structure and composition of the bacillus anthracis …fig. 1. bacillus anthracis capsular cells...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Oct., 1965Copyright © 1965 American Society for Microbiology
Vol. 90, No. 4Printed in U.S.A.
Structure and Composition of the Bacillusanthracis Capsule
A. A. AVAKYAN, L. N. KATZ, K. N. LEAVINA, AND I. B. PAVLOVALaboratory of Morphology of Microorganisms and Electron Microscopy, Gamaleya Institute of Epidemiology
and lMicrobiology, Academy of Mledical Sciences, M1Ioscow, USSR
Received for publication 29 May 1965
ABSTRACTAVAKYAN, A. A. (Academy of Medical Sciences, Moscow, USSR), L. N. KATZ, K. N.
LEVINA, AND I. B. PAVLOVA. Structure and composition of the Bacillus anthracis cap-sule. J. Bacteriol. 90:1082-1095. 1965.-Observations by various methods of light micros-copy (phase contrast, dark-field, and fluorescence) revealed the complex structureof the Bacillus anthracis capsule, which changes regularly during the growth cycle of theculture. Special cytological methods of staining the capsule made it possible to studyits fine structure, which is not revealed by negative staining with India ink. For ex-ample, the capsule shows a membranelike outline, fine transverse lines, and inter-ruptions and transverse septa traversing the entire capsule. By using cytochemicalmethods, it was found that the capsule has a stratified structure and that the variouslayers of the capsule differ as to the value of the isoelectric point, metachromaticability, sensitivity to various enzymes, and, consequently, chemical composition. Itwas thus shown that the membranelike outline of the capsule consists of peptides andneutral mucopolysaccharides. The middle part of the capsule consists of a complex ofsubstances of both polysaccharide and protein nature, and the inner part consists ofacid mueopolysaceharides. Observation of the capsular forms of B. anthracis by meansof an electron microscope revealed differences in the osmiophilia and submicroscopicstructure of the membranelike outline and the middle and inner parts of the capsule.Immunochemical studies coniducted by the fluorescent-antibody method revealed lo-calization of antigenis in different parts of the capsule, and made it possible to differ-entiate the capsular antigens according to their serum-staining ability and accordingof their relations to enzymes, i.e., their chemical composition. This paper concerns thepossibility of studying the fine structure of bacterial capsules in fixed preparations, andthe differences and similarities of the antigens of the capsule and cell wall of B. anthracisand of the related species, B. megaterium.
It is well known that Bacillus anthracis formsa capsule in special nutrient media and in in-fected animals. Many investigators associatethe virulence of the microorganism with thecapsule and its ability to produce the anthraxtoxin (Thorne, 1956; Keppie and Smith, 1963).However, the literature contains limited infor-
mation on the structure and composition of theB. anthracis capsule. In cytological studies ofthe capsules, most investigators have resortedto negative staining with India ink without pre-liminary fixation (e.g., Duguid, 1951). Thismethod does not lermit observation of structuraldetails in the capsule, because it reveals thecapsule negatively, as a white trace againsta dark background. Klieneberger-Nobel (1948)suggested cytological observations of the cap-sules in preparations fixed by Buin's solutionand then stained with Giemsa solution or with
crystal violet after treatment with tannin. Butthe treatment with tannin resulted in coagula-tion of the proteins and therefore made it im-possible to observe the fine structure of thecapsule. Observations of the capsular forms ofB. anthracis in ultrathin sections also failed tocontribute anything new to the studies of thecapsular structure, because the main part of thecapsule was destroyed while the preparationswere being made (Roth, Levis, and Willianms,1960; Roth and Williams, 1964).
Information on the composition of capsuleswas obtained mainly by chemical and immuno-chemical methods. Tomcsik (1954), Nordbergand Torsell (1955), and others have shown thatthe B. anthracis calpsule consists of glutamylpolypeptide and that it )ossibly also containslysozyme-sensitive polysaccharides.The present investigation was aiimned at ob-
1082
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 90, 1965 STRUCTURE AND COMPOSITION OF B. ANTHRACIS CAPSULE 1083
serving the B. anthracis capsule by light andelectron microscopy, as well as cytochemicaland immunochemical methods. An attempt wasmade to compare the data on the fine structureof the capsule with its chemical and antigeniccomposition.
MIATERIALS AND METHODS
To produce capsular forms, the 71/12 vaccinalstraini of B. anthracis, obtained from the secondTsenkovsky vaccine, was grown in Hanks mediumwith 40% ox serum (Arkhipova, 1962). To producenoncapsular forms, the culture was inoculated onbeef extract-agar.
The cytological study was conducted on un-fixed slide preparations with the aid of a phase-contrast microscope or dark-field illuminiationi. Insome cases, negative staining with India ink wasresorted to. Staining with crystal violet aftertreatment with tannin (Gutstein, 1924) or withLoffler's alkaline methylene blue, after fixationwith ethyl alcohol, methaniol, or formalin, was alsoutilized.
The cytochemical study made use of the iso-electric poinit (IEP) method (Pischinger, 1926). Todetermine the relative IEP value of the capsule,the capsular cells were stained with diluted solu-tions of Toluidine Blue and Acid Fuchsin at pH10.0 to 1.4 at 0.2 intervals. To reveal the poly-saccharides, the periodic acid-Schiff technique ofHotchkiss was used (Pearse, 1960); to reveal themetachromatic substances (mainly polysaccha-rides), Schmorl's thionine method was employed(Pearse, 1960). The Alcian Blue method (afterSteedman) and metachromatic staining with thio-niine and Toluidine Blue at pH 4.0 and 2.0 wereused (Pearse, 1960) to reveal acid mucopoly-saccharides; Fast Green at pH 2.2 after Alfert andGeschwind (1953) or bromophenol blue at pH 2.3(Bloch and Hew, 1960) was used to reveal proteins;the propylene glycol-Sudan Black method ofChiffelle and Putt was used to reveal lipids(Pearse, 1960).
