stimulation of human cytotoxic t cells with hiv-1 -derived...
TRANSCRIPT

lnternatimal lmmunology, W. 9, No. 3, pp. Wffi
Stimulation of human cytotoxic T cells withHIV-1 -derived peptides presented byrecombinant HLA-A2 peptide complexes
Jrirgen B. Watter, Christian Branderl, Mathai Mammen2, David N. Garboczi3,Spy-ros A. Kalamsl, George M. Whitesides2, Bruce D. Walkerl andHerman N. Eisen
Center for Cancer Research and Department of Biology, Massachusetts lnstrtute of Technology,Cambridge, MA 02139, USAltnfect ious Disease Unit , Massachusetts Generai Hosprtal and Harvard Medical School , Boston, MA0 2 1 1 4 , U S AzDepartment of Chemistry and Chemrcal Brology, Harvard Universi ty, Cambridge, MA02138, USA3Department of Moleculai and Cel lu lar Bioiogy, Harvard Universrty, Cambrrdge, MA 02138, USA
Keywords: cytotoxic T cells, HlV, HL-A-A2
Abstract
HLA-A2 heavy chain and fo-microglobulin were expressed In Escherichla coll, and refolded In the
presence of peptldes derived from HIV-1 RT and gag proteins. When recomblnant HLA-A2
molecules were at tached to cel ls lacking Hl A-A2, the cel ls became suscept ib le to lysis by HLA-A2
restr icted cytotoxic T lymphocyte (CTL) c lones speci f lc for pept ldes der ived from RT and gag
proteins. L imit ing di lut lon analyses of per ipheral b lood mononuclear cel ls f rom HIV-1- lnfected
lndiv iduals showed that the recombinant HLA-A2 pept lde complexes covalent ly immobi l ized on
microspheres stimulated the development of HLA-A2 peptide-specific CTL. Preformed HtA-peptide
complexes may provide an al ternat ive to immunizat ion procedures that depend upon intracel lu larprocessing of ant igen to el ic i t T cel l responses-
lntroduct ion
MHC-encoded class I molecules serve as pept lde binding,transport and display proterns on the cer l sunace, and can
evoke T cet l responses when recogntzeo bv ant lgen-soecrf lcreceptors on cytotoxrc T lymphocytes (CTL). l '"4HC class Imolecules are glycoproteins constst tng oi f 'vo non-covalent lyassoclated polypeptrde chains, a larger or heavy chatn and
a smal le r o r l igh t cha in , te rmed p2-mrcroQlobu l rn (1 ) . Human
class I molecules have been refolded f rom suounrts proouced
in Escherrchra col i in associatron wrth syntnet lc pepttdes
(2). The three-dimenstOnal StrUctures of severat recombtnant
complexes, determined by X-ray crystal lography (3), are
vrr tual ly ident ical to the Structure of the c laSS I molecule
isolated from human cel ls (4). However the MHC protelns
produced in E. col ido not contatn the ccmprex carbonyoratethat is normal ly at tached to asoaragrne 86. The aim of thtsr to rk i s to de termtne l f an MHC c lass I o ro ie rn , speCt f i ca l l yHLA-A2 (A2), produced in non-glycosylateo form rn E. colr
and assembled w i th svn the t ic nonapeot loes can ( i ) e l i c t t
@ 1996 Oxfotd Uniwrsitv Press
cytolyttc responses by CDB* rT cell clones arising tn thecourse of natural infect ion wrth HIV-1 and ( i i ) st imulate lheprolr feratron of pept ide-specrf ic CDB* T cel ls in perrpheral
blooo lymphocytes of HIV-1 seropostt tve donors.The HLA-A2 (HLA-A'0201) heavy chain and p2-mrcro-
globul in l ight chain v{ere assembled in the presence of fwoanttgenrc pept ides ber ived l rom HIV- ' l : a pQtide from reversetranscr lptase (RT-1V9, amrno actds 4THB4,ILKEPVHGV) (5)and a peptrde from the gag protein (Gag-SL9, amrno acros7745, SLYNTVATL) (6) Both peptrdes represent diseaseresponse targets in HLA-A2* HIV-1- infected pat ients (7),
conform to the H[-A-A2 consensus motif (8), and have theoptrmal sequence recognized by RT-|V9- 'and Gag-SL9-specrf ic CTL clones obtained from asymptomdtic HIV-1-sero-posrtrve pat ients (9,10). Both pept ides are recogntzed bycytotoxic T cel l c lones at ptcomolar concentrat ions, wi th SD.6values (r .e. peptrde concentrat ion givrng hal f -maximal specrf rclysis) of -3 pM for Gag-SL9 and -0 1 pM for Ff f - lV9 (5,9)
Corresponclence to: J. B. Walter
Transmrfttne edtor. D. Y. Loh Recetved 19 Seotember 1996, accepted 5 December 1996

102 lnduction of CTL responses by recombinant HU-42 peptide complexes
V V V
(v
Log I r l uo rescence In tens i t y
Flg. 2. Adsorptron of purrf ied soluble H[-A-AZ peptide complexes tothe surface of Ht-A-A2-negaltve cells monrtored by FACS anatysrs.MHC class I expressron on unmoorf ied HLA-A2-negative C1R cetts(HLA-8,C) was determtned wirh mAb 887.7-FITC conjugate (doneoline, 887.7 antr.HLA-A,B,C). To adsorb the Ht-A+€ptide comptexeswe covalently anached mAb BBMl (antr-pTmicroglobul in) and mAbMA2.1 (antr-HLA-A2,B 1 7) to the cel l sur{ace using a heterobifunctronalcross-lrnker (SPDP). Sudace bound mAb-SPDP conjugates weredetected using FITC-goat anti-mouse F(ab)r fragments (dasheC l ine)Adsorption of HLA-A2 peptrde complexes to mAb-coated cetts wasshown by an increase in MHC class I expression using mAb 887.7-FITC conlugate (sol id l ine).
mAb W6/32 (11) ( lgG2a specrf ic for HI-A-A,B,C) and PA2.1(12) ( lgG1 specr i ic for HLA-M ano A2B), both reactrve wrrhnatrve Ht-A-A2 complexes, wts detected with goat antr-mouselg conjugated to horseradish peroxrdase (1 :5000: Amersham)After washing the ni t rocei lu lose was incubated for 1 mrn wrthreagents for ECL immunodetectron (1:1; Amersham) andexposeo to X-ray fi lm. HLA-A2 peptrde concentrations weremeasureo by mrcro-BCA (bic inchonrntc acid) assay (Pierce,Rockford, lL) , uslng BSA as a stanoard:
Covalent attachment of mAb to the cell sur't'ace
mAb BBMl (13) (an t r - p2-mrcrog iobu i ln ) and mAb MA2.1 (14)(antr-HLA-A2, 817) were covalent ly at tached to cel ls usrng aheterobi funct lonal cross- l inker, SPDP IMsuqcinimidyl 3-(2-pyr idyldi th io)propionate, Piercel , as descr ibcd (15). SPDP(50 pl, 20 mM in ethanol) was added to mAb in PBS, pti 7 2,(1 mg/ml) for 30 min at room temoerature and then dialyzedagainst PBS Cel ls (106) were reduceo with 50 pM di th iothrertolfor 30 mrn, washed twrce with P8S and incubated wrth 200pl of mAb-SPDP at 0 5 mg/ml (1 h at room temperature),wasned and sublected to FACS analvsis.
