stability synthesis of penicillin-binding proteins duringjb.asm.org/content/156/2/545.full.pdf ·...
TRANSCRIPT

Vol. 156, No. 2JOURNAL OF BACTERIOLOGY, Nov. 1983, P. 545-5510021-9193/83/110545-07$02.00/0Copyright © 1983, American Society for Microbiology
Stability and Synthesis of the Penicillin-Binding ProteinsDuring Sporulation
CHRISTINE E. BUCHANAN* AND MARGARET 0. SOWELLtDepartment ofBiology, Southern Methodist University, Dallas, Texas 75275
Received 22 June 1983/Accepted 30 August 1983
The penicillin-binding proteins (PBPs) of Bacillus subtilis were examined afterincubation of vegetative and sporulating cultures with chloramphenicol, aninhibitor of protein synthesis. The results indicate that the sporulation-specificincreases in vegetative PBPs 2B and 3 and the appearance of two new PBPs, 4*and 5*, depend on concurrent protein synthesis, which is most likely to be denovo synthesis of the PBPs rather than synthesis of an activator or processingenzyme. It was also learned that in vivo the PBPs differ in their individualstabilities, which helps to explain some of the quantitative changes that occur inthe PBP profile during sporulation. All the membrane-bound PBPs, exceptpossibly PBP 1, were found to be stable in the presence of crude extracts ofsporulating cells that contained proteolytic activity.
Both vegetative growth and sporulation canbe inhibited by penicillin, so it is likely that thefunctions of some penicillin-binding proteins(PBPs) are normally required for both processes(1, 6, 9, 13, 24). We previously examined thechanges that occur in the membrane-boundPBPs during the early stages of sporulation inBacillus subtilis to determine which of the vege-tative PBPs are most likely to be involved insporulation and whether the penicillin-sensitiveevents in sporulation require the synthesis ofany new PBPs (20). We found that vegetativePBPs 1, 2A, 4, and 5 decrease in amount duringsporulation; PBPs 2B and 3 increase at specifictimes; and two new PBPs, 4* and 5*, appear.From a comparison of the PBP profiles in sporu-lating cells and two stage 0 (asporogenous) mu-tants, we concluded that the changes in PBPs 2Band 3, as well as the appearance of PBPs 4* and5*, are likely to be sporulation-related eventsrather than the consequences of stationaryphase aging.
Several questions were raised by these obser-vations. First, what event induced the sharpdecline in the membrane concentrations of PBPs1, 2A, 4, and 5 at the beginning (to) of sporula-tion? Since the in vivo stabilities of the PBPs ofB. subtilis have never been measured, the turn-over of any of these proteins during sporulationis open to several explanations. Second, what isthe nature of the sporulation-related increases intwo vegetative PBPs? That is, are there substan-tial amounts of PBPs 2B and 3 in some cryptic
t Present address: Medical University of South Carolina,Charleston, SC 29425.
form throughout vegetative growth which areonly activated by a specific trigger formed dur-ing sporulation, or does de novo synthesis ofthese two PBPs occur? Finally, what is therelationship of the two new PBPs to preexistingproteins? Although the previously reported datado not support the notion that PBPs 4* and 5*are generated by proteolytic processing of any ofthe vegetative PBPs present at to, the possibilityremains that either or both of these PBPs couldhave been derived from undetectable pre-PBPslocated in the membrane.
In this paper we describe the effects of treat-ment with a protein synthesis inhibitor on thePBP changes that normally occur during sporu-lation. Chloramphenicol (CAM) was used be-cause it has access to the sites of protein synthe-sis until at least 8 h after the start of sporulation(t8) (10). We found that protein synthesis wasrequired for the sporulation-specific increases inPBPs 2B and 3 and for the appearance of PBPs4* and 5*. Other results suggest that this reflectsde novo synthesis of these proteins. In addition,we learned that the PBPs have different in vivostabilities; PBP 2A was the least stable and PBPs3 and 5 were always the most stable.
