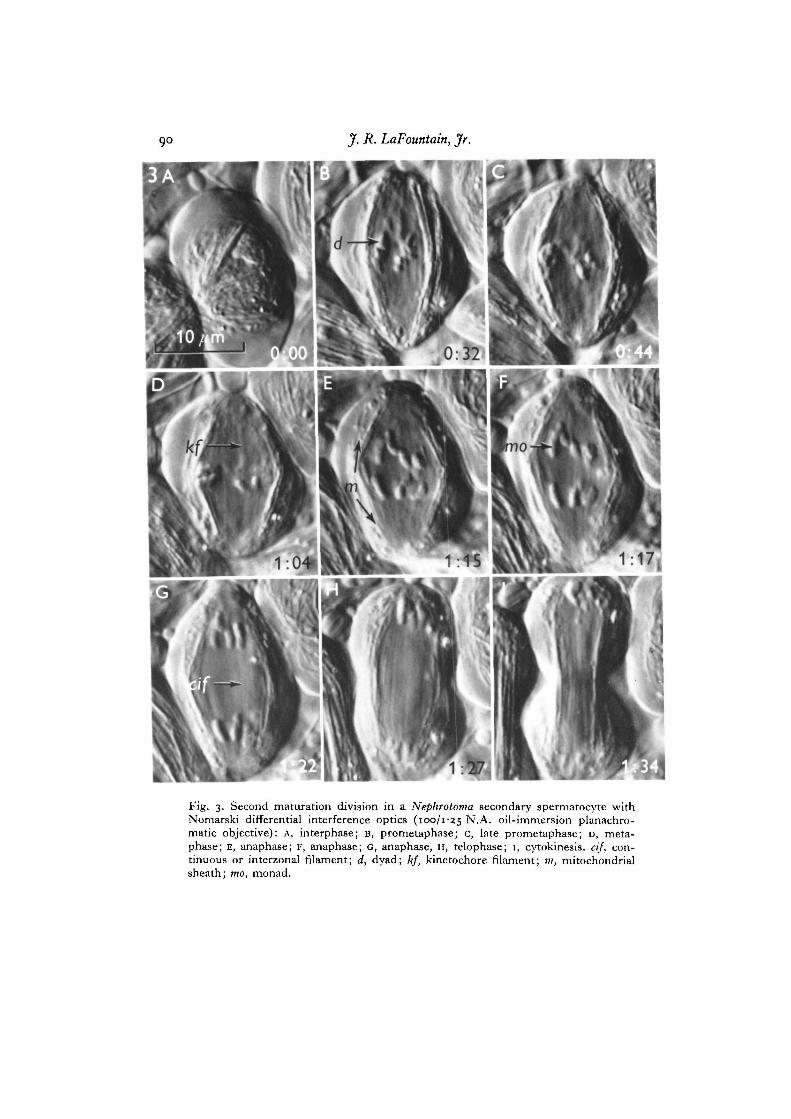
Spindles in secondary spermatocytes of Nephrotoma show shape changes similar tothose observed in meiosis I (Fig. 3). Just before the monads are transported to thepoles, the equatorial diameter of secondary spindles reaches a maximum and thelength is reduced. During anaphase, spindle shape gradually changes to a prolatespheroid. Also, as was the case in meiosis I, spindle length does not increase to morethan what it was during prometaphase until after cytokinesis ensues.
Exploration of Nephrotoma spermatocyte spindles with the Nomarski system hasrevealed several new details about the linear elements in these spindles. Both primary

82 J. R. LaFountain, Jr.
and secondary spindles contain numerous longitudinal fibrils which are visible asrefractile bodies (Figs. 2-7) with a diameter of 0-2-0-3 /tm- (Iftne reader is unfamiliarwith the interpretation of differential interference images, an earlier description of thestructure and organization of the mitotic spindle in Haemanthus endosperm (Bajer &Allen, 1966 a), as well as a description of the Nomarski system (Allen et al. 1969),should be consulted for details.) These elements can be observed extending from thekinetochore regions of the chromosomes toward the poles and also extending betweenthe poles without any apparent connexions to the chromosomes. Because these ele-ments are not similar in either size or distribution to the longitudinal spindle ele-ments previously observed with the light microscope (i.e. birefringent spindle fibresdescribed by Forer, 1966, 1969), the term spindle filament has been applied tothese newly revealed structures to mark this distinction. Spindle filaments thatextend from the kinetochore regions of the bivalents and univalents to the poles arecalled kinetochore filaments, and filaments that extend between the poles and do notterminate at kinetochores are called continuous filaments.
