structures of receptor complexes formed by hemagglutinins ... · structures of receptor complexes...
TRANSCRIPT

Structures of receptor complexes formedby hemagglutinins from the Asian Influenzapandemic of 1957Junfeng Liua, David J. Stevensa, Lesley F. Hairea, Philip A. Walkera, Peter J. Coombsa, Rupert J. Russellb,Steven J. Gamblina,1, and John J. Skehela,1
aMRC National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, United Kingdom; and bInterdisciplinary Centre for Human andAvian Influenza Research, School of Biology, University of St. Andrews, Fife KY16 9ST, United Kingdom
Edited by Stephen C. Harrison, Children’s Hospital, Boston, Boston, MA, and approved August 24, 2009 (received for review June 18, 2009)
The viruses that caused the three influenza pandemics of thetwentieth century in 1918, 1957, and 1968 had distinct hemagglu-tinin receptor binding glycoproteins that had evolved the capacityto recognize human cell receptors. We have determined the struc-ture of the H2 hemagglutinin from the second pandemic, the‘‘Asian Influenza’’ of 1957. We compare it with the 1918 ‘‘SpanishInfluenza’’ hemagglutinin, H1, and the 1968 ‘‘Hong Kong Influ-enza’’ hemagglutinin, H3, and show that despite its close overallstructural similarity to H1, and its more distant relationship to H3,the H2 receptor binding site is closely related to that of H3hemagglutinin. By analyzing hemagglutinins of potential H2 avianprecursors of the pandemic virus, we show that the human recep-tor can be bound by avian hemagglutinins that lack the human–specific mutations of H2 and H3 pandemic viruses, Gln-226Leu, andGly-228Ser. We show how Gln-226 in the avian H2 receptor bindingsite, together with Asn-186, form hydrogen bond networksthrough bound water molecules to mediate binding to humanreceptor. We show that the human receptor adopts a very similarconformation in both human and avian hemagglutinin-receptorcomplexes. We also show that Leu-226 in the receptor binding siteof human virus hemagglutinins creates a hydrophobic environ-ment near the Sia-1-Gal-2 glycosidic linkage that favors binding ofthe human receptor and is unfavorable for avian receptor binding.We consider the significance for the development of pandemics, ofthe existence of avian viruses that can bind to both avian andhuman receptors.
The second influenza pandemic of the 20th century, ‘‘AsianInfluenza,’’ began in 1957 (1–3) and ended with the appear-
ance of ‘‘Hong Kong Influenza,’’ in 1968. The viruses responsiblefor the 1957 pandemic were formed by reassortment betweenhuman and avian viruses (4, 5) and belong to the H2N2 subtype,signifying that, by comparison with the H1N1 viruses of the 1918‘‘Spanish Influenza,’’ the virus surface glycoproteins, hemagglu-tinin (‘H’) and neuraminidase (‘N�), were antigenically different(6). Both of these glycoproteins, and the polymerase componentPB1, were derived from avian viruses (4, 5). Two groups of H2avian hemagglutinins (HA) that have been characterized bygenetic and antigenic analyses, are components of viruses lo-cated either in Europe and Asia or in North America (7, 8). TheHA of the 1957 pandemic is a member of the Eurasian group.
Hemagglutinin is the receptor binding glycoprotein of influ-enza viruses (9). The receptors are sialic acids (10) that areterminal residues of carbohydrate side-chains of cellular glyco-proteins and glycolipids, and receptor recognition has beenshown to vary in specificity depending on the species infected(11, 12). The focus of this specificity is the nature of theglycosidic linkage between sialic acid and the penultimate sugarof the side-chains (11, 13). HAs of avian viruses prefer sialic acidin �2,3-linkage to galactose, the avian receptor, which predom-inates in the avian enteric tract (14, 15), while those of humanviruses prefer sialic acid in �2,6-linkage, the human receptor,which is predominant in the upper respiratory tract (16–18). The
avian origin of pandemic influenza virus HAs, therefore, impliesa change in receptor binding specificity. Genetic analyses haveindicated that for ‘‘Asian’’ and ‘‘Hong Kong’’ pandemic viruses,the same amino acid substitutions in the receptor binding sitesof HA correlate with their acquired preference for the humanreceptor; Gln-226Leu and Gly-228Ser (12). The three sites forsialic acid binding are at the membrane-distal tips of the identicalmonomers that form the HA trimer (9) (Fig. 1). Each sitecomprises a pocket of conserved amino acids that is edged by themembrane-distal 190-helix at the top of the site, and the 130- and220-loops located at the front edge and the left side of the site,respectively (9, 19, 20). Thus the 226/228 pair of substitutions arelocated at the bottom left of the receptor binding site.
