su pplementar y information - media.nature.com · 3.3 dna-protein ladder model (dpl) we now turn to...
TRANSCRIPT

SupplementaryMethods
Quantitative analysis of cooperative binding of cGASalongDNA
1 Introduction
ThecatalyticactivityofcGASisactivatedbybindingtoDNA,whichinducesastructuralchangethatproperlyformstheactivesite. InitialstructuralstudiesshowedthattwocGASmoleculesform a heterotetrameric complex with two DNA ligands. Hereby, the two cGAS moleculessandwich2,approx.30°angledDNAduplexmolecules.cGASbindsapprox.16-20basepairsofeachDNAligand.However,dsDNAthatinprinciplefullyspansthecGASdimer(20bp)doesnotactivatetheenzymetoanyappreciableamountsinvitroandinhumancellsinvivoaswell.AnactivatingtransitionisobservediftheDNAlengthisincreasedto40-50bp.LongerDNAactivatescGASevenmorerobustly,andlongplasmidDNAisa"goldstandard"forcGASactivation.Evenifweaccountfor“DNAend”effects,i.e.theenzymefallingoffendsmorerapidlythandissociationfrominternalDNAbindingsitesthroughBrownianmotion,theevidentincapabilityofshortbluntendedDNAstoactivatecGASandtheincreasingactivitywithincreasingDNAlength(keepingthemolarityofbasepairsconstant)isnotexplainedbythecGASdimermodel.OurstructuralresultsindicatethatoncetheDNAligandsarelongenoughtobindtwocGASdimersnexttoeachother,aDNA:proteinnetworkisformedthatresemblesatwistedladder.
2 DNA-ProteinLadderModel(DPL)
OurnewcrystalstructuressuggestthatpairsofcGASdimersmutuallystabilizeeachotherviaaDNA:proteinnetwork:cGASdimersarepositionedliketherungsofaladder,withthetwoDNAstrandsbeingthebeams.AmathematicaltreatmentforthecooperativebindingofcGASdimersto DNA can be formulated along the general ideas of theMonod-Wyman-Changeux (MWC)modelforcooperativetransitionsinproteins:proteinsbindasmonomersalongasingleduplexofDNAwithadissociationconstantK1[mol/l].AlongtwoparallelDNAduplexes,suchasthoseprearrangedinthevicinityofacGAS:DNAheterotretramer,twocGASmoleculesbindwithanoverallmicroscopicdissociation constantK2[mol/l]. Sinceweassume for themodel that twocGASmolecules bind simultaneously, themacroscopic dissociation constant for assembly ofcGAS dimerswith two parallel DNAmoleculeswould be K22, analogous to the empirical Hillequation. Furthermore, the binding of two cGAS:DNA heterodimer into a cGAS2:DNA2heterotetramerischaracterizedbyadissociationconstantK3[mol/l].Theoverallbindingschemeisdepictedin(Fig.3a).Themodelincludessimplifications.Inparticular,weneglectbindingofsingle cGASmolecules to the prearranged "dimeric"DNA lattice and the equilibrium can beformulated as: 2𝑐𝐺𝐴𝑆 + 2𝐷𝑁𝐴
*++ 𝑐𝐺𝐴𝑆,: 𝐷𝑁𝐴,. Furthermore, we neglect interactions betweenDNAmoleculeswithunequalnumberofboundcGASmolecules. Suchanassumptioncanbejustified,ifinteractionbetweenDNAmoleculeswithequalnumberofboundcGASmoleculesismuch stronger than interaction between DNA molecules with unequal numbers of cGAS,becauselatterhas"less"interactingcGASdimersbetweenthetwoDNAmolecules.