Adsorption of soluble A2-pepttde complexes to mAb pre-coated cells
Cel ls (106) wi th covalent ly at tacheo mAb (using SPDP anddi th iothrei to l , see above) were incubated with 10 pg solubleHLA-A2 peptide complexes in 500 pl PBS for 30 min at 4'C,washed in 10 ml of cold RPMI, counted and adjusted to theappropnate cell number.
o!
oo
L 9
oc r . i
r \
cot^l
l 5 0
m A b W 6 / 3 :
m A b P A l . I
Flg. 1. Gel lr itratron HPLC profi les of HI-A-A2 peptrde complexes.HPLC prolries of Ht-A-M reconstttutton lrom recombrnant healy charnand p2-mrcroglobuiln in associatron wrth (a) HIV-1 Gag-SL9 peptrde77-€5, SLYNTVATL and (b) HIV-'l RT-lvg peptrde 47G-484,ILKEPVHGV Specrfic brnding of mAb W6/32 and mAb PA2.1 (bothspecrlically reactrve with nattve HLA-A2 complexes)to HPLC fractionswas used to monitor refolded A2-peptrde complexes.
Methods
CTL clones and cell lines
CTL clones from asymptomatic HIV-1 seroposrt ive patrentsand the corresponding (autologous) Epstein-Barr vrrus-trans-formed B lymphoblastoid cel l l ines (B-LCL) were establ ishedand marntained as descnbed prevrously (6,7). Subiects '115i ,
15760 and 16' lJ were previously shown to have srgnrf icantgagl .specrfrc CTL actrv i fy (6 and unpubl ished). HlA-typrngwas performed by the Massachusetts General Hospi ta lTlssueTyping Laboratory usrng standard serologrcal technrques(sub jec t 161J : A2, 43 ; 87 , 860; C3; DR2, DR4: sub jec t 010-1 1 5 i : A 2 , A 2 B ; B 1 a , 8 5 2 ; C w B ; D R 1 , 2 ; D O 1 a n d s u b y e c t15760: A2,844,851; C3; DR11) RPMI 16a0 (S igma, S t Lou is ,MO) contarning 20 % (vlv) heat- inact ivated FCS (Sigma) wassupplemented with u-glutamine (2 mM), penic i l i in (50 U/ml) ,streptomycrn (50 Fg/ml) and used for al l cel l l ines.
Gel filtration HPLC of HLA-A2 peptide complexes
Peptides were synthesized at the MIT Bropolymers laboratory.A2 refolding and complex format ion was in i t iated by di lut ionof the fwo oenatured subunrts in the presence of peptrdes asprevrously oescrtbed (2).
mAb bindtng to HPLC fracilons
mAb binorng to HPLC fractrons was used to monrtor refoldedA2-peptrde complexes. Samples of HPLC peak fract ions(20 trl) were pipetted onto a nitrocellulose fi l ter (Amersham,Santa Clearbrook, lL) and allowed to adsorb for 2 h at roomtemperature. The membrane was then incubated with 3%BSA in PBS followed by the respective mAb in 50 mM Tris-HCl, pH 8, 100 mM NaCl, 0 05% Tween. Specific binding of

(a) anti Gag/SLg clone D23
hjuction of CTLresponses by recombinant HLA-A| peptide complexes 103
(b) anti RT/[V9 clone 68A62
4
t(,
20
catn
3o'6q.)
v1
N
mCIR-mAtrA2-SL9
CIRA2+SL9 60
CIR-mAbAz-lvg
ClM2+lV9 {
P8l5-mAbA2-SL9
- O r : : '
#
- - c l - -
*
- - o - -
4-
- - - -E l - - " P8 l tmA-bA2- lV9
- - z - - P 8 l 5
(c ) anti Gag/SLg clone D23E :T
FACS staining
MHC c lass I express ion on unmodr f ieo C1R, K562 and PB15
cel ls was determined with mAb 887 7-FITC coniugate IB87'7( 1 6), ant i -HLA-A,B,Cl. Surface bound mAb-.SFDP coniugateswere detected using FITC-goat antr-mouse F(ab)z fragments(Jackson lmmunoresearch, Avondale, PA). Adsorptron of HLA-
A2 peptrde complexes to mAb-coated cel ls was shown by
an increase in MHC I expressron usrng the mAb 837.7-
FITC conjugate. lmmunof luorescent StalnrnQ was performed
according to standard protocols, usrng a FACScan f low
cytometer (Becton Dickinson, Mountarn Vierv ' CA).
Cytotoxicity assays
The slCr- labeled target cei ls (107 cel ls) lvere washed twrcewith cold PBS, incubated with 50 pM drthiotnrei to l for 30 min,
again washed with cold PBS ano resuspenoed in 200 pl of
SPDP-+nAb at 0.5 mg/ml. After 30 min, the cells were washed
twice with cold PBS, resuspenoed wrth 10 pg soluble HLA-
A2 peptide in 500 pl PBS at 4"C, and after 30 min cells were
E:T
*- C1R-MAFA2-SL9
-# I$62'mAbA2'SI-9
4- PsltmAbA2'SLg
- - o - - C I R ' E A b
- -.- - K562-EAb
- - X - - P 8 l 5 ' m A b
washed with cold RPMI and adiusted to 2X10s cet ls/mlCytotoxicity assays were performed in duplicate wtth CTLclones added at ef fector- to- target ratros (E:T) of 10, 5, 2 5and 1 25:1 to mAb-A2 pept ide-coated slCr- labeled cei lsand to pept ide-pulsed cel ls (prerncubated with a peptrde
concentration of 1 pg/ml for 30 mrn and *Ettiu-O) After 4 hat 37"C, cet l supernatants were assayed for 51Cr release.Total s lCr release was determtned by detergent lysis andpercent speci f ic lys is was calculated as [ (stCr ' samolerelease - Spontaneous release) i ( total re leaSe - spontaneous
release)]x100. Spontaneous release was always <209' . ofmaxrmal release.