To examine the structural significance of these changes wehave determined by X-ray crystallography the structures of theHAs of human and avian H2 viruses complexed with sialopen-tasaccharide analogues of human and avian receptors. The HAschosen for study were from the two prototype human viruses of1957, A/Singapore/1/57 (Singapore) (1) and A/Japan/305/57(Japan) (2) and from one Eurasian A/ck/Potsdam/84 and twoNorth American avian viruses, A/dk/Ontario/77 and A/ck/NewYork/91 (7, 8). Our results for the HA of the 1957 pandemic arecompared with others derived from studies of HAs of the 1918and 1968 pandemics, reported here and before, (21–23). Tofacilitate this comparison we have adopted an ‘H3’ numberingscheme for the H2 HAs.
ResultsStructure Determination. Diffraction datasets were collected fromcrystals of two human and three avian H2 HAs, both fromnative, as well as human or avian receptor sialopentasaccharideanalogue-soaked, crystals (summarized in Table 1). The struc-tures were solved by molecular replacement and subsequentlyrefined by automated methods interspersed with manual re-building using COOT (24). Relevant crystallographic statisticsare given in the SI.
As seen for other HA subtypes, (21, 25) there is a closecorrespondence in the structures of H2 HA from human andavian viruses reflecting the high level of sequence identitybetween them (about 90%) (Fig. 1C). As expected from phylo-genetic clustering (26), H2 HAs are most closely related in
Author contributions: J.L., D.J.S., L.F.H., P.A.W., P.J.C., R.J.R., S.J.G., and J.J.S. designed andperformed research and wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The atomic coordinates have been deposited in the Protein Data Bank,www.pdb.org (PDB ID codes 2WR0–2WR5, 2WR7, 2WRB–2WRH).
1To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0906849106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0906849106 PNAS � October 6, 2009 � vol. 106 � no. 40 � 17175–17180
MIC
ROBI
OLO
GY

overall structure to H1 with root mean square deviation(r.m.s.d.) in C� positions of about 1.3Å between monomers, inboth cases. Thus the H1 and H2 HAs associated with the first twopandemic viruses in the last century are closely related but quitedistinct from the third, the H3 HA of the 1968 pandemic virus.The r.m.s.d. (on all C�s) between avian H2 and human H3 HAis 2.4Å (22), reflecting the lower sequence identity betweenthese proteins (about 40%). This comparison is of specialinterest because, as mentioned above, H2 and H3 HAs share thesame avian to human host adaptation mutations, Gln-226Leuand Gly-228Ser, but H1 does not (27).
Avian H2 Complexes with Avian and Human Receptors. Avian HA withavian receptor. The complex between avian H2 HA and an avianreceptor analogue reveals that the ligand binds in a similarmanner to that seen in nearly all other avian HAs, with thesialopentasaccharide adopting a trans conformation about theglycosidic bond between Sia-1 and Gal-2 (Fig. 2A) (21, 22, 25,28). Consequently, the trajectory of the second and third sugarrings, relative to the sialic acid, is also similar to other avianHA/avian receptor complexes (Fig. 2 A) (21, 25, 28). The keyinteractions involved in avian receptor binding are between theside-chain of the ‘avian-signature’ residue Gln-226 (28, 29) andthe glycosidic oxygen between Sia-1 and Gal-2, and the 4�OH ofGal-2.Avian haemagglutinin with human receptor. The structure of avian H2in complex with human receptor (Fig. 2B), also shows strongelectron density extending to at least the third sugar, GlcNAc-3,of the sialopentasaccharide, which has a similar conformation tothat observed for this ligand bound to human H2 HA (Fig. 2C).The Sia-1-Gal-2 glycosidic bond adopts a cis conformation andthe pentasaccharide exits the binding site in approximately theopposite direction to the avian receptor (Fig. 2 A). There areextensive hydrogen bond interactions between avian HA and
Gal-2 of the human receptor; particularly noteworthy are theroles of two water molecules in mediating these interactions.Wat-1 links the 3� OH of Gal-2 with the side-chain of Lys-222and the main-chain carbonyl at 225, while Wat-2 hydrogen bondsto Gln-226 and Asn-186, as well as to the 4�OH of Gal-2 and the9�OH of Sia-1 (Fig. 2B).
