status of proinflammatory and anti-inflammatory cytokines in different brain regions of a rat model...
TRANSCRIPT

ORIGINAL RESEARCH PAPER
Status of proinflammatory and anti-inflammatory cytokinesin different brain regions of a rat model of Japanese encephalitis
Ruchi Srivastava • Jayantee Kalita •
Mohammad Yahiya Khan • Usha Kant Misra
Received: 6 August 2011 / Revised: 19 December 2011 / Accepted: 19 December 2011 / Published online: 30 December 2011
� Springer Basel AG 2011
Abstract
Objectives This study evaluated cytokines and chemokine
in different regions of the rat brain at different time points
following Japanese encephalitis virus (JEV) infection.
Design Twelve-day-old Wistar rats were infected by
intracerebral inoculation of 3 9 106 plaque-forming units
of JEV 78668A strain. Expression of cytokines and che-
mokine was assayed using cytokine bead array in different
regions of the brain at 3, 6, 10 and 20 days post-inoculation
(dpi). Pathological changes including immunohistochem-
istry for JEV antigen were observed.
Results The cytokine levels were increased in the acute
stage and declined on follow-up. In the acute stage (6 dpi), the
levels of TNF-a, IFN-c, IL-6 and IL-10 were maximum in the
cortex, whereas the level of IL-4 was maximum in the stria-
tum. Lower levels of these cytokines were observed in
mid-brain and thalamus compared to other regions studied.
Maximum expression of JEV antigen and histopathological
changes as well as cytokines and chemokine were observed at
6 dpi in all the brain regions studied, but declined thereafter.
Conclusion This experimental study revealed maximum
expression of cytokines and chemokine at 6 dpi of JEV
infection which corresponded with histopathological
changes in different brain regions.
Keywords Viral encephalitis � JEV � Cytokine �Animal � Brain regions � Acute stage � Inflammation �Immunopathogenesis
Introduction
Japanese encephalitis (JE) is an important endemic
encephalitis prevalent in South-East Asia. About 50,000 JE
patients present annually; 15,000 of them die and half the
survivors live with major neurological sequelae [1]. The
clinical picture of JE is characterized by fever and altered
sensorium with or without seizure or focal neurological
deficit. The clinical manifestations of viral encephalitis are
determined by the location and severity of central nervous
system injury. Computed tomography scan and magnetic
resonance imaging study in 56 patients with JE revealed a
high frequency of involvement of thalamus (83%), basal
ganglia (46%), brain stem (35%) and cerebral cortex (23%)
[2]. These results are similar to autopsy studies [3, 4].
Inflammatory response associated with cell damage is
responsible for the clinical and histopathological changes
in viral encephalitis. Following JEV infection, overpro-
duction of free radicals by neurons in the acute phase of
illness has been reported [5]. Free radicals are general
mediators of signal transduction pathways, which are
responsible for cytokine production [6–8]. Cytokine levels
are surrogate markers of the inflammatory cascade in the
brain. However, an association between cytokine produc-
tion and free radical generation has not been reported in JE.
The role of inflammatory markers has been studied in viral
encephalitis patients and high levels of tumor necrosis factor
alpha (TNF-a) and interleukin (IL)-6 have been reported in
serum and cerebrospinal fluid [9–11]. In these studies,
however, the cytokine profile in different brain regions was
Responsible Editor: Graham Wallace.
R. Srivastava � J. Kalita � U. K. Misra (&)
Department of Neurology, Sanjay Gandhi Postgraduate Institute
of Medical Sciences, Raebareily Road, Lucknow 226014, India
e-mail: [email protected]; [email protected]
J. Kalita
e-mail: [email protected]
M. Y. Khan
Department of Biotechnology, Babasaheb Bhimrao Ambedkar
University, Lucknow, India
Inflamm. Res. (2012) 61:381–389
DOI 10.1007/s00011-011-0423-5 Inflammation Research
123

not possible. The severity of disease, its clinical manifesta-
tions and sequelae depend on the viral tropism. Such
information can only be obtained by autopsy or experimental
studies. Experimental studies in mice by Ghoshal and col-
leagues have reported microglial activation and expression
of a number of cytokines and free radicals which are
responsible for neuronal degeneration and death. They
reported changes in cytokine levels in cortex, hippocampus,
striatum and thalamus. In this study, involvement of the
midbrain, which is commonly involved in humans and is
responsible for various movement disorders, was not studied
[12]. The mouse model of JEV is short-lived, and therefore
the delayed effect of JEV cannot be studied. We have found a
rat model of JE to be more suitable for studying subacute and
chronic changes [5, 13]. Our study was undertaken to eval-
uate the cytokine and chemokine [monocyte chemotactic
protein-1 (MCP-1)] levels in different regions of the brain at
different time points and their relation to histopathological
changes following JEV infection.
