succinate-glycine cycle. i a-carbon atom of glycine to yield a-amino-/3-ketoadipic acid. ......
TRANSCRIPT

THE SUCCINATE-GLYCINE CYCLE*
I. THE MECHANISM OF PYRROLE SYNTHESIS
BY DAVID SHEMIN, CHARLOTTE S. RUSSELL, AND
TESSA ABRAMSKY
(From the Department of Biochemistry, College of Physicians and Surgeons, Columbia University, New York, New York)
(Received for publication, January 10, L955)
We have recently published a series of reactions which appear to be concerned with the metabolism of glycine (1). In this group of reactions, called the succinate-glycine cycle (Fig. l), “active” succinate condenses on the a-carbon atom of glycine to yield a-amino-/3-ketoadipic acid. This /3-keto acid on decarboxylation yields d-aminolevulinic acid, in which the d-carbon atom is originally the a-carbon atom of glycine (1, 2). The aminoketone, which serves as the source for all the carbon and nitrogen atoms of the porphyrin molecule,’ is also metabolized to yield its &carbon atom for the ureido groups of purines (1) and for the p-carbon atom of serine for methyl groups and is converted to formic acid2 (3). This meta- bolic pattern for the b-carbon atom of &aminolevulinic acid is the same as that known for the a-carbon atom of glycine. These findings provide a pathway of glycine metabolism by which these dissimilar compounds are metabolically related. Further, the succinate-glycine cycle provides a pathway by which glycine is converted to carbon dioxide. The original carboxyl group of glycine is converted to carbon dioxide on decarboxylation of a-amino-/3-ketoadipic acid, and the a-carbon atom of the glycine is con- verted to carbon dioxide subsequent to the conversion of the &aminolevu- linio acid to or-ketoglutaric acid2 (3). The a-carboxyl group of the latter compound arises from the a-carbon atom of glycine, and on oxidative de-
* This work was supported by grants from the National Institutes of Health, United States Public Health Service (RG-1128(C6)), from the American Cancer Society on the recommendation of the Committee on Growth of the National Re- search Council, and from the Rockefeller Foundation.
1 The succinate moiety of &aminolevulinic acid is utilized for the same carbon atoms of the protoporphyrin molecule (Schiffman, E., and Shemin, D., unpublished results) as is succinate (4).
z It was found that the methyl group of methionine (Foster, G. L., and Shemin, D., unpublished results) and the p-carbon atom of serine (Gatt, S., and Shemin, D., unpublished results) were radioactive after injection of &aminolevulinic acid-5-W into a duck. The administration by injection of &aminolevulinic acid-504 into a rat resulted in the urinary excretion of highly radioactive formic acid (Gatt, S., and Shemin, D., unpublished results). Also or-ketoglutaric acid contained Cl4 activity after incubation of a duck red blood cell hemolysate with &aminolevulinic acid-5-C’*.
613
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

614 SUCCINATE-GLYCINE CYCLE. I
carboxylation of the ketoglutarate a 2nd mole of carbon dioxide is formed. The succinate-glycine cycle is similar to the citric acid cycle in that they are both concerned with the metabolism of a 2-carbon compound. Oxal- acetate is the substrate catalyst in the citric acid cycle, whereas succinate plays this rale in the reactions in which glycine is concerned.
This communication is concerned with only part of the succinate-glycine cycle; namely, the conversion of &aminolevulinic acid to porphyrin.
We have previously demonstrated the source of each of the carbon atoms of protoporphyrin, 26 being derived from “active” succinate (4, 5) and 8 from the or-carbon atom of glycine (6-8). These two compounds are found in particular positions in the porphyrin; the a-carbon atom of glycine is the source of the methene bridge carbon atom (7, 8) and the 2 position in each of the pyrrole rings (7) (Fig. 2 of the preceding paper (9)). It is also noteworthy that the carboxyl group of glycine is not uti- lized for porphyrin synthesis (6, 10). This finding was an important clue in the elucidation of the mechanism of condensation of the succinate and glycine.