Human H2 Complexes with Human and Avian Receptors. Human HAwith human receptor. The complex between human H2 HA and thehuman receptor (Fig. 2C), shows that the ligand adopts a cisconformation at the glycosidic bond between Sia-1 and Gal-2and an overall orientation similar to that seen in complexes withhuman H3 haemagglutinin (Fig. 3A) (22). There is a hydrogenbond between the 3�OH of Gal-2 and the side-chain of Lys-222(Fig. 2C), but there is no electron density for water moleculesthat could mediate further hydrogen bond interactions betweenthe HA and ligand. The leucine residue at position 226 leads toa more hydrophobic environment than that present in avian HAsand likely accounts for the absence of water molecules in thisregion. Further, the shortest distance between the side-chain ofLeu-226 and C6 of the ligand is about 4.2 Å, which is too longto generate a significant hydrophobic interaction to favor thebinding of the �2,6-linked ligand. By contrast, in unligandedavian H2 HA Gln-226 coordinates a water molecule that is veryclose to the position occupied by Gal-2 in the human HA-humanreceptor complex. This suggests that human HAs benefit fromacquiring leucine at position 226 because human receptors canthen bind without the need to displace water.Human HA with avian receptor. In the complex between human H2HA and the avian receptor analogue, there is relatively poorelectron density for just the sialic acid of the receptor, indicativeof weak binding to this ligand (Fig. 2D). The most obviousexplanation for the poorer binding of avian receptor, that wasalso observed in binding assays (30), lies in the positioning of the
Fig. 1. Ribbons representation of different H2 HA monomers and receptor binding sites. (A) Superposition of the monomers of two human H2 HAs:A/Singapore/1/57 and A/Japan/305/57 colored green and yellow respectively. (B) Three avian H2 HAs: A/ck/New York/29878/91 colored gray, A/dk/Ontario/77colored in blue and A/ck/potsdam/4705/84 colored orange red. (C) Overlap of monomers of a human H2 HA colored in green and avian H2 HA colored in blue.The region highlighted by the gray ellipse at the top of the panel shows the receptor binding domain, an expanded version of which is shown in (D). Conservedresidues such as Tyr-98, Ser-136, Trp-153, and His-183 are shown in stick representation together with other residues important in receptor binding specificitysuch as Asn-186, Glu-190, and Leu-194, as well as the Gln/Leu-226, Gly/Ser-228 pair.
17176 � www.pnas.org�cgi�doi�10.1073�pnas.0906849106 Liu et al.
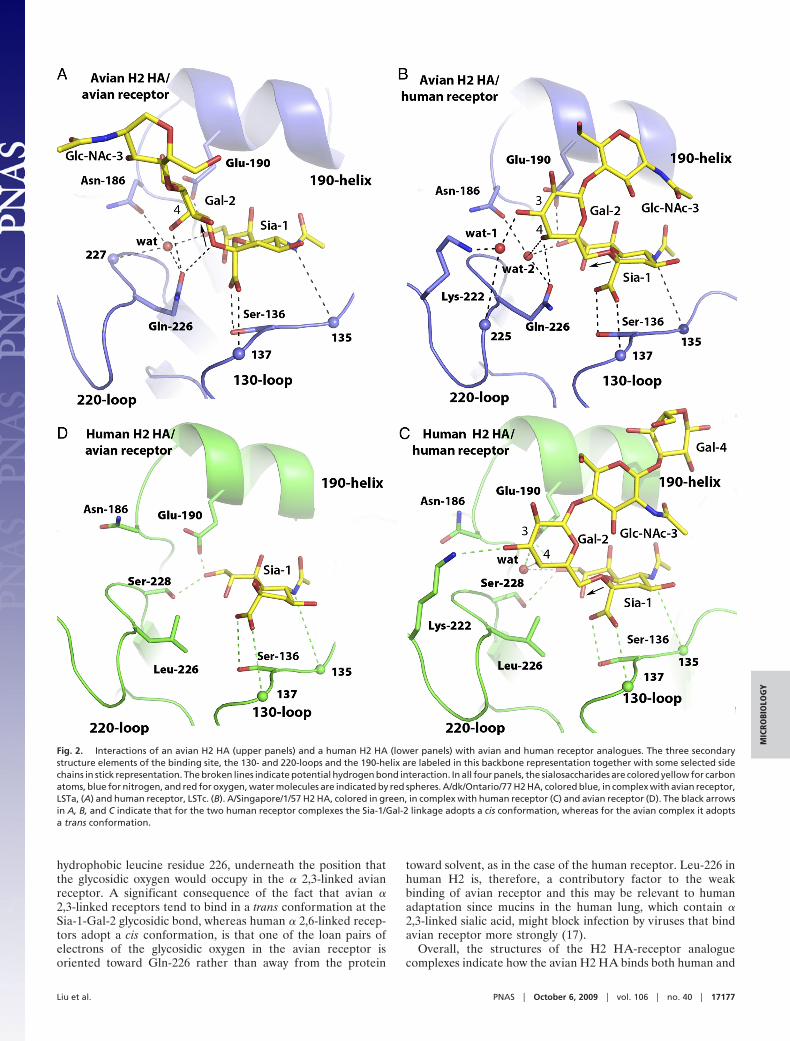
hydrophobic leucine residue 226, underneath the position thatthe glycosidic oxygen would occupy in the � 2,3-linked avianreceptor. A significant consequence of the fact that avian �2,3-linked receptors tend to bind in a trans conformation at theSia-1-Gal-2 glycosidic bond, whereas human � 2,6-linked recep-tors adopt a cis conformation, is that one of the loan pairs ofelectrons of the glycosidic oxygen in the avian receptor isoriented toward Gln-226 rather than away from the protein
toward solvent, as in the case of the human receptor. Leu-226 inhuman H2 is, therefore, a contributory factor to the weakbinding of avian receptor and this may be relevant to humanadaptation since mucins in the human lung, which contain �2,3-linked sialic acid, might block infection by viruses that bindavian receptor more strongly (17).