For immuniochemical studies, the fluorescent-antibody method (Coons and Kaplan, 1950; Riggset al., 1955) was used. The smears were fixed onglass slides with ethyl alcohol, and were treatedwith fluorescent sera labeled with fluorescein-isothiocyanate. The sera were obtained by im-munizing rabbits with the capsular cells, and wereprepared from commercial precipitable sera(Levinia and Arkhipova, 1964).
The cytological and serological studies wereconducted on preparations treated with enzymes,as well as on untreated preparatioins. The follow-ing enzymes were used: pepsin (E. Gurr), trypsin(Merck), chymotrypsin (Chempl), lysozyme(Lawsoni), anid hyaluroniidase (Light).
For electron microscopy, whole unsectioinedcapsular cells were fixed in formalin vapor, andwere later overfixed in osmium tetroxide vapor orwere negatively stained with 2% phosphotungsticacid (pH 11.5) without preliminary fixation. The
preparations were examined with a JEM-6Celectron microscope, at 80 kv and X 10,000 magni-fication.
The light-microscope studies were carried outwith a MBI-6 microscope; the preparations werephotographed with an FMH-3 microcamera (oilimmersion, 10 X ocular, 100 X objective). A usualaplanatic condenser, an OI-13 dark-field con-denser, and a KF-4 phase-contrast device wereused for photography. An ML-2 fluorescencemicroscope was used, and the photographs weremade on PF-3 fluorographic film.
RESULTS
Cytological study of capsules by light micros-copy. Within 2 to 3 hr after inoculation of thenutrient medium with B. anthracis spores, theformation of capsules begins. Young capsulesare easily seen with phase contrast. At first theyform in various areas near the transverse septa,and have an interrupted structure. Within 12to 18 hr, the capsules completely surround thestreptobacilli, and have no interruptions. Often,streptobacilli form pairs or chains inside acapsule. The lysis of the capsules begins within24 to 30 hr. The capsules lyse first in areas cor-responding to the dead cells. The capsules ofold cultures, therefore, also have an interruptedstructure, like the newly forming capsules ofyoung cultures.An 18-hr B. anthracis culture with well-formed
capsules was used for the cytological study. Theliving cells, observed with a phase-contrastnmicroscope, clearly show capsules in the form ofa white halo about the streptobacilli. They arebounded on the outside by a barely observedmembranelike outline (Fig. 1). In preparationsnegatively stained with India ink, the capsuleis of the same size as in unstained preparations.Preliminary treatment of the calpsular cells withox serum leads to a noticeable nonspecific capsu-lar swelling (Fig. 2). This phenomenon wasdescribed by Tomesik and Guex-Holzer (1953,1954a) for various capsular bacteria as a non-specific swelling reaction, and was made a specialbasis for revealing capsules by treating them withsera containing proteins. All of the above-described methods of studying capsules in livingcells have failed to show the internal organiza-tion of the capsules.
In observations of living organisms by dark-field illumination, capsular cells show whitepunctate structures in the membranelike outline(Fig. 3) and in the inner lpart of the capsule (Fig.4). In noncapsular forms of B. anthracis, onlythe cell wall is clearly seen in dark-field illumi-nation (Fig. 5).
Fixed capsular cells were also studied in afluorescence microscope. Careful fixation of non-
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

FIG. 1. Bacillus anthracis capsular cells from an 18-hr culture. Membranelike outline of capsule indi-cated by arrows. Phase contrast. X 2,000.
FIG. 2. Capsular cells from a culture of the same age after treatment with ox serum. Negative stainingwith India ink. Note swelling of capsule. Light microscope. X 1,200.
FIG. 3 and 4. Capsular cells in dark-field. Granules showing in the cytoplasm, membranelike outline,and middle part of the capsules. X 1,500.
FIG. 5. Noncapsular cells in dark-field. The cell wall is easily seen. X 1,500.FIG. 6. Capsular cells after treatment with capsular serum labeled with fluorescein isothiocyanate. Punc-
tate fluorescence of the capsule can be seen. Fluorescence microscope. X 2,000.FIG. 7. Capsular cells after treatment of the cell-wall serum labeled with fluorescein isothiocyanate.
Fluorescence of transverse septa in the capsule and of punctate strurctures in the membranelike outline ofthe capsules is noticeable. Fluorescence microscope. X 2,000.
1084
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

STRUCTURE AND COMPOSITION OF B. ANTHRACIS CAPSULE
desiccated prel)arations in formalin vapors withmethanol or ethyl alcohol revealed a slightshrinkage of the capsule, compared with livingcells. For study under the fluorescence micro-scope, the preparations were stained with anthraxsera labeled with fluorescein-isothiocyanate.After this treatment, the whole capsule wasbrilliantly fluorescent (Fig. 6), or a weak pune-tate fluorescence of the membranelike outline,the transverse septa in the capsule, and the cellwall was observed (Fig. 7).
Various staining techniques were used instudying fixed capsular cells under the lightmicroscope. On staining with Loffler's blue, themiddle part of the capsule stained metachro-matically a light purple, the membranelikeoutline of the capsule stained dark blue, and theinner part stained violet (Fig. 8). The membrane-like outline of the capsule (Fig. 9, 10, and 29),which sometimes has filaments, is seen whenstained with Toluidine Blue or crystal violetafter treatment with tannin.