During the first maturation division, numerous spindle filaments can be seen inNephrotoma spindles soon after the nuclear membrane breaks down, and within a fewminutes after the onset of spindle formation, the entire spindle region becomespacked with filaments. Optical sections through the spindle during prometaphasedemonstrate that continuous filaments appear somewhat arced (Fig. 2B) and forma cage-like framework within which the chromosomes move. It is possible in somecells to trace single continuous filaments from one pole to the other. However, due tothe slight curvature of these elements, micrographs of single filaments extending frompole to pole are not available. Kinetochore filaments are connected to the bivalentchromosomes at the kinetochores and extend toward the poles. Although kinetochorefilaments have not been traced all the way to the poles, they can be traced very closeto the poles (Fig. 5 A), and they are distributed one per autosomal chromatid (Figs. 5 A,6 A). Thus, birefringent chromosomal fibres of sister dyads are made up of 2 kineto-chore filaments (Fig. 5). In the cases of unpaired sex univalents, each chromatid has2 kinetochore regions with a kinetochore filament radiating from each (Fig. 6B).Although kinetochore filaments are distributed 2 per univalent chromatid, a ratioof 2 kinetochore filaments per chromosomal fibre is maintained.
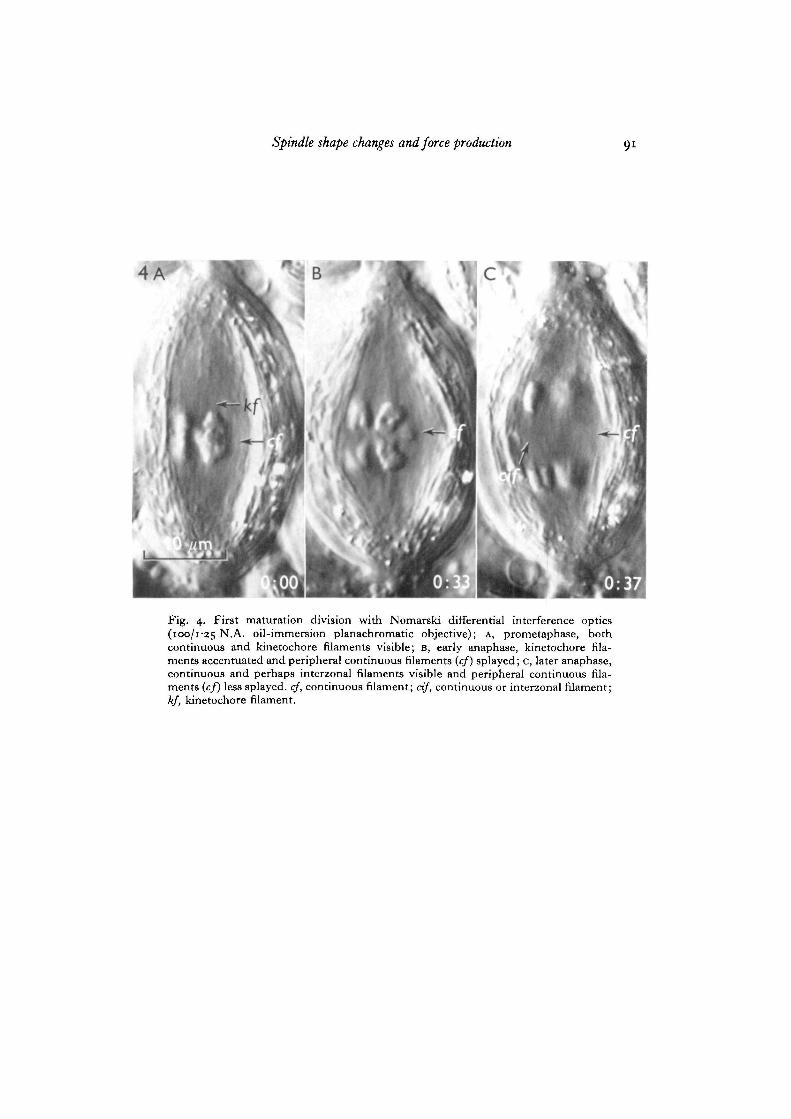
As metaphase approaches and the shape of the spindle gradually changes to abicone, the distance between the centrally located chromosomes and the mitochondrialsheath generally increases, and some of the more peripheral continuous filamentssplay out in this region between the chromosomes and the mitochondrial sheath(Fig. 7). In some instances the appearance of the continuous filaments suggests thatthey actually bend at the spindle equator (Fig. 4). During this time the visibility of,or contrast generated by, kinetochore filaments becomes markedly enhanced overthat during prometaphase (Fig. 7). This increased contrast takes place without anyapparent change in filament thickness. During anaphase, kinetochore filaments be-come shorter and remain distinct from continuous filaments (Fig. 4). The kinetochorefilaments of the lagging sex univalents are easily distinguished from continuous andperhaps interzonal filaments because they maintain a state of enhanced contrast until

Spindle shape changes and force production 83
