structure of the ovarian lactogen receptors. analysis with
TRANSCRIPT

Vol. 259, No. 7. Issue of April 10, pp. 4542-4549. 1984 Printed in U S A.
Structure of the Ovarian Lactogen Receptors ANALYSIS WITH BIFUNCTIONAL CROSS-LINKING REAGENTS*
(Received for publication, April 13, 1983)
Juan S. Bonifacino and Maria L. Dufau From the Section on Molecular Endocrinology, Endocrinology and Reproduction Research Branch, National Institute of Child Health and Human Deuelopment, National Institutes of Health, Bethesda, Maryland 20205
‘251-H~man growth hormone was cross-linked with high efficiency to lactogen receptors in membranes and in Triton X-100-solubilized preparations from lutein- ized rat ovaries using the bifunctional reagent disuc- cinimidyl suberate. Cross-linked samples were sub- jected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis under reducing conditions followed by autoradiography of dried gels. Analysis of cross-linked hormone-receptor complexes in membranes revealed the presence of a major radioactive band at M. = 60,000. Cross-linking of detergent-solubilized com- plexes resulted in the appearance of two major radio- active bands of M, = 60,000 and M, = 100,000. Excess unlabeled lactogenic hormones inhibited the labeling of the major bands while non-lactogenic hormones had no effect. The same major radioactive bands were ob- served when the cross-linked samples were analyzed under nonreducing conditions. However, in solubilized preparations, the ratio between the intensities of the lower and the higher M, bands was markedly lower under nonreducing than under reducing conditions. Two-dimensional sodium dodecyl sulfate-polyacryl- amide gel electrophoresis studies (first dimension, non- reducing; second dimension, reducing) demonstrated that a species with M, = 60,000 can be released from the M, = 100,000 species upon cleavage of disulfide bonds. Since the M, of ‘251-h~man growth hormone is 22,000, these results suggest that the ovarian lactogen receptors are molecules with an approximate M, of 80,000, containing a subunit with a M, of approxi- mately 40,000 bearing the recognition site for the hormone. Some of these subunits appear to be linked to the rest of the molecule by disulfide bonds.
The presence of prolactin receptors in the ovary has been demonstrated in various animal species, including rats (1-6), cows (1, 7), pigs (8, 9), and humans (1, 7, 10). Using autora- diographic (11) and immunocytochemical (12) techniques and direct binding measurements to different fractions isolated from rat ovary (a) , prolactin receptors were found to be predominantly located in the corpora lutea. Granulosa cells of medium and large sized follicles have also been shown to possess a small number of prolactin receptors (2 , 12, 13). Hormonal induced maturation of granulosa cells and their subsequent transformation into luteal cells are accompanied by a marked increase of the content of prolactin receptors (2, 13-15). An intense luteinization of the ovaries can be induced
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
~ _ _ _ ~ ~~~~ ~~ ~~ ~~
in uiuo by sequential treatment of immature rats with preg- nant mare serum gonadotropin and human chorionic gonad- otropin. The resulting superovulated ovaries weigh 8-9 times more than normal ovaries, and membranes prepared from them show an enhanced binding capacity for lactogenic hor- mones (16). Hence, luteinized ovaries are an excellent source of, prolactin receptors for studies on their physical-chemical and structural characteristics. These receptors can be ex- tracted from membranes in an active form with 1% (w/v) Triton X-100, and such treatment causes the unmasking of a population of cryptic receptors. As a consequence, the solu- bilized preparations display a binding capacity 6 times higher than that of the membranes (6). To initiate studies on the characteristics of the molecular components involved in the binding of lactogenic hormones in the ovary, we employed bifunctional cross-linking reagents to covalently attach the labeled hormones to the receptor proteins. This approach was originally used in the study of insulin receptors (17-21) and proved to be successful in the molecular characterization of various other hormone receptors (22-29).
MATERIALS AND METHODS
Hormones and Chemicals-hGH’ (NIADDK-hGH-I-l), oPRL (NIH-P-S-lo), hPL, bGH (NIH-GH-B-18), oLH (NIAMDD-oLH- 23), oFSH (NIH-FSH-l5), and rTSH (rTSH-RP-1) were obtained from the Hormone Distribution Program, National Institute of Ar- thritis, Metabolism, and Digestive Diseases. DSS and di- thiobis(succinimidy1 propionate) were purchasedfrom Pierce Chem- ical Co. Phenylmethylsulfonyl fluoride was from Boehringer Mann- heim. Trypsin inhibitor, leupeptin, aprotinin, Nu-p-tosyl-L-lysine chloromethyl ketone, N-ethylmaleimide, and TritonX-100 were from Sigma. The molecular weight standards and reagents for sodium dodecyl sulfate-polyacrylamide gel electrophoresis wereobtained from Bio-Rad.
Iodination of Hormones-hGH and oPRL were labeled with lZ5I by a modification of the lactoperoxidase method (30) as described pre- viously for human chorionic gonadotropin (31). Purification of the tracers was performed on a Sephadex G-100column (1.5 X 90 cm) equilibrated and eluted a t 4 “C with 50 mM sodiumphosphate buffer, pH 7.4, containing 0.1% (w/v) bovine serum albumin. The specific activity of the labeled hormones ranged from 30 to 74 pCi/pg.
Preparation of Ovarian Membranes-Superovulated ovaries, ob- tained as described before (6), were finely minced with scissors and homogenized in 10 volumes (v/w) of ice-cold 0.25 M sucrose, 0.1 mg/ ml of trypsin inhibitor by means of a Potter-Elvehjem homogenizer fitted with a motor-driven Teflon pestle (10 strokes of 2,100 rpm). All subsequent fractionation procedureswere performed a t 4 “ C . The homogenate was centrifuged for 10 min at 1,500 x g(Beckman J-62B centrifuge, JS-4.2 rotor, 2,750 rpm). The resulting supernatant was decanted and recentrifuged for 30 min a t 34,000 x g (Beckman L5- 65 ultracentrifuge, type 60 Ti rotor, 22,000 rpm). The supernatant
’ The abbreviations used are: hGH, human growth hormone; oPRL, ovine prolactin; hPL, human placental lactogen; bGH, bovine growth hormone; oLH, ovine luteinizing hormone; oFSH, ovine follicle- stimulating hormone; rTSH, rat thyroid-stimulating hormone; DSS, disuccinimidyl suberate; PBS, phosphate-buffered saline.
4542
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Structure of Ovarian Lactogen Receptors 4543
was discarded and the pellet was resuspended in 0.5 ml of homoge- nization solution/g of ovary.