MATERUILS AND METHODSOrganisms. B. subtilis 168T (a prototroph) was ob-
tained from Indiana BioLabs, Palmyra, Ind. Theasporogenous mutant AspoOA677 is Bacillus GeneticStock Center strain 1S53.
Sporulation conditions. Both organisms were rou-tinely grown at 37°C in a supplemented nutrient broth(Difco Laboratories, Detroit, Mich.) (19) until late-exponential phase and then were shifted down to thenutrient-deficient medium of Sterlini and Mandelstam
545
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

546 BUCHANAN AND SOWELL
(22) to induce sporulation (20). The time at which thecells were resuspended was designated to. Sporulationwas monitored by assaying for the production ofalkaline phosphatase, a late stage II marker, at eachhour after induction (7) and by examining samples ofthe culture with a phase-contrast microscope for theappearance of refractile forespores, a stage IV marker.As reported previously (20), alkaline phosphatase nor-mally begins to increase at 3.5 h after a downshiftinduction (t35), and refractile forespores first appear atabout t6.5 in this particular strain.
Preparation of membranes and assay for PBPs. Themembranes were prepared from sonically disruptedcells as described previously (3, 20). The membrane-bound PBPs were detected by incubation of the mem-branes with [3H]benzylpenicillin (20) followed by sodi-um dodecyl sulfate-polyacrylamide gel electrophoresisand fluorography (2, 5). The individual PBPs werequantitated by scanning the fluorographs with a softlaser scanning densitometer (Biomed Instruments,Inc., Chicago, Ill.). Duplicates of the labeled to mem-branes were run on each gel. Every point in Fig. 1 and2 represents the average of several binding assays witheach of four membrane preparations. Only the statisti-cally significant differences or similarities in theamounts of the PBPs are discussed in this paper.
Assay for protease. The protease activity in theculture supernatants was measured as described byDancer and Mandelstam (7) with 20 mg of hide powderazure substrate (17) per tube. The extracellular serineprotease was distinguished from the metalloproteaseby its resistance to a 1 mM concentration of theinhibitor 1,10-phenanthroline (7).The preparation of crude cell extracts for assay of
intracellular protease activity followed the method ofReysset and Millet (16). This method was designed tominimize contamination of the extracts with any extra-cellular protease. The protein concentration of eachextract was determined by the Lowry technique (14).The protease assay was usually performed on extractsamples corresponding to 500 ,ug of protein in a finalvolume of 5 ml. All solutions used during the extractpreparation and assay contained 2 mM CaC12 to avoidinactivation of the calcium-dependent intracellular en-zyme.
Chemicals. Reagents for the protease assays werepurchased from Calbiochem, La Jolla, Calif., all elec-trophoresis reagents were from Bio-Rad Laboratories,Richmond, Calif., and chloramphenicol, 1,10-phen-anthroline, and alkaline phosphatase substrate werepurchased from Sigma Chemical Co., St. Louis, Mo.[3H]benzylpenicillin (ethylpiperidine salt, 25 Ci/mmol)was a gift from P. J. Cassidy of Merck Sharp & DohmeLaboratories, West Point, Pa.
RESULTSStability of the vegetative PBPs in vivo. Turn-
over of the PBPs was not measured by theconventional pulse-chase methods because thePBPs are relatively minor components of the cellmembrane that can only be distinguished fromother membrane proteins by their ability to bindradiolabeled penicillin. Instead, we treated thecells in culture with an inhibitor of proteinsynthesis for 4 h to permit the disappearance of
unstable proteins. By comparing the PBPs pre-sent in the cells before and after such treatment,it was possible to determine the stability of theindividual PBPs relative to the majority of mem-brane proteins and relative to one another (4).CAM (final concentration, 100 jig/ml) was
added to cultures of B. subtilis under two condi-tions: when the cells were in the mid-exponen-tial phase of vegetative growth in a supplement-ed nutrient broth (19), and immediately after thegrowing cells had been shifted to the sporulationmedium of Sterlini and Mandelstam (22). Thislatter culture was equivalent to the to sample insporulation studies. There was very little in-crease in the optical density of either culture andno apparent lysis of cells during the 4 h oftreatment. The results from the two differentcultures were similar (compare the dashed anddotted lines extending from 0 to 4 h in Fig. 1 and2). The rate of loss of a PBP from membranes ofcells in the rich medium was either not signifi-cantly different from the loss of the correspond-ing PBP in sporulation medium or else was morerapid.