Thus, the use of various imethods of lightmicroscopy and specific staining has made itpossible to observe the fine structure of the B.anthracis capsule, which is not revealed whennegatively stained with India ink.
Cytochemical study of the capsule. The use ofcytochemical methods has made it possible todetermine the chemical composition of the B.anthracis capsule, and to establish its stratifiedstructure.
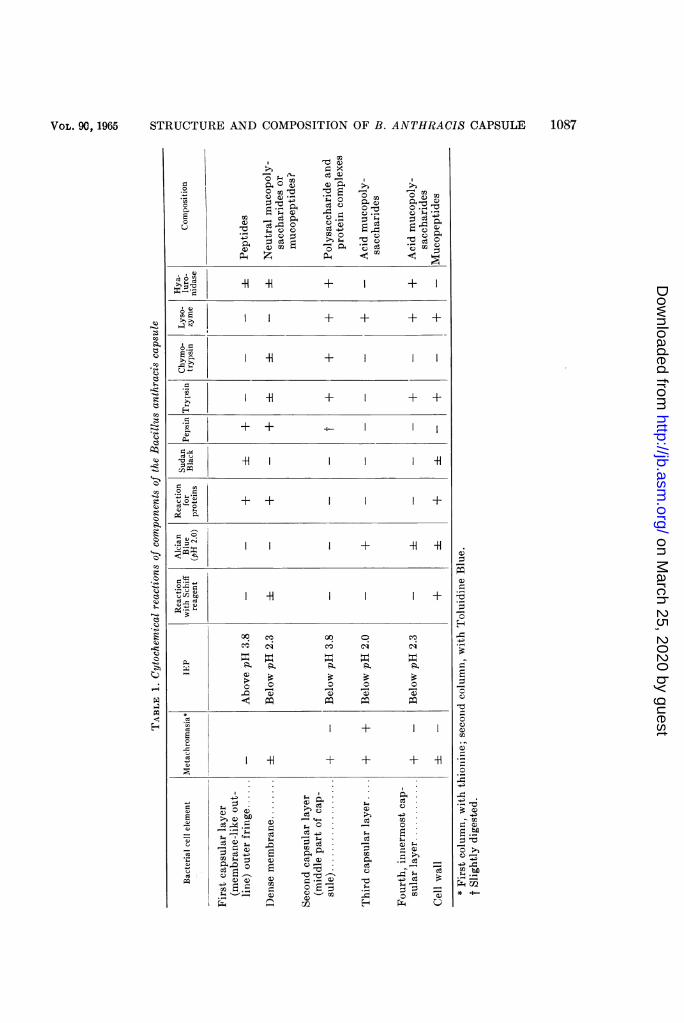
Figure 35 diagrammatically shows the strati-fied structure of the B. anthracis capsule, andTable 1 summarizes the results of the cyto-chemical studies of the various layers of thecapsule, obtained by IEP determination andby cytochemical staining methods specific forpolysaccharides, acid mucopolysaccharides, lipids,and proteins, as well as by treatment of fixedcapsular cells with enzymes.
Staining with Toluidine Bllue at pH 7.0 to 8.0often reveals a delicate outer fringe whichbounds the capsule on the outside and which iseasily destroyed when the preparations arebeing made. This fringe is not metachromaticwhen stained by Toluidine Blue and thionine(which is characteristic of polysaccharides),does not react with Schiff's reagent (test forpolysaccharides), has an IEP at pH above 3.8,does not stain with Alcian 13lue (test for acidmucopolysaccharides), gives positive reactionsfor proteins and lipids, and is destroyed by pepsin(Table 1). All of the foregoing suggests that thispart of the capsule is formed by peptides, lpos-sibly along with lipoproteins.
Directly under the fringe is a thin dense mem-brane which is not metachromatic, weakly re-acts with Schiff's reagent, has an IEP at pH
below 2.3, does not stain with Alcian Blue,gives a positive reaction for proteins, and givesa negative reaction for lipids. This suggests thatthe membrane contains polysaccharide and pro-tein components which are bound to each other,apparently after the fashion of neutral muco-polysaccharides or mucopeptides. The muco-polysaccharides of this membrane resemble themucopeltides of the cell wall, but differ fromthem by their susceptibility to destruction bypepsin and insusceptibility to lysozyme. Theouter fringe and the aforesaid membrane to-gether form the membranelike outline.The second layer forming the main, middle
part of the capsule is located under the meem-branelike outline. This l)art of the capsule is'y-metachromatic when stained with thionine(Fig. 12) and 0-metachromatic when stainedwith Toluidine Bllue; it does not react withSchiff's reagent, has an IEP in the very acidregion (below pH 2.0), stains with Alcian 13lue(Fig. 13), does not give positive reactions forproteins or lipids, and is destroyed by lysozyme.It is well known that, as a rule, acid mueopoly-saccharides do not stain with Schiff's reagentbut are y-metachromatic at pH below 4.0 to 3.0(Pearse, 1960). It may therefore be quite logi-cally assumed that this layer of the capsule isformed by acid mucopolysaccharides.
Lastly, the fourth and innermost layer of thecapsule, directly adjoining the cell wall, is clearly,y-metachromatic in the acid and neutral regions,has an IEP at pH below 2.3, does not react toSchiff's reagent, weakly stains with AlcianBlue, and is destroyed by lysozyme and hya-luronidase. This capsular layer is thus also formedby acid mucopolysaccharides, which differ,however, from the acid mucopolysaccharides ofthe preceding capsular layer by their sensitivityto hyaluronidase and the position of the IEP ina somewhat more alkaline region. By its positionin the cell, this innermost layer of the capsuleresembles the cell wall, but we ascribe it to thecapsule because it is absent in noneapsular formsand stains with crystal violet without prelimi-nary treatment with tannin. (The bacterial cellwall stains, as is well known, only after treatmentwith tannin.)