In some experiments, fresh luteinized ovaries were homogenized in ice-cold 0.25 M sucrose solution containing a mixture of protease inhibitors that included 0.1 mg/ml of trypsin inhibitor, 1 mM leupep- tin, 1 mM phenylmethylsulfonyl fluoride, 2 mM N"-p-tosyl-L-lysine chloromethyl ketone, and 10 trypsin inhibitor units/ml (9000 Kalli- krein inhibitor units/ml) of aprotinin. The homogenate was subjected to a fractionation procedure similar to that described above. The resulting crude membrane pellet was resuspended in homogenization solution containing protease inhibitors. In some other experiments, crude membranes were obtained from fresh ovaries including 20 mM N-ethylmaleimide in the homogenization solution. Purified plasma membranes were prepared as described by Gospodarowicz (32).
All membrane suspensions were fractionated in 0.5-ml aliquots and stored a t -80 "C until used. The concentration of proteins of each preparation was determined by the method of Lowry et al. (33). 5'-Nucleotidase (EC 3.1.3.5) was assayed according to Heppel and Hilmoe (34). The specific activity of 5'-nucleotidase/unit of protein in crude membrane preparations was 2-3 times higher than that in the homogenate. Preparation of purified membranes brought about a 9-12-fold increase of the specific activity of 5'-nucleotidase/unit of protein with respect to the homogenate.
Solubilization of Ouarian Membranes-An aliquot of the crude membrane suspension (about 2.4 mg of proteins in 1.2 ml) was diluted 1/10 with Dulbecco's PBS and centrifuged at 4 "C for 30 min a t 34,000 X g (Beckman L5-65 ultracentrifuge, type 60 Ti rotor, 22,000 rprn). The pellet was resuspended in 12 ml of ice-cold Dulbecco's PBS containing 1% (w/v) Triton X-100 by means of a Dounce-type homogenizer (five strokes, tight pestle). This suspenion was centri- fuged a t 4 "C for 90 min a t 240,000 X g (Beckman L5-65 ultracentri- fuge, type 65 rotor, 61,000 rprn). After removing the lipids floating on top, the supernatant was decanted. Aliquots (0.5 ml) of the solubilized extract were stored at -80 "C until used.
Binding of '251-hCH to Membranes-Unless otherwise indicated, binding of 12sII-hGH to membranes was performed as follows. Crude membranes (200-480 pg of proteins) were incubated with lz5I-hGH (0.6-1.5 X lo6 cpm, final concentration 1.2-4.5 nM) for 5 h a t 25 "C in 0.3 ml of Dulbecco's PBS, pH 7.4, containing 0.1% (w/v) bovine serum albumin. After incubation, samples were diluted with 10 ml of ice-cold Dulbecco's PBS and centrifuged at 4 "C for 30 min a t 14,500 X g (Sorvall RC-5B centrifuge, SM-24 rotor, 12,000 rpm). The pellet was washed once with 10 ml of the same buffer and centrifuged under identical conditions. The washed membranes were then resuspended in 0.4 ml of Dulbecco's PBS by means of a Dounce-type homogenizer (five strokes, tight pestle), counted for radioactivity, placed on an ice bath, and subjected to cross-linking (see below). In some cases, membranes prelabeled with '2sI-hGH were treated with 1% (w/v) Triton X-100 prior to cross-linking. Specific binding was calculated as the difference between the radioactivity bound in the absence and in the presence of 5 pg of unlabeled hGH, determined in parallel incubations. Under these conditions, specific binding of Iz5I-hGH to membranes ranged from 11 to 23% of the total radioactivity added.
Binding of '2s1-hCH to the Solubilized Extract-Solubilized extract (61-67 pg of proteins) was incubated with '2sI-hGH (1.3-2.0 X lo5 cpm, final concentration 0.5-2.1 nM) for 5 h at 25 "C in 0.1 ml of Dulbecco's PBS, pH 7.4, containing 0.1% (w/v) bovine serum albu- min. Unlike experiments with membranes, unbound 'T-hGH was not removed after incubation and the mixture was directly subjected to cross-linking. Specific binding was determined in parallel incuba- tions by polyethylene glycol precipitation (6). Under these conditions, specific binding was found to range from 56 to 62% of the total radioactivity added.
Cross-linking of Hormone-Receptor Complexes-The standard pro- cedure employed for cross-linking was as follows. Freshly prepared 25 mM DSS in dimethyl sulfoxide was added to prelabeled membrane (in 0.4 ml) or solubilized (in 0.1 ml) preparations to a final concen- tration of 0.5 mM. Samples were then incubated for 15 min at 0 "C. The reaction was terminated by the addition of 0.15 volumes of ice- cold 1 M Tris/HCl, pH 7.4. The solubilized preparation was then prepared for analysis by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (see below). Cross-linked membranes were diluted with 4 ml of Dulbecco's PBS and centrifuged for 30 min a t 2000 x g (Beckman J-6B centrifuge, JS-4.2 rotor, 3200 rprn). The pellet was then subjected to sodium dodecyl sulfate-polyacrylamide gel electro- phoresis. Conditions of cross-linking varying from those described above are specified in the figure legends.
Electrophoresis and Autoradiography-Cross-linked membranes and solubilized preparations were heated at 100 "C for 15 min in 60 mM Tris/HCl, pH 6.8, 10% (v/v) glycerol, 2% (w/v) sodium dodecyl sulfate, 5 mM EDTA in the presence or in the absence of 0.7 M mercaptoethanol. Samples were analyzed by sodium dodecyl sulfate- polyacrylamide slab gel electrophoresis using the discontinuous buffer system described by Laemmli (35). Resolving gels contained 7.5% (w/ v) acrylamide (41:l acrylamide/bisacrylamide ratio) and were 3 mm thick. Electrophoresis was carried out for 15 h at 13 mA/gel constant current. After the dye (bromphenol blue) front reached the bottom of the gel, the electrophoresis was allowed to continue for 30 additional min. This permitted most of the free '"I-hGH to run off the gel. Gels were stained with 0.2% Coomassie brilliant blue R-250 in 50% meth- anol, 10% acetic acid followed by destaining with 25% methanol, 7.5% acetic acid. The gel slabs were then dried and subjected to autoradiography with Kodak X-Omat AR film. Standards used in electrophoresis and their molecular weights were myosin heavy chain (200,000), 0-galactosidase (116,250), phosphorylase b (92,500), bovine serum albumin (66,200), ovalbumin (45,000), and carbonic anhydrase (31,000). For quantitation of the amount of labeled hormone cova- lently coupled to the receptor preparations, the cross-linking samples were electrophoresed as described above, the resulting gels were sliced, and the fractions were counted for radioactivity. All the radioactivity detected at molecular weights higher than that of the free labeled hormone was added and related to the amount of hormone-receptor complexes to yield the efficiency of the cross-linking reaction. In some cases, cross-linked samples were subjected to two-dimensional electrophoresis. The technique employed is described in the legend to Fig. 4.