In the absence of new protein synthesis, dif-ferences in the relative stabilities of the PBPscould be detected. PBPs 3 and 5 were clearly themost stable since very little of either protein waslost from the membranes during the period whengrowth was inhibited. The small increase in PBP3 that appeared to occur in the sporulationmedium (Fig. 2) was not statistically significant(the average relative amount of PBP 3 at t4 in thetreated culture was 1.17 ± 0.19 for six assays)and thus is not likely to be due to CAM-resistantprotein synthesis. PBP 2A was consistently themost unstable PBP, whereas the stabilities of theremaining vegetative PBPs could be classified asintermediate. Nearly identical results were ob-tained when tetracycline was used to inhibitprotein synthesis for 4 h (C. E. Buchanan andM. 0. Sowell, in R. Hakenbeck, J. V. Holtje,and H. Labischinski, ed., The Target ofPenicil-lin, in press).
Effect of CAM on PBP synthesis and turnoverduring sporulation. CAM was added for differenttitne intervals (to to t4, t2 to t4, and t4 to t6) tocultures of sporulating cells to determine whichof the PBP changes depended on concurrentprotein synthesis. The appearance of the twonew PBPs 4* and 5* as well as the sporulation-related increases in vegetative PBPs 2B and 3were completely inhibited by CAM (Fig.. 2).When CAM was added after these PBPs hadbegun to increase, the amount of each proteinimmediately began to decline. This result wasparticularly striking for PBP 3 since it was stablein CAM-treated cultures at an earlier stage ofsporulation. For all four of these PBPs, it ap-peared as if synthesis and turnover were simul-
J. BACTERIOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STABILITY AND SYNTHESIS OF THE PBPs 547
PBP 2A1.0
).8
).2
).2 .. ",0
O 2 4 6Hours
1.0
0.8
0.6F04 F
PBP 4
' 's0.21-
2 4 6Hours
1.0,
0.8
0.6
PBP 5____
0.4-
0.2
O2 4 6Hours
FIG. 1. Stability of the vegetative PBPs in the early stages of sporulation. The amounts of each membrane-bound PBP are given relative to the amount present at to. Sporulating cultures treated with CAM for various timeintervals after to are indicated by the dashed lines, whereas the single vegetative culture treated for 4 h with CAMis indicated by the dotted line. The solid lines represent the amount of the PBP in the untreated control cultureduring the first 6 h after induction of sporulation. Vegetative cells were grown in supplemented nutrient broth(19) and sporulating cells were grown in nutrient-deficient sporulation medium (22).
taneous events, with the rate of synthesis ex-
ceeding the rate of turnover for a few hours.Very soon after induction of sporulation, the
membrane-bound amount of PBP 5 declined toapproximately 60% of its vegetative (to) level(Fig. 1). The decline then continued until at leastt1o but at a much slower rate (Fig. 1) (20). Otherproteins have also been reported to exhibit bi-phasic decay during sporulation, which probablyreflects changes in the enzyme composition ofsporulating cells (21). Unlike the response ofPBPs 1, 2A, and 4, no change in the amount ofPBP 5 occurred when CAM was present from to
C
0
E
PBP 2B
(Fig. 1). CAM did not prevent the slower rate ofloss of this protein when it was added to theculture at a later stage of sporulation.At about stage III of sporulation, which corre-
sponds to t4 or earlier in Fig. 1 and 2, the rate ofloss of vegetative PBPs 1, 2A, and 4 becamegreater in the presence of CAM than in itsabsence. This change in the pattern of responseof these three proteins to the inhibitor suggeststhat the small increases in PBPs 1 and 2A andthe cessation in the drop-off ofPBP 4 at this timemay be due to a resumption of their synthesis inthe untreated cultures and could be a reflection
PBP 3
PBP 4* PBP 5*
0.8 1 0.8 0.Q2 L 02 -
0.60
4600
24
Q6-. S 0.4 ~ ~ °O 2 4 6 °O 2 4 6Hours Hours
0.4L ' ' ' 0.4 .0 2 4 6 0 2 4 6
Hours HoursFIG. 2. Effect of CAM on sporulation-specific synthesis of PBPs. The amounts of vegetative PBPs 2B and 3
are given relative to their to levels, whereas the amounts of the new PBPs 4* and 5* are relative to their maximumvalues achieved during sporulation. The dashed, dotted, and solid lines correspond to the same culturesdescribed in the legend to Fig. 1.