Overall, the structures of the H2 HA-receptor analoguecomplexes indicate how the avian H2 HA binds both human and
Fig. 2. Interactions of an avian H2 HA (upper panels) and a human H2 HA (lower panels) with avian and human receptor analogues. The three secondarystructure elements of the binding site, the 130- and 220-loops and the 190-helix are labeled in this backbone representation together with some selected sidechains in stick representation. The broken lines indicate potential hydrogen bond interaction. In all four panels, the sialosaccharides are colored yellow for carbonatoms, blue for nitrogen, and red for oxygen, water molecules are indicated by red spheres. A/dk/Ontario/77 H2 HA, colored blue, in complex with avian receptor,LSTa, (A) and human receptor, LSTc. (B). A/Singapore/1/57 H2 HA, colored in green, in complex with human receptor (C) and avian receptor (D). The black arrowsin A, B, and C indicate that for the two human receptor complexes the Sia-1/Gal-2 linkage adopts a cis conformation, whereas for the avian complex it adoptsa trans conformation.
Liu et al. PNAS � October 6, 2009 � vol. 106 � no. 40 � 17177
MIC
ROBI
OLO
GY
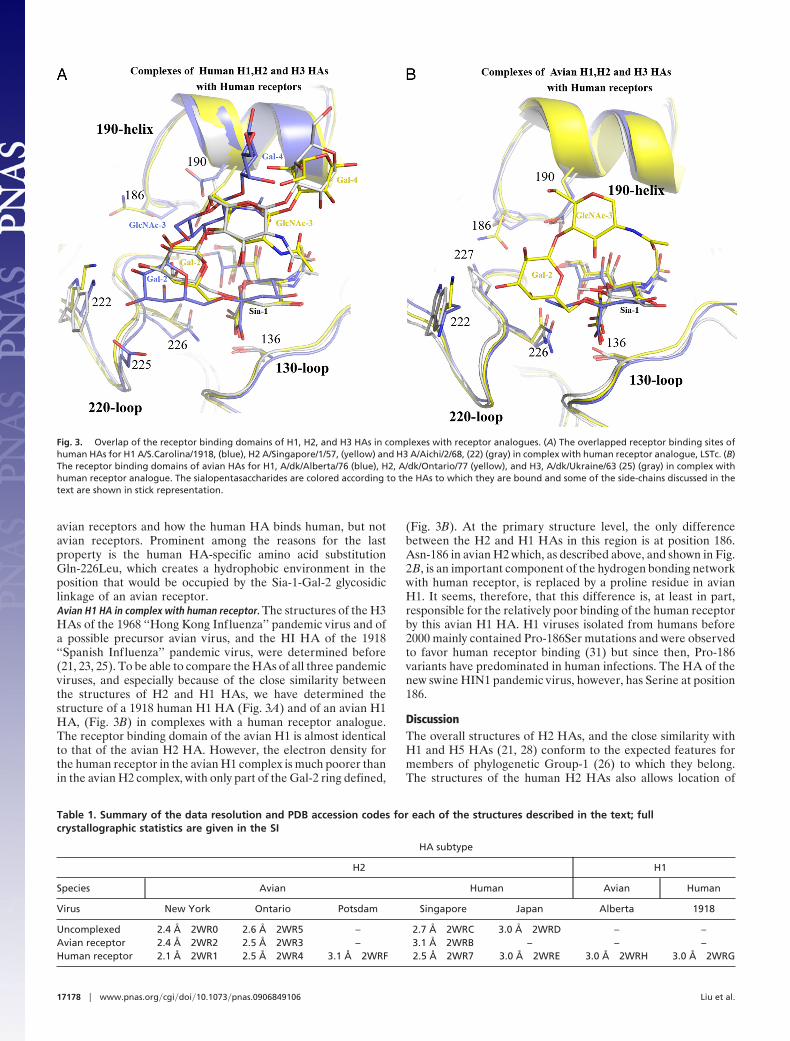
avian receptors and how the human HA binds human, but notavian receptors. Prominent among the reasons for the lastproperty is the human HA-specific amino acid substitutionGln-226Leu, which creates a hydrophobic environment in theposition that would be occupied by the Sia-1-Gal-2 glycosidiclinkage of an avian receptor.Avian H1 HA in complex with human receptor. The structures of the H3HAs of the 1968 ‘‘Hong Kong Influenza’’ pandemic virus and ofa possible precursor avian virus, and the HI HA of the 1918‘‘Spanish Influenza’’ pandemic virus, were determined before(21, 23, 25). To be able to compare the HAs of all three pandemicviruses, and especially because of the close similarity betweenthe structures of H2 and H1 HAs, we have determined thestructure of a 1918 human H1 HA (Fig. 3A) and of an avian H1HA, (Fig. 3B) in complexes with a human receptor analogue.The receptor binding domain of the avian H1 is almost identicalto that of the avian H2 HA. However, the electron density forthe human receptor in the avian H1 complex is much poorer thanin the avian H2 complex, with only part of the Gal-2 ring defined,
(Fig. 3B). At the primary structure level, the only differencebetween the H2 and H1 HAs in this region is at position 186.Asn-186 in avian H2 which, as described above, and shown in Fig.2B, is an important component of the hydrogen bonding networkwith human receptor, is replaced by a proline residue in avianH1. It seems, therefore, that this difference is, at least in part,responsible for the relatively poor binding of the human receptorby this avian H1 HA. H1 viruses isolated from humans before2000 mainly contained Pro-186Ser mutations and were observedto favor human receptor binding (31) but since then, Pro-186variants have predominated in human infections. The HA of thenew swine HIN1 pandemic virus, however, has Serine at position186.