RESULTS
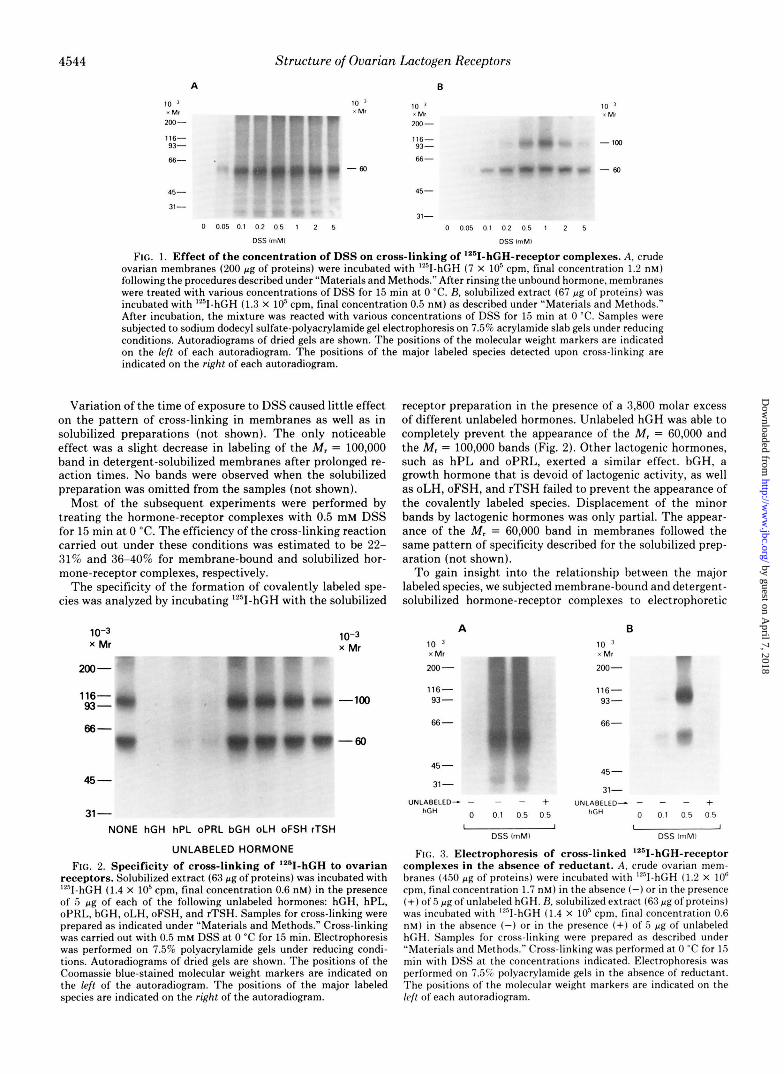
Initial studies were conducted to explore the conditions for cross-linking of membrane-bound or solubilized hormone- receptor complexes. Membranes labeled with lz5I-hGH were treated with increasing concentrations of DSS for 15 min at 0 "C and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis under reducing conditions. Autoradi- ographs of the gels showed the presence of a major band corresponding to a species with a molecular weight of approx- imately 60,000 (Fig. 1A). The extent of labeling of this band was found to be a function of the DSS concentration, reaching a maximal level at 0.5 mM DSS. At higher concentrations of cross-linker, the intensity of this band decreased slightly. Several minor bands with approximate M , = 100,000,82,000, 40,000, 37,000, and 30,000 were usually observed. At a DSS concentration of 0.5 mM, the M , = 30,000 band showed 3-5% and the M , = 37,000 band showed 1-2% of the intensity of the Mr = 60,000 band, as determined by densitometric meas- urements of the autoradiograms. All the other bands were less than 1% as intense as the major band. Cross-linking of detergent-solubilized lZ5I-hGH-receptor complexes resulted in the appearance of two major bands (Fig. 1B). One of these bands corresponded to a species with M , = 60,000, similar to that observed in cross-linked membranes. The other band had an approximate M, of 100,000. The extent of labeling of both bands was dependent upon the concentration of DSS (Fig. 1B). Labeling of the Mr = 60,000 band progressively increased reaching a maximum at 0.5 mM DSS. This level was maintained with concentrations of DSS up to 2 mM. Higher concentrations (5 mM) caused a slight decrease in the intensity of this band. Optimal conditions for labeling the M , = 100,000 band were found over a narrower range of DSS concentrations (Fig. 1B). At 0.1 and 0.2 mM, this band ap- peared faint in the autoradiograms. Its intensity increased at 0.5 mM DSS, reaching a maximum at 1 mM DSS. Higher concentrations of the cross-linker caused a marked decrease in the radioactivity associated with the Mr = 100,000 band. Minor bands having the same M , as in membranes were also detected. The radioactivity background of the gels was lower when solubilized preparations were used.
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4544 Structure of Ovarian Lactogen Receptors
A B
10 3 x Mr 200 - 21r- 116- 93-
66-
45-
116- 93 - 66-
K* 1pI rg .... - 60 45-
- loo
"60
31-
0 0.05 0.1 0 2 0 5 1 2 5 31-
0 0.05 0.1 0.2 0.5 1 2 5
OSS ImM) OSS ImMl
FIG. 1. Effect of the concentration of DSS on cross-linking of I2"I-hGH-receptor complexes. A, crude ovarian membranes (200 pg of proteins) were incubated with IZ5I-hGH (7 X IO5 cpm, final concentration 1.2 nM) following the procedures described under "Materials and Methods." After rinsing the unbound hormone, membranes were treated with various concentrations of DSS for 15 min at 0 "C. B, solubilized extract (67 pg of proteins) was incubated with "'I-hGH (1.3 X lo5 cpm, final concentration 0.5 nM) as described under "Materials and Methods." After incubation, the mixture was reacted with various concentrations of DSS for 15 min at 0 "C. Samples were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 7.5% acrylamide slab gels under reducing conditions. Autoradiograms of dried gels are shown. The positions of the molecular weight markers are indicated on the left of each autoradiomam. The Dositions of the major labeled species detected upon cross-linking are indicatedon the right of each autoradiogram.
Variation of the time of exposure to DSS caused little effect on the pattern of cross-linking in membranes as well as in solubilized preparations (not shown). The only noticeable effect was a slight decrease in labeling of the M, = 100,000 band in detergent-solubilized membranes after prolonged re- action times. No bands were observed when the solubilized preparation was omitted from the samples (not shown).
Most of the subsequent experiments were performed by treating the hormone-receptor complexes with 0.5 mM DSS for 15 min a t 0 "C. The efficiency of the cross-linking reaction carried out under these conditions was estimated to be 22- 31% and 36-40% for membrane-bound and solubilized hor- mone-receptor complexes, respectively.