Preparation of silica microspheres (beads) covalently mooiiedwith HlA-peptide complexes
We prepared si l ica beads (microspheres, 5 pm dtameter)whose surfaces are covaiently modified with the HLA-A2peptide complexes using either the RT ot gag nonapeptrdes.
Beginning with commercrally available (Bangs, LocatronU,
0 6 - n-
a a
v,
3(J
= {(J
ct
v,
S r (
O 6 - h
t r : r
Flg. 3. susceptibilaty ol target cells coareo wln oufified A2-peptide complexes lo lysis by RT- and Ga,q-specilic 9-T! clones. cytotoxrcrtva"i.vi n"i" ciri,ei out t",in the Gag-Sl9-specrtrc CTL clone D 23 (a_and c; ano ihe Rl'lvg-specriic ClL-rlone 18030 (b) on CIR cerls.precoated wrth mAb and loaded w(fi sotubte A2.oeptrde complexes (c1R-mAd-A2-peptide). as well as.c1R-A2-transfectants pulsed wrth
b"pr,o"lCrn-ez + peptroe. I [g/ml). CTL crones were arso tested agarnsr P815 cells and Pgl5-mAb-A2-peptrde as well as K562 and K562'mAO-n2-peptrde. nLe-A2 pepirO! complexes were bound to slCr-labeted larget cetls atter covalent anachmenl oi antl-HLA-A2-specrfic mAbMA2.l (a ano 3) or antr-F2-mrcrogtobutrn-soecrirc mAb BBMl to the cell sudace (c). Background lysrs for both T cell clones was determrneoby testr;g them agaanst u;ransle;ied clF and C1R cetls pulsed wrth the resoectve peptlde (data not shown)'

104 lnduction of CTLresponses by recombinant-r^-Ot peptide complexes
(a )
p sr.txtrfI lpirroIpcd
nqtap.pld.
o subLnl9lfrnorrrpo6rd
( c )
A2fc{tldo comfler
lN) aminopropyt s i l ica beads (4 NH2 groups/100 A2), we f i rst
prepareo an acuvated surface usrng the bis N-hydroxysuccini-
mide der ivatrve of a diacid of o l igoethyleneglycol (Fig. 4a).
The beaOs were first washed twice wrth deiontzed water,
twice wi th ethanol , then resuspended in a smal l amount of
anhydrous ethanol and added to a tube containing a solut ion
of 10 mM bis(NHS)ol igoethyleneglycoldicarboxyl ic acid (a
mixture of d iesters contatning seven to 14 etnyleneglycol
uni ts) ano 10 mM tr iethyl amtne in anhydrous drmethyl for-
mamide. The react ion tube was inverted gent ly ( for mixrng)
for 24 h at room temperature. The beads were then wasneo
three trmes with deionized water and modrf ied in one of
two ways:( i )The act ivated beads were converted into non- interact tng
control beads by reacting them as above with 10 mM amtno-
ethanol, yielding a OH-terminated oligoethyleneglycol-derivat-ized surface. lt has been shown prevlously that surfaces
o'o
oN
( ) oo
oOJ
'o
(.)o
F G .
(.)r \
ol
.,o2I r tc
Log Fluorescence IntensitY
Fig.4. Schematic represenratton oi beads covalently modilied with HLA-A2 peptrde complexes and FACS analysls HLA-peplide complexes
,u"-r" io*rJnty corpred to 5lm srtica oeioidenvatized with actve (NHS) ester orouos at the temrnal posmdns ol oligoethylensgtycol {seven
to 14 EG orougs per chain). The drstnbutron ol potentrally reactlve e'amrno grouoiol.lysne residues oilhe HlA-A2__peptide complex is shown
on tt" ,lg;t. fh" bornons ol the tysrnes were rdentrlied usrng 15e structure ol lhe HLA-A2 peptide comptex deposrled in the ptotein data banK
iili "rii"L,v"t,i-rir;g
ouanta. HLA-A2 peptrde-coaled beads wefe prepared lor FAcs analvsrs exaclly as were the cells These beads
(5x105) were lncubalecl wrth a porycronat ,"bb't serum (11200) agalnst heavy chatn (b) and_1 ltg ot HLA-A2-soecrfiC mAb MA2 1 (c) ano
ii"in"o'r",rn irrc-go"t anrr+auoit Fiab)zlrag-"nt. 1"otid tin", b) aino goat anir-mouse F(ab)z kagments (solid.line. c) , Beads harborrng onlv
a OH-lermrnared olrgoetny€negtycor wlre useo as a negatNe control ano slarned as deacrrbed above (dashed line in b and c) The
lluorescence Inlensity oi c1R ce s wrth r;c;i-J rulic ir""ir "rpression
(HLA-B.C) starned with polyclonal rabbrt serum-('l-:2OO) against heavv
chain is srmrlar to (b) (data not shown)
r 0 4',03ro2r o lroot 0 4, 0 3' 'o t. ,00
modrfied covalently with short oligoethyleneglycols are oftenhighly resistant to protein adsorptron (17).