and during the time they are transported to the poles during telophase (Fig. 2D, E).(Interzonal fibres are clearly visible at the level of the light microscope (Schrader,i953;Bajer, 1968; Forer, 1969). At present, however, one cannot exclude the possibilitythat these are segments of continuous filaments, the courses of which could have beenchanged within chromosomes.)
The arrangement of spindle filaments in secondary spermatocytes closely resemblesthat found in meiosis I (Fig. 3). Continuous filaments appear to be distributed through-out the spindle forming a kind of longitudinal framework, while kinetochore filamentsare attached to the chromosomes at the kinetochores. Enhanced contrast in kinetochorefilaments can also be observed before and during anaphase, but the appearance of thisphenomenon is not as striking as in meiosis I.
DISCUSSION
Regarding the mechanism of chromosome transport during cell division, it is gener-ally believed that poleward-pulling forces applied to the kinetochores are responsiblefor anaphase movement (for review: Forer, 1969; Nicklas, 1970). Chromosomal fibresundoubtedly are involved in either generating or transmitting the tensile forces thatresult in movement (Forer, 1966, 1969). Since the spindle poles approach one anotherto a limited extent, the spindle must contain supporting elements that resist these ten-sile forces. The presence and arrangement of continuous or interpolar microtubules inmany plant and animal spindles suggests that the role of these elements is to providethe supporting framework within which chromosomes move (Bajer, 1968; Nicldas,1970; Nicklas & Staehley, 1967). The findings obtained here in living crane-fly meioticspindles offer additional evidence that continuous elements provide the structuralframework needed to support the intraspindle forces that bring about chromosomalmovement. In addition, the appearance and distribution of continuous filaments inNephrotoma spindles, coupled with the dramatic shape changes of the entire spindleduring the course of division, are compatible with a simple hypothesis regarding thedevelopment and deployment of forces within the spindle from prometaphase throughanaphase. That is, tensile forces between the chromosomes and poles develop longbefore anaphase movement commences, and the different stages of division (prometa-phase, metaphase, and anaphase) are results of continued action of the same mechanism.Accordingly, metaphase is a state of equilibrium when the forces counterbalanceone another (Ostergren, 1945, Ostergren, Mole-Bajer & Bajer, i960; Mclntosh,Hepler & VanWie, 1969).