In addition to the aforementioned layers ofthe B. anthracis capsule, we obsersved delicatetransverse lines which are y-metachromatic atpH 2.3 to 6.0 and are apparently also of a poly-saccharide nature (Fig. 11). Similar lines in thecapsules were observed by Ivanovics and Hor-vath (1953a) in B. megaterium, and by Tomesikand Guex-Holzer (1952) in Bacillus M (an or-ganism allied to B. anthbracis) treated withserum.
Thus, the cytochemical study made it possible
Vo L. 90 s 1965 108.;
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

V
E .ws z1 i Eu .._ ,M,
1 n ..f n
.t "w
s¢#
a S.,2F
*d*.
N
s:.X _ _w
,* '1i v j.tiRl wS|.ey
.s
:t
11 ..tb B:: A
so
'Sw
4 -Alsje ~a
.'
FIG. 8. Capsular cells from an 18-hr culture. Stained with Loffler's blue. Membranelike outline (firstlayer), middle part (second layer), and innermost (fourth layer) part of capsules are seen. Light microscope.X 1,300.
FIG. 9. Capsular cells from a 30-hr culture, stained by Gutstein's method. Several cells with lysed capsuleand cell wall in the middle part of the streptobacillus. Light microscope. X 1,300.
FIG. 10. Capsular cells from an 18-hr culture. Stained with Toluidine Blue at pH 6.0. Membranelikeoutline and innermost (fourth) layer of capsule are seen. Light microscope. X 2,000.
FIG. 11. Capsular cells from an 18-hr culture. Stained with Toluidine Blue at pH 2.3. Transverse linescan be seen in capsule. X 1,700.
FIG. 12. Capsular cells stained by Schmorl's thionine method. Mainly the inner part (third layer) of thecapsule is seen. X 1,500.
FIG. 13. Capsular cells stained with Alcian Blue, also showing the third layer of the capsule. X 1,300.
1086
J
"-mwqwp--
6
1w
.9
'o oo'*F
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
STRUCTURE AND COMPOSITION OF B. ANTHRACIS CAPSULE 1087
"0n()
~$ .05+ -SM
Q. x(:d CL
o)44-ICPv
° 2 ° a) m$=,
COu .
*-s C :s* CQ
C.) C) P4
IC0 7 0coe
2 ~ ~ -°-HI+ + +
oF~~ + + + +2; ~I -H ±
-H + + +
+ + -I+-+-
cu ce d ~I I -H
o 4=
*u)Q + + +cd _ Ce
ccu
oo cACo I
00 0 0
Ce cli Cy: cl
LI +E +
: Q= E-
d E E t ~~~~~c ~o ,r +.4s_o5~~ ~ ~ ~ ~ ~ ~~~r
-5
VOL. 90, 1965
g._._
0CQEc
u
Co
Co
Co0s
0t
0
H)
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

AVAKYAN ET AL.
to reveal the stratified structure of the B. an-thracis capsule, and showed the localization ofvarious substances in the capsule.
Investigation of the capsule by means of electronmicroscopy. Investigation of capsular cells, fixedby formalin and overfixed by osmium, with theaid of an electron microscope has shown thecapsule to consist of a membranelike outline,a middle part, and an inner part. The membrane-like outline of the capsule is observed as a thinosmiophilic layer, but is not seen all along thecell (Fig. 14). Sometimes we observed filamentscoming out from the membranelike outline ofthe capsule. These filaments consist of osmio-philic substance, and show an osmiophobic spiralstructure (Fig. 14, 15). The main middle partof the capsule is more electron-dense than are themembranelike outline and inner part of the cap-sule; it often shows inclusions of less-dense ma-terial in the form of small round or angularbodies (Fig. 14), or sometimes in the form offine transverse septa (Fig. 15).
Negative staining of capsular cells with phos-photungstic acid reveals the structure of theinner capsular layer. It consists of materialwith low electron density, but contains inclusionsof small electron-dense particles (Fig. 16). Verythin filaments extend from this layer into themiddle part of the capsule; these filaments ap-parently create the impression of the transverselines seen in the capsule stained with ToluidineBlue (Fig. 11).
Thus, the studies with the electron microscopehave shown that the different layers of the cap-sule differ in electron density, osmiophilia, andsubmicroscopic structure.
Immunochemical studies of the capsule. Theluminescent sera obtained by immunizing rab-bits with whole capsular cells labeled the capsulebut not the cell wall or the protoplasm. Thisenabled us to give the luminescent serum thecode name of capsular luminescent serum (CLS).The luminescent sera prepared from commercialprecipitable sera selectively labeled the cell wallof noncapsular cells, and on this basis were given
the code name of cell-wall luminescent sera(CWLS).
Studies of CLS-treated capsular cells with thefluorescence microscope have shown the outerfringe and the middle part of the capsule (thefirst and second capsular layers revealed bycytological methods) to be brilliantly fluorescent.The inner part of the capsule adjacent to thecell wall (the third and fourth capsular layers)was not fluorescent (Fig. 19). CLS-treated non-capsular cells were practically nonfluorescent.The cell wall of CWLS-treated noncapsularcells also showed a green fluorescence. The cellwall of capsular cells, the thin fringe on theperiphery of the capsule, and the transverse linestraversing the body of the capsule along thetransverse septa of streptobacilli or between thesepta fluoresced less brightly (Fig. 7, 20). Asalready mentioned, we observed similar trans-verse septa in the capsule with an electronmicroscope (Fig. 15).