(i i) The activated beads were reacted with either the A2-
RT pept ide complex or the A2-gag pept ide complex usrng-50 pg of e i ther of the A2-pept ide complexes (1 nmol)
dissolved in PBS (1.5 mi) wi th ' l mM tr iethylamine. Approxr-
mately 3 mg of the actrvated beads (10 nmol) were useoper reactton mrxture. ASSumrng a maximum COVerage ol i
complex/3g nmz, at most 0.1 nmol of protein would form a
monolaver on the sudace of 3 mg beads; therefore, theprotetn was added at 1O-fold molar eXceSS over reactrvegroups in the reaction. The solution was allowed to tnvertgently for 6 h at room temperature and the beads were
washed as described above. They were finally Stored at 4"C
in PBS. 0.1"/o sodium azide and 1 1r9/mt of the appropflalepeptide. The residual concentration of soluble peptide aher
washing and resuspension of the beads was calculated to
(b )
A - l
l r/ rt 1
t l
,- t
tniuction of CTLresponses by recombinant HLA-A2 peptide canplexes
(a) HLA-A2 peptide coated Beads (b) peptide pulsed autologous PBMC
c e l l s / w e l l c e l l s / w e l l
t 0 0 0 0 t 5 0 0 0 o 5 0 0 0 t 0 0 0 0 t t c o c0 5 0 0 0
1 0 5
o'aa
oazxtl
aa
oa
x
( c )
pep t ide pu lscd au to logous PBI \ tCHLA-A2 pep t lde coa ted Beads
pep t ide pu lscd C IR-A2 ce l l sHLA-A2 pep t idc coe ted Beads
pcpt ide pulscd autotogous PBI\ ICHLA-A2 pcp t idc coa ted Beads
precursor CTL per 106 PRMC
I (N ) 19 (12 -31 ) 326 (24e428)| (o-8) 2 (0{) 1e8 ( ts} '2s6)
4 (1 -11 ) 2 (0 -s ) l l 0 (84 '144)il (620) 6 (3-14) s2 (37-72)
r 9 (t t -31) t2 (&22) /t '19 (r!6.591)20 (1233) 6 (914) ZE (7enn
su bject
1 6 r J
1 15i
1 t/60
lv-9 sL.9
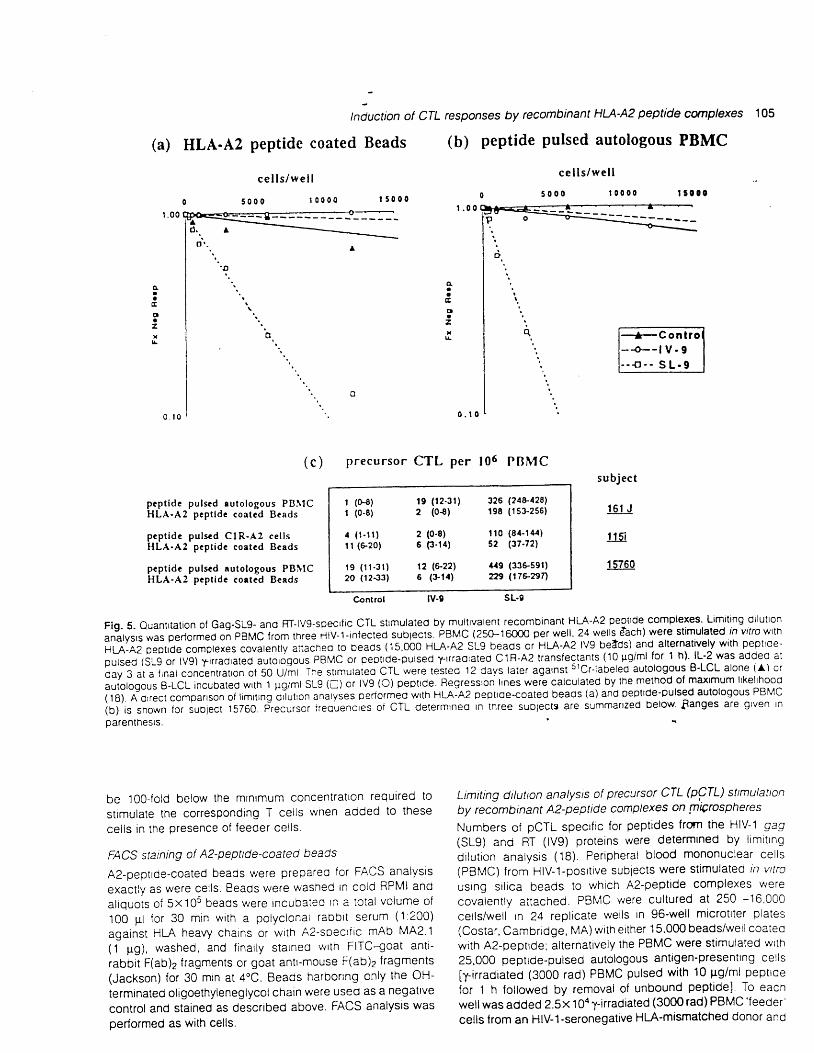
Flg. S- Ouantrtation ol Gag-SLg- and RT-lvg-specric CTL strmulated bymultivalenl recombinant HLA'A2 peptrde complexes' Limiting dilullon
"ni'r,, n lr p"Ao,."O oi pgpl6 lrom tl,iee ilv-l-rnfecled sublects. eAUC lZSO-l OOOO per well. 24 wells Aach) were stimulated rh vitto wth
HLA-A2 Deotrde complexes covalenlly anached to beads {15.OOO HLA-A2 SLg beads or HLA-A2 lvg betds) and alternatvely w$ peplroe-
pulsed {SLg or lvg) l-rr.adrated autologous PBMC of ceptrde-pulsed trrraoraled CIR'A2 translectanls (10 Fg/ml lor t h} lL'2 was added al
l i ig "i"
i,n"r concenrrarron of so U/;t The srrmurareo cTL were rested 12 days later against srcr-labeled auloiogous B-LCL alone ia) or
ir-riotogous a-rcr incubaled wft 1 pg/ml sLg (c) or lv9 (O) peptlde. Fleg.essron lrnes were calculated by the melhod ol maximum lrkelrhood
fr dl n'ou""t "orp"nson
ot limrring dlluro; aniryies oeaormeo w'th HLA:A2 geptrde-coated beaos (a) and pepttde-pulsed autologous PBMc
ioj' i" ano*n lor subjecl 15760. precu,st, irequenc,"s oi CTL delermrneo In tnree suoiecte are summanzed below Sanges are grven In
parenlhesrs.
be 1QQ-fold below the minimum concentratron reqUlre6 to
St imulate the corresponding T cel ls wnen aoded to these
cel ls in the presence of feeder cel ls.
FACS staining of A2'pepttde-coated beaos
A2-oeptrde-coated beaOs were prepareo for FACS analysis
exact ly as were cel ls. Beads were washec In cold RPMI and
a l iquots o f 5x10s beaOs were incubateo In a to ta l vo iume o f
100 p l fo r 30 min w i th a po lyc iona l raobr t serum (1 :200)
against HLA heavy charns or wrth A2-specr i ic mAb MA2' 1(1 pg), lvashed, and f inal ly starneo wrth FITC-goat ant i -
rabbi t F(ab)z f ragments or goat antr-mouse F(ab)z f ragments(Jackson) for 30 min at 4"C. Beacjs harborrnQ only the OH-
terminated oligoethyleneglycol chain were useO aS a negatlve
control and Stained as described above. FACS analysis was
performed as with cells.