Materials and methods
Animal
Twelve-day-old Wistar strain rats of either sex were used in
this study and were obtained from the animal house of the
Sanjay Gandhi Postgraduate Institute of Medical Sciences,
Lucknow, India. A maximum of five rats were kept in each
cage. The animals were maintained in an air-conditioned
room (25 ± 2�C) with 12-h light (7 a.m. to 7 p.m.) and dark
cycle. All the experiments were performed during the light
cycle, between 10 a.m. and 2 p.m., and were normalized in all
rats. Housing and procedures involving animals and their
care were conducted in accordance with the Principles of
Laboratory Animal Care (National Institutes of Health
publication number 86-23). The study was approved by the
local ethics committee, and all the experiments were carried
out in accordance with the institutional guidelines on the care
and use of experimental animals.
Virus
The JEV 78668A strain which was used was isolated from
the post-mortem brain tissue of a 38-year-old woman
patient who died during an epidemic of encephalitis in
Gorakhpur, India in 1978 [14]. The genome of GP78 is
10,976 nucleotides long. An open reading frame of 10,296
bases, capable of coding 3,432 amino acid polyproteins, is
flanked by 95- and 585-base-long 50- and 30-non-coding
regions, respectively [15]. The JEV was propagated in
3–4 days old suckling mice brain. After 4 days of infec-
tion, the rats were killed. Brain was removed aseptically,
homogenized in sterile phosphate-buffered saline (PBS)
and centrifuged at 160609g for 30 min at 8�C. The
supernatant was collected, aliquoted and stored at -70�C
until further use. The virus was plaque-purified twice on
monolayers of porcine stable kidney (PS) cells and the
isolate was subsequently referred to as the GP78 strain.
Infection of rats
The rats were divided into experimental and control groups. In
the experimental group, rats were inoculated intracerebrally
with 3 9 106 plaque-forming units of JEV according to a
fixed protocol [13]. The inoculation was done at the midpoint
of the line connecting the ears. Control rats were inoculated
with sterile PBS (19, Sigma). The rats were killed at 3, 6, 10,
and 20 days post-inoculation (dpi). Their brains were excised
aseptically and the regions of interest such as cortex, striatum,
thalamus and mid-brain were dissected using the standard
procedure, and cryopreserved for cytokine and chemokine
assay by cytometric bead array (CBA).
Histopathological studies
The rats were anaesthetized with ether and perfused with
150 ml of 0.1 M, pH 7.2, PBS followed by 250 ml of ice-
cold 4% paraformaldehyde in PBS for fixation of tissues.
The brains were removed and preserved in 10% parafor-
maldehyde. Serial coronal sections of 5-lm thickness were
cut on a freezing microtome (Slee Mainz Co., Germany).
The sections were then transferred to gelatinized slides and
stained with haematoxylin and eosin (H&E).
Immunohistochemical studies
The rats were anaesthetized with ether and perfused with
150 ml of 0.1 M, pH 7.2, PBS followed by 250 ml of ice-
cold 4% paraformaldehyde in PBS for fixation of tissues.
The brains were removed and postfixed in the same fixative
overnight, followed by cryopreservation in 10, 20, and 30%
(w/v) sucrose in PBS. Five serial coronal sections of 20-lm
thickness were cut on a freezing microtome (Slee Mainz
Co., Germany). These tissue sections were treated with
0.3% H2O2–methanol to suppress endogenous peroxidase
activity followed by 10% normal goat serum and allowed
to react with primary antibody (anti-mouse JEV antibody,
1:100, obtained from National Institute of Virology, Pune,
India) at 4�C overnight. After removing the primary anti-
body, sections were washed three times with PBS and
incubated in biotinylated peroxidase-linked secondary
antibody (1:200) for 2 h at room temperature followed by
three washes with PBS. Color was developed for peroxi-
dase-linked antibody with 3,3-diaminobenzidine as
chromogen. Sections were dehydrated in a graded series of
382 R. Srivastava et al.
123

alcohol followed by xylene, mounted in DPX, cover-slip-
ped, and then examined under the microscope.