The occurrence of active chromosome movement and chromosome stretchingduring prometaphase in Nephrotoma spermatocytes suggests that tensile forces actingbetween the chromosomes and poles are already active at that time (Dietz, 1956;Nicklas, 1970). In early prometaphase continuous filaments are distributed more orless uniformly throughout the spindle and some of the more peripheral ones appearslightly arced. As prometaphase proceeds and the chromosomes became stabilizedon to the spindle equator, the outer continous filaments, lying close to the peripheryof the spindle, tend to splay out, or bend to a greater degree, causing the spindle to
6-2

84 J- R- LaFountain, Jr.
widen and assume the shape of a bicone. These changes can be explained in terms ofincreased compression applied to continuous filaments due to increased tensile forcesbetween the chromosomes and the poles. As tension toward one pole is balanced byforces pulling in the opposite direction, the chromosomes equilibrate at a point halfway between the 2 poles. The result of this is metaphase, during which time spindledeformation reaches a maximum, and there is no net chromosome movement to thepoles.
According to this interpretation, it would be expected that tension on the kineto-chore filaments and the balancing compression on continuous filaments would bothdiminish upon separation of sister dyads and the onset of anaphase. The results bearthis out. When the chromosomes move to the poles during anaphase, spindle widthdecreases and the continuous filaments become less splayed (Fig. 4). Also, interpolardistance generally increases (Fig. 1), but never beyond the maximum establishedduring prometaphase. Spindle elongation during anaphase may then be interpretedas a result of the release of continuous filaments from compression. If this entireinterpretation is correct, then the shape of the spindle may be a sensitive indicator ofthe magnitude of the balanced forces acting within the spindle.
Included in this interpretation is an assumption that the apparent contraction ofchromosomes during metaphase is not a result of a reduction in the tensile forcesapplied to them (as was suggested by Dietz, 1956) but is due to force-independentchanges within the chromosomes themselves. Since chromosome condensation occursduring other stages (i.e. prophase and telophase) without the influence of motiveforces, it is concluded that the above assumption calling for self-condensation is valid.
The discovery of both continuous and kinetochore filaments as refractile bodies inNephrotoma spindles has raised several interesting questions. First, what is therelationship between spindle filaments and microtubules? That is, are spindle fila-ments composed of several microtubules (Bajer & Jensen, 1969), and if so, how manymicrotubules are there per filament? Secondly, what is the relationship betweenspindle filaments and birefringent spindle fibres seen with the polarization microscope?Continuous filaments can be found in regions where little or no birefringence can bedetected (the interzone (see Forer, 1969) and the more peripheral regions of thespindle between the chromosomes and the mitochondrial sheath). These findings plusother data suggest that the interpretation of birefringence solely in terms of orientedfilamentous material may not be acceptable (J. R. LaFountain, in preparation).Thirdly, what is the significance of the marked increase in contrast generated by kine-tochore filaments during the later stages of prometaphase through anaphase? Sincethe steepness of the shadows cast across linear elements with the Nomarski system is afunction both of the gradient in optical path difference in the object and the instru-mental bias retardation (Allen et al. 1969), changes in the contrast of an object at con-stant bias retardation reflect size and/or refractive index changes. If kinetochorefilaments change in size at this time, the change must be a subtle one. In order toaccount for an increase in contrast, the elements comprising the spindle filamentwould have to become more condensed, thus increasing the mean refractive index inthe filament area. Alternatively, the same effect could be achieved by the addition of

Spindle shape changes and force production 85
dry mass to the filament. One might speculate whether some undetected' last elements'(e.g. microtubule cross-bridges, additional microtubules, microfilaments, etc.) of theforce-producing mechanism might be laid down at this stage and contribute sufficientdry mass to be observed as enhanced contrast.
Despite the extensive study of Behnke & Forer (1966) on Nephrotoma spindles, ourknowledge of spindle ultrastructure in these cells is very incomplete. In order toanswer any of the questions raised above, a more thorough investigation of theultrastructure of Nephrotoma spindles must be made. Studies toward this end are nowbeing conducted.