Limiting ditution analysis of precursor CTL (O,Cfq silmulatton
by recombrnant A2-peptide complexes on {nQrospheres
Numbers of pCTL specific for peptides frcm the HIV-1 gag(sL9) and RT ( lvg) proteins were determined by l tmitrng
dr lut ion analysis (18). Per ipheral b lood mononuclear cel ls(PBMC) from HIV-1-posrt ive subjects were st imulated in vr t ro
usrng sr l ica beads to which A2-pept ide complexes were
covaleni ly at tached. PBMC vrere cul tured at 250 -16,000
cel ls/wel l tn 24 repl icate lvel ls in 96-wel l microtr ter p lates
(costar, cambridge, MA) wi th erther 15,000 beads/wel l coateowith A2-peptrde, ai ternat lvely the PBMC were St imulated wrth
25,000 peptrde-pulsed autologous ant igen-presentrng cel is
[y- i r radiated (3000 rad) PBMC pulsed with 10 [g/ml peptroe
for t h foilowed by removal of unbound peptidel To eacrr
well was added 2.5x 10a y-irradiated (3000 rad) PBMC 'f eeder
cells {rom an HIV-1-seronegative Hl-{-mismatched donor and

106 lnduction of CTL responses by recombinant HIA-42 peptide complexes
IV9/SL9 Beads+ fecder cclls
IV9 Beads+ feeder cells
SL9 Bcads+ feeder cells
IV9/SL9 Beads
soluble pcpt lde/SL9( 5 n g m l ' l ) + f e e d e r c e l l s
or ig inal Beads unmodi f icd+ feeder cel ls
Bcads modi f ied ( l inkcr)+ feeder ccl ls
7. specilic kllllng
Flg. 6. T cert stimutation by multivalent recombinant HLA-A2 peptide complexes is specilic. PBMC (20.000 'vell) from HlV.1-positive subjectt5760 were ptaced in culture along with the 5 F n srlrca beads (20,000/well) modilied covalenlly wath the HLA-A2 peptid€ complexes in thegresence or absence ol HLA-mismatched y-rradrated feeder cells (7x lor/well). lL-2 was added at day 4 at a linal concentration ol 50 U/mlAfter 12 davs culture. lyltc activtty was assayed agarnst 5lCr-labeled autologous 8-LCL alone (solrd bars) or autologous B-LCL incubateo wnh1 frg/mt lvg (open bars) or SLg (shaded bars) peptde. Data are expressed as a percenlage of specrlic srcr release and values.epresentthe average ol lour welis.
. r o 807060504 03020' t0
recombrnant lL-2 (50 U/ml added on day 3) After 12 dayseach weil was split and assayed for cytotoxtcity on slcr-
labeled autologous B-LCL that were ei ther Incubated withsynthet ic SL9 or lV9 pept ide (1 pg/ml peptrde for t h andwashed) or B-LCL without added peptrde. The fractron ofnon-responding wel ls was the number of wet ls in which s lCr
release did not exceed the mean + 3 SD of the release Inthe 24 control wel ls (no pept ide) div ided by the total numberof assayed wel ls (19,20). pCTL freeuency was estrmated bythe maxrmum l ikel ihood method (18).
T cell stimulation by recombinant Hl-A-peptide complexes
PBMC (20,000/wel l ) f rom HIV-1-posi t ive subiect 15760 wereplaced in 96-wel l mtcrot i ter p lates along wrth the 5 pm st l icabeads (20,0004,,/ell) modified covalently wrth the A2-peptrdecomplexes tn the presence or absence of 1- i r radiated HLA-mismatched feeder cel ls (3000 rad, 7x t04/wel l ) . lL-2(Hoffman La Roche) was added at day 3 at a f inal concentra-tron of 50 U/ml. Af ter 12 days lyt ic actrvrry was assayed onpept ide-pulsed slcr- labeled target cel ls (autologous B-LCLpulsed wrin 1 Fg/ml pept ide as descrrbeo above).
Resul ts
Refolding of HIA-A2 in the presence of HIV-1-derived pepttdes
HLA-A2 complexes with RT-lvg (A2-lV9) and Gag-Slg (A2-
SL9) were formed by dilution of recombinant heavy chain(1 pM) and p2-microglobulin (2 pM) in the presence of each
peprrde (100 pM) (2). The complexes appeared as sharppeaks on HPLC gel f i l tration at €lutron times consistent wrththe mot wt of the ternary complex (Fig. 1). HPLC fractions wereplpened onto ni t rocel lu lose and stained with mAb specrfrcfor c iass I MHC proterns (MHC l) The f i rst peak fractron,corresponding to the fo lded complex, was recognized bymAb W6/32 (11), whrch recognrzes human MHC I moleculesin general , and by'mAb PA2.1 (12), whictr recognrzes thenatrve HLA-A2 molecule specifically Thdrecognition by mAbsensttrve to conformatron rmpl ies that the recombinant com-plexes contain native epitopes, consrstent with the presenceof correct ly fo lded molecular complexes.
Adsorptton of A2-peptide complexes to target cells
To determine if the A2-peptide complexes assembled in vrtrocan activate Az-restricted T cell clones, we coated a panelof A2-negatrve cel ls wi th the complexes and used them astargets for RT-|V9 and Gag-SL9 specrf ic T cel l c lones derrvedfrom HIV-1- infected indiv iduals (c lone 68462: antt-A2- lvg anoclone D23: ant i -A2-SLg) in cytotoxrc T cel l assays Themoorfiecj target cells were the Ht-A-A negative human B celll i ne C1R (21) , the MHC Inegat rve numan ce l l l i ne K562, anoP815, a mouse mastocytoma cel l l rne. To bind the A2-peptrdecomplexes on these cells, we first covalently attached theA2-soecrf ic mAb MA2.1 (14) to the surface of d i th iothrertol-t reated C1R and PB15 cel ls using the heterobi funct ional cross-linker SPDP. The antibody coated cells were subsequentlyincubated with a saturating concentration of soluble A2-

ndlrtion of crL
peptide comptexes. The binding of A2 to the cell surface was
demonstrated byflowcytometry, using mAb B87'7 (16)' which
reacts wrth a combinatorial determlnant of H|'-A-A,B,C heavy
chain ano p2-microglobulin (Fig. 2).
specific lysis of A2-peptide-coated target cells by cTL clones.
As shown in Fig. 3, the cTL ctones lysed c1R cells coated
with the relevant A2-peptide complexes. rndicatrng that the
Hlv-1 pepttdes were effectively presenteo and recogntzed by
T ceils bearing receptors of the approprrate specificiry. ClR
cells coated with the reconstituted A2'pepttde complexes
were lyseo essentially as rvell aS were C1R cells transfected
with A2 and pulsed with pept ides (1 pg/ml) ' Both T cel l c lones
also lysed the murine P815 cel ls to whrch the approprrate A2-
pept ide complex had been adsorbed. al though to a lesser
extent than human target cel ls modrf ieo wrth the same com-
plexes. As expected. the T cel ls far led to lyse unrrrodrf ied
P815 cer ls and c1R-A2 t rans fec tan ts pu lsed wt th an
irrelevant pePt ide.Recognrt ion of A2-pept ide complexes by TCR was evident ly
not imparred by the binding of these complexes to mAb
MAz.1, even though this ant ibody brnos to the polymorphtc
regron of HI-A-A2 close to the peptrde brnding groove (22)'
HLq-nz pept ide molecules held by mAb MA2.1 at the cel l
surface mrght not be accessrble for optrmal engagement wi th
the TCR. Accordingly mAb BBM1, an ant i -p2-microglobul in
ant ibody (13) , was cova len t ly l inked to C1R, K562 and P815
cei ls. and used to bind refolded complexes. A2-pept ide
complexes on each of these target cel ls were iust as suscept-
ib le to lysrs (Fig. 3c) as target cel ls that made use of the
MA2.1 mAb.