WWW.NATURE.COM/NATURE | 1
SUPPLEMENTARY INFORMATIONdoi:10.1038/nature23890

3 MathematicalModelling
3.1 NomenclatureDNA-ProteinLadderModel(DPL)
InderivinganexpressionfortheactivityasafunctionoftheconcentrationandlengthsofDNAligandsandtheconcentrationofprotein,weusethefollowingterminology:
p0: totalconcentrationofprotein(cGASmonomer)l0: totalconcentrationofDNAmoleculess: numberofbindingsitesperDNAmoleculepilj: complexofjDNAwithicGASmoleculesK1: equilibriumdissociationconstantcGASmonomerwithDNAK2: equilibriumdissociation constant for binding of cGAS to prearrangedDNAmolecules,
formingthecGAS2:DNA2complexK3: equilibrium dissociation constant for the interaction of two cGAS:DNA complexes,
formingthecGAS2:DNA2complexa: aconstantrelatingtheconcentrationofDNAboundcGAStotheobservedrateofproduct
formation
3.2 Generaldimerequilibrium
Ingeneral,theformationofadimer,e.g.theassociationoftwoDNAmoleculesbycGAS,canbedescribedinitssimplestformwiththefollowingequilibriumscheme:
𝑙 + 𝑙*𝑙, (1.1)
with𝐾 = 1off1on.Usingtheresultingequations
𝑙 ∗ 𝑙 = 𝐾𝑙, (1.2)𝑙6 = 𝑙 + 2𝑙, (1.3)
onecanderiveexpressionforlandl2asafunctionofl0andK:
𝑙 = − 89𝐾 − 𝐾 𝐾 + 8𝑙6 (1.4)
𝑙, =8;𝐾 + 4𝑙6 − 𝐾 𝐾 + 8𝑙6 (1.5)
Iflhassomeformofassociatedactivitya1(e.g.fluorescence,enzymaticactivity)andl2arelatedproperty 2*a2 (since l2 has two "active" sites), one can combine equations (1.4 and 1.5) tocomputetheoverallactivityA:
𝐴 = 89
𝑎, − 𝑎8 𝐾 + 4𝑎,𝑙6 − 𝑎, − 𝑎8 𝐾 𝐾 + 8𝑙6 (1.6)
With𝑎 = 𝑎, − 𝑎8,𝑏 = 𝑎,andK'=K/4oneobtains
𝐴 = 𝑎𝐾′ + 𝑏𝑙6 − 𝑎 𝐾′ 𝐾′ + 2𝑙6 (1.7)
WWW.NATURE.COM/NATURE | 2
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

3.3 DNA-ProteinLadderModel(DPL)
Wenowturntotheschemeoutlinedin(Fig.3a)andderiveanequationfortheconcentrationofDNAboundcGAS.Indoingso,weassumethatundersteadystateconditions,ATP/2APTP,GTPandcGAMP/fGAMPhavenosubstantialinfluenceonK1,K2andK3.Theexperimentallyobservedsteady state activity A of product formation (in our case measured by (∆F/∆t: change influorescencepertime)canbewrittenas
𝐴 = 𝑎8 ∗ 𝑖 ∗ 𝑝B𝑙si=1 + 𝑎, ∗ 2𝑖 ∗ 𝑝,B𝑙,s
i=1 (2.1)
witha1anda2constantsthatrelatetheconcentrationofDNAboundcGASmonomers(e.g.p1l)anddimers(e.g.p2l2)totherateofproductformation.Thefactorof2inthesecondsumaccountsforthetwo"active"sitesineachp2il2.Foragivennumberofbindingsitess,atequilibrium,theschemein(Fig.3a)hasthefollowingequilibriumequations:
a) conservationofproteinandDNAligand:
𝑝6 ≈ 𝑝 (2.2)
𝑙6 = 𝑙 + 2𝑙, + 𝑝B𝑙GBH8 + 2𝑝,B𝑙, (2.3)
To derive a mathematical expression, in 2.1 a simplification is used: we designed ourexperimentswithforthemostpartasurplusofproteinoverDNAbindingsites,assumingthatone molecule of cGAS covers ~20 bp of DNA. For the purpose of deriving an analyticmathematicalequation,weassumep0>>s*l0,i.e.p≈p0.
b) bindingofcGAStoDNA:weassumethatcGAScanbindanywhereonDNA.TheDNAhassnon-overlappingbindingsites(e.g.oneper20bp)allsbindingsitesareequal.
G∗IJ∗KILK
= 𝐾8 (2.4)
GM8 ∗IJ∗ILK,∗I+K
= 𝐾8 (2.5)
...
GM BM8 ∗IJ∗INOLKB∗INK
= 𝐾8 (2.6)
...
IJ∗IPOLKG∗IPK
= 𝐾8 (2.7)
LikewiseforthecGASdimersboundtotwoDNAmolecules(l2denotestheprearranged“DNAdimer”):
G∗IJ+∗K+I+K+
= 𝐾,, (2.8)
...