Until recently the mechanism by which succinate and glycine condense to form the pyrrole unit was not known. Any explanation of this con- densation should not only be concerned with pyrrole formation, but should also afford a reasonable biochemical mechanism by which the carboxyl group of glycine is separated from its a-carbon atom. Further, the mech- anism of pyrrole formation should account for the positional distribution of the a-carbon atom of glycine in the porphyrin molecule. These consid- erations must be included in any approach because the a-carbon atom of glycine is equally distributed in the 2 position of the pyrrole rings and in the methene bridges, and because no “G” compound has been found to substitute for glycine (3). It is therefore evident that the whole molecule of glycine is involved in its initial condensation with succinate. If the “active” succinate condensed on the a-carbon atom of glycine to form a new intermediate, cr-amino-/?-ketoadipic acid, all the experimental findings and conclusions would be satisfied (Fig. 1). This compound, being a P-keto acid, can then readily be decarboxylated, and thus a mechanism is provided for the utilization of the whole molecule of glycine in the initial step and a mechanism for subsequent separation of the carboxyl group of glycine from its a-carbon atom. Further, condensation of 2 moles of the resulting compound, b-aminolevulinic acid, by a Knorr type of mechanism would yield a pyrrole in which the a-carbon atom of glycine would be in the 2 position and in the o( substituent (aminomethyl) of the pyrrole, and would thus constitute the source of the methene bridge carbon atoms (Fig. 2). This formulation is supported by our recent finding that the molar radioactivity of porphobilinogen enzymatically synthesized from &aminolevulinic acid-5-Cl4 is twice that of the latter compound (11).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

D. SHEMIN, C. S. RUSSELL, AND T. ABRAMSKY 615
In order to test the above hypothesis, 6-aminolevulinic acid was synthe- sized and its role in the biosynthesis of protoporphyrin was investigated. In the initial experiments, non-radioactive &aminolevulinic acid was added to a duck red blood cell hemolysate along with either U4-labeled succinate or with glycine-2-CY4. If &aminolevulinic acid is formed as an inter-
TRICARBOXYLIC
UREIDO group of purines,
HOOC-CH2-CH2-$-COOH A
d - keto-glutaric :cid PORPHYRIN
FIG. 1. Succinate-glycine cycle; a pathway for the metabolism of glycine
mediate8 in the biosynthesis of porphyrins from succinate and glycine, the addition of unlabeled &aminolevulinic acid should theoretically be reflected in a lowered Cl4 activity of the newly formed heme, since a dilution would occur. It can be seen from Table I, in which a few of the experiments are given, that the Cl4 activities of the isolated hemin samples synthesized from either CY4-labeled succinate or from CY4-labeled glycine in the presence of
8 On incubation of hemolysates with glycine-2-C’* in the presence of unlabeled &aminolevulinic acid, the formaldehyde formed on treating the protein-free super- natant fluid with periodic acid was highly radioactive (see “Experimental”).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

616 SUCCINATE-GLYCINE CYCLE. I
non-radioactive &aminolevulinic acid were much lower than those of the experiments in which the diluent was not added. This effect was not due to an inhibition, since in Experiment 2, Table I, in which the Saminolevulinic acid was labeled with N15, there was a simultaneous excellent incorporation of N16 into the porphyrin. The incorporation into heme of N16 from the &aminolevulinic acid was much greater than that found in the experi- ment in which 6 to 7 times as much N15-labeled glycine was the substrate.
This utilization of Saminolevulinic acid for porphyrin formation was further demonstrated by incubation of the duck red blood cell hemolgsates with b-aminolevulinic alid-5-C14. As can be seen from Table II, not only did the hemin samples contain CY4, but their radioactivities were about
COOH
I COOH CH.2
I I
COOH
I CH2 I
CH2 I
CH2 I
COOH
I CHZ
I y2
~-AMINO LEVULINIC ACID
(El + (n)
FIG. 2. The mechanism for the formation of the monopyrrole, porphobilinogen, by condensation of 2 moles of S-aminolevulinic acid. The carbon atoms bearing the closed circles (0) were originally the a-carbon atom of glycine.
65 times greater than those of samples made from glycine-2-Cl4 having the same Cl4 activity.
