studies insect segmentation: from teratology to...
TRANSCRIPT

Development 104 Supplement, II2-I21, (1988)Prinred in Great Britain @ The Company of Biologists Limited 1988
Studies insect segmentation: from teratology to phenogenetics
K. SANDER
Biologisches Institut 1 (Zoologie), Albert-Ludwigs Universitrit, D-78 Freiburg im Breisgau, Albertstrasse 21a, FRG
111
IN
Summary
Old and new data on the segmentation process inarthropods are considered mainly with reference tothe genetical basis of segmentation in Drosophila.Problems discussed are the universality (or otherwise)of parasegmental metam€ry, homeotic genes in insects
other than Drosophila, pair-rule traits in myriapods,various segmentation anomalies related to polar mor-phogenetic centres, and sequential segment determi-nation in the horseshoe crab. The opening and closing
paragraphs address the conceptual and terminologicalprogress from the study of 'developmental monsters'to the study of 'developmental genes' as a means ofanalysing segmentation.
Key words: insect segmentation, pair-rule organization,morphogenetic centres, progress zone model,experimental embryology.
lntroduction
Insect segmentation, for long of importance mainly tothe taxonomist and evolutionary biologist, has
increasingly attracted developmental and molecularbiologists interested in its ontogenetic generation. Inthis field, the last decade is characterized by a shift instrategies from teratology and experimental embry-ology (for their relationship see Oppenheimer, L968)
towards phenogenetics, from the formal interpret-ation of 'monsters' (see Cappe de Baillon, 1933;
Seidel , 196I; Sand er, 1976) to probing gene action indevelopment. The herculean efforts of Lewis (1978)
and Ntisslein-Volhard & Wieschaus (1980) at eluci-dating gene activities involved in Drosophila segmen-tation have triggered a veritable goldrush in this field,and knowledge both genetical and molecular is ac-
cumulating exponentially. The data on stripe patternexpression of segmentation genes (reviewed byAkam , L987) provide just one example, and one thathas led very close to a goal set by Valentin Haecker(1918) for the discipline he named 'Phtinogenetik': toestablish for each heritable variant the stage ofontogenesis when it begins to deviate from normaldevelopment. Another of Haecker's goals still seems
far distant, namely to describe and understand thecourse of aberrant ontogenesis from its first manifes-tations to the ultimate mutant phenotype. Most topics
to be discussed below fall into this category. As a pre-goldrush veteran in the field of experimental insectembryogenesis, I shall venture (1) to view some'teratological' data in the light of findings aird con-cepts that have sprung up durittg this last decade , (2)to discuss the validity of some of these conceptsbeyond the realm of Drosophila, and (3) to drawattention to some data from other arthropods thatmight shed light on insect, segmentation and itsevolution.
For easier going, I shall work my way back from themetameric endproducts of the segmentation process
to its beginnings. Before this, however, a glimpse ofthe degree of variation to be expected might help toprepare the reader's mind. Some parasitic hymenop-terans desynchronize the segmentation process to theextreme. The best-known example so far is providedby the genus Trichogramma where development fromthe egg stage to the minuscule adult occurs within theeggs of various host insects. In Trichogramma evanes-cens (Bront6 Gatenby, I9I7; Lee , 196I; Tanaka,1985), the tiny blastoderm turns directly into a bag-like larva barely capable of any movements otherthan gulping up the host's yolk. No signs of metamerycan be recognized before the imaginal discs formprior to pupation, except for a faint segmentation ofthe central nervous system. Which segmentationgenes and homeotic genes will have persisted in this

II2 K. Sander
system, and how and when do they interact? Suchquestions might be approached some day startingfrom honeybee molecular probes (see below).
(1) Are parasegments universal in insects?
Parasegmental metamery (see Lawrence, this vol-ume) has been demonstrated so far only in Dros-ophila. The question whether other insects, or insectsin general, also pass through a stage of parasegmentalorganization before visible segments form (Fig. 1)remains open. The answer may partly depend oncriteria and definitions. As the early expression ofcertain segmentation genes and homeotic genes inDrosophila occurs in parasegmental domains (see
e.g. Lawrence et al. 1987), one might try to visualizeparasegments in other insects by in situ hybridizationof suitable molecular probes. The discovery of thehomeobox (McGinnis et al. 1984; Laughon & Scott,1984) has provided the means to 'fish' for homeobox-containing genes in other insects (and eukaryotes in
Fig. l. Segmented insect germbands as seen in thescanning electron microscope. Arrows mark tracheal pitsin the 4th abdominal segment. (A) Colorado beetle,length about 1.1mm (courtesy of R. Fleig, Freiburg).(B) Honeybee, length about 1.5mm. The pits in thesegmental grooves will give rise to the oenocytes (Nelson,1915), the others are the tracheal pits (from Fleig &Sander, 1986). Note that in both species, the tracheal pits(at least in the abdomen) are located well within thesegmental territories whereas in Drosophila they arethought to occupy intersegmental positions (Martinez-Arias & Lawrence, 1985).
general, see Gehring, 1987). This approach has beenchosen in the migratory locust by Akam, Kornbergand Martinez-Arias (see their contributions to thisvolume), while we have started work on the honey-bee in cooperation with Walter Gehring's laboratoryat Basel.
In screens using the Antennapedia and engrailedhomeobox sequences from Drosophila for fishinghomeobox-containing sequences from a genomicDNA library of the honeybee, So far eight genes havebeen identified (Walldorf et al. unpublished results).Several of these are homologous to known homeoticgenes from Drosophila (see section 2) and might beused for in situ hybridizations that could revealparasegment boundaries. A further sequence showshomology to engrailed from Drosophila. However, a
fushi tarazu homologue has failed to show up in thehoneybee screens. Whether this means that meta-mery is generated by (slightly?) different gene inter-actions in the honeybee must remain open for thetime being. A potentially relevant finding is that thehoneybee shows no trace of germband retraction, theprocess that in Drosophila aligns the mesodermalparasegments with the segmental domains of theectoderffi, and thus provides for the proper attach-ment of somatic muscles (Akam & Martinez-Arias,1985; Lawfence & Martinez-Arias, 1985). This mightmean that (some?) genes expressed parasegmentallyin Drosophila could be expressed in the honeybeecoincident with segmental domains of the ectodermright from the beginning. If SO, would their ex-pression then be considered parasegmental or seg-mental? In any case, it is worth remembering theprofound differences between the modes of segmentformation in different insects (see Sander, 1976,,
1983), suggesting that gene interactions guiding de-velopment up till the germband stage might differmore between various insect forms than the genesactive in the germband itself.