WWW.NATURE.COM/NATURE | 3
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

GM BM8 ∗IJ+∗I+NO+K+
B∗I+NK+= 𝐾,, (2.9)
... IJ+∗I+PO+K+
G∗I+PK+= 𝐾,, (2.10)
Finally,wehavetheinteractionoftwocGASboundDNAmoleculesp1l.Weonlyneedoneequation;allothersareredundantduetothermodynamiclinkage. ILK∗ILK
I+K+= 𝐾Q (2.11)
Foragivens,theequations2.1a,2.2,...2.7cannowbeusedtoeliminateallbutonepiloronep2il2.Repeatingthisprocedureforeachofthepilandp2il2,weobtainasetofexpressionsfortheconcentrationsofthepilandp2il2(i=1...s)asfunctionsofs,p0,l0,K1,K2andK3.For example, let's look at the case s=3. In Mathematica code, we formulate the followingexpressionsforequations2.2,...2.10:eq1 = l0 - (l + p1l + p2l + p3l +2*l2+2*p2l2 + 2*p4l2 + 2*p6l2) eq2 = 3*p0*l - k1*p1l eq3 = 2*p0*p1l - 2*k1*p2l eq4 = p0*p2l - 3*k1*p3l eq5 = 3*p0*p0*l2 - k2^2*p2l2 eq6 = 2*p0*p0*p2l2 - 2 k2^2*p4l2 eq7 = p0*p0*p4l2 - 3 k2^2*p6l2 eq8 = p1l*p1l - k3*p2l2
In equilibrium, eq0=0, eq1=0, eq2=0 ... We now use these expressions in Mathematica toeliminatelandallpilbutpl:Eliminate[{eq1==0,eq2==0,eq3==0,eq4==0,eq5==0,eq6==0,eq7==0,eq8==0},{l,l2,p2l,p3l,p2l2,p4l2,p6l2}]
Thisprocedureresultsinthefollowingequation:p1l (k1^3 k2^4 k3 p0 + 3 k1^2 k2^4 k3 p0^2 + 3 k1 k2^4 k3 p0^3 + k2^4 k3 p0^4 + 2 k1^2 k2^6 p1l + 6 k1^2 k2^8 p0^2 p1l + 6 k1^2 k2^4 p0^4 p1l + 2 k1^2 p0^6 p1l) == 3 k1^2 k2^4 k3 l0 p0^2
Theequationcanbesolvedforp1lwithSolve[p1l (k1^3 k2^4 k3 p0 + 3 k1^2 k2^4 k3 p0^2 + 3 k1 k2^4 k3 p0^3 + k2^4 k3 p0^4 + 2 k1^2 k2^6 p1l + 6 k1^2 k2^4 p0^2 p1l + 6 k1^2 k2^2 p0^4 p1l + 2 k1^2 p0^6 p1l) == 3 k1^2 k2^4 k3 l0 p0^2, {p1l}]
resultinginasolutionthatdescribestheconcentrationofpilasafunctionofs=3,p0,l0,K1,K2:p1l = (k1^3 k2^4 k3 p0 - 3 k1^2 k2^4 k3 p0^2 - 3 k1 k2^4 k3 p0^3 - k2^4 k3 p0^4 + Sqrt((k1^3 k2^4 k3 p0 + 3 k1^2 k2^4 k3 p0^2 + 3 k1 k2^4 k3 p0^3 + k2^4 k3 p0^4)^2 + 12 k1^2 k2^4 k3 l0 p0^2 (2 k1^2 k2^6 + 6 k1^2 k2^4 p0^2 + 6 k1^2 k2^2 p0^4 + 2 k1^2 p0^6)))/ (4 (k1^2 k2^6 + 3 k1^2 k2^4 p0^2 + 3 k1^2 k2^2 p0^4 + k1^2 p0^6)) p2l = (-k1^3 k2^4 k3 p0^2 - 3 k1^2 k2^4 k3 p0^3 - 3 k1 k2^4 k3 p0^4 - k2^4 k3 p0^5 + Sqrt((k1^3 k2^4 k3 p0^2 + 3 k1^2 k2^4 k3 p0^3 + 3 k1 k2^4 k3 p0^4 + k2^4 k3 p0^5)^2 + 12 k1 k2^4 k3 l0 p0^4 (2 k1^3 k2^6 + 6 k1^3 k2^4 p0^2 + 6 k1^3 k2^2 p0^4 + 2 k1^3 p0^6)))/ (4 (k1^3 k2^6 + 3 k1^3 k2^4 p0^2 + 3 k1^3 k2^2 p0^4 + k1^3 p0^6)) p3l = (-k1^3 k2^4 k3 p0^3 - 3 k1^2 k2^4 k3 p0^4 - 3 k1 k2^4 k3 p0^5 - k2^4 k3 p0^6 +
WWW.NATURE.COM/NATURE | 4
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

Sqrt(4 k2^4 k3 l0 p0^6 (6 k1^4 k2^6 + 18 k1^4 k2^4 p0^2 + 18 k1^4 k2^2 p0^4 + 6 k1^4 p0^6) + (k1^3 k2^4 k3 p0^3 + 3 k1^2 k2^4 k3 p0^4 + 3 k1 k2^4 k3 p0^5 + k2^4 k3 p0^6)^2))/(12 (k1^4 k2^6 + 3 k1^4 k2^4 p0^2 + 3 k1^4 k2^2 p0^4 + k1^4 p0^6))