Further proof (2) that b-aminolevulinic acid was synthesized by the re- action outlined in Fig. 1 and that it was an intermediate in porphyrin syn- thesis was obtained by degradation of hemin samples synthesized from &aminolevulinic acid-5-C14. The &carbon atom of the latter compound should, according to the hypothesis, metabolically arise from the a-carbon atom of glycine. Thus the Cl4 distribution pattern among the carbon atoms of protoporphyrin synthesized from b-aminolevulinic acid-5-U4 should be exactly the same as that previously found for glycine-2-C4; 50 per cent of the activity should reside in the four pyrrole rings and 50 per cent should reside in the 4 methene bridge carbon atoms (Fig. 3). It can be seen from Table III that the same Cl4 distribution pattern in the porphy- rin was realized for b-aminolevulinic acid-5-C14 and glycine-2-CY4.
The utilization of &aminolevulinic acid for porphyrin formation provides
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

D. SHEMIN, C. S. RUSSELL, AND T. ABRAMSKY 617
the evidence for the condensation of “active” succinate and glycine to form ol-amino-P-ketoadipic acid. The latter compound is therefore an obligatory intermediate in porphyrin formation,4 and its clecarboxylation to Caminolevulinic acid provides the mechanism by which the carboxyl group
TABLE I
Comparison of 04 Activities of Hemin Samples Synthesized from Glycine-2-P (0.06 me. per mmole) or Succinic Acid-2-C14 (0.06 me. per mmole) in Presence
and Absence of Non-Radioactive &Aminolevulinic Acid
Experi merit No.
C”-labeled N”-labeled Unlabeled
Glycine-2-C” (0.05 mmole)
Glycine-2-Cl4 (0.05 mmole)
Glycine-2-P (0.05 mmole)
Glycine-2-CP (0.05 mmole)
Succinate-2-Cl4 (0.1 mmole)
Succinate-2-Cl4 (0.1 mmole)
Substrates
&Aminolevulinic acid (0.05 n-mole) *
Glycine (0.33 mmole) *
&Aminolevulinic acid (0.05 mmole)
&Aminolevulinic acid (0.1 mmole)
Isotope concentra- tion in hemin
C’”
c.g.m.
125
15
230
48
660
180
N’S
atom per ent CXCf3.w
0.21
0.06
* The isotopic concentration of these samples was 34 atom per cent excess Nr6. In each of the experiments the volume of the hemolyzed preparation was 30 ml. Unlabeled succinate (0.1 mmole) was added to the flasks in which labeled glycine was the substrate, and unlabeled glycine (0.33 mmole) was added to the flasks in which labeled succinate was the substrate. Each flask contained 1 mg. of iron (ferric).
of glycine is detached from its a-carbon atom. Subsequent to the initial publication (1) of the conversion of &aminolevulinic acid to protoporphy- rin, Neuberger and Scott (12) and later Dresel and Falk (13) published experiments which were in agreement with this observation.
It is also of interest to note the degree of organization of the cluck red
4 The intraperitoneal injection into a rat of the diethyl ester of a-amino-fi-keto- adipic acid gives rise to porphobilinogen, as does the injection of b-aminolevulinic acid (Weliky, I., and Shemin, D., unpublished observations).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

618 SUCCINATE-GLYCINE CYCLE. I
blood cell preparation necessary for the condensation of the “active” suc- cinate with glycine to form eventually b-aminolevulinic acid. It has been
demonstrated that not only can whole red cells (14) effect the synthesis of porphyrin from these two substrates, but also that the synthesis occurs in a gently hemolyzed preparation (4, 15, 16). However, more drastic dis- ruption of the preparation, namely homogenization in a Waring blendor,
TABLE II
Comparison of Cl4 Activities of Hemin Samples Synthesized from Glycine-904 (0.06 me. per mmole) and &Aminolevulinic Acid (0.06 me. per mmole)
Glycine-2-W (0.05 mmole) &Aminolevulinic acid (0.05 mmole)
I‘ “ (0.05 “ ) *
Other additions Cl” activity
of hemin
c.p.m.