Covalent coupling of A2-peptide complexes to mrcrospheres.
A c r u c i a | q u e s t i o n i s w h e t h e r t h e r e c o m b | n a n t m o i e c u | e s c a nst imulate pCTL present in blood samples of HIV-1-seropostt tve
sublects, i .e. in natural ly prrmed indrvrouals. To examtne this
possrbi l i ty , we covalent ly coupled the A2-peptrde complexes
to s i l ica beads der ivat ized wrth act lve ester groups at the f ree
ends of o l rgoethyleneglycol charns tnat extenoed -3O-50 A
from the bead surface.Figure 4 shows the coupl ing proceoure ano the dlstr lbut ton
of potenttal ly react ive e-amrno groupS of lysrne resrdues of
the A2-peptrde complex. The partrcular amtno groups that
react with acttvated ester groups on tf ' le surface of the bead
determrnes the or ientat ton of the MHC complex. Posi t ive
staining rvr th 3 rabbi t serum against human heavy chain (Fig
4b) showed attachment of HLA complexes on the surface.
Stainrng wrth the ant i -Ht-A-A2 mAb MA2 1 (Fig 4c) indtcated
that a srgni f icant f ract ion of the HLA-peptrde complexes
matntaineo a nat ive conf igurat ion and were exposed to the
solvent, and are thus presumably accessrble to TCR on T cel ls.
Recombtnant HlA-pepttde complexes are rmmunogenic
To assess the immunogentc ef fectrveness of the A2-pepttde
complexes on beads, we used them to st tmulate PBMC of
th ree seroposr t rve sub lec ts (161J , 1151 and 15760) measur ing
the frequency of peptrde speci f ic pCTL af ter 12 days by
l imit ing or lut ion analyses (Fig. 5) . The resui ts show that the
1.ecorntinant A2-peptide complexes were effective and as
specific as pepttde-pulsed autologous antrgen-presentlng
responses by recombinant HU-A2 peptide complexes 107
cells in inducing functional responses that normally depend
upon ceil<ell interactions, i.e. they triggered antigen{epend-
ent responses of primed pCTL. A direct comparison of l imiting
dilution analyses performed with A2-peptide complexes dis-
played on beads (Fig. 5a) and autologous peptide-pulsed
PBMC (Fig. 5b) revealed that the responses were of compar-
able magnitude.
Expansion of pCTL is peptide specific
To rule out non-specific mitogenlc effects of the bead surface,
PBMC were stimulated with a mixture of A2-lV9- and M-sL9-
coated beads and tested for A2-lv9- and M-sL9-specific
responses (Fig. 6). Only SL9 specific responses were detect-
able. culture of the same PBMC with beads bearing only A2-
lV9 induced no response; only A2-sL9 beads stimulated a
specific SL9 response. The apparent absence of lV9-specificpcTL in the responder population was confirmed using the
same batch of beads as in the l imiting dilution analysis
(Fig. 5). The recovery of SL9-specific CTL suggests a true
expansron of peptrde-specrfic pcTL, confirming other data (6
and unpublished), that the dominant Gag-specific HLA-A2
restncted response is directed against the SL9 peptide in
these subjects.Titratron of peptide revealed that a peptide concentration
over 5 ng/ml SL9 was requrred to stimulate pcTL expansion
from PBMC in the presence of feeder cel ls (Fig. 6 and data
not shown). By contrast much less pept ide is needed when
it is presented by HI-A-A2 on the microspheres. Thus with
maximal packrng a 5 pm microsphere could accommodate
about 2x 106 HLA-A2 peptrde complexes in a monolayer. This
numoer corresponds to 0.35 ng/ml peptide if all the A2 on
the beads were loaded with the peptide originally and all the
peptrde adduct then dissociated (given 2X 101 beads, 0.2 ml l
wel l and a maxtmum of 2x103M-pept ide Complexes per
beao) By comparing the fluorescence intensity of HLA-A2
oeot ide-coated mtcrosoheres wi th C1R cel ls stained wrth
polyclonal antrbodres to MHc I heavy chain i t appears,
however, that the actual amount of surface-bound A2-pepttde
complexes on the pnrcrospheres is -50 t t rqes lower than the
calculated maxtmum. re. the f luorescence intensi ty ts stmt lar
to C1R ce ls v r i th -3x104 HLA 835 and Cw4 per ce l l (21)
Thus r t appears that the T cel l response was el ic i ted by far
less peptide associated wrth H[-A-A2 on the microspheres
than by pepttde addeo rn solut ion. Non-speci f ic mrtogenrc
effects of the beads modrfied wrth only OH-terminated oligo-
ethylenglycol groups were not observed (Fig ' 6) ', .
D iscuss ion
This is the first demonstratron of biological activity of E' co1-
derrved HLA-A2 peptroe complexes' In contrast to nattve
HLA-A2, the recombrnant protern used here ts not glycosylated
at Asn-86 and i ts p2-mrcroglobul in subuni t has an aodrt ional
N-termrnal methionrne resrdue not c leaved fol lOwing synthesrs
(3). These di f ferences have previously been shown not to
significantly affect the crystal structure of A2 in the form of
A2-pept ide complexes (3 '4) .We show here that these differences also do not prevent
recombinant A2-Oeptrde complexes from el ic i t ing spectf ic
cytolytic responses of CDB* CTL clones from HIV-1-infected

1OB tnduction of CTL responses by recombinant HtA-42 peptide complexes
individuals. Thus, the antigen-specrfic receptor (TCR)of these
clones can recognize recombinant E. colr-produced A2'proteins refolded in assoctation wrth approprlate peptides'
This study shows furthermore that mtcrospheres coated withrecombinant A2-peptide complexes can specifically stimulateT cel ls in PBMC from HIV-1- infected indivrduals. The st imulatedcells, presumably primed by natural infectron, develop into
an expanded population of CTL that specrfically responds tothe A2-peptide complex that trlggered this development.Since the Az-peptide-coated microspheres lack B7 and otherco-stimulatory molecules, their immunogentciN has to beconsidered in the l ight of the large body of evrdence that, in
addition to TCR ligation, the actrvation of quiescent T cellsdepends upon secondary signals that trrgger the expressronof lL-2 receptors and the production of lymphokrnes (35). In
the present system which makes use of PBMC from HIV-1-rnfected indiv iduals, i t is possrble that the responding T
cel ls already express lL-2 receptors and that the exogenousrecombrnant lL-2 substrtutes for endogenous production oflymphokines. In addi t ion, the HLA-mrsmatched feeder cel lsmight provide a co-strmulatory functron and, by elicit ing analloreactron, tnduce cells in the respondrng PBMC population
to produce endogenous lymphoktnes. In any event, wnatever
the detai led mechantsm may be, r t seems clear that the beadscoated covalently with A2-peptrde complexes are activeimmunogenical ly wi th PBMC of HIV-1- infected indiv iduals.