1.0O
0.8
PBP 1
-1..
bX0
.-
c
0
E
10-a
0.6[
04F
I
C
C
a0.21
O 2 4 6Hours
VOL. 156, 1983
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
548 BUCHANAN AND SOWELL
of the membrane growth that occurs duringstage III engulfment of the forespore. However,their original to membrane concentrations arenever regained before the amounts of thesePBPs again decrease (20).
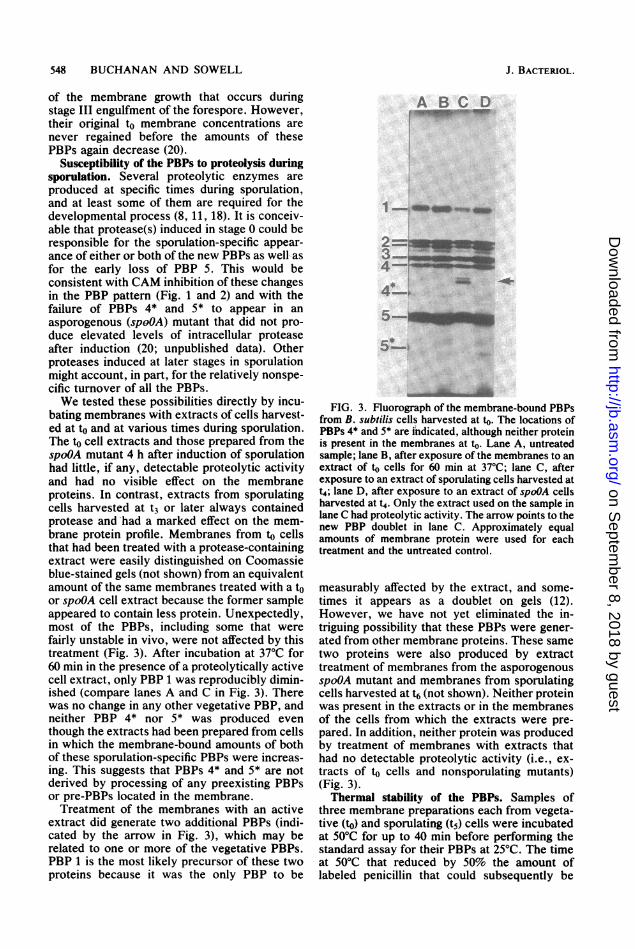
Susceptibility of the PBPs to proteolysis duringsporulation. Several proteolytic enzymes areproduced at specific times during sporulation,and at least some of them are required for thedevelopmental process (8, 11, 18). It is conceiv-able that protease(s) induced in stage 0 could beresponsible for the sporulation-specific appear-ance of either or both of the new PBPs as well- asfor the early loss of PBP 5. This would beconsistent with CAM inhibition of these changesin the PBP pattern (Fig. 1 and 2) and with thefailure of PBPs 4* and 5* to appear in an
asporogenous (spoOA) mutant that did not pro-duce elevated levels of intracellular proteaseafter induction (20; unpublished data). Otherproteases induced at later stages in sporulationmight account, in part, for the relatively nonspe-cific turnover of all the PBPs.We tested these possibilities directly by incu-
bating membranes with extracts of cells harvest-ed at to and at various times during sporulation.The to cell extracts and those prepared from thespoOA mutant 4 h after induction of sporulationhad little, if any, detectable proteolytic activityand had no visible effect on the membraneproteins. In contrast, extracts from sporulatingcells harvested at t3 or later always containedprotease and had a marked effect on the mem-brane protein profile. Membranes from to cellsthat had been treated with a protease-containingextract were easily distinguished on Coomassieblue-stained gels (not shown) from an equivalentamount of the same membranes treated with a toor spoOA cell extract because the former sampleappeared to contain less protein. Unexpectedly,most of the PBPs, including some that werefairly unstable in vivo, were not affected by thistreatment (Fig. 3). After incubation at 37°C for60 min in the presence of a proteolytically activecell extract, only PBP 1 was reproducibly dimin-ished (compare lanes A and C in Fig. 3). Therewas no change in any other vegetative PBP, andneither PBP 4* nor 5* was produced eventhough the extracts had been prepared from cellsin which the membrane-bound amounts of bothof these sporulation-specific PBPs were increas-ing. This suggests that PBPs 4* and 5* are notderived by processing of any preexisting PBPsor pre-PBPs located in the membrane.Treatment of the membranes with an active