DiscussionThe overall structures of H2 HAs, and the close similarity withH1 and H5 HAs (21, 28) conform to the expected features formembers of phylogenetic Group-1 (26) to which they belong.The structures of the human H2 HAs also allows location of
Fig. 3. Overlap of the receptor binding domains of H1, H2, and H3 HAs in complexes with receptor analogues. (A) The overlapped receptor binding sites ofhuman HAs for H1 A/S.Carolina/1918, (blue), H2 A/Singapore/1/57, (yellow) and H3 A/Aichi/2/68, (22) (gray) in complex with human receptor analogue, LSTc. (B)The receptor binding domains of avian HAs for H1, A/dk/Alberta/76 (blue), H2, A/dk/Ontario/77 (yellow), and H3, A/dk/Ukraine/63 (25) (gray) in complex withhuman receptor analogue. The sialopentasaccharides are colored according to the HAs to which they are bound and some of the side-chains discussed in thetext are shown in stick representation.
Table 1. Summary of the data resolution and PDB accession codes for each of the structures described in the text; fullcrystallographic statistics are given in the SI
HA subtype
H2 H1
Species Avian Human Avian Human
Virus New York Ontario Potsdam Singapore Japan Alberta 1918
Uncomplexed 2.4 Å 2WR0 2.6 Å 2WR5 – 2.7 Å 2WRC 3.0 Å 2WRD – –Avian receptor 2.4 Å 2WR2 2.5 Å 2WR3 – 3.1 Å 2WRB – – –Human receptor 2.1 Å 2WR1 2.5 Å 2WR4 3.1 Å 2WRF 2.5 Å 2WR7 3.0 Å 2WRE 3.0 Å 2WRH 3.0 Å 2WRG
17178 � www.pnas.org�cgi�doi�10.1073�pnas.0906849106 Liu et al.

previously deduced antigenically significant amino acids nearresidues 137, 189, 140, 200, and 214 (7, 8) and all five H2 HAstructures confirm the group-specific identity and positions ofresidues near the ‘‘fusion peptide’’ that are involved in mem-brane fusion activation (26). Our focus here, on the receptorbinding properties of the HAs of the prototype 1957 viruses andof their potential avian precursors, reflects the importance ofHA-receptor interactions in the initial stages of infection and ininterspecies transmission. Having 1918 H1 and 1957 H2 HA-receptor analogue complexes to complement those available for1968 H3 HA (22, 25), enables comparison of the receptorbinding sites and of the basis for the receptor binding specificityof the HAs of all three pandemic viruses.
On the basis of amino acid sequence and structure, H2 HA is verysimilar to H1 (21) but quite distinct from H3 (23). However,comparison of the complexes formed between human H1, H2, andH3 HAs with a human receptor shows that the H2 binding modeis much more similar to H3 than H1. Two main features distinguishthe complexes: the lower position of Gal-2 in the H1 binding site,and the different orientation of the third, fourth, and fifth residuesof the bound sialopentasaccharide, which is almost parallel with the3-fold axis of symmetry of the H1 HA, compared with a morefolded-back conformation in the H2 and H3 sites (Fig. 3A). HumanH2 and H3 HAs share the amino acid substitutions Gln-226Leu andGly-228Ser by comparison with avian HAs, and with human H1HA, which retains the avian Gln-226/Gly-228 sequence. The dif-ferences in receptor positioning might, therefore, be a directconsequence of these differences in sequence. However, examina-tion of complexes formed by avian H2 HA with human receptor(Fig. 3B) suggests an alternative. Despite the characteristic avianGln-226/Gly-228 sequence in the avian H2 HA binding site, theconformation adopted by the human receptor is the same as that inthe human H2 HA human receptor complex. The positioning of thereceptor, therefore, appears not to be related directly to theGln-226/Gly-228 sequence but to an alternative feature of the site.