In CLS-treated capsular cells examined elec-tron microscopically, we observed the appearanceof an osmiophilic fringe around the capsule. Themiddle part of the capsule acquired a delicateradial stratified structure (Fig. 17). We did notobserve this picture in capsular cells treated withnormal horse serum (Fig. 18).The immunochemical studies have thus shown
heterogeneity of the capsular antigens and haveenabled us to localize them in the capsule. Thecapsule contains antigen complexes consistingof antigens revealed by CLS and homogeneouslydistributed in the outer and middle parts of thecapsule, and antigens revealed by CWLS andlocalized in the membranelike outline and septaof the capsule and in separate punctate structuresin the middle part of the capsule.
Action of enzymes on capsular cells. To studythe action of enzymes on capsular cells of B.anthracis, cytological observations were madewith a light microscope, and antigenic composi-tion was studied with a fluorescence microscope.In the former, the preparations, both treated anduntreated with enzymes, were stained with
FIG. 14 and 15. Capsular cells from an 18-hr culture. Formalin fixation with osmium overfixation. Mem-branelike outline (osmiophobic), as well as middle and inner (also osmiophobic) parts of capsules are seen.Note filaments with spiral structure extending from the membranelike outline of capsules. Electron micro-scope. X 5,600.
FIG. 16. Capsular cells negatively stained with phosphotungstic acid. The structure of the inner layerof the capsules and thin filaments extending into the middle part of capsules is noticeable. Electron micro-scope. X 15,000.
FIG. 17. Capsular cells after treatment with capsular serum. The middle part of the capsules has a deli-cate, radial, stratified structure. The membranelike outline of the capsules is somewhat swollen. Electronmicroscope. X 8,000.
FIG. 18. Capsular cells after treatment with normal horse serum. Electron microscope. X 6,500.
1088 J. BACTERIOL.
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

S t';17 '
AM*8~;/--
iAtt s:?:UP -r.
i:I*
4..
...._1-' ..
VW..,.
I
FIGS. 14-18
1089
t-r-
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

AVAKYAN ET AL.
crystal violet after treatment with tannin (thisstained mainly the membranelike outline and thecell wall) and with Toluidine Blue (this stainedall layers of the capsule). In the latter case, thecontrol preparations and the enzyme-treatedpreparations were stained with CLS (mainlythe middle part and outer fringe of the capsulewere labeled) and with CWVLS (the cell wall,the punctate structures in the membranelikeoutline, and the middle part of the capsule, as wellas the transverse septa of the capsule were labeled).The action of pepsin makes the membranelike
outline and, partly, the middle l)art of the capsuledisappear, the inner part of the capsule and thecell wall remaining intact (Fig. 29, 30, Table 1).In capsular cells treated with CLS after pepsin,the capsule is unlabeled; the inner layer of thecapsule is intact and appears as a dark halo (Fig.19, 21). The cell wall of capsular cells treatedwith CWLS after pepsin shows the same clearfluorescence (Fig. 22) as do preparations un-treated with the enzyme (Fig. 20). The fluorescentpoints in the membranelike outline and the trans-verse septa of the capsule are absent.The action of trypsin destroys primarily the
middle and inner parts of the capsule and thecell wall, especially on the side of the longitudinalsepta; the membranelike outline remains intact(compare Fig. 31 with Fig. 29). In CLS-treatedcells, we observed a slight fluorescence of themain part of the capsule; the fluorescence dis-appeared under prolonged action of trypsin (Fig.23). In CWLS-stained preparations, it is possibleto observe an intensely fluorescent cell wall,fluorescent punctate structures on the peripheryand in the middle part of the capsule, and trans-verse lines in the capsule (Fig. 24). It is undoubt-edly of some interest that, despite the visible
destruction of the cell wall under the action oftrypsin, its fluorescence after treatment withCWLS does not decrease.The action of hyaluronidase is observed to
destroy both the main middle part of the capsuleand its inner part. The membranelike outlineof the capsule usually remains intact. The cellwall is not affected, although the streptobacillioften break up into separate cells owingto destruction of the substance which joins thecells of the streptobacilli (Fig. 32). In CLS-stained preparations, the intact part of thecapsule appears as a brilliantly fluorescent rim,whereas the main part of the capsule does notfluoresce (Fig. 25). In CWLS-treated prepara-tions, the cell wall is brilliantly fluorescent, andtransverse lines and fluorescent points are seenin the middle part of the capsule (Fig. 26).The action of lysozyme is observed to loosen
the entire capsule, although, as a rule, no com-plete lysis of the capsule occurs because themembranelike outline of the capsule remainsintact. The cell wall is also affected (Fig. 33, 34).It is interesting that lysozyme only slightlyaffects the cell wall of noneapsular cells. In CLS-treated preparations, the capsule is fluorescent,but is deformed and is sometimes separated fromthe cell body (Fig. 27). In CWLS-treated prepa-rations, a dim punctate fluorescence of the cellwall is observed (Fig. 28). The periphery of thecapsule sometimes shows a brilliantly fluorescentborder which corresponds to the membranelikeoutline of the capsule. This border is not soclearly observed in control preparations. Thetransverse septa and separate fluorescent pointsin the middle part of the capsule are absent.Other workers also make mention of loosening ofthe cell wall and of lysis of the capsule under the
FIG. 19. Capsular cells from an 18-hr culture, stained with capsular serum labeled with fluorescein iso-thiocyanate. Punctate fluorescence of outer and middle parts of capsules is seen. Fluorescence microscope.X 2,000.