extract did generate two additional PBPs (indi-cated by the arrow in Fig. 3), which may berelated to one or more of the vegetative PBPs.PBP 1 is the most likely precursor of these twoproteins because it was the only PBP to be
A B C D
1n _ _ _
z-
4-i=
*5
FIG. 3. Fluorograph of the membrane-bound PBPsfrom B. subtilis cells harvested at to. The locations ofPBPs 4* and 5* are indicated, although neither proteinis present in the membranes at to. Lane A, untreatedsample; lane B, after exposure of the membranes to anextract of to cells for 60 min at 37°C; lane C, afterexposure to an extract of sporulating cells harvested att4; lane D, after exposure to an extract of spoOA cellsharvested at t4. Only the extract used on the sample inlane C had proteolytic activity. The arrow points to thenew PBP doublet in lane C. Approximately equalamounts of membrane protein were used for eachtreatment and the untreated control.
measurably affected by the extract, and some-times it appears as a doublet on gels (12).However, we have not yet eliminated the in-triguing possibility that these PBPs were gener-ated from other membrane proteins. These sametwo proteins were also produced by extracttreatment of membranes from the asporogenousspoOA mutant and membranes from sporulatingcells harvested at t6 (not shown). Neither proteinwas present in the extracts or in the membranesof the cells from which the extracts were pre-pared. In addition, neither protein was producedby treatment of membranes with extracts thathad no detectable proteolytic activity (i.e., ex-tracts of to cells and nonsporulating mutants)(Fig. 3).Thermal stability of the PBPs. Samples of
three membrane preparations each from vegeta-tive (to) and sporulating (t5) cells were incubatedat 50°C for up to 40 min before performing thestandard assay for their PBPs at 25°C. The timeat 50°C that reduced by 50% the amount oflabeled penicillin that could subsequently be
J. BACTERIOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STABILITY AND SYNTHESIS OF THE PBPs
bound was estimated from the inactivation curvefor each PBP (Table 1). PBPs 4* and 5 wereclearly the most thermostable, whereas some ofthe other proteins (PBPs 2A, 2B, 4, and 5*) wereat the opposite extreme. PBPs 1 and 3 werealways somewhat more stable than this lattergroup and were usually more stable in the vege-tative membranes than in the membranes fromsporulating cells.