The specificity of the formation of covalently labeled spe- cies was analyzed by incubating '*'I-hCH with the solubilized
10-3 X Mr
200-
116- 93-
66-
10-3 X Mr - . ""
"100
w p -60
45-
31 - NONE hGH hPL oPRL bGH oLH oFSH rTSH
UNLABELED HORMONE FIG. 2. Specificity of cross-linking of I2"I-hGH to ovarian
receptors. Solubilized extract (63 pg of proteins) was incubated with 'YsII-hGH (1.4 X lo5 cpm, final concentration 0.6 nM) in the presence of 5 p g of each of the following unlabeled hormones: hGH, hPL, oPRL, bGH, oLH, oFSH, and rTSH. Samples for cross-linking were prepared as indicated under "Materials and Methods." Cross-linking was carried out with 0.5 mM DSS at 0 "C for 15 min. Electrophoresis was performed on 7.5% polyacrylamide gels under reducing condi- tions. Autoradiograms of dried gels are shown. The positions of the Coomassie blue-stained molecular weight markers are indicated on the left of the autoradiogram. The positions of the major labeled species are indicated on the right of the autoradiogram.
ab,@ "Y
receptor preparation in the presence of a 3,800 molar excess of different unlabeled hormones. Unlabeled hGH was able to completely prevent the appearance of the M, = 60,000 and the M, = 100,000 bands (Fig. 2). Other lactogenic hormones, such as hPL and oPRL, exerted a similar effect. bGH, a growth hormone that is devoid of lactogenic activity, as well as oLH, oFSH, and rTSH failed to prevent the appearance of the covalently labeled species. Displacement of the minor bands by lactogenic hormones was only partial. The appear- ance of the M, = 60,000 band in membranes followed the same pattern of specificity described for the solubilized prep- aration (not shown).
To gain insight into the relationship between the major labeled species, we subjected membrane-bound and detergent- solubilized hormone-receptor complexes to electrophoretic
A B 10 ' 10 x Mr " X MI -
200- 200-
116- 93 - 66 -
45 - 31-
116- 93 - d#
66 -
45-
31- UNLABELED- - - - UNLABELED- - - - +
hGH 0 0.1 0.5 0.5 hGH 0 0.1 0.5 0 5 I 1
DSS lmMl I I
DSS lmMl
FIG. 3. Electrophoresis of cross-linked I2"I-hGH-receptor complexes in the absence of reductant. A, crude ovarian mem- branes (450 pg of proteins) were incuhated with '"I-hGH (1.2 X lo6 cpm, final concentration 1.7 nM) in the absence (-)or in the presence (+) of5 pg of unlabeled hGH. H. solubilized extract (63 pg of proteins) was incubated with '"I-hGH (1.4 X lo5 cpm, final concentration 0.6 nM) in the absence (-) or in the presence (+) of 5 pg of unlabeled hGH. Samples for cross-linking were prepared as described under "Materials and Methods." Cross-linking was performed at 0 "C for 15 min with DSS at the concentrations indicated. Electrophoresis was performed on 7.5% polyacrylamide gels in the absence of reductant. The positions of the molecular weight markers are indicated on the kfl of each autoradiogram.
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Structure of Ovarian Lactogen Receptors 4545
analysis under nonreducing conditions. When 1251-hGH cross- linked to the membrane preparation with 0.5 mM DSS was analyzed by sodium dodecyl sulfate-polyacrylamide gel elec- trophoresis under nonreducing conditions (Fig. 3A), a band of M , = 60,000 was preferentially labeled. The radioactivity incorporated into this band increased as the concentration of DSS was increased from 0.1 to 0.5 mM. After prolonged exposure times, minor bands with approximate M , = 100,000, 75,000, 37,000, and 30,000 could be detected. Labeling of the major band was completely abolished by incubation in the presence of unlabeled hGH.
Analysis of 12sI-hGH cross-linked to the solubilized lactogen receptors in the absence of reducing agents (Fig. 3B) revealed that l2'1-hGH was covalently attached to two predominant species to yield complexes with M , = 60,000 and 100,000. A concentration-dependent increase in the radioactivity incor- porated into both bands was observed using DSS. Only one minor band at M , = 37,000-41,000 could be detected even after long exposures of the films to the dried gels. Excess unlabeled hGH abolished the labeling of the M , = 60,000 and the M , = 100,000 bands. By contrast, the intensity of the M , = 37,000-41,000 band remained unchanged. Similar results were obtained using the cleavable cross-linking reagent di- thiobis(succinimidy1 propionate) (not shown). Electrophore- sis of the covalently labeled membranes or solubilized prepa- rations under nonreducing conditions in gels containing 5% acrylamide did not result in the appearance of any labeled species other than those indicated above (not shown).
Even though the same bands appeared in the autoradi- ograms obtained under either reducing or nonreducing con- ditions, significant differences in the relative intensities of the bands were noted. When solubilized lZ5I-hGH-receptor complexes were reacted with 0.5 mM DSS and analyzed under reducing conditions (Fig. 1B, fifth lune), densitometric scan- ning of the autoradiograms showed that the M , = 60,000 band was 50% more intense than the M, = 100,000 band. By contrast, when analysis was performed in the absence of reductant (Fig. 3B, third lane), the M , = 60,000 band was 40% less intense than the M , = 100,000 band. These observations suggested that a fraction of the M, = 60,000 protein could be part of the M , = 100,000 protein, being released after cleavage of disulfide bonds with mercaptoethanol.