(21 Homeotic genes
That homeosis (Bateson, 1894) can be heritable wasfirst shown in Drosophila (Bridges & Morgan, 1923)and the domestic silk moth (Sasaki, 1930, reviewed inTazima, 196/.) (Fig.2). More recently, severalhomeotic mutations have been described in flourbeetles of the genus Tribolium (see Sokoloff &Soliman, 1986; Beem&tr, 1987) and may now bestudied with molecular methods. Extensive homeotictransformations of unknown genetic causation wereobserved by M. Verdier in a locust strain selected forparthenogenetic development (see fig. 39 in Sander,re76).
In the honeybee, some homeotic genes are remark-
able for their degrees of homology to the respectiveDrosophila homeoboxes. In Scr, homology is 86 % atthe DNA level and 98 % in the derived peptidesequence, while in Dfd. the respective values are 82%and I00"/o. In situ hybridizatron has been achieved so
far only with the Dfd, homologous probe H42 (FIeig et
al. 1988). The cytoplasmic expression pattern in theblastoderm was found to resemble that described forDrosophila (McGinnis et al. 1984). One expectationtriggering our studies was that the honeybee shouldprovide better spatial and temporal resolution thanDrosophila since its egg is much larger and spends
many hours in the blastoderm stage (Fleig & Sander,1985). This difference might indeed account for somenovel details observed with the Dfd-homologousprobe. For instan ce, transcripts can be demonstratedall over the blastoderm for many hours, but only in arestricted patch inside each nucleus, and later onnuclear labelling is repressed in the immediate vicin-ity of the blastoderm zone where transcripts appear inthe cytoplasm (Fleig et al. 1988).
(3) Pair-rule organizalion: evolutionary originsand ontogenetic functions
In view of the surprising pair-rule genes (Ntisslein-Volhard & Wieschaus, 1,980), I would like to drawattention to segmentation in the myriapods. Both themajor taxa combined in this group (known as theDiplopoda and Chilopoda, respectively) reveal traitsof double-segment organization which merit atten-tion in their own right but even more so because ofthe widespread belief that insects could have origi-nated from the juvenile stages of diplopod-like ances-
tors.Among the (centipedes), some double-
Fig. 2. Silk moth germbands representing the wild-type(right) and the hom ozygotrs EN phenotype with eightabdominal segments bearing thorax-type leg buds. Thelarge anterior appendage buds (3 pairs) will give rise to
. the mouth parts (from Itikawa , 1952).
Studies in insect segmentation 113
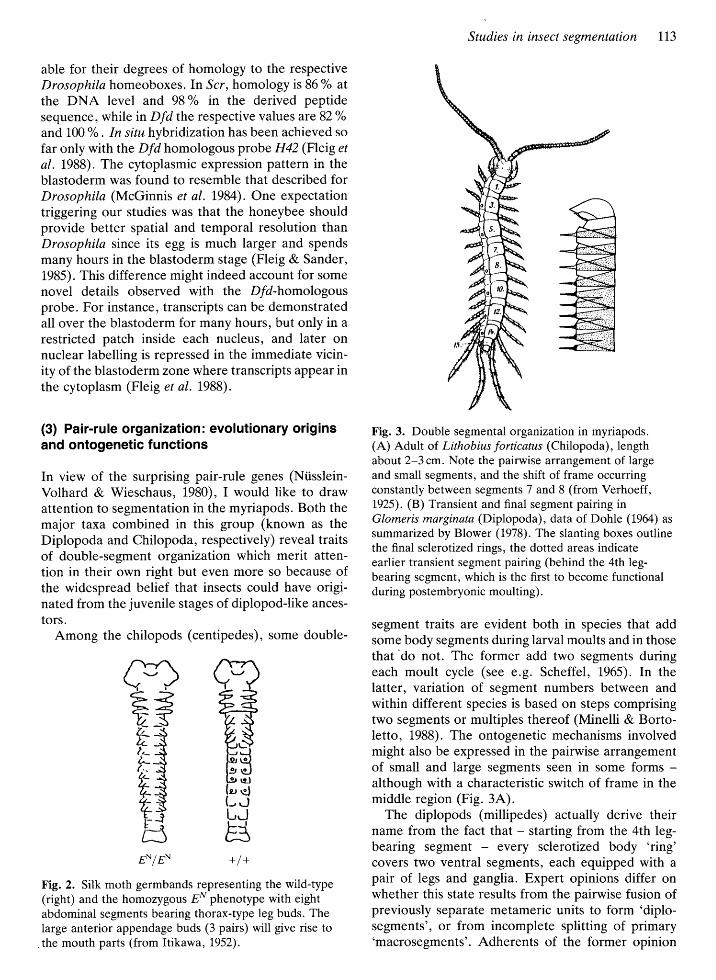
Fig. 3. Double segmental organization in myriapods.(A) Adult of Lithobius forticatus (Chilopoda), lengthabout 2-3 cm. Note the pairwise arrangement of largeand small segments, and the shift of frame occurringconstantly between segments 7 and 8 (from Verhoeff,L925). (B) Transient and final segment pairing inGlomeris marginata (Diplopoda) , data of Dohle (1964) as
summarrzed by Blower (1978). The slanting boxes outlinethe final sclerotized rings, the dotted areas indicateearlier transient segment pairing (behind the 4th leg-bearing segment, which is the first to become functionalduring postembryonic moulting).
segment traits are evident both in species that addsome body segments during larval moults and in thosethat'do not. The former add two segments duringeach moult cycle (see e.g. Scheffel , 1965). In thelatter, variation of segment numbers between andwithin different species is based on steps comprisingtwo segments or multiples thereof (Minelli & Borto-letto, 1988). The ontogenetic mechanisms involvedmight also be expressed in the pairwise arrangementof small and large segments seen in some formsalthough with a characteristic switch of frame in themiddle region (Fig. 3A).