FIG. 20. Capsular cells from an 18-hr culture, stained with cell-wall serum. Note fluorescence of punctatestructures in membranelike outline, middle part of capsules, and in the transverse septa. Fluorescence micro-scope. X 2,000.FIG. 21. Capsular cells stained with CLS after treatment with pepsin. Outer and inner parts of capsules
are nonfluorescent. X 2,000.FIG. 22. Capsular cells stained with CWLS after treatment with pepsin. Cell wall is fluorescent; punctate
structures and septa in the capsules are nonfluorescent. X 2,000.FIG. 23. Capsular cells stained with CLS after trypsin. Middle part of capsules is dimly fluorescent.
X 2,000.FIG. 24. Capsular cells stained with CWVLS after trypsin. X 2,000.FIG. 25. Capsular cells stained with CLS after hyaluronidase. Only outerfringe of capsules is fluorescent.
X 2,000.FIG. 26. Capsular cells stained with CWlLS after hyaluronidase. X 2,000.FIG. 27. Capsular cells stained with CLS after lysozymne. X 2,000.FIG. 28. Capsular cells stained with CWLS after lysozyme. Note dim punctate fluorescence of the cell
walls; transverse septa in capsules and the punctate structures in them are nonfluorescent. X 2,000.
1090 J. BACTERIOL.
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from


-------- - .
- lUe
i- .s-j .|s-vs-
29m h s=f8 tW0 ......................... i30
FIG. 29. Capsular cells from an 18-hr culture stained by Gutstein's method. Cell walls and membranelikeoutline of capsules are stained. Light microscope.VE'I.FG. 30. Capsular cells stained by Gutstein's method after pepsin. Cell walls intact, outer part of capsulesdestroyed. X 1,300.
FIG. 31. Capsular cells stained by Gutstein's method after trypsin. Destruction of inner and middle partsof capsules and of cell walls in evidence. X 1,300.
FIG. 32. Capsular cells stained by Gutstein's method after hyaluronidase. Destruction of capsules andsubstance joining the cells into streptobacilli are seen. X 1,300.
FIG. 33 and 34. Capsular cells stained by Gutstein's method after lysozyme. Capsules and cell wallsdestroyed. X 1,300.
1092
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

STRUCTURE AND COMPOSITION OF B. ANTHRACIS CAPSULE 1093
cytop/osvna._,- cell wall
'- two inner/ayersof capsule
- Mr;od/de ,arroKs- meindroee l;*e
autiin-of CapSule
FIG. 35. Diagrammatic representation of theBacillus anthracis capsule.
action of lysozyme and trypsin in B. anthracisand allied spore-bearing aerobic microorganisms(Tomcsik and Guex-Holzer, 1952; Nordbergand Thorsell, 1955).
Thus, our cytological and immunochemicalstudies of the action of enzymes on capsular cellshave enabled us to ascertain the chemical com-
position of the different layers of the capsule,and to differentiate the capsular antigens ac-cording to their relations to enzymes. We haveshown, for example, that the capsular antigensmay be divided into at least two groups: thesuperficial antigens of the capsule, revealed byCLS and localized in the outer fringe of thecapsule, possess selective sensitivity to pepsin;the antigens of the capsule proper, revealed bythe action of CLS and CWLS and concentratedin the middle part of the capsule, are destroyedby hyaluronidase, lysozyme, and proteolyticenzymes.
DISCUSSION
The present study is one of the few attemptsto investigate the structure and compositionof bacterial capsules by cytological, cytochemical,and immunochemical methods. It is no mereaccident that the capsule of B. anthracis waschosen as the model for this study. It is wellknown that the capsule of this microorganismhas a complex composition, which includespolysaccharide as well as peptide components(Tomesik, 1954; Nordberg and Thorsell, 1955),and that the antigens of the capsule in largemeasure determine the antigenic properties ofthe causative agent of anthrax (Tomcsik, 1956).To investigate the capsular structure of this
microorganism, we used both light and electronmicroscopes. Our investigations have shownthe B. anthracis capsule to have a complexstructure with a well-defined membranelikeoutline, which is observed by phase contrast inliving cells and with electron and light micro-scopes in fixed preparations. This is at variancewith Tomcsik's (1956) statement to the effectthat the existence of the outer membranelike
outline is doubtful. We cannot fully agree withTomesik's (1956) and Duguid's (1951) opinionsabout the inferior character of the cytologicalstudies conducted with fixed capsular bacteria.As our investigations of the capsular forms ofB. anthracis and the investigations of the capsulesof other bacteria (Klieneberger-Nobel, 1948;Katz, 1964; Katz, Solovyov, and Volkova, Zh.Microbiol. Epidemiol. i Immunobiol., in press)have shown, the studies of capsules in fixed prepa-rations extend the present state of our knowledgeof the structure and composition of bacterialcapsules.To study the chemical composition of the B.