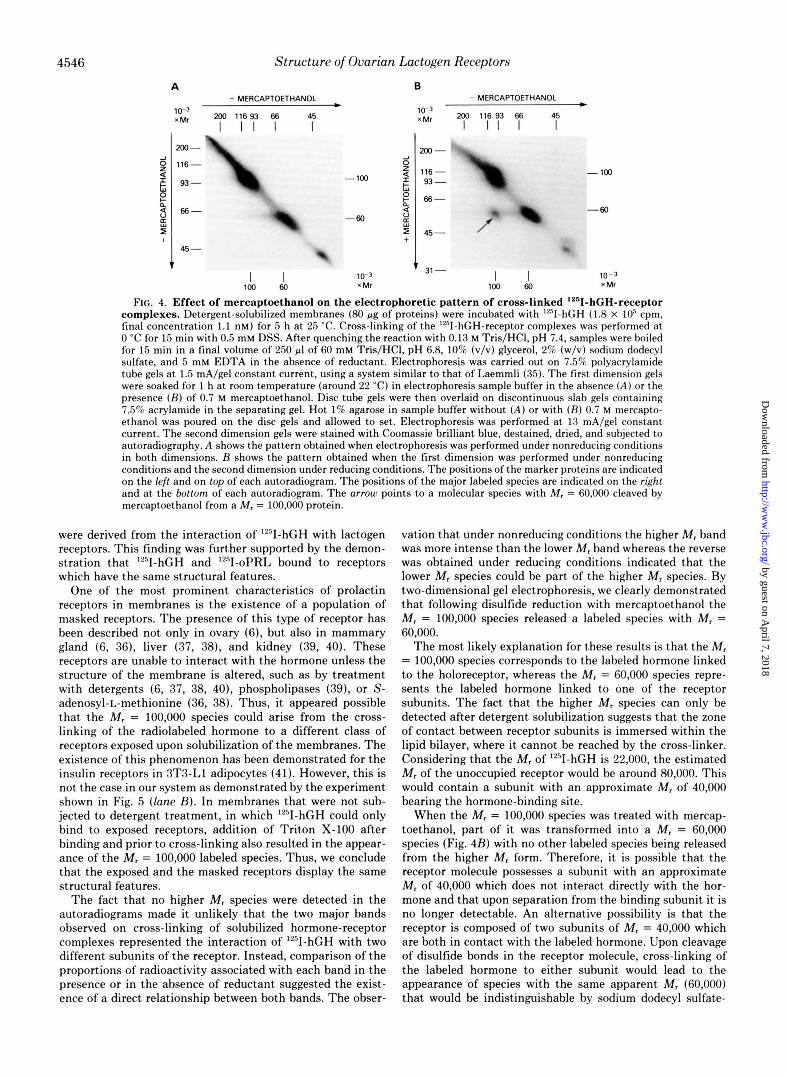
This proposal was further examined by subjecting cross- linked hormone-receptor complexes to two-dimensional elec- trophoresis. Fig. 4A shows the pattern obtained when electro- phoresis in both dimensions was performed under nonreduc- ing conditions. In this case, all the labeled species are located along a diagonal. The two major species with M , = 60,000 and 100,000 as well as the minor M, = 40,000 band depicted in the third lune of Fig. 3B were clearly seen. When electropho- resis in the second dimension was performed following reduc- tion of the tube gel with mercaptoethanol, an additional radioactive spot with M, = 60,000 could be seen deriving from the M , = 100,000 species (Fig. 4B). This demonstrated that at least part of the M , = 60,000 protein is contained within the M, = 100,000 species from which it can be released upon treatment with reducing agents. The fact that not all of the M , = 100,000 labeled protein could be cleaved to a lower molecular weight species could be attributed to cross-linking between the receptor subunits. Thus, the M , = 60,000 species can only be derived from the M , = 100,000 molecules held together by disulfide bonds but not by the suberate linker.
Previous studies (6) have demonstrated that treatment of ovarian membranes with 1% (w/v) Triton X-100 caused a significant increase in the number of lactogen receptors. This observation, together with similar findings in other organs
(36-40), led to the concept that besides the "exposed recep- tors which can interact directly with the labeled hormone in membranes, there is a population of "masked receptors which can only bind hormone after treatment with detergent. To assess whether the different patterns of labeling obtained using membranes or a solubilized extract were due to different structural characteristics of the exposed and masked recep- tors, we analyzed the effect of cross-linking on solubilized 1251- hGH-receptor complexes preformed in membranes (e.g. ex- posed receptors). Analysis of these preparations showed the presence of both the M , = 60,000 and the M , = 100,000 bands in the autoradiograms (Fig. 5, lune B), thus demonstrating that the exposed and masked receptors have similar struc- tures. It is also clear that in order to detect the Mr = 100,000 band, the receptors have to be solubilized either before or after binding of the hormone and prior to cross-linking. Similar results were observed using a purified membrane preparation as source of receptors (Fig. 5, lanes C and D).
Addition of Triton X-100 to prelabeled membranes after cross-linking and prior to electrophoretic analysis had no effect on the cross-linking pattern (Fig. 5, lune E ) . No major changes were observed when the membranes were prepared and incubated with 1251-hGH in the presence of a mixture of protease inhibitors including trypsin inhibitor, phenylmeth- ylsulfonyl fluoride, leupeptin, N"-p-tosyl-L-lysine chloro- methyl ketone, and aprotinin (Fig. 5, lunes F and G). The same minor radiolabeled bands described above were observed in these autoradiograms, suggesting that they are not products of proteolysis derived from the major species. The only change observed at this level was an increase in the labeling of a band with M , = 30,000.
The addition of 20 mM N-ethylmaleimide to the homoge- nization medium for the purpose of alkylating "SH groups resulted in an increased detection of the higher M , species in membranes when samples were analyzed under nonreducing conditions (Fig. 5, lune H).
To further demonstrate that the receptors to which 1251- hGH was cross-linked were lactogenic in nature, similar stud- ies were carried out using 1251-oPRL as tracer. Membranes and solubilized extract previously incubated with 1251-oPRL were subjected to cross-linking with 0.5 mM DSS. A major band at M , = 60,000 was observed in membranes, and bands at M, = 60,000 and 100,000 were observed in solubilized preparations (Fig. 6). Unlabeled oPRL caused a significant decrease in the intensity of these bands (Fig. 6). Unlabeled hGH was also able to prevent the appearance of the major bands in membranes as well as in solubilized preparations (not shown). The similarity of the patterns of cross-linking obtained using either lZ5I-hGH or 1251-oPRL suggests that both hormones bind to the same receptor in the ovary and that this receptor has specificity for lactogenic hormones.
DISCUSSION
These studies have demonstrated that 1251-hGH can be covalently linked to the lactogen receptors in luteinized rat ovaries with high efficiency using the bifunctional N-hy- droxysuccinimide ester DSS. Analysis of the cross-linked membrane-bound complexes by sodium dodecyl sulfate-poly- acrylamide gel electrophoresis under reducing conditions fol- lowed by autoradiography revealed the presence of a major band with M , = 60,000. The same labeled species was detected when solubilized '251-hGH-receptor complexes were cross- linked. Interestingly, a second major band at M , = 100,000 was also clearly detectable in this case. Incubation with 3,800- fold excess lactogenic hormones prevented the appearance of these bands in the autoradiograms, demonstrating that they
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
4546
A
Structure of Ovarian Lactogen Receptors
B - MERCAPTOETHANOL
lo-? 200 11693 66 45 x Mr I I I I I
,I\ 93 - -
66-
45 -
I I 100 60
h
. 6 0 9 Y LC
4
10-3 x Mr
- MERCAPTOETHANOL
LLt 200 116 93 66 45 I I I I I
200-
116 - 93 - 66-
45 -
- loo
-60
31 - I I 10-3 100 60 x Mr
FIG. 4. Effect of mercaptoethanol on the electrophoretic pattern of cross-linked '*"I-hGH-receptor complexes. Detergent-solubilized membranes (80 pg of proteins) were incubated with 9 - h G H (1.8 X IO5 cpm, final concentration 1.1 nM) for 5 h a t 25 'C. Cross-linking of the "'I-hGH-receptor complexes was performed a t 0 "C for 15 min with 0.5 mM DSS. After quenching the reaction with 0.13 M Tris/HCl, pH 7.4, samples were boiled for 15 min in a final volume of 250 pl of 60 mM Tris/HCI, pH 6.8, 10% (v/v) glycerol, 2% (w/v) sodium dodecyl sulfate, and 5 mM EDTA in the absence of reductant. Electrophoresis was carried out on 7.5% polyacrylamide tube gels a t 1.5 mA/gel constant current, using a system similar to that of Laemmli (35). The first dimension gels were soaked for 1 h a t room temperature (around 22 "C) in electrophoresis sample buffer in the absence ( A ) or the presence ( R ) of 0.7 M mercaptoethanol. Disc tube gels were then overlaid on discontinuous slab gels containing 7.5% acrylamide in the separating gel. Hot 1% agarose in sample buffer without ( A ) or with ( R ) 0.7 M mercapto- ethanol was poured on the disc gels and allowed to set. Electrophoresis was performed a t 13 mA/gel constant current. The second dimension gels were stained with Coomassie brilliant blue, destained, dried, and subjected to autoradiography. A shows the pattern obtained when electrophoresis was performed under nonreducing conditions in both dimensions. H shows the pattern obtained when the first dimension was performed under nonreducing conditions and the second dimension under reducing conditions. The positions of the marker proteins are indicated on the left and on top of each autoradiogram. The positions of the major labeled species are indicated on the right and at the bottom of each autoradiogram. The arrow points to a molecular species with M, = 60,000 cleaved by mercaptoethanol from a M, = 100.000 protein.