The avian H2 HA-human receptor complex is remarkablywell-ordered by comparison with equivalent avian H1 and avianH3 complexes. The protein environment near Gal-2 is quitesimilar in all three cases but it is only in H2 that there iswell-defined electron density for the second and third sugars ofthe human receptor analogue. The most significant sequencedifferences that might be involved in this feature appear to be atresidue 186, which is an asparagine in H2, proline in H1, andserine in H3. The role of Asn-186 in contacting Gln-226 and the4� OH of Gal-2 through a water molecule, is illustrated in Fig.2B, and it is likely that the absence of this residue and of thehydrogen-bonded network that involves Gln-226, contributes tothe relatively poor binding of human receptor by these avian H1and H3 HAs. Interestingly, Asn-186 does not play this role inhuman H2 binding to human receptor. In that case the Gln-226Leu substitution means that the hydrogen bond networkinvolving Asn-186 is not formed. Instead the Asn-186 side-chainpoints away from the receptor binding site, adopting the sameorientation as in the unliganded structure.
The significance of the ability of HAs to bind both avian andhuman receptors has been the subject of debate, particularlyconcerning whether mixtures of human and avian viruses havebeen isolated (17, 32) or whether modifications have occurredduring adaptation to growth in the laboratory in avian ormammalian cells (11, 17). Nevertheless, it would appear to be aclear advantage for an avian virus, with the overall geneticpotential to cause a pandemic, to be able to bind to humanreceptors like the H2 avian HA described here, to gain an initial
foothold for infection of humans. Under selective pressure thesubsequent Gln-226Leu mutation would be expected to increaseaffinity for the human receptor and decrease affinity for theavian receptor (Fig. 2 C and D). The latter effect would be anadvantage in avoiding interaction with avian receptor-rich, res-piratory-tract mucins that would otherwise block infection (17).Early in H1, H2, and H3 pandemics, single receptor binding sitemutants were detected, Gln-226Leu for H2 and H3 and Glu-190Asp for H1 (30, 33). The additional characteristic mutationsGly-228Ser in H2 and H3 and Gly-225Asp in H1 seem to havemore limited effects on affinity and specificity and were pre-sumably secondary acquisitions (30). It is also feasible that someavian viruses containing H1, H2, and H3 HAs have a particularpropensity, compared with those containing HAs of othersubtypes, for trans-species infection. Consistent with this possi-bility, there have been suggestions of ‘‘recycling’’ of these threesubtypes in humans: in 1890 and 1968 for H3, and 1918, 1977, and2009 for H1 viruses, although the proposal for an H2 epidemicat the end of the nineteenth century, as well as in 1957 (34)appears less likely (35). At this stage it could be informative, notleast with regard to pandemic planning, to extend investigationsof this possibility, (12, 31, 36, 37) using robust assays of bindingspecificity on a substantial number of avian viruses from all 16HA subtypes, to see if there are other subtypes that haverepresentatives with the ability to bind human receptors.
MethodsAll HAs, except the 1918 H1 human, were from viruses grown in hens’ eggs andprepared by bromelain digestion, as previously described (28). The gene for theH1 human 1918 HA was made synthetically (38) and the protein expressed inrecombinant vaccinia virus infected CV1 cells, as previously described (39) andisolated by bromelain digestion. Crystallization conditions were screened by thesitting-drop vapor diffusion method using Crystal Clear strips (Douglas Instru-ments). The nano-drops were set up with 0.1 �L of BHA protein solution (10mg/mL in 10 mM Tris�HCl, pH 8.0) and 0.1 �L of reservoir solution by using anOryx-8 robot (Douglas Instruments). Crystals were grown at 18 °C using thefollowing reservoir solutions; H2 human Singapore 32% PEG 400, 2% PEG3350,0.1M Mes pH6.5; H2 human Japan 30% PEG 400, 0.1M Mes pH6.0; H2 avian duckOntario 30 PEG 2K MME, 0.1M Bis Tris propane at pH 6.5; H2 avian New York 25%PEG 3350, 0.2M Ammonium acetate, 0.1M HEPES at pH 6.8; H2 avian Potsdam22% PEG 3350, 0.1M Tris at pH 8.0; H1 avian 1976 2.3 M AS, 1% PEG 400, 0.1MPipes pH 7.0; H1 human 1918 2.2M Ammonium sulfate, 1% PEG 400, 0.1M Bis Trispropane pH 5.5. Crystals were prepared for flash cooling by serial transfer intoreservoir solution augmented by 20–25% PEG400. Crystals for datasets withbound receptor were prepared by soaking the crystals over night in either 4 mMLSTa(�2,3-linkedgalactose)or4mMLSTc (�2,6-linkedgalactose)madeupincryobuffer.All crystalswerescreenedonaRaxis4detector (100-mmscan)mountedona Rigaku MicroMax 007 HF generator. Ligand soaked crystals of H1 avian 1976 HAwere collected at Daresbury SRS stations 14.2 and 9.6 using ADSC Quantum Q4detectors. All of the other data were collected on beamlines IO3/IO4 at theDiamond Light Source at 100K with an ADSC Q315. Diffraction data were inte-grated using Denzo and scaled with Scalepack (40). Ellipsoidal Truncation andAnisotropic Scaling are performed using the server: http://www.doe-mbi.u-cla.edu/�sawaya/anisoscale/ (41). H2 A/dk/ Ontario/77 HA was solved by molec-ular replacement with Phaser (42) using H1, A/PR/8/34 HA, (21). All other H2 HAswere solved using H2 A/dk/ Ontario/77 HA as the search model. Standard refine-ment, with Refmac (43) and PHENIX (44), and manual model building with Coot(24), was performed on all of the structures. Crystallographic statistics are givenin the SI, together with the relevant PDB accession codes. Molecular figures werecreated with Pymol (http://pymol.sourceforge.net/; 45).