DISCUSSIONAnalysis of the membrane-bound PBPs after
incubation of vegetative and sporulating cultureswith CAM established that protein synthesis isrequired for the appearance and accumulation ofsporulation-specific PBPs 4* and 5* as well asfor the sporulation-specific increases in vegeta-tive PBPs 2B and 3 (Fig. 2). These studies havealso revealed that the PBPs are not all equallystable in vivo, which helps to explain some ofthe quantitative changes in the PBP profile thatoccur during sporulation. For example, the ten-tative conclusion (20) that the decline in PBPs 1,2A, and 4 shortly after induction does not de-pend on an early sporulation event has beenstrengthened by the observations that these pro-teins appeared relatively unstable even undervegetative conditions, and CAM added at to hadlittle effect on their subsequent loss (Fig. 1). Incontrast, the loss of PBP 5 early in sporulationcannot be attributed to its intrinsic instability orto a preexisting inactivation mechanism becausethis PBP was quite stable in the presence ofCAM.There are at least two interpretations possible
for the observation that CAM prevented theincreases in PBPs 2B, 3, 4*, and 5*. The antibi-otic could have inhibited their de novo synthesisor it could have inhibited the synthesis of aprocessing enzyme that normally generates oneor more of these proteins from other PBPs orundetectable precursors during sporulation. Thedata do not support this latter interpretation. Aprevious quantitative analysis of PBP changes(20) demonstrated that modification of the PBPspresent at to could not account for the PBPincreases that occur after induction of sporula-tion. Consistent with this conclusion are thecurrent results from treatment of cell mem-branes with proteolytically active extracts ofsporulating cells (Fig. 3), which failed to pro-duce any of the four PIPs from proteins alreadypresent in the membrane. Furthermore, none ofthese PBPs was detectable in the soluble frac-tion of sporulating cells, which weakens theargument that the putative processing eventoccurred before insertion of the PBP into themembrane. The strongest evidence against theproteolytic origin of any PBP and in favor oftheir de novo synthesis, however, is the fact that
TABLE 1. Thermal stability of the PBPs
Half-life (min) at 50°CPBP
to sample t5 sample
1 -15 5-102A <1 <12B <1 <13 5-10 2-54 <1 <14* -405 >40 >405* <1
CAM immediately interrupted the increases inPBPs 2B, 3, 4*, and 5* when it was added afterthe amount of each had begun to rise (Fig. 2).The intracellular protease activity present at thetime the inhibitor was added was stable for theduration of the treatment.The membrane concentrations of PBPs 1, 2A,
4, and 5 began to drop from their to levels verysoon after induction of sporulation (Fig. 1), andlater the other PBPs also declined. We could notdistinguish inactivation from physical loss of thePBPs since currently the only practical methodfor detecting these proteins depends on theirpenicillin-binding activity. It is generally as-sumed that loss of penicillin-binding activityreflects loss of any other enzymatic activity ofthe PBP as well, because there is good evidencewith several PBPs that penicillin and substratesbind to the same active site of penicillin-sensi-tive enzymes (26, 27, 29, 30). Nevertheless, itwould be useful to know whether our inability todetect a PBP is the result of its inactivationwithin the membrane, its release from the mem-brane, or its degradation by proteases. Thestriking in vitro resistance (Fig. 3) of all thePBPs, except possibly PBP 1, to the majorprotease(s) induced during sporulation suggeststhat any degradation which does occur may bespecific rather than the result of generalizedproteolysis. A similar conclusion was reached inmuch the same way about the inactivation ofaspartate transcarbamylase during sporulationof B. subuilis (15, 25). There is more directevidence that some of the PBPs in Bacillusmegaterium are susceptible to limited proteoly-sis during sporulation; some of the fragmentsstill bind penicillin and are easily detected onfluorographs of radioactive penicillin-labeledmembranes (23). Such fragments are only rarelyobserved with B. subtilis preparations.Even though the process remains obscure, the
results presented here have helped to distinguishbetween those PBPs (PBPs 1, 2A, 2B, and 4)that are inactivated by preexisting mechanismsor conditions and those whose inactivation/lossdepends on protein synthesis during sporulation
VOL. 156, 1983 549
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

550 BUCHANAN AND SOWELL
(PBPs 3 and 5). Also, the results indicate that thechanges in PBPs 3 and 5 probably depend ondifferent cellular events, since the loss of PBP 5occurred early after induction, whereas PBP 3was stable until at least t2 (Fig. 1 and 2).