were derived from the interaction of '*'I-hGH with lactogen receptors. This finding was further supported by the demon- stration that "'I-hGH and '2sI-oPRL bound to receptors which have the same structural features.
One of the most prominent characteristics of prolactin receptors in membranes is the existence of a population of masked receptors. The presence of this type of receptor has been described not only in ovary (6), but also in mammary gland (6, 36), liver (37, 38), and kidney (39, 40). These receptors are unable to interact with the hormone unless the structure of the membrane is altered, such as by treatment with detergents (6, 37, 38, 401, phospholipases (39), or S- adenosyl-L-methionine (36, 38). Thus, it appeared possible that the M , = 100,000 species could arise from the cross- linking of the radiolabeled hormone to a different class of receptors exposed upon solubilization of the membranes. The existence of this phenomenon has been demonstrated for the insulin receptors in 3T3-Ll adipocytes (41). However, this is not the case in our system as demonstrated by the experiment shown in Fig. 5 (lane 23). In membranes that were not sub- jected to detergent treatment, in which '*'I-hGH could only bind to exposed receptors, addition of Triton X-100 after binding and prior to cross-linking also resulted in the appear- ance of the M , = 100,000 labeled species. Thus, we conclude that the exposed and the masked receptors display the same structural features.
The fact that no higher M , species were detected in the autoradiograms made it unlikely that the two major bands observed on cross-linking of solubilized hormone-receptor complexes represented the interaction of I2'I-hGH with two different subunits of the receptor. Instead, comparison of the proportions of radioactivity associated with each band in the presence or in the absence of reductant suggested the exist- ence of a direct relationship between both bands. The obser-
vation that under nonreducing conditions the higher Mr band was more intense than the lower M , band whereas the reverse was obtained under reducing conditions indicated that the lower M , species could be part of the higher M , species. By two-dimensional gel electrophoresis, we clearly demonstrated that following disulfide reduction with mercaptoethanol the M , = 100,000 species released a labeled species with M , = 60,000.
The most likely explanation for these results is that the M , = 100,000 species corresponds to the labeled hormone linked to the holoreceptor, whereas the Mr = 60,000 species repre- sents the labeled hormone linked to one of the receptor subunits. The fact that the higher M , species can only be detected after detergent solubilization suggests that the zone of contact between receptor subunits is immersed within the lipid bilayer, where it cannot be reached by the cross-linker. Considering that the M , of '""IhGH is 22,000, the estimated M , of the unoccupied receptor would be around 80,000. This would contain a subunit with an approximate M , of 40,000 bearing the hormone-binding site.
When the M , = 100,000 species was treated with mercap- toethanol, part of it was transformed into a M , = 60,000 species (Fig. 4B) with no other labeled species being released from the higher M , form. Therefore, it is possible that the receptor molecule possesses a subunit with an approximate M , of 40,000 which does not interact directly with the hor- mone and that upon separation from the binding subunit it is no longer detectable. An alternative possibility is that the receptor is composed of two subunits of M , = 40,000 which are both in contact with the labeled hormone. Upon cleavage of disulfide bonds in the receptor molecule, cross-linking of the labeled hormone to either subunit would lead to the appearance of species with the same apparent M, (60,000) that would be indistinguishable by sodium dodecyl sulfate-
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10 x Mr
200 -
116- 93 - 66-
45 -
31 -
Structure of Ovarian Lactogen Receptors
A B C D E
10 3 x Mr
200- "
116- 93-
66-
4547
10 x Mr
200 - w - 116-
93- *
66 -
45 - 45-
31- F G H I
FIG. 5. Autoradiograms showing control experiments. Lane A, crude ovarian memhranes (480 pg of proteins) were incubated with "'I-hCH (1.0 X 10" cpm, final concentration 1.4 nM) for 5 h at 25 "C. After incubation, membranes were washed, resuspended in 0.4 ml of Dulhecco's PBS, and treated with 0.5 mM DSS for 15 min at 0 "C. Lane H. same as lane A, except that after washing of unbound '?'I-hGH, the memhranes were resuspended in Dulhecco's I'HS containing 1% (w/v) Triton X-100 prior to cross-linking. Lune C, purified ovarian plasma membranes (58 pg of proteins) were incubated with lZ'l-hCH (1.0 X lofi cpm, final concentration 1.4 nM) for 5 h at 25 'C. Labeled membranes were washed, resuspended in 0.4 ml of Dulbecco's PRS, and cross-linked with DSS as descrihed for lane A. Lane I) . same as lane C, except that labeled membranes were resuspended in Dulhecco's PHS containing 1% (w/v) Triton X-100 before cross-linking. Imze E , same as lane A, except that after cross- linking. membranes were resuspended in 1% (w/v) Triton X-lOO/Dulhecco's PBS. Lane b', crude memhranes were prepared in the presence of a mixture of proteolysis inhibitors containing 0.1 mg/ml of trypsin inhibitor, 1 mM leupeptin, 1 mM phenylmethylsulfonyl fluoride, 2 mM N"-p-tosyl-l,-lysine chloromethyl ketone, and 10 trypsin inhibitor units/ml of aprotinin. An aliquot of the resulting suspension (480 pg of proteins) was incubated with "'I- hGH (1.0 X IO6 cpm, final concentration 1.4 nM). Protease inhihitors were present in the incuhation medium at concentrations of one fifth of those used for preparation of the membranes. After incubation, memhranes were washed and resuspended in 0.4 ml of Dulhecco's PBS and reacted with 0.5 mM DSS under standard conditions. Lane G, same as lane F, except that Dulhecco's PBS employed to resuspend the labeled membranes contained 1% (w/v) Triton X-100. Lane H. crude membranes were prepared in the presence of 20 mM N-ethylmaleimide. An aliquot of the resulting suspension (480 pg of proteins) was incubated with "'I-hCH (1.0 X 10" cpm, final concentration 1.4 nM). The incubation medium contained 5 mM N-ethylmaleimide. The labeled memhranes were then resuspended in 0.4 ml of Dulbecco's PHS and cross-linked with 0.5 mM DSS. Lane I, same as lane H , except that membranes were resuspended in Dulhecco's PBS containing 1% (w/v) Triton X-100 prior to cross-linking. Samples in lanes A-C were electrophoresed under reducing conditions, whereas samples in lanes H and I were electrophoresed under nonreducing conditions. The positions of the molecular weight markers are indicated on the left of each autoradiogram.