anthracis capsule, we used cytochemical methodswhich enabled us to ascertain the chemical na-ture of the capsule and to locate the varioussubstances in it, which could not be done bychemical methods alone. It was thus shown thatthe B. anthracis capsule has a stratified structure,the various layers of the capsule differing intheir IEP, sensitivity to enzymes, and, conse-quently, in their chemical composition. Earlierliterature gave contradictory indications con-cerning the position of the IEP in the B. anthraciscapsule. For example, Tomcsik and Guex-Holzer (1953), who studied the interaction of thecapsule with various proteins, determined theposition of the IEP at pH 2.0, whereas Ivanovicsand Horvath (1953b) established its position,with the aid of stains, below pH 3. Lastly, ac-cording to electrophoresis, the IEP of the B.anthracis capsule is at pH 3.1 (Harden, quotedby Tomcsik and Guex-Holzer, 1954a). Thesedata apparently pertain to different parts of thecapsule, which would explain such considerabledifferences in IEP.To study the antigenic composition of the
capsule and cell wall of B. anthracis and alliedspecies, most investigators have employed eitherchemical or immunochemical methods of re-search. However, the chemical methods of re-search resulted in mechanical destruction ofthe cell, and therefore made it impossible to lo-cate the antigens in the intact capsular cell. Theimmunological investigations were conductedmainly on living capsular cells treated with"polypeptide" and "polysaccharide" sera; thesestudies revealed the topographical distributionof the "polysaccharide" and "polypeptide"antigens in the capsules of B. megaterium andBacillus M, species closely related to B. anthracis,thereby making a definite contribution to ourknowledge of the fine structure and antigeniccomposition of bacterial capsules (Ivanovics andHorvath, 1953a; Tomesik and Guex-Holzer,1954b; Baumann-Grace and Tomesik, 1958).However, the terms "polypeptide serum" and
VOL. 90, 1965
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

AVAKYAN ET AL.
"polysaccharide serum" suggested by theseinvestigators do not reflect either the morphologi-cal localization of the corresponding antigensin the cell or the chemical nature of these anti-gens, since the capsule and cell wall of thesemicroorganisms contain l)olysaccharide andlprotein components. It would be more ap)pro-priate to use the ternis "calpsular antibody serum"and "cell-wall antibody serum," which was donein the present study.To study the arrangement of antigens in the
B. anthracis capsule, we used the fluorescent-antibody method; to investigate the chemicalnature of antigens, we used treatment withenzymes. As is well known (Dixon and W1'ebb,1958), pepsin hydrolyzes only the peptide hondsof carboxyl groups which belong to aromaticamino acids, especially if glutamyl is the adjacentamino acid residue. Trypsin and chymotrypsinhydrolyze not only leptide, but also ether bonds.Hyaluronidase acts on acid mucopolysaccharideswhich contain hyaluronic acid and chondroitin-sulfate (Dixon and Wrebb, 1958). Lysozvineapparently hydrolyzes the bond between A-ace-tylmuramic acid and 2-acetylamino-2-deoxy-D-glucose in mucopolysaceharides and mucopep-tides (Report of the Commission on Enzymes ofthe International Union of Biochemistry, 1961).Thus, the use of enzymes makes it possible todetermine the chemical nature of antigens in theintact bacterial cell. Our overall study has shownthat the outer frinoe of the capsule contains so-called superficial capsular antigens, revealed byCLS; these antigens are of a peptide nature anddetermine the immunochemical specificity of thecapsule. The dense membrane of the capsule,which together with the foreg'oing fringe formsthe membranelike outline of the capsule, con-tains antigens closely related in coml)osition tothe antigens of the cel] wall. Like the antigensof the cell wall, they are revealed by CWLS, butdiffer from them in that they are insensitive tolysozyme and sensitive to pepsin. The middlepart of the capsule contains antigens of the cap-sule proper; these antigens are revealed by bothCLS and CWLS, and contain polysaccharide andprotein components (digested by lysozyme andproteolytic enzymes). Lastly, the inner part ofthe capsule, formed by acid mucopolysaccharides,either contains no antigens at all or contains anti-gens which are not revealed by the methods wehave used. Thus, as regards antigens, the B.anthracis capsule is heterogeneous and containssubstances of both protein and pJolysaccharidenature. However, the antigenic specificity ofthe capsule is determined only by pelptides whichpossess selective sensitivity to leIpsin. tIsingimmunochemical methods of studying nonfixed
bacteria, Ivanovics and Horvath (1953b),Toincsik (1956), and others have also shown thatthe antigenic specificity of the B. anthracis andB. negaterinm calsules is dletermined by glutamyllpolypel)tide.Our overall immunochemical and cytochemi-
cal studies have also enabled us to obtain newdata on antigenic an(d other lproperties of thecell wall of cal)sular and noncapsular forms of B.anthlracis. For examl)le, it was shown that thecell wall of capsular cells is poorly seen in dark-field illuminatioin, comnpared with the cell wall ofnoncap)sular forms, that it is more sensitive tolysozyine, and that it possesses weaker fluo-rescence when stained with CWLS obtaine(d byimmunization of animals with noncapsularcells of B. anthracis. All this warrants the as-sumlj)tion that there are significant differencesbetween the chemical composition and antigeniclproperties of the cell wall of calpsular and non-
calpsular form.s of B. anthracis. It is also lpossible,however, that the revealed differences are dueto the fact that the cal)sule l)revents enzyme andserum from penetrating into capsular cells.
It is also of some interest that the morphologi-cal destruction of the cell wall is not always ac-coml)anied by disturbances in its antigens.Under the action of lysozyme, B. anthracis ex-hibits morphological destruction of the cell walland a change in its antigenic l)rolerties (weak-ened staining when treated with CWrLS). Underthe action of trypsin, the morphological destruc-tion of the cell wall is not accompanied by achange in its antigenic properties. This suggeststhat in B. anthracis the antigenic l)rolertiesof the cell wall are, unlike those of the capsule,determiiined not by a protein, but by a l)olysac-charidle component. Guex-Holzer and Tomesik(1956) and Tomcsik (1956) similarly indicatedthat the serological specificity of the BacillusM cell wall is also determined by a polysaccharidecomponent.
LITERATrURE CITrED
ALFERT, M. andC I. J. GESCHWIND, 1953. A selec-tive staininlg method for the basic proteins ofcell nuclei. Proc. Natl. Acad. Sci. U.S. 39:991.