Protein extraction and quantitation
Snap-frozen brain tissues were homogenized in 19 PBS
containing proteinase inhibitor (10 ml ice-cold 19 PBS ?
5 ll proteinase inhibitor) chilled buffer using a glass
homogenizer under ice. Typically, each sample was
homogenized in 700 ll of buffer, followed by sonication
on ice for 30 s and centrifugation at 136809g for 30 min at
4�C [16]. Supernatant was collected, aliquoted and stored
at -80�C for further use in CBA after protein quantitation
by the standard Lowry assay [17].
Cytometric bead array
Cytometric bead array (CBA, rat flex kit; BD Biosciences,
Singapore) was used to quantitatively measure cytokine
expression levels in brain tissue lysates of different brain
regions (cortex, striatum, thalamus and mid-brain) of the
control and JEV-infected rats. The assay was performed
according to the manufacturer’s instructions and analyzed
on an FACS Calibur (Becton–Dickinson, UK). Analysis
was performed using CBA software (FCAP Array v1.01)
which allows the calculation of cytokine concentrations in
unknown samples [12].
Enzyme-linked immunosorbent assay (ELISA)
To examine the MCP-1 level in the tissue lysates of
different brain regions (cortex, striatum, thalamus and mid-
brain) of control and JEV-infected rats, ELISA was per-
formed. A commercially available kit from BD Biosciences
was used according to the manufacturer’s guidelines.
Statistical analysis
All the statistical analyses were done using GraphPad
Prism (v3.03) software. Mean differences in more than two
groups were analyzed using one-way analysis of variance
(ANOVA) followed by Newman–Keul’s multiple com-
parison test. The variable was considered significant if
p \ 0.05. Data were expressed as mean ± SD.
Results
Forty-five rats were included in the study group and six in
the control group.
Clinical signs
The clinical signs were monitored daily up to 20 dpi. The
clinical signs appeared from 4 dpi in the study group. All
the rats showed hypokinesia, pelvic elevation and hind
limb disability which was observed up to 20 dpi. However,
tremors and convulsions were not frequently observed.
Mortality was observed from 6 to 10 dpi; none of the rats
died after 10 dpi. The control group did not exhibit any of
these clinical symptoms (Table 1).
Histopathological findings
The histopathological features of JE such as inflammatory
cell infiltration, necrosis and neuronal shrinkage were
observed in the JEV-inoculated rats (Fig. 1). These
pathological changes were not present in the control rats.
Three rats in each group were used for the histopatholo-
gical study.
Immunohistochemical study
JEV antigen was present in different brain regions such as
thalamus, mid-brain, striatum and cortex (representative
photomicrographs are shown in Fig. 2). On the basis of
visual perception, the intensity of JEV antigen was noted
and is presented in Table 2. The intensity of JEV antigen
was maximum at 6 dpi in all the brain region and gradually
declined up to 20 dpi.
Table 1 Clinical signs and
symptoms and mortality in
12-day-old rats challenged with
3 9 106 pfu of JEV
Numbers in parenthesesindicate the number of rats
? denotes present; - denotes
absent; ± denotes poor or
inconsistent
pfu plaque-forming units,dpi days post-inoculation
Dpi Clinical signs Mortality
Hypokinesia Pelvic
elevation
Hind limb
disability
Tremor Convulsions
4 (45) ? (45) ± (45) ± (45) - - -
6 (41) ? (41) ? (41) ? (41) ? (7) ? (5) ? (4)
7 (35) ? (35) ? (35) ? (35) ? (7) ? (5) ? (6)
9 (32) ? (32) ? (32) ? (32) ? (4) ? (3) ? (3)
10 (30) ? (30) ? (30) ? (30) - - ? (2)
15 (30) ? (30) ? (30) ? (30) - - -
20 (30) ? (30) ? (30) ? (30) - - -
Cytokines in different brain regions of JE infected rat 383
123

Cytokines and chemokine levels in different brain
regions
Levels of cytokines and MCP-1 in cortex
The cytokine and chemokine levels were highest at 6 dpi
and lowest at 20 dpi in the cortex of rat brain after JEV
infection. Levels of IFN-c reached maximum at 6 dpi but
gradually declined significantly at 10 dpi and further at
20 dpi when compared between the study groups
(Fig. 3a). Similar trends were observed for IL-6, IL-10,
TNF-a and MCP-1 (Fig. 3a–e). For IL-4, a different
pattern was observed. There was insignificant increase in
the level of IL-4 at 6 dpi compared to control. However,
there was a significant decrease in levels at 10 dpi and
20 dpi, like other cytokines, but the level at 20 dpi was
also significantly lower compared to control (p \ 0.001)
(Fig. 3b).