polyacrylamide gel electrophoresis. Similar to recent findings on the insulin receptors (18, 21,
42) and insulin-like growth factor receptors (23, 24, 26), disulfide bonds are involved, a t least in part, in the interaction between the subunits of the ovarian lactogen receptors. How- ever, the fact that a fraction of the M, = 60,000 species is detected to a certain degree under nonreducing conditions indicates that a proportion of the subunits are either free or associated by noncovalent interactions. Therefore, the native receptors could consist of two populations: one in which the subunits are linked by disulfide bonds and another in which the interaction between subunits is of noncovalent nature. Reduced forms of the insulin receptor have also been found in native plasma membranes from rat liver even when elec- trophoresis was performed under nonreducing conditions (21). Provided that some of the receptor subunits are held together by disulfide bonds, it is not clear why the higher M, form was not more evident when cross-linked hormone-receptor com- plexes in membranes were analyzed under nonreducing con- ditions (Fig. 3A). A likely explanation for this fact is the occurrence of cleavage of disulfide bonds during incubation of the membranes with the labeled hormone. This cleavage could take place by thiol-disulfide exchange with non-protein thiols that contaminate membrane preparations and could be either spontaneous or catalyzed by an enzyme. Enzymes that are able to catalyze this type of reaction have been found in microsomal membranes from various tissues (43), and at least
one of them has been demonstrated to possess a "SH group essential for its catalytic activity (44). N-Ethylmaleimide appears to inhibit the cleavage of the disulfide-linked form of the receptor since in its presence the higher M, species be- comes clearly evident in membranes (Fig. 5 , lane H). Inhibi- tion of the cleavage by N-ethylmaleimide could be due to depletion of non-protein thiols or to inactivation of the en- zymes necessary for reduction of disulfide bonds.
In addition to the major bands, several other minor bands were observed in the autoradiograms. However, their intensity was much lower and more variable and their appearance tended to be less dependent on the presence or absence of excess unlabeled hormone in the incubation medium. These bands do not seem to be products of proteolysis of the major species, since most of them were still present when prepara- tion of the membranes and hormone binding were carried out in the presence of several protease inhibitors. Competitive binding studies (6) have shown that besides the high affinity receptors there was a second population of sites in the ovary with about 100 times less affinity for "'I-hGH. Thus, it is likely that at least some of the minor bands derive from nonspecific interaction of the labeled hormone with proteins other than the lactogen receptors. At any rate, the possibility that some of these minor bands are related in some way to the receptor molecule cannot be completely ruled out. The appearance of other bands in addition to those derived from the hormone-receptor complexes is an ordinary finding in
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4548 Structure of Ovarian Lactogen Receptors
10-3 10-3 10-3 10-3 x Mr x Mr x Mr x Mr
200 -
116 -
93 -
200 -
116- 93 - ,100
66- 66- ' 6 0
-60
45 - 45 -
UNLABELED - - oPRL
+ UNLABELED - - oPRL
+ MEMBRANES SOLUBILIZED
EXTRACT FIG. 6. Cross-linking of '261-oPRL to the ovarian lactogen
receptors. Crude ovarian membranes (450 pg of proteins) were incubated with 12'I-oPRL (1.5 X lo6 cpm) for 5 h a t 25 "C in the absence (-) or in the presence (+) of 5 pg of unlabeled oPRL. After washing out the unbound labeled hormone, membranes were resus- pended in 0.4 ml of Dulbecco's PBS and treated with 0.5 mM DSS for 15 min a t 0 "C. Solubilized extract (100 pg of proteins) was incubated with "'I-oPRL (2 X 1O'cpm) for 5 h a t 25 "C in the absence (-) or in the presence (+) of 5 pgof unlabeled oPRL. After incubation, the mixture was cross-linked with 0.5 mM DSS under conditions similar to those described for membranes. Samples were electropho- resed under reducing conditions. The positions of the molecular weight markers are denoted on the left of each autoradiogram. The positions of the major labeled species are indicated on the right of each autoradiogram.
cross-linking and photoaffinity labeling techniques (19, 21,
Recently, it has been reported that '2"I-hGH can be cova- lently attached to lactogen receptors in rat liver and rabbit mammary gland microsomal fractions using the bifunctional reagent ethylene glycol bis(succinimidy1 succinate) (48). Anal- ysis of the cross-linked samples showed the presence of labeled bands of M, = 66,000 and 61,000 respectively. l""IoPRL has also been cross-linked to the liver lactogen receptors using the photoactivable reagent N-hydroxysuccinimidyl 4-azido- benzoate (49). A M, = 60,000 band was detected in this case. Lactogen receptors from rabbit mammary gland were pho- toaffinity labeled with l""IhGH derivatized with ethyl-4-azi- dophenyl-1,2-dithiobutyrimidate to yield a complex with M, = 57,000 (50). These results are similar to what we have observed upon cross-linking of "'I-hGH-receptor complexes in ovarian membranes. In addition, prolactin receptors have been purified from mouse liver (51), rabbit liver (50), and rabbit mammary gland (50) by affinity chromatography. The major molecular species detected in the purified preparations had M, of 37,000, 35,000, and 35,000, respectively. These values are in close agreement with the molecular weight of the species detected by cross-linking techniques, once the contribution of the labeled hormone is subtracted. No evi- dence was presented in the studies cited above for the exist- ence of a receptor species with higher molecular weight. In the case of the cross-linking experiments, a complex with M, - 100,000 could have escaped detection because the covalent linking was performed in membranes and not in solubilized preparations. Relevant to this point is the work of Jaffe (52) on the physical properties of detergent-solubilized "'I-hCH- lactogen receptor complexes from rat liver. By performing hydrodynamic measurements, this author demonstrated com- plexes of M, = 148,000, of which 48,200 corresponded to
25, 27, 28, 45-47).
Triton X-100 bound and 99,800 to the protein "core." This is consistent with our observation of a hormone-receptor com- plex at M, = 100,000.