ACKNOWLEDGMENTS. Research at the National Institute for Medical Re-search is funded by the Medical Research Council (U.K.). R.J.R. thanks theScottish Funding Council. We thank R.G. Webster of St Jude Research Institute,Memphis for seeds of the avian H2 viruses and the World Health OrganizationCollaborating Centre for Influenza Reference and Research at National Insti-tute for Medical Research for HA sequences.
1. Lim KA, Smith A, Hale JH, Glass J (1957) Influenza outbreak in Singapore. Lancet273:791–796.
2. Meyer HM, Jr, Hilleman MR, Miesse ML, Crawford IP, Bankhead AS (1957) New antigenicvariant in Far East influenza epidemic, 1957. Proc Soc Exp Biol Med 95:609–616.
3. Chu CM (1958) The etiology and epidemiology of influenza: An analysis of the 1957epidemic. J Hyg Epidemiol Microbiol Immunol 2:1–8.
4. Scholtissek C, Rohde W, Von Hoyningen V, Rott R (1978) On the origin of the humaninfluenza virus subtypes H2N2 and H3N2. Virology 87:13–20.
Liu et al. PNAS � October 6, 2009 � vol. 106 � no. 40 � 17179
MIC
ROBI
OLO
GY

5. Kawaoka Y, Krauss S, Webster RG (1989) Avian-to-human transmission of the PB1gene of influenza A viruses in the 1957 and 1968 pandemics. J Virol 63:4603– 4608.
6. WHO (1980) A revision of the system of nomenclature for influenza viruses: A WHOmemorandum. Bull World Health Organ 58:585–591.
7. Schafer JR, et al. (1993) Origin of the pandemic 1957 H2 influenza A virus and thepersistence of its possible progenitors in the avian reservoir. Virology 194:781–788.
8. Makarova NV, Kaverin NV, Krauss S, Senne D, Webster RG (1999) Transmission ofEurasian avian H2 influenza virus to shorebirds in North America. J Gen Virol 80:3167–3171.
9. Skehel JJ, Wiley DC (2000) Receptor binding and membrane fusion in virus entry: Theinfluenza hemagglutinin. Annu Rev Biochem 69:531–569.
10. Gottschalk A (1959) Chemistry of virus receptors. The Viruses 3:51–61.11. Rogers GN, Paulson JC (1983) Receptor determinants of human and animal influenza
virus isolates: Differences in receptor specificity of the H3 hemagglutinin based onspecies of origin. Virology 127:361–373.
12. Connor RJ, Kawaoka Y, Webster RG, Paulson JC (1994) Receptor specificity in human,avian, and equine H2 and H3 influenza virus isolates. Virology 205:17–23.
13. Paulson JC, Sadler JE, Hill RL (1979) Restoration of specific myxovirus receptors toasialoerythrocytes by incorporation of sialic acid with pure sialyltransferases. J BiolChem 254:2120–2124.
14. Webster RG, Yakhno M, Hinshaw VS, Bean WJ, Murti KG (1978) Intestinal influenza:Replication and characterization of influenza viruses in ducks. Virology 84:268–278.
15. Naeve CW, Hinshaw VS, Webster RG (1984) Mutations in the hemagglutinin receptor-binding site can change the biological properties of an influenza virus. J Virol 51:567–569.
16. Baum LG, Paulson JC (1990) Sialyloligosaccharides of the respiratory epithelium in theselection of human influenza virus receptor specificity. Acta Histochem Suppl 40:35–38.
17. Couceiro JN, Paulson JC, Baum LG (1993) Influenza virus strains selectively recognizesialyloligosaccharides on human respiratory epithelium; The role of the host cell inselection of hemagglutinin receptor specificity. Virus Res 29:155–165.
18. Matrosovich MN, Matrosovich TY, Gray T, Roberts NA, Klenk HD (2004) Human andavian influenza viruses target different cell types in cultures of human airway epithe-lium. Proc Natl Acad Sci USA 101:4620–4624.
19. Weis W, et al. (1988) Structure of the influenza virus haemagglutinin complexed withits receptor, sialic acid. Nature 333:426–431.
20. Sauter NK, et al. (1989) Hemagglutinins from two influenza virus variants bind to sialicacid derivatives with millimolar dissociation constants: A 500-MHz proton nuclearmagnetic resonance study. Biochemistry 28:8388–8396.