ARKHIPOVA, V. 11. 1962. The use of niutrienit me-dium for early diagntosis of capsular forms ofanithrax [in IRussian]. Probl. Vet. Sci. Sanit.21 :171-175.
BAUMANN-G1RACE, J. B., AND J. TomcSIKs. 1958.Elektronenmikroskopische Untersuchung derkomplexen Kapselstruktur bei B. megaterium.Schweiz. Z. Allgem. Pathol. Bakteriol. 21:906-914.
BLOCII, 1). P., AND 11. I. HEW. 1960. Changes innuclear histons during fertilization, and earlyembryonic development in pulmonate sniail
1094 J. BA~CTERIOL.
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 90, 1965 STRUCTURE AND COMPOSITION OF B. ANTHRACIS CAPSULE 1095
Helix aspera. J. Biophys. Biochem. Cytol. 8:69-82.
COONS, A. H., AND M. H. KAPLAN. 1950. Localiza-tion of antigen in tissue cells. II. Improvementsin a method for the detection of antigen bymeans of fluorescent antibody. J. Extl. Med.91 :1-13.
DIXON, M., AND E. C. WEBB. 1958. Enzymes.Academic Press, Inc., New York.
DUGUID, I. P. 1951. The demonstration of bac-terial capsules and slime. J. Pathol. Bacteriol.63:673-685.
GUEX-HOLzER, S., AND J. TOMCSIK. 1956. Isolationand chemical nature of capsules and cell-wallhaptens in a Bacillus sp. J. Gen. Microbiol.14:14-21.
GUTSTEIN, M. 1924. Das Ektoplasma der Bak-terien. Zentr. Bakteriol. Parasitenk. Abt. IOrig. 93:393-402.
IVANOVICS, G., AND S. HORVATH. 1953a. Thestructure of the capsule of B. megaterium. ActaPhysiol. Acad. Sci. Hung. 4:175-183.
IVANOVICS, G., AND S. HORVATH. 1953b. On thechemical structure of the capsulae of B. anthra-cis and B. megaterium. Acta Physiol. Sci. Hung.4 :401-408.
KATZ, L. N. 1964. A cytological and cytochemicalstudy of the capsule and cell-wall of B. anthra-cis [in Russian]. Mikrobiologiya 33:836-843.
KEPPIE, J., AND H. SMITH. 1963. The chemicalbasis of the virulence of Bacillus anthracis. IX.Its aggressius and their mode of action. Brit.J. Exptl. Pathol. 44:446-453.
KLIENEBERGER-NOBEL, E. 1948. Capsules andmucoid envelopes of bacteria. J. Hyg. 46:345-348.
LEVINA, K. N., AND V. R. ARKHIPOVA. 1964. Pro-duction of capsular Anthrax sera and determina-tions of their immunological activity [in Rus-sian]. Lab. Delo 10:619-621.
NORDBERG, B. K., AND W. THORSELL. 1955. Theeffect of certain enzyme systems on the capsuleof Bacillus anthracis. J. Bacteriol. 69:367-371.
PEARSE, A. G. E. 1960. Histochemistry: theoreti-
cal and applied, 2nd ed. Little, Brown and Co.,Boston.
PISCHINGER, A. 1926. Die Lage des isoelektrischenPunktes histologischer Elemente als Ursacheihrer verschiedenen Farbbarkeit. Z. Zellforsch.3:169-197.
RIGGS, J. L., R. J. SEIWALD, J. H. BURCKHALTER,C. M. DOWNS, AND T. G. METCALF. 1955. Iso-thiocyanate compounds as fluorescent labelingagents for immune serum. Am. J. Pathol.34:1081-1097.
ROTH, I. L., C. W. LEVIS, JR., AND R. P. WILLIAMS.1960. Electron microscope study of Bacillusanthracis in mouse spleen. J. Bacteriol. 80:772-782.
ROTH, I. L., AND R. P. WILLIAMS. 1964. Nature ofthe cytopathic area surrounding virulent cellsof Bacillus anthracis in mouse spleen. J. Bac-teriol. 88:523-530.
THORNE, C. 1956. Capsule formation and synthesisof glutamyl polypeptide by Bacillus anthracisand Bacillus subtilis. Symp. Soc. Gen. Micro-biol. 6:68-80.
ToMcSIK, J. 1954. Ueber die Oberfliachenstruk-turen des Bacillus anthracis. Schweiz. Z.Allgem. Pathol. Bakteriol. 17:457-468.
ToMcSIK, J. 1956. Bacterial capsules and their re-lations to the cell wall. Symp. Soc. Gen. Micro-biol. 6:41-67.
ToMcSIK, J., AND S. GUEX-HOLZER. 1952. Ande-rung der Struktur der Bakterienzelle im Verlaufder Lysozym-Einwirkung. Schweiz. Z. Allgem.Pathol. Bakteriol. 15:517.
ToMCSIK, J., AND S. GUEX-HOLZER. 1953. EinPrinzip zur Fiirbung der Bakterienkapsel.Schweiz. Z. Allgem. Pathol. Bakteriol. 16:882-891.
ToMCSIK, J., AND S. GuEx-HOLzER. 1954a. Demon-stration of the bacterial capsule by means of apH-dependent salt-like combination with pro-teins. J. Gen. Microbiol. 10:97-109.
ToMcsIK, J., AND S. GUEX-HOLZER. 1954b. Geneseder komplexen Kapselstructur. Schweiz. Z.Allgem. Pathol. Bakteriol. 17:221-240.
on March 25, 2020 by guest
http://jb.asm.org/
Dow
nloaded from