REFERENCES
1. Saito, T., and Saxena, B. B. (1975) Acta Endocrinol. 30,126-137 2. Richards, J. S., and Williams, J. J. (1976) Endocrinology 99,
3. Cheng, K. W. (1976) J. Reprod. Fertil. 48, 129-13s 4. Solano, A. R., Garcia-Vela, A., Catt, K. J., and Dufau, M. L.
5. Davies, T. F., Katikineni, M., Chan, V., Harwood, J. P., Dufau, M. L., and Catt, K. J. (1980) Nature (Land.) 283, 863-865
6. Koppelman, M. C. S., and Dufau, M. L. (1982) Endocrinology
7. Poindexter, A. N., Buttram, V. C., Besch, P. K., and Smith, R.
8. Rolland, R., and Hammond, J. M. (1975) Endocr. Res. Commun.
9. Rolland, R., Gunsalus, G . L., and Hammond, J. M. (1976) En-
10. Ben-David, M., and Schenker, J. G . (1982) Fertil. Steril. 38,182-
11. Midgley, A. R. (1973) Ado. Exp. Med. Biol. 36,365-378 12. Dunaif, A. E., Zimmerman, E. A., Friesen, H. G., and Frantz, A.
13. Wang, C., Hsueh, A. J. W., and Erickson, G. F. (1979) J. Biol.
14. Knecht, M., Amsterdam, A., and Catt, K. (1981) J . Biol. Chem.
15. Navickis, R. J., Jones, P. B. C., and Hsueh, A. J. W. (1982) Mol. Cell. Endocr. 27,77-88
16. Harwood, J. P., Clayton, R. N., and Catt, K. J. (1980) in Endo- crinology 1980 (Cumming, I. A., Funder, J. w., and Mendelsohn, F. A. O., eds) pp. 436-439, Australian Academy of Sciences, Canberra
17. Pilch, P. F., and Czech, M. P. (1979) J. Biol. Chem. 254, 3375- 3381
18. Pilch, P. F., and Czech, M. P. (1980) J. Biol. Chem. 255, 1722- 1731
19. Massague, J., Pilch, P. F., and Czech, M. P. (1980) Proc. Natl. Acad. Sci. U. S. A. 77, 7137-7141
20. Massague, J., Pilch, P. F., and Czech, M. P. (1981) J. Biol. Chem.
21. Massague, J., and Czech, M. P. (1980) Diabetes 29,945-947 22. Rebois, R. V., Omedeo-Sale, F., Brady, R. O., and Fishman, P.
H. (1981) Proc. Natl. Acad. Sci. U. S. A. 78,2086-2089 23. Kasuga, M., Van Obberghen, E., Nissley, S. P., and Rechler, M.
M. (1981) J. Biol. Chem. 256,5305-5308 24. Kasuga, M., Van Obberghen, E., Nissley, S. P., and Rechler, M.
M. (1982) Proc. Natl. Acad. Sci. U. S. A. 79, 1864-1868 25. Johnson, G. L., MacAndrew, V. I., and Pilch, P. F. (1981) Proc.
Natl. Acad. Sci. U. S. A. 78,875-878 26. Massague, J., and Czech, M. P. (1982) J. Biol. Chem. 257,5038-
5045 27. Paglin, S., and Jamieson, J. D. (1982) Proc. Natl. Acad. Sci.
28. Glenn, K., Bowen-Pope, D. F., and Ross, R. (1982) J. Biol. Chem.
29. Massague, J., Czech, M. P., Iwata, K., DeLarco, J. E., and Todaro, G. J. (1982) Proc. Natl. Acad. Sci. U. S. A. 79,6822-6826
30. Thorell, J. K., and Johansson, B. A. (1971) Bbchim. Biophys. Acta 25 1,363-369
31. Dufau, M. L., Podesta, E. J., and Catt, K. J. (1975) Proc. Natl. Acad. Sei. U. S. A. 72, 1272-1275
32. Gospodarowicz, D. (1973) J. Biol. Chem. 248,5050-5056 33. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
34. Heppel, L. A., and Hilmoe, R. J. (1951) J . Biol. Chem. 188,665-
35. Laemmli, U. K. (1970) Nature ( L o n d . ) 227, 680-685 36. Bhattacharya, A., and Vonderhaar, B. K. (1979) Proc. Natl. Acad.
37. Bonifacino, J. S., Sanchez, S. H., and Paladini, A. C. (1981)
38. Alhadi, T., and Vonderhaar, B. K. (1982) Endocrinology 110,
1571-1581
(1980) FEBS Lett. 122, 184-188
11 1, 1350-1357
G . (1979) Fertil. Steril. 31, 273-277
2,281-298
docrinology 98, 1083-1091
186
G. (1982) Endocrinology 110, 1465-1471
Chem. 254, 11330-11336
256,10628-10633
256,3182-3190
U. S. A. 79,3739-3743
257,5172-5176
(1951) J. Biol. Chem. 193, 265-275
676
Sci. U. S. A. 76,4489-4492
Biochem. J. 194.385-394
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

39.
40.
41.
42.
43. 44.
45.
Structure of Ovarian Lactogen Receptors 4549
254-259 Roguin, L. P., Sanchez, S. H., Bonifacino, J . S., and Paladini, A.
Roguin, L. P., Bonifacino, J. S., and Paladini, A. C. (1982)
Deutsch, P. J., Wan, C. F., Rosen, 0. M., and Rubin, C. S. (1983)
Jacobs, S., Hazum, E., and Cuatrecasas, P. (1980) J. Biol. Chem.
Freedman, R. E. (1979) FEBS Lett. 242,201-210 Fuchs, S., De Lorenzo, F., and Anfinsen, C. B. (1967) J. Biol.
Ji, I., and Ji, T. H. (1980) Proc. Nutl. Acad. Sei. U. S. A. 77 ,
C . (1981) Biochem. J . 2 0 0 , 257-264
Biochim. Biophys. Acta 715,222-229
Proc. Natl. Acud. Sci. U. S. A. 80, 133-136
255,6937-6940
Chem. 242,398-402
46.
47.
48.
49.
50.
51.
52.
7167-7170 Moseley, J. M., Findlay, D. M., Martin, T. J., and Gorman, J. J.
Yip, C. C., Yeung, C . W. T., and Moule, M. L. (1980) Biochemistry
Hughes, J. P., Simpson, J . S. A,, and Friesen, H. G. (1983)
Borst. D. W.. and Savare. M. (1982) Biochem. BioDhvs. Res.
(1982) J. Biol. Chem. 257 , 5846-5851
19,70-76
Endocrinology 112, 1980-1985
Cokmun. 105,194-io1 ’
. . _ - Haeuptle. M.-T., Aubert, M. L., Djiane, J., and Kraehenbuhl,
J.-P. (1983) J. Biol. Chem. 2 5 8 , 305-314 Liscia, D. S., and Vonderhaar, B. K. (1982) Proc. Nutl. Acud. Sci.
Jaffe, R. C. (1982) Biochemistry 21, 2936-2939 U. S. A. 79,5930-5934
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

J S Bonifacino and M L Dufaucross-linking reagents.
Structure of the ovarian lactogen receptors. Analysis with bifunctional
1984, 259:4542-4549.J. Biol. Chem.
http://www.jbc.org/content/259/7/4542Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/259/7/4542.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from