Fig. 1 Photomicrograph showing H&E staining in different brain
regions at different days post-inoculation in 12-day-old rats inocu-
lated with 3 9 106 pfu/ml of JEV. a control rat, b neuronal shrinkage
in cortical region (broken arrow shows shrunken neuron, arrowindicates normal neuron), c inflammatory cells infiltration with
necrosis in the cortex (arrow)
Fig. 2 Representative photomicrograph showing presence of JEV antigen in cortex, striatum, thalamus and mid-brain in 12-day-old rats
inoculated with 3 9 106 pfu/ml of JEV solution
384 R. Srivastava et al.
123

Levels of cytokines and MCP-1 in striatum
The striatum of JEV-infected rat brain showed the maxi-
mum level of all cytokines and chemokine at 6 dpi. There
was a significant increase in IFN-c, IL-6, IL-10, TNF-a and
MCP-1 levels at 3, 6, 10 and 20 dpi compared to controls.
These levels declined at 10 dpi and further at 20 dpi when
compared between the groups. IL-4 levels, however, were
not significantly different between the experimental and the
control groups (Fig. 4).
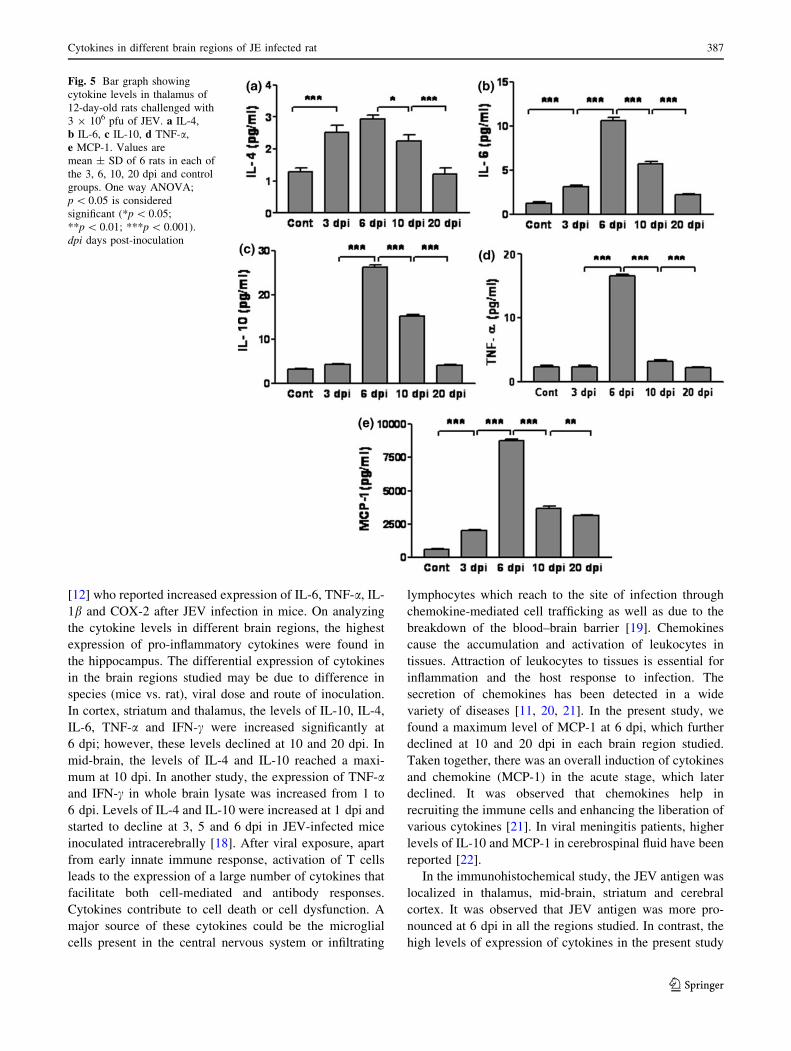
Levels of cytokines and MCP-1 in thalamus
Expression patterns of different cytokines and chemokine
in the thalamus of JEV-infected rats were similar to the
expression patterns in the brain regions described above.