The diplopods (millipedes) actually derive theirname from the fact that - starting from the 4th leg-bearing segment every sclerotized body 'ring'covers two ventral segments, each equipped with a
pair of legs and ganglia. Expert opinions differ onwhether this state results from the pairwise fusion ofpreviously separate metameric units to form 'diplo-segments', or from incomplete splitting of primary'macrosegments'. Adherents of the former opinion
c"#usHHl, elLJUJE3+l+
chilopods
CAEF&* -sh{
a$!-{trf
r*/r*
LI4 K. Sander
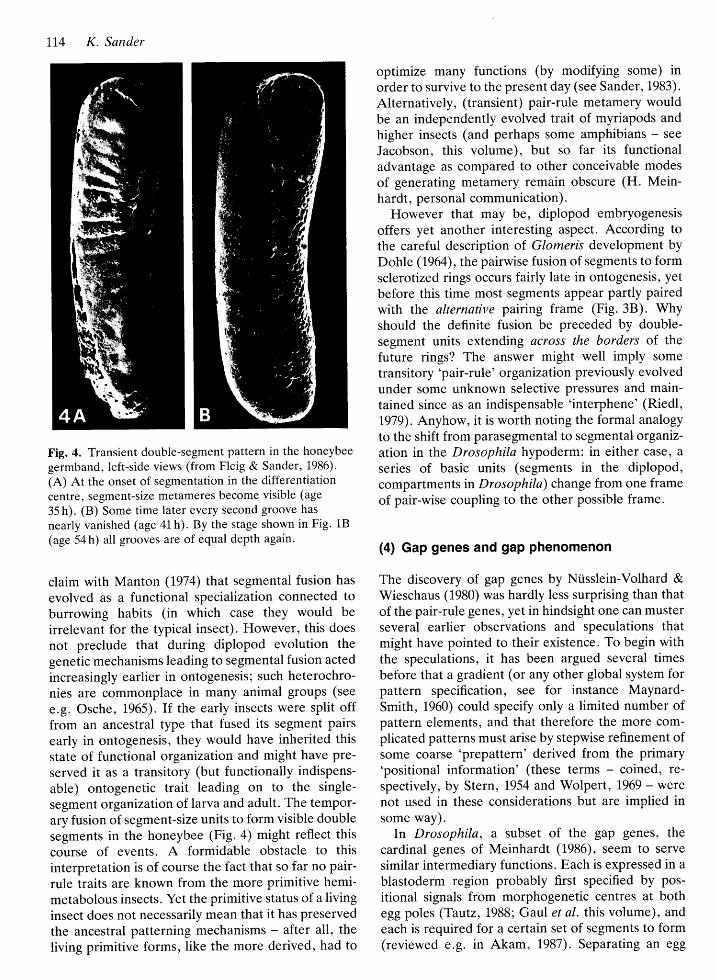
Fig. 4. Transient double-segment pattern in the honeybee
germband, left-side views (from Fleig & Sander, 1986).
(A) At the onset of segmentation in the differentiationcentre, segment-size metameres become visible (age
35 h). (B) Some time later every second groove has
nearly vanished (age 41 h). By the stage shown in Fig. 18
(age 54 h) all grooves are of equal depth again.
claim with Manton (1974) that segmental fusion has
evolved as a functional specializatron connected toburrowing habits (in which case they would be
irrelevant for the typical insect). However, this does
not preclude that during diplopod evolution thegenetic mechanisms leading to segmental fusion acted
increasingly earlier in ontogenesis; such heterochro-nies are commonplace in many animal groups (see
e.g. Osche, 1965). If the early insects were split offfrom an ancestral type that fused its segment pairsearly in ontogenesis, they would have inherited thisstate of functional organrzation and might have pre-served it as a transitory (but functionally indispens-
able) ontogenetic tratt leading on to the single-
segment organtzation of larva and adult. The tempor-ary fusion of segment-size units to form visible doublesegments in the honeybee (Fig . 4) might reflect thiscourse of events. A formidable obstacle to thisinterpretation is of course the fact that so far no pair-rule traits are known from the more primitive hemi-metabolous insects. Yet the primitive status of a livinginsect does not necessarily mean that it has preservedthe ancestral patterning mechanisms - after all, the
living primitive forffiS, like the more derived, had to
optimize many functions (bV modifying some) inorder to survive to the present day (see Sander, 1983).
Alternatively, (transient) pair-rule metamery wouldbe an independently evolved trait of myriapods andhigher insects (and perhaps some amphibians - see
Jacobsor, this volume), but so far its functionaladvantage as compared to other conceivable modes
of generating metamery remain obscure (H. Mein-hardt, personal communication).
However that may be, diplopod embryogenesisoffers yet another interesting aspect. According tothe careful description of. Glomeris development byDohle (1964), the pairwise fusion of segments to formsclerottzed rings occurs fairly late in ontogenesis, yetbefore this time most segments appear partly pairedwith the alternative pairing frame (Fig.3B). Whyshould the definite fusion be preceded by double-segment units extending across the borders of thefuture rings? The answer might well imply some
transitory'pair-rule' organi zation previously evolvedunder Some unknown selective pressures and main-tained since as an indispensable 'interphene' (Riedl,1979). Anyhow, it is worth noting the formal analogy
to the shift from parasegmental to segmental organiz-ation in the Drosophila hypoderm: in either case, a
series of basic units (segments in the diplopod,compartments in Drosophila) change from one frameof pair-wise coupling to the other possible frame.
(4) Gap genes and gap phenomenon
The discovery of gap genes by Ni.isslein-Volhard &Wieschaus (1980) was hardly less surprising than thatof the pair-rule genes, yet in hindsight one can musterseveral earlier observations and speculations thatmight have pointed to their existence. To begin withthe speculations , it has been argued several timesbefore that a gradient (or any other global system forpattern specification, see for instance Maynard-Smith, 1960) could specify only a limited number ofpattern elements, and that therefore the more com-plicated patterns must arise by stepwise refinement ofsome coarse 'prepattern' derived from the primary'positional information' (these terms coined , te-spectively, by Stern, 1954 and Wolpert, 1969 - werenot used in these considerations but are implied insome way).
In Drosophila, a subset of the gap genes, thecardinal genes of Meinhardt (1986), seem to serve
similar intermediary functions. Each is expressed in ablastoderm region probably first specified by pos-
itional signals from morphogenetic centres at bothegg poles (Tautz, 1988; Gaul et al. this volume), andeach is required for a certain set of segments to form(reviewed e.g. in Akam, 1987). Separating an egg

into two parts (fragments), a key experiment in thedays of experimental teratology (reviewed in Sander,1976), should interfere with the local interactionsbetween some cardinal genes, especially hb, Kr andkni. These genes seem to inhibit their expressionmutually during blastoderm formation and thus each
restricts the region(s) where its neighbour(s) willfinally be expressed (Jiickle et al. 1986) .
If one of the cardinal genes should fail to becomeactive because part of the egg cell was removed byligaturing, the 'adjacent' cardinal gene might beexpressed over an extended territory, as was found byJiickle et al. (1986) in mutants lackin g a cardinal gene.Consequently, the incomplete segment patternsforming in egg fragments should comprise a constantsegment number over a certain range of fragmentsizes, and should grow by the simultaneous additionof several segments once the fragment is made so
large that the cardinal gene required for those seg-
ments can start functioning; similar considerations,but focusing on interfaces rather than territories,have been published by Meinhardt (L986).