21. Gamblin SJ, et al. (2004) The structure and receptor binding properties of the 1918influenza hemagglutinin. Science 303:1838–1842.
22. Eisen MB, Sabesan S, Skehel JJ, Wiley DC (1997) Binding of the influenza A virus tocell-surface receptors: Structures of five hemagglutinin-sialyloligosaccharide com-plexes determined by X-ray crystallography. Virology 232:19–31.
23. Wilson IA, Skehel JJ, Wiley DC (1981) Structure of the haemagglutinin membraneglycoprotein of influenza virus at 3 A resolution. Nature 289:366–373.
24. Emsley P, Cowtan K (2004) Coot: Model-building tools for molecular graphics. ActaCrystallogr D Biol Crystallogr 60:2126–2132.
25. Ha Y, Stevens DJ, Skehel JJ, Wiley DC (2003) X-ray structure of the hemagglutinin of apotential H3 avian progenitor of the 1968 Hong Kong pandemic influenza virus.Virology 309:209–218.
26. Russell RJ, et al. (2004) H1 and H7 influenza haemagglutinin structures extend astructural classification of haemagglutinin subtypes. Virology 325:287–296.
27. Winter G, Fields S, Brownlee GG (1981) Nucleotide sequence of the haemagglutiningene of a human influenza virus H1 subtype. Nature 292:72–75.
28. Ha Y, Stevens DJ, Skehel JJ, Wiley DC (2001) X-ray structures of H5 avian and H9 swineinfluenza virus hemagglutinins bound to avian and human receptor analogs. Proc NatlAcad Sci USA 98:11181–11186.
29. Rogers GN, et al. (1983) Single amino acid substitutions in influenza haemagglutininchange receptor binding specificity. Nature 304:76–78.
30. Matrosovich M, et al. (2000) Early alterations of the receptor-binding properties of H1,H2, and H3 avian influenza virus hemagglutinins after their introduction into mam-mals. J Virol 74:8502–8512.
31. Matrosovich MN, et al. (1997) Avian influenza A viruses differ from human viruses byrecognition of sialyloligosaccharides and gangliosides and by a higher conservation ofthe HA receptor-binding site. Virology 233:224–234.
32. Choppin PW, Tamm I (1960) Studies of two kinds of virus particles, which compriseinfluenza A2 virus strains: I. Characterisation of stable homogeneous substrains inreactions with specific antibody, mucoprotein inhibitors, and erythrocytes. J Exp Med112:895–920.
33. Reid AH, Fanning TG, Hultin JV, Taubenberger JK (1999) Origin and evolution of the1918 ‘‘Spanish’’ influenza virus hemagglutinin gene. Proc Natl Acad Sci USA 96:1651–1656.
34. Mulder J, Masurel N (1958) Pre-epidemic antibody against 1957 strain of Asiaticinfluenza in serum of older people living in the Netherlands. Lancet 1:810–814.
35. Dowdle WR (2006) Influenza pandemic periodicity, virus recycling, and the art of riskassessment. Emerg Infect Dis 12:34–39.
36. Gambaryan A, et al. (2005) Receptor specificity of influenza viruses from birds andmammals: New data on involvement of the inner fragments of the carbohydrate chain.Virology 334:276–283.
37. Blixt O, et al. (2004) Printed covalent glycan array for ligand profiling of diverse glycanbinding proteins. Proc Natl Acad Sci USA 101:17033–17038.
38. Stemmer WP, Crameri A, Ha KD, Brennan TM, Heyneker HL (1995) Single-step assemblyof a gene and entire plasmid from large numbers of oligodeoxyribonucleotides. Gene164:49–53.
39. Chen J, et al. (1998) Structure of the hemagglutinin precursor cleavage site, a deter-minant of influenza pathogenicity and the origin of the labile conformation. Cell95:409–417.
40. Otwinowski Z, Minor W (1993) Data Collection and Processing. In Data Collection andProcessing, eds Sawyer L, Isaacs N, Bailey S (SERC Daresbury Laboratory, Warrington,U.K.), pp 556–562.
41. Strong M, et al. (2006) Toward the structural genomics of complexes: Crystal structureof a PE/PPE protein complex from Mycobacterium tuberculosis. Proc Natl Acad Sci USA103:8060–8065.
42. McCoy AJ, et al. (2007) Phaser crystallographic software. J Appl Crystallogr 40:658–674.
43. Anonymous (1994) The CCP4 suite: Programs for protein crystallography. Acta Crys-tallogr D Biol Crystallogr 50:760–763.
44. Adams PD, et al. (2002) PHENIX: Building new software for automated crystallographicstructure determination. Acta Crystallogr D Biol Crystallogr 58:1948–1954.
45. DeLano WL (2002) The PyMOL Molecular Graphics System DeLano Scientific, (Palo Alto,CA).
17180 � www.pnas.org�cgi�doi�10.1073�pnas.0906849106 Liu et al.