Levels of all cytokines and chemokine reached a maximum
at 6 dpi in the thalamus. These levels significantly declined
at 10 and 20 dpi when compared to 6 dpi. Interestingly, we
were not able to detect IFN-c in the thalamus of JEV-
infected rats at any time point (Fig. 5).
Table 2 Presence of JEV antigen in different brain regions in 12-day-old rats challenged with 3 9 106 pfu of JEV at 3 and 6 dpi
JEV staining 3 dpi 6 dpi 10 dpi 20 dpi
St Co MB Th St Co MB Th St Co MB Th St Co MB Th
? ? ? ? ??? ??? ??? ??? ? ? ? ? ± ± ± ±
? denotes low, ?? denotes moderate, ??? denotes high, ± denotes faint staining
pfu plaque-forming units, dpi days post-inoculation, St striatum, Co cortex, MB mid-brain, Th thalamus
Fig. 3 Bar graph showing
cytokines and chemokine levels
in cortex of 12-day-old rats
challenged with 3 9 106 pfu of
JEV. a IFN-c, b IL-4, c IL-6,
d IL-10, e TNF-a, f MCP-1.
Values are mean ± SD of 6 rats
in each of the 3, 6, 10, 20 dpi
and control groups. One way
ANOVA; p \ 0.05 is
considered significant
(*p \ 0.05, **p \ 0.01,
***p \ 0.001). dpi days
post-inoculation
Cytokines in different brain regions of JE infected rat 385
123

Levels of cytokines and MCP-1 in mid-brain
The mid-brain of JEV-infected rats showed different
expression patterns for cytokines and chemokine compared
to patterns in cortex, striatum and thalamus. Levels of
IFN-c, IL-6, TNF-a and MCP-1 were found to be maxi-
mum at 6 dpi in the mid-brain; however, the level of IL-10
reached a maximum at 10 dpi. A significant increase in
levels of IL-6, IL-10, TNF-a and MCP-1 compared to
controls was noted at 3, 6, 10 and 20 dpi (p \ 0.001).
Levels of IFN-c increased significantly compared to
controls only at 6 dpi (p \ 0.001). IL-4 levels were sig-
nificantly decreased at 3 dpi (p \ 0.05); however, no
significant changes were observed at 6, 10 and 20 dpi
compared to controls. IFN-c was increased significantly at
6 dpi compared to 3 dpi (p \ 0.01), and significantly
declined at 10 dpi compared to 6 dpi (p \ 0.001). TNF-aand MCP-1 levels were reduced significantly at 10 dpi
compared to 6 dpi (p \ 0.001). An insignificant increase
was noted in IL-10 level from 6 to 10 dpi (Fig. 6).
Discussion
In the present study, we observed increased levels of pro-
and anti-inflammatory cytokines in the different brain
regions of rats after JEV infection, compared to controls.
These cytokine levels varied from the early (3 to 10 dpi) to
the late (20 dpi) phases of infection. We used the rat model
of JE rather than the mouse model because the life span of
rats is longer; thereby, it is feasible to study the clinical
sequence and its underlying pathophysiological mecha-
nisms. We used younger rats because of greater
susceptibility and longer survival of younger rats has been
reported compared to older ones following intracerebral
inoculation of JEV [18].
In the acute stage (at 6 dpi), the maximum levels of
TNF-a, IFN-c, IL-6 and IL-10 were observed in the cortex.
However, the level of IL-4 was maximum in striatum.
Lower levels of these cytokines were observed in mid-
brain and thalamus compared to the other studied regions.
Our results are in agreement with those of Ghoshal et al.
Fig. 4 Bar graph showing
cytokine levels in striatum of
12-day-old rats challenged with
3 9 106 pfu of JEV. a IFN-c,
b IL-4, c IL-6, d IL-10,
e TNF-a, f MCP-1. Values are
mean ± SD of 6 rats in each of
the 3, 6, 10, 20 dpi and control
groups. One way ANOVA;
p \ 0.05 is considered
significant (*p \ 0.05;
**p \ 0.01; ***p \ 0.001).
dpi days post-inoculation
386 R. Srivastava et al.
123
[12] who reported increased expression of IL-6, TNF-a, IL-
1b and COX-2 after JEV infection in mice. On analyzing
the cytokine levels in different brain regions, the highest
expression of pro-inflammatory cytokines were found in
the hippocampus. The differential expression of cytokines
in the brain regions studied may be due to difference in
species (mice vs. rat), viral dose and route of inoculation.