In an effort to relate the turnover of certainPBPs that occurred in both rich and nutrient-deficient media to some intrinsic property of theproteins, we examined the thermal stabilities ofall the membrane-bound PBPs. Those vegetativeproteins (PBPs 2A, 2B, and 4) that consistentlyappeared to be the most unstable in CAM-treated cultures were among the most thermola-bile in vitro (Table 1). In contrast, PBP 5 wasone of the most stable PBPs in vivo and was alsothe most resistant to thermal inactivation, with ahalf-life of greater than 40 min at 50°C. AfterPBP 5 is extracted from the membrane withdetergent, it has a half-life of only about 5 min at50°C (28), which suggests that its normal mem-brane association does more than simply pro-vide an anchor for correct orientation of theprotein with respect to its cell wall substrate.
In summary, the results of this study indicatethat the sporulation-specific increases in vegeta-tive PBPs 2B and 3 and the appearance andaccumulation of new PBPs 4* and 5* are depen-dent on concurrent protein synthesis duringsporulation of B. subtilis. In addition, analysis ofthe PBPs after treatment of cells with CAMrevealed that these proteins differ in their indi-vidual stabilities. In many cases, the in vivostability of a PBP was consistent with its thermallability in vitro. An effort to generate the sporu-lation-specific PBP changes in vitro by treatingmembranes with extracts of sporulating cells ledto the conclusion that none of the PBPs, exceptpossibly PBP 1, was subject to the generalizedproteolysis that occurs during sporulation.These results have helped to distinguish amongseveral possible explanations for the changes inthe PBP profile that occur in the early stages ofsporulation.
ACKNOWLEDGMENTSThis work was supported by National Science Foundation
grant PCM-7921673 and Public Health Service grant AI-19829from the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED1. Blumberg, P. M., and J. L. Strominger. 1974. Interaction
of penicillin with the bacterial cell: penicillin-bindingproteins and penicillin-sensitive enzymes. Bacteriol. Rev.38:291-335.
2. Bonner, W. M., and R. A. Laskey. 1974. A film detectionmethod for tritium-labeled proteins and nucleic acids inpolyacrylamide gels. Eur. J. Biochem. 46:83-88.
3. Buchanan, C. E. 1979. Altered membrane proteins in aminicell-producing mutant of Bacillus subtilis. J. Bacteri-ol. 139:305-307.
4. Buchanan, C. E. 1980. In vivo stability of the Escherichiacoli penicillin-binding proteins. FEMS Microbiol. Lett.7:253-256.
5. Buchanan, C. E., and J. L. Strominger. 1976. Alteredpenicillin-binding components in penicillin-resistant mu-tants of Bacillus subtilis. Proc. Natl. Acad. Sci. U.S.A.73:1816-1820.
6. Cooper, P. D. 1956. Site of action of radiopenicillin.Bacteriol. Rev. 20:28-48.
7. Dancer, B. N., and J. Mandelstam. 1975. Production andpossible function of serine protease during sporulation ofBacillus subtilis. J. Bacteriol. 121:406-410.
8. Hageman, J. H., and B. C. Carlton. 1973. Effects ofmutational loss of specific intracellular proteases on thesporulation of Bacillus subtilis. J. Bacteriol. 114:612-617.
9. Hitchins, A. D., and R. A. Slepecky. 1969. Antibioticinhibition of the septation stage in sporulation of Bacillusmegaterium. J. Bacteriol. 97:1513-1515.
10. Jenkinson, H. F., D. Kay, and J. Mandelatam. 1980.Temporal dissociation of late events in Bacillus subtilissporulation from expression of genes that determinethem. J. Bacteriol. 141:793-805.
11. Kerjan, P., E. Keryer, and J. Szulmajster. 1979. Charac-terization of a thermosensitive sporulation mutant ofBacillus subtilis affected in the structural gene of anintracellular protease. Eur. J. Biochem. 98:353-362.
12. Kleppe, G., and J. L. Strominger. 1979. Studies of thehigh molecular weight penicillin-binding proteins of Bacil-lus subtilis. J. Biol. Chem. 254:4856-4862.