The author thanks Dr Robert D. Allen for his advice and criticism during the course of thisinvestigation; Dr F. A. Muckenthaler for help during the early stages; and Robert Speck andDale Rice for technical assistance.
This work was supported by a predoctoral fellowship from the National Institute of Health(1-F1-GM-40, 135-01) and Program Project Grant GM 08691 from the National Institute ofGeneral Medical Sciences administered by Dr R. D. Allen.
REFERENCES
ALLEN, R. D., DAVID, G. B. & NOMARSKI, G. (1969). The Zeiss/Nomarski differential inter-ference equipment for transmitted-light microscopy. Z. iviss. Mikrosk. 69, 193-221.
BAJER, A. (1968). Behavior and fine structure of spindle fibers during mitosis in endosperm.Chronwsoma 25, 249—281.
BAJER, A. & ALLEN, R. D. (1966a), Structure and organization of the living mitotic spindle ofHaemanthus endosperm. Science, N.Y. 151, 572-574.
BAJER, A. & ALLEN, R. D. (19664). Role of phragmoplast filaments in cell-plate formation.J. Cell Sci. 1, 4SS-462.
BAJER, A. & JENSEN, C. (1969). Detectability of mitotic spindle microtubules with the light andelectron microscopes. J. Microscopie 8, 343-354.
BEHNKE, O. & FORER, A. (1966). Some aspects of microtubules in spermatocyte meiosis in acrane fly (Nephrotoma suturajis, Loew): Intranuclear and intrachromosomal microtubules.C. r. Trav. Lab. Carhberg 35, 437-455-
DIETZ, R. (1956). Die Spermatocytenteilungen der Tipuliden II. Graphische Analyse derChromosomenbewegung wahrend der Prometaphase I im Leben. Cliromosoma 8, 183-211.
DIETZ, R. (1969). Bau und Funktion des Spindelapparatus. Naturwissenschaften 56, 237-248.DUNCAN, R. E. & PERIDSKY, M. D. (1958). The achromatic figure during mitosis in maize
endosperm. Am. J. Bot. 45, 719-729.FORER, A. (1964). Evidence for Two Spindle Fiber Components: A Study of Chromosome Move-
ment in Living Crane-fly (Nephrotoma suturalis) Spermatocytes using Polarization Microscopyand an Ultraviolet Microbeam. Doctoral Thesis, Dartmouth College.
FORER, A. (1966). Characterization of the mitotic traction system and evidence that birefringentspindle fibers neither produce nor transmit force for chromosome movement. Cliromosoma19, 44-98.
FORER, A. (1969). Chromosome movements during cell-division. In Handbook of MolecularCytology (ed. A. Lima-de-Faria), pp. 553-601. Amsterdam: North-Holland Publishing Co.
JACQUEZ, J. A. & BIESELE, J. J. (1954). A study of Michel's film on meiosis in Psophus stridulus,L. Expl Cell Res. 6, 17-29.
LAFOUNTAIN, J., MUCKENTHALER, F. & ALLEN, R. D. (1968). Argon-ion laser as a source forphysical microscopy. Biophys. J. 8, A159.
MCINTOSH, J. A., HEPLER, P. K. & VANWIE, D. G. (1969). A model for mitosis. Nature, Lond.224, 659-663.
NICKLAS, R. B. (1970). Mitosis. In Advances in Cell Biology, vol. 2 (ed. D. M. Prescott, L.Goldstein & E. H. McConkey). New York: Appleton-Century-Crofts.

86 J. R. LaFountain, Jr.
NICKLAS, R. B. & STAEHLEY, C. A. (1967). Chromosome micromanipulation. I. The mechan-ism of chromosome attachment to the spindle. Chromosoma 21, 1-16.
OSTERGREN, G. (1945). Equilibrium of trivalents and the mechanism of chromosome movement.Hereditas 31, 498.
OSTERGREN, G., MOLE-BAJER, L. & BAJER, A. (i960). An interpretation of transport phenomenaat mitosis. Ann. N.Y. Acad. Sci. 90, 381-408.