In cortex, striatum and thalamus, the levels of IL-10, IL-4,
IL-6, TNF-a and IFN-c were increased significantly at
6 dpi; however, these levels declined at 10 and 20 dpi. In
mid-brain, the levels of IL-4 and IL-10 reached a maxi-
mum at 10 dpi. In another study, the expression of TNF-aand IFN-c in whole brain lysate was increased from 1 to
6 dpi. Levels of IL-4 and IL-10 were increased at 1 dpi and
started to decline at 3, 5 and 6 dpi in JEV-infected mice
inoculated intracerebrally [18]. After viral exposure, apart
from early innate immune response, activation of T cells
leads to the expression of a large number of cytokines that
facilitate both cell-mediated and antibody responses.
Cytokines contribute to cell death or cell dysfunction. A
major source of these cytokines could be the microglial
cells present in the central nervous system or infiltrating
lymphocytes which reach to the site of infection through
chemokine-mediated cell trafficking as well as due to the
breakdown of the blood–brain barrier [19]. Chemokines
cause the accumulation and activation of leukocytes in
tissues. Attraction of leukocytes to tissues is essential for
inflammation and the host response to infection. The
secretion of chemokines has been detected in a wide
variety of diseases [11, 20, 21]. In the present study, we
found a maximum level of MCP-1 at 6 dpi, which further
declined at 10 and 20 dpi in each brain region studied.
Taken together, there was an overall induction of cytokines
and chemokine (MCP-1) in the acute stage, which later
declined. It was observed that chemokines help in
recruiting the immune cells and enhancing the liberation of
various cytokines [21]. In viral meningitis patients, higher
levels of IL-10 and MCP-1 in cerebrospinal fluid have been
reported [22].
In the immunohistochemical study, the JEV antigen was
localized in thalamus, mid-brain, striatum and cerebral
cortex. It was observed that JEV antigen was more pro-
nounced at 6 dpi in all the regions studied. In contrast, the
high levels of expression of cytokines in the present study
Fig. 5 Bar graph showing
cytokine levels in thalamus of
12-day-old rats challenged with
3 9 106 pfu of JEV. a IL-4,
b IL-6, c IL-10, d TNF-a,
e MCP-1. Values are
mean ± SD of 6 rats in each of
the 3, 6, 10, 20 dpi and control
groups. One way ANOVA;
p \ 0.05 is considered
significant (*p \ 0.05;
**p \ 0.01; ***p \ 0.001).
dpi days post-inoculation
Cytokines in different brain regions of JE infected rat 387
123

was observed in those areas where the viral load and the
presence of viral antigen was low, suggesting the possible
role of cytokines in viral clearance. These findings may
suggest immune-mediated pathogenesis, i.e. the host
immune response may kill the infected cells in an effort to
clear the virus. In our earlier study, we reported the his-
topathological changes in thalamus, mid-brain, striatum
and cortex, in which maximum damage evidenced by
neuronal shrinkage, cellular infiltration and inflammatory
cells was observed at 6 dpi [13]. The extent of cellular
damage could be associated with the viral load and the
presence of JEV antigen in different brain regions.
In the present study, IFN-c was not detectable in thalamus.
The antiviral effect of IFN-c has been reported in West Nile
virus infection [23]. As part of innate immunity, IFN-c is
produced in response to viral infections. Instead of being
directly antiviral, it induces the production of effector pro-
teins in the cells, which inhibits various stages of viral
replication, assembly, or release. We hypothesize that the
absence of IFN-c might contribute to the vulnerability of the
thalamic region to viral infection. These observations are
supported by the high frequency of thalamic involvement in
JE as seen on magnetic resonance imaging [2].
Besides the proinflammatory cytokines, anti-inflamma-
tory cytokines are also expressed simultaneously to balance
the action of proinflammatory cytokines. The highest levels
of IL-10 and IL-4, which are immunosuppressive cyto-
kines, were noted at 6 dpi. It might be possible that the
expression of these cytokines would suppresses the
immune response and contributes to the recovery. The role
of the immune response in recovery from JEV infection is
poorly understood. The decline in cytokine levels at 10 and
20 dpi may be due to an associated immune response and
neutralizing antibodies. Early host defence against JEV
infections is mediated by phagocytic cells, followed by a
complex mechanism which involves B and T effector cells
[14, 24, 25]. The reciprocal relationship between cytokine
and virus load raises an important question of whether the
activation of cytokines results in viral proliferation and cell
damage, or absence of cytokines at a particular site results
in more viral load. We found different levels of cytokines
in different brain regions (cortex, striatum, thalamus and
mid-brain) of JEV-infected rat at different time points.