The joint formation or failure of several segmentshas indeed been observed in experiments on thehoneybee embryo by Schnetter (1934). He coined theterm 'Potenzbereich' (potency region) for egg regionswhose presence in a posterior egg fragment would berequired for a certain set of segments gnathal,thoracic or anterior abdominal - to form (reviewed inSander, L976, fig .27); however, experiments on otherstrains of the honeybee (Maul , 1970; see Sander,1976) have failed to confirm this concept. A trend forcertain segments to form either together or not at allhas also been recognized in certain body regions ofthe leafhopper Euscelis and of Drosophila, and was
indeed ascribed to requirements for local interaction(Vogel, 1977 , L978). Another instance might be
reflected in the frequencies of heat-shock pertur-bance that vary between different abdominal seg-
ments of the migratory locust (see Fig. 9). Whysimilar symptoms were not observed in other bodyregions or in other species (e.g. Brltchidius: Jung,L966; Protophormia: Herth & Sander, L973), mustremain open for the present. Possible explanationsrange from the regression of undersized segmentanlagen (after their initial specification, see e.g.Carroll et al. 1986; Frohnh6fer & Ntisslein-Volhardt,1987) to the possibility that cardinal genes are notuniversal among insects. This possibility might at firstsight seem remote because the primary subdivision ofthe insect body in regions called tagmata (gnatho-cephalor, thorax and abdomen) seems to call forgenes specifying groups of segments. But these genes
could (at least initially) have acted in combinatorialrather than hierarchical fashion, specifying not thetagma as a functional unit but rather, in each segment
Studies in insect segmentation 115
anlage, the properties distinguishing all segments inthe tagma from the others (see Akam, this volume).
Finally, it should be noted that, by present knowl-edge, the gap genes cannot entirely account for the'gap phenomenon' defined earlier (Sander, 1976).The gap in the middle of the segment file is observedalmost universally when insect eggs or embryonicrudiments are separated into two fragments early indevelopment, and it decreases in extent with increas-ing age at fragmentation (Fig. 5). Primarily thisphenomenon must be due to the interruption of long-range effects from anterior and posterior morphogen-etic centres (see below) while gap-gene functions, os
discussed above, would account for a stepwise ratherthan continuous variation of gap size. The 'asymmet-ric' reduction of pattern gaps shown in Fig. 5 couldreflect an asymmetry of polar centre (andf or gapgene) interactions similar to that revealed by fatemaps of mutant Drosophila eggs (Ntisslein-Volhard etal. 1987).
(5) Polar morphogenetic centres, polarfocusing and terminal prevalence
In the long-germ type of insect development (Krause,1939), the metameric subdivision of the blastoderm is
Fig. 5. 'Asymmetric' decrease of the segment gap in eggs
of Pimpla turionellae (Hymenoptera) ligatured at variousstages and levels (data of Jung et al. 1977; from Krause,1981). Scale at the left represents blastodermal fate map(P, procephaloni G, mouth part segments; T, thorax; A,abdomen). I-4 stages at ligation (early cleavage, latecleavage, early blastoderm, late blastoderm) . n, numberof cases. Columns correspond to segments formed inanterior (hatched) and posterior fragments (cross-hatched) of eggs ligatured at about 70, 50 and 40 % egglength, respectively (panels from left to right) (posteriorpole : 0 o/"). Note age-invariant patterns formed inanterior fragments, and the decrease of the pattern gap
due to the increase with age in the number of segmentsformed in the posterior fragments.

116 K. Sander
Fig. 6. Results from centrifugation of Drosophila eggs
soon after egg deposition, reversing once the position inthe centrifuge (first one pole centrifugal, then the other).(A) Double-abdomen cuticle from centrifuged eg9,ventral side at the left. Note reduction of terminalstructures despite double polarity revealing posteriorcentre activities at both poles. (B) The only doublecephalon obtained so far, same orientation (compositephotograph, the thin line at the right represents the edgeof the broken vitelline envelope). Preparation courtesy P.
Aichele, Freiburg.
governed by long-range influences from the egg poles(Sander, 1976). In Drosophila, the role of morpho-genetic polar centres was demonstrated by ChristianeNtisslein-Volhard and her coworkers in a masterlycombination of genetical analysis and transplantationexperiments (reviewed by Frohnh6fer et al. 1986 andNi.isslein-Volhard et al. 1987). The centres owe theirspecific activities to molecules synthesized in thenurse cells and exported to the growing oocyte duringoogenesis. In the oocyte, these molecules becomelocalized at the appropriate poles. The mechanismsinvolved differ apparently between anterior and pos-terior centre (Frigerio et ol. 1986; Sander 8L Leh-mann, 1988; see also Lehmatrn, this volume) and arecritically dependent on nurse cell functions (Bohr-mann & Sander, 1987).
We have tested the subsequent stability of polarlocalizations by centrifugation experiments withnewly deposited eggs (P. Aichele & K. Sander,unpublished results) (Fig. 6). Although the distri-bution of putative morphogens after centrifugationcould not yet be checked, the larval cuticle patternsdeveloping in centrifuged eggs permit several con-clusions. First, global alterations in the segment
pattern are frequent, but only if (1) the egg iscentrifuged within 15 min after deposition and (2) itsorientation in the centrifuge was reversed once ortwice ('reverse centrifugation'). Both these con-ditions seem to indicate that global changes in thesegment-determining system require strong disrup-tions in oocyte architecture. Secotrd, anterior andposterior factors differ greatly with respect to theirexpression in centrifuged eggs. Posterior centre ac-tivity is frequently expressed at both poles, leading tothe double-abdomen or double-posterior pattern(Fig. 6A), while anterior expression occurred, if atall, only at one of the poles (the original anterior orposterior pole); only in a single case was a doublecephalon formed (Fig. 68). This difference may re-flect the strict localization of anterior-determining bcdRNA (Frigerio et al. 1986) as contrasted to thewidespread initial distribution of posterior factorsthat can be inferred from transplantations of nursecell and oocyte cytoplasm (Sander & Lehmailtr,lggg).
The third conclusion concerns what might be called'polar focusing'. Strong deviations of the embryo'santeroposterior pattern axis from the longitudinal eggaxis were very rare in centrifuged Drosophila eggs.
Since one cannot assume that centrifugation enrichesanterior and posterior determinant molecules just inthe egg poles and nowhere else, this result mustdepend on some secondary mechanisms that allocateanterior, and especially posterior, centre activities tothe polar regions of the egg. Such focusing effectswere also observed by Ni,isslein-Volhard et al. (1987)after transplantation of posterior cytoplasm: if effec-tive at all, these transplantations lead to polarizingactivities centred right in the polar region even if thematerial itself was injected not in this pole but at somedistance from it. Injected anterior material, on theother hand, influenced patterning only around theinjection site (Niisslein-Volhard et al. 1987); themechanisms responsible for this restriction could bereflected in the low incidence (mentioned above) ofanterior expression in centrifuged eggs. Focusingmight be based on signal exchange between incipientblastoderm cells, and it could have evolved frommechanisms that serve to establish the initial polardifferences in the epithelial germ anlage of short-germ insects (see below).