Inserting the three expression into 𝐴8 = 𝑎8 ∗ 𝑖 ∗ 𝑝B𝑙s
i=1 (first half of (eq. 2.1)), one canformulatethefollowingsum(s=3):
𝐴8 = 𝑎8 ∗ 𝑓8 ∗3𝑖
𝑖𝑝6B
𝐾8BT8
Q
BH8
with (3.1)
𝑓8 =−𝐾,9𝐾Q 𝐾8 + 𝑝6 Q + 𝐾,9𝐾Q 𝐾,9𝐾Q 𝐾8 + 𝑝6 U + 24𝐾89𝑙6 𝐾,, + 𝑝6, Q
12 𝐾,, + 𝑝6, Q
Generalizingsresultsinthefollowingexpression:
𝐴8 = 𝑎8 ∗ 𝑓8 ∗𝑠𝑖
𝑖𝑝6B
𝐾8BTGM,
G
BH8
with (3.2)
𝑓8 =−𝐾,,GM,𝐾Q 𝐾8 + 𝑝6 G + 𝐾,,GM,𝐾Q 𝐾,,GM,𝐾Q 𝐾8 + 𝑝6 ,G + 8𝑠𝐾89GM9𝑙6 𝐾,, + 𝑝6, G
4𝑠 𝐾,, + 𝑝6, G
Thesummationhasanexplicitexpression:
𝑠𝑖
𝑖𝑝6B
𝐾8BTGM,
G
BH8
= 𝐾8,𝑠𝑝6
𝐾8 + 𝑝6𝐾8 + 𝑝6𝐾8,
G
Withthisexpressionandreformulation,3.2canbewrittenas: (3.3)
𝐴8 = −𝑎84
𝑝6𝐾8 + 𝑝6
∗𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G
−𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G 𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G
+ 8𝑠𝑙6
Notethatthisexpressionhasthegeneralformof(1.4).Usingthesameprocedure,oneobtainsanexpressionforA2: (3.4)
𝐴, =𝑎,4
𝑝6,
𝐾,, + 𝑝6,∗
𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G
+ 4𝑠𝑙6
−𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G 𝐾8,𝐾Q𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G
+ 8𝑠𝑙6
Inourexperimentswestudy theactivationof cGASas functionofDNA lengthandkeep theeffectiveconcentrationofbindingsitesleff=l0*sconstant.CombiningA1andA2resultsinthefinalformulafortheconcentrationofcGASboundtoDNAandhencesteadystateactivityAofproductformationasafunctionofs:
WWW.NATURE.COM/NATURE | 5
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

𝐴(𝑠) = 𝑎𝐾′(𝑠) + 𝑏𝑙eff − 𝑎 𝐾′(𝑠) 𝐾′(𝑠) + 2𝑙eff (3.5)with
𝑙eff = 𝑠𝑙0
𝐾′(𝑠) =𝐾8,𝐾Q4𝐾,,
𝐾,, 𝐾8 + 𝑝6 ,
𝐾8, 𝐾,, + 𝑝6,
G
𝑎 =𝑎,𝑝6,
𝐾,, + 𝑝6,−
𝑎8𝑝6𝐾8 + 𝑝6
𝑏 = [+IJ+
*++TIJ+
3.4 DPLModel:SpecialCasesandSimplifications
a)K3→∞(no“dimer”state)ATaylorseriesexpansionof(3.5)aroundleffresultsaround0yields
𝐴 𝑠 = −𝑎 + 𝑏 𝑙eff +𝑎2𝐾′
𝑙eff, −𝑎
2𝐾′,𝑙effQ ⋯
henceforK3→∞(thereforeK’→∞),𝐴 𝑠 = −𝑎 + 𝑏 𝑙eff = 𝑎8
𝑙eff𝑝6𝐾8 + 𝑝6
which describes bind of cGAS monomers on a single DNA ligand with s binding sites andconcentrationl0.b)K3→0(no“monomer”state)SettingK3=0in(3.5)yields
𝐴 𝑠 = 𝑏𝑙eff = 𝑎,𝑙eff𝑝6,
𝐾,, + 𝑝6,
withdescribesa(maximally)cooperativeformationofcGASdimerson(prearranged)DNAwithsbindingsitesandaconcentrationofl0.BindingalongDNAofcGASdimersisnon-cooperative.c)K1≫p0ThisconditionisareasonableassumptionforourstudiessincetheexperimentallydeterminedDNAbindingaffinityforshortDNAisonly~20µM.Inthiscase(3.5)simplifiesto: 𝐴(𝑠) = 𝑎 𝐾′(𝑠) + 𝑙eff − 𝐾′(𝑠) 𝐾′(𝑠) + 2𝑙eff (3.6)with
𝑙eff = 𝑠𝑙0
𝐾′(𝑠) = 𝐾8′𝐾,,
𝐾,, + 𝑝6,
G
𝐾8] =𝐾8,𝐾Q4𝐾,,
𝑎 = 𝑎,𝑝6,
𝐾,, + 𝑝6,
WWW.NATURE.COM/NATURE | 6
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

Note,inthiscase,thereareonlytwoindependentbindingconstantsK2andK1’=K12K3/4K22.Eq.3.6istheoneusedtofittheexperimentaldatainourstudy.