In our earlier study, we observed an increase in free
radical generation in the initial phase of JEV infection,
which declined at a later stage. Maximum levels of
Fig. 6 Bar graph showing
cytokine levels in mid-brain of
12-day-old rats challenged with
3 9 106 pfu JEV. a IFN-c,
b IL-4, c IL-6, d IL-10,
e TNF-a, f MCP-1. Values are
mean ± SD of 6 rats in each of
the 3, 6, 10, 20 dpi and control
groups. One way ANOVA;
p \ 0.05 is considered
significant (*p \ 0.05;
**p \ 0.01; ***p \ 0.001).
dpi days post-inoculation
388 R. Srivastava et al.
123

neuronal reactive oxygen species and peroxinitrite were
observed at 6 dpi and declined significantly at 10 and
20 dpi as compared to control [5]. In the present study, we
noted the same pattern of expression of cytokines, which
peaked at 6 dpi and declined by 20 dpi. Many studies
reported an association of free radical generation with
cytokine production [26, 27]. Free radicals are general
mediators of signal transduction pathways which are able
to induce cytokine production [6–8]. In JE, free radicals
may also act as a signal for the release of different cyto-
kines. A detailed study, however, is needed to prove this
hypothesis.
This study highlights that the cytokines are expressed
maximally during the acute phase of JEV infection, which
is associated with histological changes, and decline sub-
sequently. Confirmation of these results may help in the
treatment of JE to reduce cellular damage.
Acknowledgments We acknowledge the Indian Council of Medical
Research for financial support to Ruchi Srivastava as Senior Research
Fellow, and Council of Science and Technology, Uttar Pradesh, India
for establishing the Encephalitis Research Center. We thank Mr.
Rakesh Kumar Nigam for secretarial help.
References
1. Tabatabaie T, Vasquez-Weldon A, Moore DR, Kotake Y. Free
radicals and the pathogenesis of type 1 diabetes: beta-cell cyto-
kine-mediated free radical generation via cyclooxygenase-2.
Diabetes. 2003;52:1994–9.
2. Kalita J, Misra UK, Pandey S, Dhole TN. A comparison of
clinical and radiological findings in adults and children with
Japanese encephalitis. Arch Neurol. 2003;60:1760–4.
3. Saxena V, Mathur A, Krishnani N, Dhole TN. An insufficient
anti-inflammatory cytokine response in mice brain is associated
with increased tissue pathology and viral load during Japanese
encephalitis virus infection. Arch Virol. 2008;153:283–92.
4. Wang SM, Lei HY, Yu CK, Wang JR, Su IJ, Liu CC. Acute
chemokine response in the blood and cerebrospinal fluid of
children with enterovirus 71-associated brainstem encephalitis.
J Infect Dis. 2008;198:1002–6.
5. Shrestha B, Wang T, Samuel MA, Whitby K, Craft J, Fikrig E,
Diamond MS. Gamma interferon plays a crucial early antiviral
role in protection against West Nile Virus infection. J Virol.
2006;80:5338–48.
6. Ali MH, Schlidt SA, Chandel NS, Hynes KL, Schumacker PT,
Gewertz BL. Endothelial permeability and IL-6 production dur-
ing hypoxia: role of ROS in signal transduction. Am J Physiol.
1999;277:L1057–65.
7. Haddad JJ, Safieh-Garabedian B, Saade NE, Land SC. Thiol
regulation of pro-inflammatory cytokines reveals a novel immu-
nopharmacological potential of glutathione in the alveolar
epithelium. J Pharmacol Exp Ther. 2001;296:996–1005.
8. Kosmidou I, Vassilakopoulos T, Xagorari A, Zakynthinos S,
Papapetropoulos A, Roussos C. Production of interleukin-6 by
skeletal myotubes: role of reactive oxygen species. Am J Respir
Cell Mol Biol. 2002;26:587–93.
9. Burke DS, Morill JC. Levels of interferon in the plasma and
cerebrospinal fluid of patients with acute Japanese encephalitis.
J Infect Dis. 1987;155:797–9.
10. Mathur A, Khanna N, Chaturvedi UC. Breakdown of blood-brain
barrier by virus induced cytokine during Japanese Encephalitis
Virus infection. Int J Exp Pathol. 1992;73:603–11.