Lower dipterans show an additional trait, namelythe absolute priority of the terminal pattern elementswith respect to final expression This 'terminal preva-lence', ?S it might be called, is documented by the factthat truncated segment patterns (lacking a terminalstructure (head or telson) at one or both ends) arerare among experimental results except after severedamage to the respective egg pole, while the loss ofsome segments in between can easily be induced
Studies in insect segmentation II7
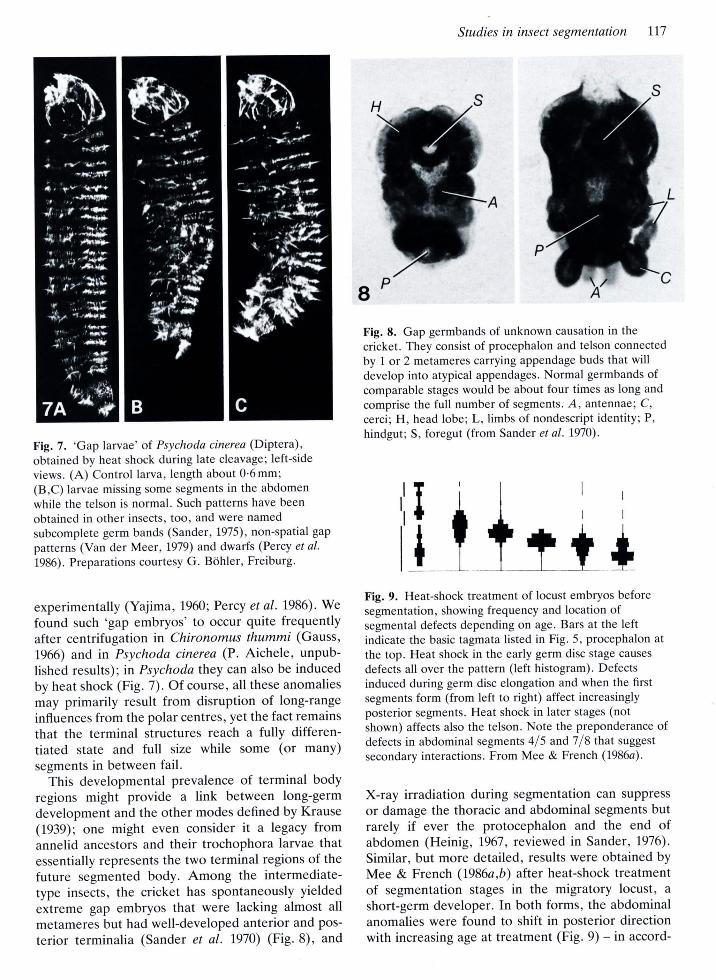
Fig. 7. 'Gap larvae' of. Psychoda cinerea (Diptera),obtained by heat shock during late cleavage; left-side
views. (A) Control larva, length about 0'6 mm;(B,C) larvae missing some segments in the abdomen
while the telson is normal. Such patterns have been
obtained in other insects, too, and were named
subcomplete germ bands (Sander,1975), non-spatial gap
patterns (Van der Me er, L979) and dwarfs (Percy et al.
1986). Preparations courtesy G. B6hler, Freiburg.
experimentally (Yajima ,1960; Percy et al. 1986). We
found such 'gap embryos' to occur quite frequentlyafter centrifugation in Chironomus thummi (Gauss,
1966) and in Psychoda cinerea (P. Aichele, unpub-lished results); in Psychoda they can also be inducedby heat shock (Fig .7). Of course, all these anomalies
may primarily result from disruption of long-rangeinfluences from the polar centres, yet the fact remains
that the terminal structures reach a fully differen-tiated state and full size while some (ot many)
segments in between fail.This developmental prevalence of terminal body
regions might provide a link between long-germdevelopment and the other modes defined by Krause(1939); one might even consider it a legacy fromannelid ancestors and their trochophora larvae thatessentially represents the two terminal regions of the
future segmented body. Among the intermediate-type insects, the cricket has spontaneously yielded
extreme gap embryos that were lacking almost all
metameres but had well-developed anterior and pos-
terior terminalia (Sander et al. 1970) (Fig. 8), and
Fig. 8. Gap germbands of unknown causation in the
cricket. They consist of procephalon and telson connected
by 1 or 2 metameres carrying appendage buds that willdevelop into atypical appendages. Normal germbands ofcomparable stages would be about four times as long and
comprise the full number of segments. A, antennae; C,
cercii H, head lobe; L,, limbs of nondescript identity; P,
hindgut; S, foregut (from Sander et al. 1970).
t+Fig. 9. Heat-shock treatment of locust embryos beforesegmentation, showing frequency and location ofsegmental defects depending on age. Bars at the leftindicate the basic tagmata listed in Fig. 5, procephalon at
the top. Heat shock in the early germ disc stage causes
defects all over the pattern (left histogram). Defectsinduced during germ disc elongation and when the firstsegments form (from left to right) affect increasinglyposterior segments. Heat shock in later stages (notshown) affects also the telson. Note the preponderance ofdefects in abdominal segments 415 and 718 that suggest
secondary interactions. From Mee & French (I986a).
X-ray irradiation during segmentation can suppress
or damage the thoracic and abdominal segments butrarely if ever the protocephalon and the end ofabdomen (Heinig, 1967, reviewed in Sander, 1976).
Similar, but more detailed, results were obtained byMee & French (I986a,b) after heat-shock treatmentof segmentation stages in the migratory locust, a
short-germ developer. In both forms, the abdominalanomalies were found to shift in posterior directionwith increasing age at treatment (Fig. 9) - in accord-
II

118 K. Sander
ance with the visible segmentation that occurs somehours later.
(6) Sequential segmentation and segmentidentity
The progress zone model of patterning as proposedfor the chick wing by Summerbell et al. (1973) mightwith due modifications account for the findings in thecricket just mentioned (Sander, 1976), and also forthe segmentation anomalies observed after ligaturinglocust embryos (Mee, 1986). As published data onpresumed progress zone mechanisms in insects arescanty, it might be useful to discuss some relevantfindings from the horseshoe crab , Tachypleus triden-tatus, one of the most primitive living arthropods.Unfortunately, genetical data on this animal will beslow in coming, because each larval instar of. Tachy-pleus takes about a year and maturity is reached onlyafter some 15 to 20 years.
The horseshoe crab produces its posterior seg-ments one after another (Itow, 1985, 1986) - in themode of short-germ-type insects but at a slower pace,requiring about 2 days per segment as compared to a
few hours in the locust (Mee & French , I986a).Another trait that Tachypleus shares with short-germinsects is its capability to form several individualgermbands in a single egg if the blastodermal cells aremade to aggregate in several patches rather than in asingle germdisc. In Tachypleus, this can be effectedby removal of Ca2+ ions (Itow, 1985), in the stonecricket Tachycines and the beetle Atrachya by mech-anical cuts or by cold treatment, respectively(Krause, L934; Miya & Kobayashi , 1974).