Several drfferent systems have been developed for stimulat-ing T cells wrth purif ied class I MHC molecules (24-26)
including al logeneic MHC proteins (27. 28) and var iousmethods have been used to assay T cell actlvation, e.g.release of BlT-esterase (29), production of lL-2 in T cellhybridomas and release of serotonrn in basophils transfectedwith a TCR-( fusion (30). The MHC-peptide complexes usedin these studies included purrfied native MHC I molecules,
cleaved from the cellular surface (28,29), and 'empty' MHC Imolecules produced in rnsect cel is and loaded with pept ides
of interest (31,32). Soluble H-2Ko-peptrde fusrons were alsoproduced in which heavy charn, p2-mrcroglobulrn and pept ide
have been fused Into a s ingle cnaln protetn and cel lsexpressing these constructs were shown to be highly immuno-genic (33). Genet ical ly engrneered soluble H-zOd moleculescoated on plast ic were also found to present HIV-1 envelopepept ide to an ant igen-specrf ic CTL clone, inducing i t toproduce IFN-y (3a).
Since the overexpression of the class I subunrts in the E. coli
system and the subsequent refolding provides an abundantsource of homogenous Ht-A-peptide complexes, the recom-
binant soluble complexes descrrbed here of fer new toolsfor explor ing drverse problems and tssues, such as s lgnal
t ransductron events el ic i ted by agonrst and antagonrst pept ide
l igands (36,37), the ident i f icatron ancj isolat ion of ant igen-speci f ic T lymphocytes ln complex cel l mixtures, and thedevelopment of vacctnes aimed at st tmulat tng part lcular antt-gen-speof ic T cel ls.
Acknowledgements
J. W. is a recrpient of a Mildred-Scheel fellowshrp from Germany andwas supported by grants lrom the NIH (Al 34247 and CA 60686).The efforts of C. 8. were supported by the Schwerzerische Stiftung
fOr medizinisch biologische Stipendien, M. M. was a Eli Lilly predoc-toral fellow and supported by an NIH grant (GM 30367). D. N. G.was supported by grant no. DAMD17-94'J'4060 from the Departmentof the Army and his eflorts were performed in the laboratory of D. C.Wiley who is a Howard Hughes Medrcal lnstitute Investigator.'Wethanx P. Cresswell lor provrdrng the C1R'A2 cells and A. Trocha iorexcerlent technical asststance.
Abbreviat ions
B.LCL
CTLPBMCSPDP
Epstein-8arr vrrus-transformed B lymphoblastoid celll inecytotoxic T lymPhocytepenpheral blood mononuclear cel lMsuccinimrdyl 3-(2-pyrrdyldithio)propionate
References' l Germarn, R. N. 1994. MHC-dependent antigen processtng ano
pepttde presentatton: provrdrng ligands lor T lymphocyteactrvatron. Cell 76.287 .
2 Garboczr. D. N., Hung D. T. and Wiley. D. C. 1992. HLA-A2peptrde complexes: relolding ano crystallization of moleculesexpressed rn Eschenchia coliand complexecl with single antlgenrcpeotldes. Proc. Natl Acad. Sci. US'A 89:3a29.
3 Madden, D., Garboczr. D. N. and Wiley, D. C. 1993. The antigenrcidentrty ol peptide/MHC complexes: a comparison of theconformatrons of five vrral peptrdes presented by HI-A-A2. Cel/75:693.
4 Biorkman, P. J., Saper, M. A., Samraoui, B'. Bennen. W S.,Stromrnger, J. L. and lVi ley, D C 1987. Structure of the humanclass I hrstocompatrbrlity antrgen, H[-A-A2. Nature 329: 506.
5 Tsomrdes, T. J.. Walker, D. B. and Eisen. H. N. 1991. An optrmalvrral peptrde recognrzed by CD8' T cel ls binds very t ightly to therestnctlng class I MHC protern on tntact cel ls but not to punfiedclass I protern. Proc. Natl Acad. Scr. USA 88:11276.
6 Johnson. R. P., Trocha, A., Yang, L., Mazzara' G. P., Panical i ,D. L.. Buchanan T. N. and Wblker, B. D. 1991. HIV-1 gaEl-specrtrccytotoxrc T lymphocytes recognrze multiple highly conservedeprtopes. J. lmmunol. 147'.151?
7 Vlhlker, 8., Flexner, C.. ParaQs, T. J.. Ful ler, T. C., Hirsch. M. S,Schootey, R T. and Moss, B. 1988. HIV-1 reverse transcrrptases a target for cytotoxrc T lymphocytes tn infected indrvrdualsScience 240'.64.
B Falk, K.. ROtzschke. O . Stevanovrc, S., Jung, G. and Rammensee,H G. 1991. Al lele-specrf ic motrfs revealed Q1t sequenctng of serfpeptroes eluted from lv' tHC molecules. Nature 351:290.
9 Tsomroes, T. J. ' , Aldovrnr, A., Johnson' R. P., $hlker, B. D., Young,R A. ano Eisen. H N 1994. Natural ly processed vlral peptrdesrecognrzed by cytotoxrc T lymphocytes on cel ls chronrcal lyinfected by human rmmunodeficiency Vrus Type 1. J. Exp.Med. 180:1283.
10 Walker, L D.. Flexner, C.. Birch-Limberger, K. ' Fischer' L., Paradrs.T. J., Aldovrni, A., Young, R., Moss, B. and Sct ' tooley, R. T. 1989.Long-term culture ano f ine specrf ici ty of hurnan cytotoxlc T'tymprrocyte c lones react rve wr th HIV-1 Pt . . Nat l Acad. Sc i .USA 86:9514.
11 Ways. J P and Parham, P. 1983. The antigenic structure of HLA-A2: an analysis with competrt ive brndrng assays and monoclonalant rboores. J . lmmunoL l3 l :856.