4 FittingofexperimentaldataForglobalfittingofthedatamatrix(8differentDNAligandsby8differentcGASconcentrations),weusedthefminsearchprocedureasimplementedinMatlab_R2015a(TheMathWorks,Inc).4.1 FittingwithHillequations
First,thedatawerefittedusingasetofempiricalHillequations (4.1)
𝑉_BKK(𝑉 [a, 𝑠B, 𝐾B) = 𝑉 [a𝑙eff,i𝑝6
GN
𝐾B + 𝑝6GN
withVmaxfittedglobally(i.e.itisassumedtobethesameforallligands)andsi,KifittedforeachDNAligandliindividually.Here,leff,iistheconcentrationofDNAliganditimesitsnumberofbpsand leff,i=constant in all reactions. fminsearchwasused tominimize the following function (iligands, j protein concentrations, rij: experimentally measured rate for ligand i and proteinconcentrationj,wij:standarddeviationofrijfromthreeindependentexperiments):
𝑅𝑒𝑠 =1𝑤Bf
𝑉_BKK 𝑉 [a, 𝑠B, 𝐾B − 𝑟B,f,
B,f
Minimizationof17parameters(Vmax,8si,8Ki)resultedinanR2=0.991.Fig.S1(leftpanel)showstheexperimentaldataalongwiththefitofthesetofHillequations,therightpanelisaplotofthe“Hillcoefficients”siasafunctionofDNAlength. Ingeneral,thedatashowanincreasein“cooperativity”forlongerDNAasexpected.
FigureS1.Leftpanel:Experimentaldata(seealsolegendforFig.3c)fittedtoasetofHillequationswithaglobalVmax,andindividualKi,si.Plottedistherateofsubstrateturnover(∆F/∆t[RFUmin-1])asa
WWW.NATURE.COM/NATURE | 7
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890
TheempiricalHillequation,althoughbeingabletogenerallyfitthedatawithanR2=0.991,hasnodirectphysicalinterpretation,becauseitisunclearhowKiandsicorrelatewiththeunderlyingmolecularevents.4.2 FittingwithDPLequation
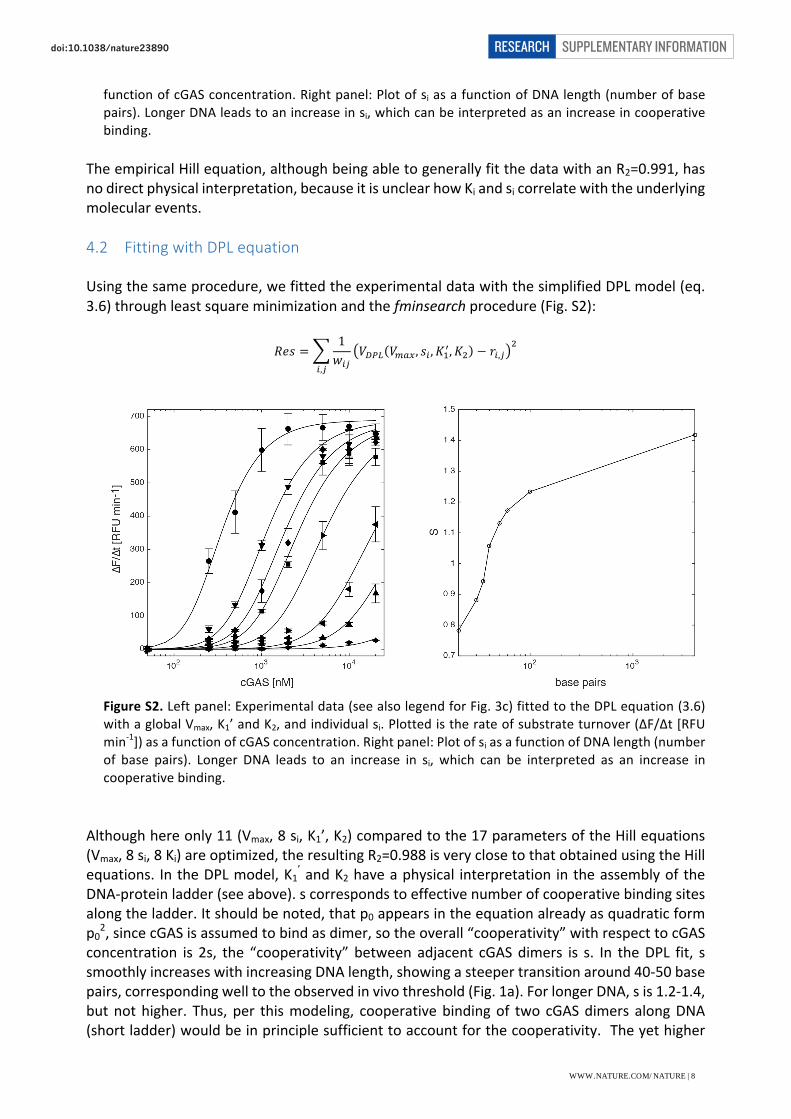