In Tachypleus, inhibition of DNA synthesis byS-phase blockers for 12 to 24h during the segmen-tation process leads to peculiar disturbances (Itow,1986). They are characterrzed by the addition of one(sometimes two) segments in excess of the normalnumber, combined with a mixed identity in one orseveral segments. The older the embryo at the time oftreatment, the more posterior starts the anomaly inthe file of segments - this rule must reflect sequentialsegment determination, os in the short-germ insectsdiscussed above.
Itow's explanation for his results (Itow, 1986) is noteasily understood but I feel what is relevant for thepresent considerations can be summarized as follows(Fig. 10). The identity of each segment is due to amaster gene or combination of genes. During or afterthe S-phase, a switch in accessibility from one seg-ment-identity gene (or combination of genes) to thenext is effected by some mechanism depending on theprevious switching step. The switch is thought torequire DNA replication. The expression of the
sr ---o-D-Tf E " Ta-b b-c c-d d-€ -e.^ar
; * --0--!-O-O- cr'ilti#t
Fta -b -c -d -e rlifiit€ s ,,il??olBit
r-T-cbr
s*Fig. 10. Interpretation of results from blocking DNAsynthesis in segmenting embryos of the horseshoe crabTachypleus (top and middle panels, based on Itow, 1986),and extension of this interpretation to cover results fromheat-shock or X-ray treatment of segmenting short-germinsect embryos (bottom panel). Capital letters denotemorphological segment identity, small letters indicate themaster gene(s) accessible at the times when cytoplasmicperiodicity (top row) triggers master gene expression.Time period of blocking treatment indicated by hatchedboxes. The model is based on the following findings(Itow, 1986). (1) The first segment forming after removalof the inhibitor acquires a mixed character correspondingpartly to the identity due to appear in this segment, andpartly to the preceding segment . (2) Behind the mixedsegment several segments may perpetuate the patterninganomaly, combining the characters of subsequentsegment pairs (e.g. AlB,BlC etc. if, to begin with, theprospective segment B was transformed to A/B). The(first) mixed segment (in the case quote d, Af B) increasesthe total number of segments by one. (3) The older theembryo at the time of treatment, the more posterior doesthe anomaly start in the file of segments. In the bottompanel, segment A has been shifted (broken arrow) to theposition of the absent segment, in order to indicate thephysical continuity of segments (non-spatial gap!).
accessible master genes, however, is coupled toperiodic changes in cell metabolism which are notslowed down by the inhibitors used. When DNAsynthesis is experimentally delayed for part of a
cytoplasmic cycle, the switch in gene accessibility getsstuck in an intermediary state, thus causing theintermediary character of the nascent segment (Itow,1986); alternatively, the cytoplasmic clock mightenforce gene expression during the period when thegenome switches from one master gene (or genecombination) to the next. Occasionally, the phasediscrepancy between DNA status and cytoplasmiccycling may continue while some subsequent seg-ments form, due to dependence of each genomic

Switch on the previous one. Itow's model assumes a
Sequential arrangement, or the chromosome, ofgenes specifying successive segments, but this view is
based on considerations of Lewis (1978) rather than
on independent evidence. However, if correct, his
model could help understanding the evolution of a
gene system like the Bithorax Complex whose (polar-ized) molecular organi zation somehow reflects the
spatial pattern of the segments under its control (see
Ingham, this volume).But the model also has implications for insect
segmentation. Itow (1986) explains the increase insegment number by assuming that segmentationcomes to an end when the gene-switching mechanism
has activated the master gene(s) specifying the ter-
minal body region (e in Fig. 10). After tempotatyinhibition of gene switching, this genomic state will be
reached with some delay which gives the cytoplasmicoscillator the time for initiating yet another segment
(Fig. 10 top). With reference to short-germ insects' itis interesting to consider the possible consequences ofthe inverse desynchronizatton A temporaty arrest ofthe cytoplasmic clock while DNA switching was
continued would cause segmentation to terminate at a
lower than normal total number of segments (Fig. 10
bottom). Morphologically the segment identities lost
should be those that were due for specification whilethe cytoplasmic clock was arrested, and the pos-
teriorly following segments might, with appropriatedurations of arrest, be of mixed identity (as after
DNA blockitrg, see Fig. L0 top). This type of anomalyresembles the effects of heat-shock treatment and
X-irradiation on short-germ segmentation described
by Heinig (1967) and Mee & French (1986a,b) as
mentioned above. Like any model, Itow's certainlyhas its limitations, but it deserves attention because ofits dependence on DNA replication and its (formal)capacity to cover two seemingly opposite experimen-
tal effects - the increase and the reduction of segment
numbers, respectively.In closing, I would like to return to the historical
aspect and venture to envision the future course ofevents. Although present feelings may be different,the genetical analysis of development rs not going toreplace teratology and experimental embryology.
Understanding development as the phenogeneticinterplay of gene activities requires both the
geneticalf molecular and the experimental approach.For instance, the phenotypes produced by maternal-
effect mutations in Drosophila cannot be fully under-
stood without transplantation experiments. In lowerinsects and other arthropods unsuitable for genetical
analysis, on the other hand, spontaneous and exper-
imentally induced 'monsters' will continue to provide
the basis for analysing ontogenesis. However, knowl-edge obtained in this way will be greatly enhanced by
Studies in insect segmentation Il9
studies with appropriate molecular probes derivedfrom work on Drosophila.
I wish to thank Mrs Margit Scherer for patiently typing
the manuscript, Dr Richard Fleig for unpublished data and
useful criticism, and the Deutsche Forschungsgemeinschaft
for supporting our investigations.
References
Arnu, M. (1987). The molecular basis for metamericpattern in the Drosophila embryo . Development l0l,1,-22.
Arnu, M. E. & MnRTINEZ-Anns, A. (1985). Thedistribution of. Ultrabithorax transcripts rn Drosophilaembryos. EMBO I. 4, 1689-1700.
BnrBsoN, W. (1894) . Materials for the Study of VariationTreated with Especial Regard to Discontinuity in the
Origin of Species. London and New York: Macmillan.BBsueN, R. W. (1987). A homoeotic gene cluster in the
red flour beetle . Nature, Lond. 327,247-249.BrowsR, J. G. (1978). Anamorphosis in the
Nematophora. Verh. naturwiss. Ver. Hamburg (NF)ztf 22,97-103.
BoHnuANN, J. & SnNoBn, K. (1987). Aberrant oogenesis
in the patterning mutant dicephalic of Drosophilamelanogaster: time lapse recordings and volumentry invitro. Roux's Arch. devl Biol. 196,279-285.
BnrocBS, C. B. & MoncnN, T. H. (1923). The third-chromosome group of mutant characters of. Drosophilamelanogaster. Publ. Carnegie Inst. Wash. 327 .
BnoNrf GnrBNny, J. (I9I7). The embryonic developmentof- Trichogramma evanescens, Westw., monembryonicegg parasite of Donacia simplex. Q. I. microsc. Scl. 62,
1,49-187 .