13. Lawrence, P. J., M. Rogolsky, and V. T. Hanh. 1971.Binding of radioactive benzylpenicillin to sporulating Ba-cillus cultures: chemistry and fluctuations in specificbinding capacity. J. Bacteriol. 108:662-667.
14. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
15. Maurizi, M. R., J. S. Brabson, and R. L. Switzer. 1978.Immunochemical studies of the inactivation of aspartatetranscarbamylase by stationary phase Bacillus subtiliscells. J. Biol. Chem. 253:5585-5593.
16. Reysset, G., and J. Millet. 1972. Characterization of anintracellular protease in Bacillus subtilis during sporula-tion. Biochem. Biophys. Res. Commun. 49:328-334.
17. Rinderknecht, H., M. C. Geokas, P. Silverman, and B. J.Haverback. 1968. A new ultrasensitive method for thedetermination of proteolytic activity. Clin. Chim. Acta21:197-203.
18. Schaeffer, P. 1969. Sporulation and the production ofantibiotics, exoenzymes, and exotoxins. Bacteriol. Rev.33:48-71.
19. Schaeffer, P., J. Millet, and J.-P. Aubert. 1965. Catabolicrepression of bacterial sporulation. Proc. Natl. Acad. Sci.U.S.A. 54:704-711.
20. Sowell, M. O., and C. E. Buchanan. 1983. Changes inpenicillin-binding proteins during sporulation of Bacillussubtilis. J. Bacteriol. 153:1331-1337.
21. Spudkch, J. A., and A. Kornberg. 1968. Biochemical stud-ies of bacterial sporulation and germination VII. Proteinturnover during sporulation of Bacillus subtilis. J. Biol.Chem. 243:4600-4605.
22. Sterlini, J. M., and J. Mandelstam. 1969. Commitment tosporulation in Bacillus subtilis and its relationship todevelopment of actinomycin resistance. Biochem. J.113:29-37.
23. Todd, J. A., and D. J. EUar. 1982. Alteration in thepenicillin-binding profile of Bacillus megaterium duringsporulation. Nature (London) 300:640-643.
24. Vlnter, V. 1962. Spores of microorganisms. Penicillin-induced destruction of sporulating cells ofBacillus cereus.Experientia 18:409-410.
25. Wadndle, L. M., and R. L. Switzer. 1973. Inactivation ofaspartic transcarbamylase in sporulating Bacillus subtilis:demonstration of a requirement for metabolic energy. J.Bacteriol. 114:517-527.
26. Waxman, D. J., D. M. Lindgren, and J. L. Strominger.1981. High-molecular-weight penicillin-binding proteinsfrom membranes of bacilli. J. Bacteriol. 148:950-955.
27. Waxman, D. J., and J. L. Strominger. 1980. Sequence of
J. BACTERIOL.
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

STABILITY AND SYNTHESIS OF THE PBPs
active site peptides from the penicillin-sensitive D-alaninecarboxypeptidase of Bacillus subtilis. J. Biol. Chem.255:3964-3976.
28. Waxman, D. J., and J. L. Strominger. 1981. Limitedproteolysis of the penicillin-sensitive D-alanine carboxy-peptidase purified from Bacillus subtilis membranes. J.Biol. Chem. 256:2059-2066.
29. Yocum, R. R., H. Amanuma, T. A. O'Brien, D. J. Wax-
man, and J. L. Strominger. 1982. Penicillin is an active-site inhibitor for four genera of bacteria. J. Bacteriol.149:1150-1153.
30. Yocum, R. R., D. J. Waxman, R. R. Rasmussen, and J. L.Strominger. 1979. Mechanism of penicillin action: penicil-lin and substrate bind covalently to the same active serinein two bacterial D-alanine carboxypeptidases. Proc. Natl.Acad. Sci. U.S.A. 76:2730-2734.
VOL. 156, 1983 551
on Septem
ber 8, 2018 by guesthttp://jb.asm
.org/D
ownloaded from