SCHRADER, F. (1953). Mitosis, Hit Movement of Chromosomes in Cell Division, 2nd edn. NewYork: Columbia University Press.
TAYLOR, E. W. (i960). Dynamics of spindle formation. Ann. N.Y. Acad. Sci. 90, 430-434.
(Received 8 January 1971—Revised 21 June 1971)
Fig. 1. Three different Nephrotoma suturalis spermatocytes during meiosis I withpolarization microscopy (40/085 N.A. oil-immereion achromatic objective and laserillumination): A-F, first cell from early prometaphase through anaphase showingprometaphase elongation (A, B), metaphase shortening (c, D), and anaphase elongation(E, F); G-I, second cell from prometaphase (G, H) to metaphase (1); J-L: third cellduring metaphase (j) and anaphase (K, L). Birefringent bands in the interzone arethe chromosomal fibres of the lagging univalent sex chromosomes (u). Arrows locatepoles. Spindle fibre birefringence appears as either positive (white) or negative(black) contrast depending on compensator setting.

Spindle shape changes and force production

J. R. LaFountain, Jr.
Fig. 2. First meiodc division in a Nephrotoma primary spermatocyte with Nomarskidifferential interference optics (100/125 N.A. oil-immersion planachromatic objec-tive) and mercury arc illumination: A, late diakinesis; B, prometaphase; c, metaphase;D, anaphase; E, telophase; F, cytokinesis, cf, continuous filament; cif, continuous orinterzonal filament; d, dyad; kf, kinetochore filament; m, mitochondrial sheath;t, tetrad; u, univalent sex chromosome.

Spindle shape changes and force production
get J. R. LaFountain, Jr.
Fig. 3. Second maturation division in a Nephrotoma secondary spermatocyte withNomarski differential interference optics (100/125 N.A. oil-immersion planachro-matic objective): A, interphase; B, prometaphase; c, late prometaphase; D, meta-phase; E, anaphase; F, anaphase; c, anaphase, H, telophase; 1, cytokinesis, cif, con-tinuous or interzonal filament; d, dyad; kf, kinetochore filament; m, mitochondrialsheath; mo, monad.
Spindle shape changes and force production
4A
Fig. 4. First maturation division with Nomareki differential interference optics(100/1-25 N.A. oil-immersion planachromatic objective); A, prometaphase, bothcontinuous and kinetochore filaments visible; B, early anaphase, kinetochore fila-ments accentuated and peripheral continuous filaments {cf) splayed; c, later anaphase,continuous and perhaps interzonal filaments visible and peripheral continuous fila-ments (cf) less splayed, cf, continuous filament; cif, continuous or interzonal filament;kf, kinetochore filament.

92 J. R. La Fountain, Jr.
Fig. 5. A Nephrotoma primary spermatocyte with both A, Nomarski optics (100/1 25 N.A. oil-immersion planachromatic objective) and B, polarization optics (40/085 N.A. oil-immersion achromatic objective and mercury arc illumination). Thebirefringent chromosomal fibre (arrow) in B contains 2 kinetochore filaments (arrowin A).
Fig. 6. A, Nephrotoma primary spermatocyte during metaphase with Nomarski optics(100/125 N.A. oil-immersion planachromatic objective); kinetochore filaments aredistributed one per autosomal chromatid, or 2 per dyad (arrow). B, Univalent sexchromosomes remain at the spindle equator during autosomal anaphase I and have2 kinetochores per chromatid with a kinetochore filament radiating from each kineto-chore (arrow). Nomarski optics (100/1-25 N.A. oil immersion planachromaticobjective).
Fig. 7. A Nephrotoma primary spermatocyte with Nomarski optics (100/1-25 N.A.oil-immersion planachromatic objective): A, prometaphase; B, metaphase. Kineto-chore filaments show enhanced contrast during metaphase (compare contrast infilaments in A (arrow) with contrast in B) and peripheral continuous filaments becomesplayed (paired arrows in B).

Spindle shape changes and force production