Usingthesameprocedure,wefittedtheexperimentaldatawiththesimplifiedDPLmodel(eq.3.6)throughleastsquareminimizationandthefminsearchprocedure(Fig.S2):
𝑅𝑒𝑠 =1𝑤Bf
𝑉hij 𝑉 [a, 𝑠B, 𝐾8], 𝐾, − 𝑟B,f,
B,f
Althoughhereonly11(Vmax,8si,K1’,K2)comparedtothe17parametersoftheHillequations(Vmax,8si,8Ki)areoptimized,theresultingR2=0.988isveryclosetothatobtainedusingtheHillequations. IntheDPLmodel,K1’andK2haveaphysical interpretation intheassemblyoftheDNA-proteinladder(seeabove).scorrespondstoeffectivenumberofcooperativebindingsitesalongtheladder.Itshouldbenoted,thatp0appearsintheequationalreadyasquadraticformp02,sincecGASisassumedtobindasdimer,sotheoverall“cooperativity”withrespecttocGASconcentration is 2s, the “cooperativity” between adjacent cGASdimers is s. In theDPL fit, ssmoothlyincreaseswithincreasingDNAlength,showingasteepertransitionaround40-50basepairs,correspondingwelltotheobservedinvivothreshold(Fig.1a).ForlongerDNA,sis1.2-1.4,butnothigher. Thus, per thismodeling, cooperativebindingof two cGASdimers alongDNA(shortladder)wouldbeinprinciplesufficienttoaccountforthecooperativity.Theyethigher
functionofcGASconcentration.Rightpanel:PlotofsiasafunctionofDNAlength(numberofbasepairs).LongerDNAleadstoanincreaseinsi,whichcanbeinterpretedasanincreaseincooperativebinding.
FigureS2.Leftpanel:Experimentaldata(seealsolegendforFig.3c)fittedtotheDPLequation(3.6)withaglobalVmax,K1’andK2,andindividualsi.Plottedistherateofsubstrateturnover(∆F/∆t[RFUmin-1])asafunctionofcGASconcentration.Rightpanel:PlotofsiasafunctionofDNAlength(numberof base pairs). Longer DNA leads to an increase in si, which can be interpreted as an increase incooperativebinding.