Cnppn pp BntttoN, P. (1933) . Tdratologie compar6e duPlatycnemis pennipes Pall. (Odonates, Agrionidae) et
du Carausius morosus Br. (Orthoptera, Phasmidae).
Bull. biol. Fr. Belg. 67 , l-33.Cannorl, S. B., WrNStow, G. M., ScHUPBAcH,T. &
Scott, M. P. (1986). Maternal control of. Drosophilasegmentation gene expression. Nature, Lond. 323,278-280.
Dourp, W. (1964). Die Embryonalentwicklung vonGlomeris marginata (Villers) i- Vergleich zurEntwicklung anderer Diplopoden. Zool. Ib. (Anat.) 81,
24r-3r0.Frnrc, R. & SaNnpn, K. (1985). Blastoderm development
in honeybee embryogenesis as seen in the scanning
electron microscope . Int. J. Invert. Reprod. Dev. 8,
279-286.Frnrc, R. & SeNosn, K. (1986). Embryogenesis of the
honeybee Apis melliferaL. (Hymenoptera: Apidae): an
SEM study. Int. J. Insect Morphol. Embryol. 15,
M9-462.Fruc, R., WILLDoRF, LJ., GEHnING, W. J. & SnNnEn, K.
(1988). In situ locahzation of the transcripts of ahomeobox gene in the honeyb ee Apis mellifera L.(Hymenoptera). Roux's Arch. devl Biol. 197,269-274.
FnrcBnIo, G., Bunnl, M., Bonr, D., BRunacARTNEn, S. &

120 K. Sander
Non, M. (1986). Structure of the segmentation genepaired and the Drosophila PRD gene set as part of agene network. Cell 47,735-746.
FnoHNHornR, H. G., LnuuANN, R. & NUssLETN-
Vornnno, C. (1986). Manipulating the anteroposteriorpattern of the Drosophila embryo. J. Embryol. exp.Morph. 97 Suppl. 1,69-179.
FnonNuornR, H. G. & NUssLETN-VorHnRo, C. (1987).Maternal genes required for the anterior localization ofbicoid activity in the embryo of Drosophila. GenesDevelop. 1, 880-890.
Gruss, IJ. (1966). Der Einfluss der Zentrtfugalkraft aufEiarchitektur und embryonale Differenzierung vonChironomus th. thummi (Diptera). Staatsexamens-arbeit, Fakultiit fi.ir Biologie, Universitrit Freiburg.
Gpsnmc, W. J. (1987). Homeo boxes in the study ofdevelopment . Science 236, 1245-1252.
Hnscrnn, V. (1918) . EntwicklungsgeschichtlicheEigenschaftsanalyse ( Phcinogenetik) . GemeinsameAufgaben der Entwicklungsgeschichte, Vererbungs- undRassenlehre. Jena: Fischer.
HerNrc, S. (1967). Die Abiinderung embryonalerDifferenzierungsprozesse durch totaleRontgenbestrahlung im Ei von Gryllus domesticus.Zool. Jb. (Anat.) 84, 425-492.
Hnnru, W. & SnNoER, K. (L973). Mode and timing ofbody pattern formation (regionalizatton) in the earlyembryonic development of cyclorrhaphic dipterans(Protophormia,, Drosophila). Wilhelm Roux' Arch.Entwmech. Org. 172, I-27.
IrxawA, N. (1952). Genetical and embryological studieson the E-multiple alleles in the silkworm, Bomby*moriL. Bull. Sericult. Exp. Station (Tokyo) 14,86-91.
Irow, T. (1935). The effect of Caz+-free sea water on thebody segmentation in the horseshoe crab (Chelicerata,Arthropoda) . Acta Embryol. Morph. exp.N. S. 6,15-29.
Irow, T. (1986). Inhibitors of DNA synthesis change thedifferentiation of body segments and increase thesegment number in horseshoe crab embryos(Chelicerata, Arthropoda) . Roux's Arch. devl Biol.1g5, 323-333.
JAcrrE, H. ,Txlrz, D., ScHuu, R., Snrrnnr, E. &LsHi\anNN, R. (1986). Cross-regulatory interactionsamong the gap genes of Drosophila. Nature, Lond.324, 668-670.
JuNc, E. (1966). Untersuchungen am Ei desSpeisebohnenkzifers Bruchidius obtectus SAY.(Coleoptera) . Wilhelm Roux' Arch. Entwmech. Org.157,320-392.
JuNc, E., Nuss, E. & Worr, R. (1977). Geschni.irtePimpla-Eier zeigen nur im hinteren TeilembryoSegmentausfall: Sind abgeiinderte Ooplasma-stromungen die Ursache? Verh. dt. Zool. Ges. 1977,307 .
Kna.use, G. (L934). Analyse erster Differenzierungs-prozesse im Keim der Gewiichshausschrecke durchkiinstlich erueugte Zwillings-, Doppel- undMehrfachbildungen . Wilhelm Roux' Arch. Entwmech.Org. 132, II5-205.
Knauss, G. (1939). Die Eitypen der Insekten. Biol. Zbl
59, 495-536.KnnusB, G. (1981). Homology studies on insect egg
systems. In Progress in Developmental Biology (ed. H.W. Sauer). Fortschr. d. Zool. 26,307-333. Stuttgart:Fischer-Verlag.
LlucuoN, A. & Scorr, M. P. (1984). Sequence of aDrosophila segmentation gene: Protein structurehomology with DNA-binding proteins. Nature, Lond.310, 25-3r.
LnwnBNCE, P. A. & MnnrrNpz-ARrAS, A. (1985). The celllineage of segments and parasegments in Drosophila.Phil. Trans. R. Soc. Lond.8.312, 83-90.
LnwnENCE, P. A., JouNSToN, P., MecooNALD, P. &Srnuur, G. (1987). The fushi-tarazu and even-skippedgenes delimit the borders of parasegments inDrosophila embryos. Nature, Lond. 328, 440-442.
LEp, Tsur-YrNc (197I). The development ofTrichogramma evanescens Westw. and its influence onthe embryonic development of its host, Attacus cynthiaricini Boisd. Acta entomologica sinica 10,352-354.
LEwIs, E. B. (1978). A gene complex controllingsegmentation in Drosophila. Nature, Lond. 276,565-570.
MnNroN, S. M. (1974). Segmentation in Symphyla,Chilopoda and Pauropoda in relation to phylogeny.Symp. zool. Soc. Lond. 32, 163-190.
Menrrxsz-Anra.s, A. & LewRENcE, P. (1985).Parasegments and compartments in the Drosophilaembryo . Nature, Lond. 3L3, 639-642.
MRur, V. (1970). Uber Schniirungsexperimente an Eiernzweier verschiedener Inzuchtstdmme der Honigbiene.Zool. Anz. Suppl. 33, 53-58.