11. Wakefield TW, Strieter RM, Wilke CA, Kadell AM, Wrobleski
SK, Burdick MD, Schmidt R, Kunkel SL, Greenfield LJ. Venous
thrombosis-associated inflammation and attenuation with neu-
tralizing antibodies to cytokines and adhesion molecules.
Arterioscler Thromb Vasc Biol. 1995;15:258–68.
12. Ghoshal A, Das S, Ghosh S, Mishra MK, Sharma V, Koli P, Sen
E, Basu A. Proinflammatory mediators released by activated
microglia induces neuronal death in Japanese encephalitis. Glia.
2007;55:483–96.
13. Kumar S, Kalita J, Saxena V, Khan MY, Khanna VK, Sharma S,
Dhole TN, Misra UK. Some observations on the tropism of
Japanese encephalitis virus in the rat brain. Brain Res.
2009;1268:135–41.
14. Mathur A, Chaturvedi UC, Tandon HO, Agarwal AK, Mathur
GP, Nag D, Prasad A, Mittal VP. Japanese encephalitis epidemic
in Uttar Pradesh, India during 1978. Indian J Med Res.
1982;75:161–9.
15. Vrati S, Giri RK, Razdan A, Malik P. Complete nucleotide
sequence of an Indian strain of Japanese encephalitis virus:
sequence comparison with other strains and phylogenetic analy-
sis. Am J Trop Med Hyg. 1999;61:677–80.
16. Vilela MC, Mansur DS, Lacerda-Queiroz N, Rodrigues DH,
Lima GK, Arantes RM, Kroon EG, da Silva Campos MA,
Teixeira MM, Teixeira AL. The chemokine CCL5 is essential for
leukocyte recruitment in a model of severe Herpes simplex
encephalitis. Ann N Y Acad Sci. 2009;1153:256–63.
17. Lowry OH, Rosebrough NH, Farr AL, RJ Randall. Protein
measurement with the Folin phenol reagent. Biol Chem.
1951;193:265–75.
18. Ogata A, Nagashima K, Hall WW, Ichikawa M, Kuroda JK,
Yasui K. Japanese encephalitis virus neurotropism is dependent
on the degree of neuronal maturity. J Virol. 1991;65:880–6.
19. Mathur A, Arora KL, Chaturvedi UC. Host defense mechanisms
against Japanese encephalitis virus infection in mice. J Gen Virol.
1983;64:805–11.
20. Tsai TF. New initiatives for the control of Japanese encephalitis
by vaccination: minutes of a WHO/CVI meeting, Bangkok,
Thailand. Vaccine. 1998;18:1–25.
21. Vilela MC, Mansur DS, Lacerda-Queiroz N, Rodrigues DH,
Arantes RM, Kroon EG, Campos MA, Teixeira MM, Teixeira
AL. Traffic of leukocytes in the central nervous system is asso-
ciated with chemokine up-regulation in a severe model of herpes
simplex encephalitis: an intravital microscopy study. Neurosci
Lett. 2008;445:18–22.
22. Lahrtz F, Piali L, Spanaus KS, Seebach J, Fontana A. Chemo-
kines and chemotaxis of leukocytes in infectious meningitis.
J Neuroimmunol. 1998;85:33–43.
23. Shankar SK, Rao TV, Mruthyunjayanna BP, Devi MG. Autopsy
study of brains during an epidemic of Japanese encephalitis in
Karnataka. Indian J Med Res. 1983;78:431–40.
24. Levine B, Hardwick JM, Trapp BD, Crawford TO, Bollinger RC,
Griffin DE. Antibody-mediated clearance of alphavirus infection
from neurons. Science. 1991;254:856–60.
25. Srivastava R, Kalita J, Khan MY, Misra UK. Free radical gen-
eration by neurons in rat model of Japanese encephalitis.
Neurochem Res. 2009;34:2141–6.
26. Srivastava S, Khanna N, Saxena SK, Singh A, Mathur A, Dhole
TN. Degradation of Japanese encephalitis virus by neutrophils.
Int J Exp Pathol. 1999;80:17–24.
27. Makay B, Makay O, Yenisey C, Icoz G, Ozgen G, Unsal E,
Akyildiz M, Yetkin E. The interaction of oxidative stress
response with cytokines in the thyrotoxic rat: is there a link?
Mediators Inflamm. 2009;2009:391682.
Cytokines in different brain regions of JE infected rat 389
123