WWW.NATURE.COM/NATURE | 8
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

activity of very longDNA could originate frommore efficient assembly of cGAS dimer in cisbecauselongDNAcouldeasilybendbackandthuscanhelpassemblecGASdimersatlowDNAconcentrations.4.3 TitrationwithinactivecGASD307N
Inthetitrationexperiment,anincreasingamountofcGAScd(D307N),i.e.amutantthatdoesnotturnoversubstrateGTPandATP,istitratedintoasolutionofafixedamountofcGAScdandafixedamountofDNA.Theladdermodelpredictsahillorbellshapedcurve.IntheabsenceofcGAScd(D307N),thelowamountofcGAScdresultsinlowfGAMPproduction,becausemostifnotallofcGAScdisnotboundtoDNAincatalyticallyactivecGASn:DNA2complexes.TitratingincGAScd(D307N) will cooperatively promote ladder formation and therefore help trap cGAScd incGASn:DNA2 ladders, therefore increasing activity. Increasing amounts of cGAScd (D307N),however, will more and more compete cGAScd away from DNA:protein ladders due to thelimitingnumberofproteinbindingsitesonDNA.Thus,afteramaximumstimulation, furtherincreaseof cGAScd (D307N)will result ina gradual reductionof theobserved rateof fGAMPproduction.Theobservedactivityisasfollows:
𝐴 =𝑝[𝑝klk
∗ 𝑓(𝑝klk)
Here,paistheconcentrationofcatalyticactivecGASandpitheconcentrationofcatalyticinactivecGAS(D307N).ptot=pa+piisthetotalconcentrationofcGASmolecules,bothactiveandinactive.f is theDPL function (3.5 or 3.6) or e.g. an empiricalHill function (4.1). Fittingwas done asdescribedfor4.2usingthefminsearchfunctionasimplementedinMatlab.Forfitting,K1’wassetfixedtothevalueobtainedfromthefitinFig.S2(toreducethenumberoffreeparamteter),allotherparameters(Vmax,K2ands)werekeptfreeandoptimized.Itshouldbenotedthatourpremisethatp0>>s*l0,i.e.p≈p0isnotexactlytruefortheexperimentalconditionsusedinFig.3d, sobothaHillequationand theDPLequationareonlyapproximations.Nevertheless,weobtainagoodfit.Theimportantpointhereisthatagoodfitofthedataleadtos=1.8.Sincetheprotein concentration scales with p2s in the DPL model, 2*1.8=3.6 indicates substantialcooperativityforthestimulationofactivecGASbytitratingininactivecGAS.
WWW.NATURE.COM/NATURE | 9
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

SupplementaryTable1Proteinconstructsandtheirdescriptions
ConstructName ProteinNameSourceOrganism
Fragment,aa
Modification
mcGAScdCyclicGMP-AMPsynthase(cGAS)
Musmusculus 141-507 -
hcGAScdCyclicGMP-AMPsynthase(cGAS)
Homosapiens 155-522 -
hcGASCyclicGMP-AMPsynthase(cGAS)
Homosapiens 1-522N-terminalHis6-MBP-tag
Flag/HA-hcGASCyclicGMP-AMPsynthase(cGAS)
Homosapiens 1-522N-terminalFlag/HA-tag
eGFP-hcGASCyclicGMP-AMPsynthase(cGAS)
Homosapiens 1-522N-terminaleGFP-tag
mTFAMTranscriptionfactorA,mitochondrial(TFAM)
Musmusculus 43-243N-terminalHis6-tag
hTFAMTranscriptionfactorA,mitochondrial(TFAM)
Homosapiens 43-246N-terminalHis6-tag
HA-TFAMTranscriptionfactorA,mitochondrial(TFAM)
Homosapiens 43-246 N-terminalHA-tag
mHMGB1HighmobilitygroupproteinB1(HMGB1)
Musmusculus 1-215N-terminalHis6-tag
mHMGB1dCTTHighmobilitygroupproteinB1(HMGB1)
Musmusculus 1-185N-terminalHis6-tag
lHUDNA-bindingproteinHU
Listeriamonocytogenes
1-121N-terminalHis6-tag
WWW.NATURE.COM/NATURE | 10
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

Supplementary Table 2 Stimulatory DNA sequences Construct Name
Sequence
20 bp-s CTACTAGTGATCTATGACTG 20 bp-as CAGTCATAGATCACTAGTAG 25 bp-s CTACTAGTGATCTATGACTGATCTG 25 bp-as CAGATCAGTCATAGATCACTAGTAG 30 bp-s CTACTAGTGATCTATGACTGATCTGTACAG 30 bp-as CTGTACAGATCAGTCATAGATCACTAGTAG 35 bp-s ATCTACTAGTGATCTATGACTGATCTGTACATGAT 35 bp-as ATCATGTACAGATCAGTCATAGATCACTAGTAGAT 40 bp-s AGTGTCTACTAGTGATCTATGACTGATCTGTACATGATCT 40 bp-as AGATCATGTACAGATCAGTCATAGATCACTAGTAGACACT 45 bp-s TACAGATCTACTAGTGATCTATGACTGATCTGTACATGATCTACA 45 bp-as TGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATCTGTA 50 bp-s GATACAGATCTACTAGTGATCTATGACTGATCTGTACATGATCTACA