12 Parham, P and Bodmer, W. F. 1978. Monoclonal antrbody to ahuman hrstocompatrbrhfy alloantigen, H[-A-A2. Nature 276 397
13 Parham, P. , Andro lewrcz, M. J . , Holmest , N. J . and Rothenberg,B. E. t 983. Arginrne 45 rs a malor part of the antigenic determrnantof human p2-microglobul in recognrzed by mouse monoclonalant rbody BBM.1 . J . B io l . chem.258:6179.
'14 McMrchael, A. J., Parham, P., Rust, N. and Brodsky, F M. 1980A monoclonal antibody that recogntzes an antigenic determtnantshared by HLA-A2 and I 17 . Hum. lmmun. 1: 121.
15 Kranz, D. M. and Eisen, H. N. 1987. Resistance of cytotoxrc Tlymphocytes to lysis by a clone of cytotoxic T lymphocytes. Proc.Natl Acad. Sci. USA 8a:3375.

16
1 7
Brodsky, F. M., Parham, P., Barnstable, C J.. Crumpton, M. J.and Bodmer. W. F. 1979. Monoclonal antrbodres for analysis ofthe Hl.-A system. lmmunol. Rev. 47.3.Prime, K. L. and Whitesides. G. M. 1991 Self assembled organicmonolayers are good model systems for studytng adsorptton ofproteins at surfaces. Science 252:1164Fazekas oe St Groth, S. 1982. The evatuatron oi l imit ing di lut ionassays. J. lmmunol. Methods 49:R11.Koup, R. A., Pikora, C. A., Luzurraga, K , Erett ler, D. 8.. Day,E. S., MazzTJs, G. P. and Sull ivan, J. L. 1991. Ltmtttng di luttonanalysrs of cytotoxic T lymphocytes to human rmmunooefrctencyvrrus 9a9 antigens in infected persons: n vilro quantltation ofeffector cel l populat ions wrth p17 and p24 specrf icttres. J. Exp.Med. 17a:1593.Lefkovrts f . and Waldmann. H. 1979. Limttng Dilution Analysis ofCells ol the lmmune System, p. 38 Camorrdge Untversrty Press,CambrrogeZemmour J., Lit t le, A. M., Schendel, D J and Parham. P' 1992.The HLA-A,B 'negative' mutant cel l l rne C' lR exgresses a novelHt-A-835 al lele, whrch also has a polnt mutatlon rn the translattoninit iatron codon. J. lmmunoL l4S:1941
22 Santos-Alguaoo, J . , Barbosa, J A, Brro . P A. and Stomrnger .J' L' 1988' Molecular characterrzatton of lhe serologrc recognittonsltes ln the human HI-A-A2 molecule J. tmmunol. 141:2811.
23 Coli lns. E. J . Garboczr, D. N and Wley, D. C. 1994. Threedimensronal structure of a peptrde extendrng from one end of aclass I MHC binding site. Nature371'.626.
24 Goldstern. S. A N. and Mescher , M. F 1988. T ce l l recognt t ionof nonpolymorphic determtnants on H-2 class I moleculesJ. lmmunol. 140:3707.
25 Kane. K. P., Sherman. L. A. and Mescher, M F 1989. Molecularinteractrons required for tr iggerrng al loantrgen-specrf ic cytotoxlcT lymphocytes. J. lmmunol. 142.4153.
26 Mescner. M. F.. O'Rourke, A. M.. Champoux, P. and Kane, K. P.199'1. Eourl ibrtum bindrng of cytotoxrc T lymphocyles to class Iantigen. J. lmmunoL l4T:36.
27 McCluskey. J.. Boyd, L. F., Hignet, P. F, Inman, J. and Margul ies,D. H. 1988. T cel l act ivatron by purrfred, soluble class I MHCmolecu les. Reout rement for po lyva lency J . lmmunoL l4 l : '1451.
28 El l iott . T. J. and Eisen, H. N. 1990. Cytotoxrc T lymphocytes
tn-duction of CTLresponses by recombinant H|A-A| peptide complexes 1Og
recognize a reconstituted class I histocompatibility antigen (Ht.A-M) as an allogeneic target motecule. Proc. Natl Acad. Sci.USA 87:5213.
29 Kane, K. P., Vit iel lo. A., Sherman. L. A. and Mescher, M. F 1989.Cytolytic T-lymphocyte response to rsolated class I H-2 proi'ernsand inf luenza peptides. Nature 340:157.
30 Abastado. J. P., Lone. Y C., Casrouge, A.. Boulot, G. andKourilsky, P. 1995. Dimenzatron of soluble maior histocompatibilityhybridoma and inductron of unresponsiveness. J. Exp. Med.182:439.
31 Godeau, F., Luescher, l . F., Oicius, D. M . Saucier. C., Mottez. 8.,Cabanie, L. and Kourtlsky, P. 1992. Purtfication and ligand bindingof a soluble class I malor hrstocompatrbility complex moleculeconsisting of the first three domarns of H-2Kd fuSed to P2-mrcroglobulin expressed In the baculovtrusinsect cell system.J. 1iol. Chem. 267 :2a223.
32 Huang, J. H., Getty, R. R., Chisarr. F. V.. Fowler, P., Greenspan,N. S. and Tykocinsky, M. L. 1994. Protein transfer of preformedMHC-peptide complexes sensrtizes target cells to cTL cytolysrs.lmmunity 1:607.
33 Monez, E., Langlade-Demoyen, P., Gournier. H., Mart inion, F.,Maryanski, J., Kouri lsky, P and Abastado, J. P. 1995. Cellsexpressing a major htstocompattbrlity complex class I moleculewrth o srngle covalently bound peptrde are highly immunogenrc.J. ExP. Med. 181'.493.
34 Takeshita, T., Kozlowski, S., England, R. D., Erower, R ,Schneck, J., Takahashr, H , DeLisr, C., Margul ies. D. H. andBerzofsky, J. A. 1993. Role of conserved regions of class I MHCmolecules in the acttvatron of CD8' cytotoxrc T lymphocytes bypeptrde and purified cell-free class I molecules. lnt. Immunol.5 :1 1 29 .
35 Hardrng, F. A. and All ison. J. P. 1993. CD28'87 interactions al lowthe induction of CD8* cytoioxrc T lympnocytes in the absence ofexogenous help. J. Exp. Med. 177.1791.
36 Sloan-Lancaster, J.. Shaw, A S., Rothbard, J. 8. and Allen, P. M.1994. Partial T cell signalling: altered phospho-( and lack of zap70 recruitment in APL-induced f cell anergy. Cell 79:913.
37 Madrenas, J., Wange, R. L., Wang, J. L.. lsakov. N.. Samelson,L. E. and Germain, R. N 1995. ( phosphorylat ion without zap-70 actrvatron induced by TCR anlagontsts or part ial agonrs(sScience 267:5'15.
1 8
1 9
2Q
z l