MeyNnno-SrvnrH, J. (1960). Continuous, quantized andmodal variation. Proc. Roy. Soc. Lond. B 152,397 -409 .
McGrNNrs, W., LBvrNB, M. S., HanBN, E., KunorwA, A.& GBHRTNG, W. J. (1984). A conserved DNA sequencein homoeotic genes of the Drosophila Antennapediaand bithorax complexes. Nature, Lond. 308, 428-433.
MnE, J. E. (1986). Pattern formation in fragmented eggsof the short germ insect Schistocerca gregaria. Rottx'sArch. devl Biol. 195, 506-512.
MnB, J. E. & FnpNcH, V. (I986a). Disruption ofsegmentation in a short germ insect embryo. I. Thelocation of abnormalities induced by heat shock. "I.Embryol. exp. Morph. 96,245-266.
Mnn, J. E. & FnBNCH, V. (I986b). Disruption ofsegmentation in a short germ insect embryo. II. Thestructure of segmental abnormalities induced by heatshock. J. Embryol. exp. Morph. 96,267-294.
Mnmulnor, H. (1986). Hierarchical inductions of cellstates: A model for segmentation in Drosophila. J. CellSci. Suppl. 4, 357-381.
MrNBru, A. & BonrolErro, S. (1988). Myriapodmetamerism and arthropod segmentation. J. Linn. Soc.33, 323-343.
MryA., K. & Kosey.LsHr, Y. (1974). The embryonicdevelopment of Atrachy a menetriesi F aldermann(Coleoptera, Chrysomelidae). II. Analyses of earlydevelopment by ligation and low temperaturetreatment. J. Fac. Agric. Iwate Univ. 12,39-55.

NsrsoN, J. A. (1915) . The Embryology of the Honey Bee.
Princeton University Press.
NUssrerN-VoTHARD , C. & WTBscHAUS, E. (1980).
Mutations affecting segment number and polarity inDrosophila. Nature, Lond. 287, 795-801.
NUssrsn-VorHARD, C., FnonNHorEn, H. G. &LsHtunNN, R. (1987). Determination of anteroposteriorpolarity in Drosophila. Science 238, L675-1681.
OppBNHEIMER, J. M. (1968). Some historical relationshipsbetween teratology and experimental embryology. Bull.History of Medicine 42, I45-L59.
OscHE, G. (1965). Grundzi.ige der allgemeinenPhylogenetik. In Handbuch der Biologie (ed. F.
Gessner), Vol. 3, pp. 8I7-906. Frankfurt: Akad.Verlagsges. Athenaion.
Pnncy, J., KuuN, K. L. & KnrrHonn, K. (1986). Scanning
electron microscopic analysis of spontaneous and UV-induced abnormal segment patterns in Chironomussamoensis (Diptera, Chironomidae). Roux's Arch. devlBiol. I95,92-102.
Rreor, R. (1979). Order in Living Organisms: a Systems
Analysis of Evolution. New York: Wiley.S^q.NoBn, K. (1975). Pattern specification in the insect
embryo. In Cell Patterning, Ciba Found. Symp . 29
(New series) 24I-263.SnNoBn, K. (1976). Specification of the basic body
pattern in insect embryogenesis. Adv. Insect. Physiol.12, 125-238.
SaNosn, K. (1983). The evolution of patterningmechanisms: gleanings from insect embryogenesis and
spermatogenesis. In Development and Evolution (ed.B. C. Goodwin, N. Holder & C. C. Wylie), PP.I37-I59. Cambridge Univ. Press.
Sl.Nonn, K., HnntH, W. & Vortuln, H. (1970).
Abwandlungen des metameren Organisationsmusters infragmentierten und in abnormen Insekteneiern. Zool.Anz. Suppl. 33, 46-52.
SnNoBn, K. & LBuMANN, R. (1988) . Drosophila nurse
cells produce a posterior signal required for embryonicsegmentation and polarity. Nature, Lond. 335, 68-70.
ScHBrrnt, H. (1965). ElektronenmikroskopischeUntersuchungen tiber den Bau der Cerebraldri.ise der
Chilopoden. Zool. Jb. (Physiol.) 71,624-&0.ScHNnrrER, M. (1934). Physiologische Untersuchungen
i.iber das Diff erenzierungszentrum in der
Studies in insect segmentation I21'
Embryonalentwicklung der Honigbiene . Wilhelm Roux'Arch. Entwmech. Org. L3L, 285-323.
SnroBr, F. (1961). Entwicklungsphysiologische Zentrenim Eisystem der Insekten. Zool. Anz. Suppl. 24,
121,-1,42.
Sorororr, A. & SorrMAN, M. H. (1986). Mutantsclassified according to body parts affected. TriboliumInform. Bulletin 26, LI9-I39.
SrnnN, C. (1954). Two or three bristles. Am. Scient. 42,
213-247.
SuurusRBELL, D., LnwIS, J. S. & Wotrnnt, L. (1973).
Positional information in chick limb morphogenesis.
Nature, Lond. 244, 492-496.TnNarA, M. (1985). Embryonic and early post-embryonic
development of the parasitic wasp , Trichogrammachilonis (Hymenoptera, Trichogrammatidae). In Recent
Advances in Insect Embryology in Japan (ed. H. Ando& K. Miya). pp. 181-189. Tsukuba: ISEBU Co. Ltd.
Tturz, D. (1988). Regulation of the Drosophilasegmentation gene hunchback by two maternalmorphogenetic centres. Nature, Lond. 332, 28I-284.
Tnzuun, Y. (1964). The Genetics of the Silkworm.London: Logos Press.
VaN DER Mnnn, J. M. (1979). The specification ofmetameric order in the insect Callosobruchus maculatus
Fabr. (Coleoptera). J. Embryol. exp. Morph. 51 ,I-26.VenHoEFF, K. W. (1925). Klasse Chilopoda. In H. G.
Bronn's Klassen und Ordnungen des Tier-Reichs. Vol.V 12. Leipzig: Akad. Verlagsges.
Vocpr, O. (1977). Regionalisation of segment-formingcapacities during early embryogenesis rn Drosophilamelanogaster. Wilhelm Roux' Arch. devl Biol. 182,
9-32.Vocnr, O. (1978). Pattern formation in the egg of the
leaflropper Euscelis plebeius Fall. (Homoptera):Developmental capacities of fragments isolated fromthe polar egg regions . Devl Biol. 67 , 357 -370
Worpsnt, L. (1969). Positional information and thespatial pattern of cellular differentiation. "I. theor. Biol.25, L-47 .
Ysnua, H. (1960). Studies on embryonic determinationof the harlequin-fly, Chironomus dorsalis. I. Effects ofcentrifugation and of its combination with constrictionand puncturing. J. Embryol. exp. Morph. 8, L98-21'5.