ATC 50 bp-as GATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATCTGT
ATC 55 bp-s TCGATACAGATCTACTAGTGATCTATGACTGATCTGTACATGATCTA
CAATCACT 55 bp-as AGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATC
TGTATCGA 60 bp-s AGTCGATACAGATCTACTAGTGATCTATGACTGATCTGTACATGATC
TACAATCACTGCA 60 bp-as TGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTA
GATCTGTATCGACT 65 bp-s CCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTGTACAT
GATCTACAATCACTGCAGT 65 bp-as ACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAG
TAGATCTGTATCGACTTGG 70 bp-s GACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTG
TACATGATCTACAATCACTGCAGT 70 bp-as ACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAG
TAGATCTGTATCGACTTGGTAGTC 75 bp-s GACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTG
TACATGATCTACAATCACTGCAGTTACCG 75 bp-as CGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATC
ACTAGTAGATCTGTATCGACTTGGTAGTC 80 bp-s GACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTG
TACATGATCTACAATCACTGCAGTTACCGTGACC 80 bp-as GGTCACGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCAT
AGATCACTAGTAGATCTGTATCGACTTGGTAGTC 85 bp-s TCCTAGACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTG
ATCTGTACATGATCTACAATCACTGCAGTTACCGTGACC 85 bp-as GGTCACGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCAT
AGATCACTAGTAGATCTGTATCGACTTGGTAGTCTAGGA 90 bp-s TCCTAGACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTG
ATCTGTACATGATCTACAATCACTGCAGTTACCGTGACCAATGT
WWW.NATURE.COM/NATURE | 11
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

90 bp-as ACATTGGTCACGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATCTGTATCGACTTGGTAGTCTAGGA
95 bp-s TCCTAGACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTGTACATGATCTACAATCACTGCAGTTACCGTGACCAATGTCGACT
95 bp-as AGTCGACATTGGTCACGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATCTGTATCGACTTGGTAGTCTAGGA
100 bp-s TCCTAGACTACCAAGTCGATACAGATCTACTAGTGATCTATGACTGATCTGTACATGATCTACAATCACTGCAGTTACCGTGACCAATGTCGACTGGATC
100 bp-as GATCCAGTCGACATTGGTCACGGTAACTGCAGTGATTGTAGATCATGTACAGATCAGTCATAGATCACTAGTAGATCTGTATCGACTTGGTAGTCTAGGA
200 bp-s ATTGTGAGCGGATAACAATTCCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGTCGTACTACCATCACCATCACCATCACGATTACATGATCGAAGAAGGTAAACTGGTAATCTGGATTAACGGCGATAAAGGCTATAACGGTCTCGCTGAAGTCGGTAAGAAATTCGAGAAAGATACCGGAAT
200 bp-as ATTCCGGTATCTTTCTCGAATTTCTTACCGACTTCAGCGAGACCGTTATAGCCTTTATCGCCGTTAATCCAGATTACCAGTTTACCTTCTTCGATCATGTAATCGTGATGGTGATGGTGATGGTAGTACGACATATGTATATCTCCTTCTTAAAGTTAAACAAAATTATTTCTAGAGGGGAATTGTTATCCGCTCACAAT
WWW.NATURE.COM/NATURE | 12
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890

Supplementary Table 3 Data processing and refinement statistics
Space group C2 (No. 5) C2 (No. 5)
Unit cell dimensions a, b,
c, α, β, γ
168.5 Å, 122.9 Å, 180.0 Å,
90º, 96.4º, 90º
168.5 Å, 122.9 Å, 180.0 Å,
90º, 96.4º, 90º
XDS STARANISO
Resolution rangea 50 – 3.6 (4.2) Å 50 – 3.6 (4.2) Å
No. of observed
reflections 212879 –
No. of unique reflections 41980 28400
Completenessb 0.99 0.67 / 0.91
I/σ(I)c 9.4 9.3 / 12.6
Rsym 22.4% (91.0%) –
No. of protein atoms 17712
No. of DNA atoms 4370
No. of zinc ions 6
R-factor / Free-R-factor 0.204 / 0.256
Rmsd bond lengths /
bond angles 0.010 Å / 1.4º
Ramachandran plot
preferred / allowed /
outliers
94.9% / 3.1% / 2.0%
aApproximate effective resolution in parentheses bResulting completeness after STARANISO for spherical / elliptical shells cResulting signal-to-noise ratio after STARANISO for spherical / elliptical shells
WWW.NATURE.COM/NATURE | 13
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature23890