Succinate (0.1 mmole) 333 23,000
Glycine (0.33 mmole) + suc- 21,000 cinate (0.1 mmole)
N H2- ;H2-COOH
Glycine-2-Cl4
NH2 -:H, -CO-CH2 - CH2-COOH
CH3 y2 7H2 CH3 6-Aminolevulinic Acid-5-Ci4
CH2
iOOH
CH2
!OOH
PROTOPORPHYRIN IX
FIG. 3. The carbon atoms of protoporphyrin which arise from the a-carbon atom of glycine and from the B-carbon atom of &aminolevulinic acid.
causes the system to lose most if not all of its synthetic capacity for heme synthesis when succinate or ketoglutarate and glycine are the substrates (Table IV). On the other hand, we have found that the homogenate can synthesize heme when &aminolevulinic acid is the substrate (2). Further, even a particle-free extract obtained by high speed centrifugation, and a lyophilized powder of the extract, can synthesize the porphyrin from Saminolevulinic acid (Table IV). It appears, therefore, that a solution of enzymes can synthesize the porphyrin from d-aminolevulinic acid and that on homogenization the functional activity of only those enzymes concerned
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from
D. SHEMIN, C. S. RUSSELL, AND T. ABRAMSKY 619
in the condensation of “active” succinate with glycine is lost. Since the conversion of &aminolevulinic acid (2) and porphobilinogen (11, 17, 18) to porphyrins is now known to occur in a soluble enzyme system, the pos- sibility has been opened up of isolating the enzymes concerned with the individual steps and thus studying the reactions in greater detail (11). Indeed, it has been found subsequently that a highly purified protein frac- tion obtained from ox liver (19) and from duck erythrocytes (11) can catalyze the conversion of &aminolevulinic acid to porphobilinogen.
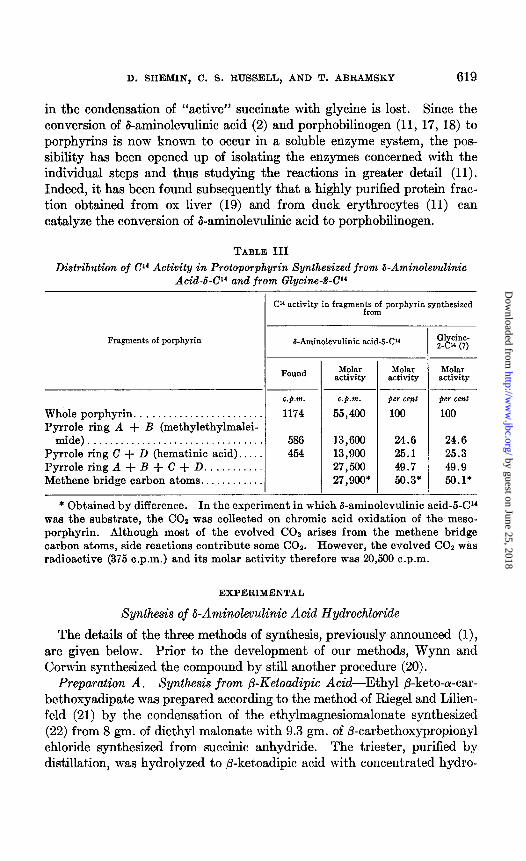
TABLE III
Distribution of 04 Activity in Protoporphyrin Synthesized from Q-Aminolevulinic Acid-b-Cl4 and from Glycine-P-Cl4
Cl’ activity in fragmer$;;f porphyrin synthesized
Fragments of porphyrin I-Aminolevulinic acid-W3 Glycine- 2-C” (7)
Found M&r Molar activity activity
~___
Whole porphyrin. . . . . . . . . . . . . . Pyrrole ring A + B (methylethylmalei-
mide)................................ Pyrrole ring C + D (hematinic acid). . PyrroleringA+B+C+D . . . . . . . . . . Methene bridge carbon atoms.. . . . . . .
c.p.m. 1174
586 454
13,600 24.6 24.6 13,900 25.1 25.3 27,560 49.7 49.9 27,900* 56.3* 56.1*
per cent 100
* Obtained by difference. In the experiment in which &aminolevulinic acid-5-04 was the substrate, the CO2 was collected on chromic acid oxidation of the meso- porphyrin. Although most of the evolved CO2 arises from the methene bridge carbon atoms, side reactions contribute some COZ. However, the evolved COZ was radioactive (375 c.p.m.) and its molar activity therefore was 20,560 c.p.m.
EXPERIMENTAL
Synthesis of b-Aminolevulinic Acid Hydrochloride
The details of the three methods of synthesis, previously announced (l), are given below. Prior to the development of our methods, Wynn and Corwin synthesized the compound by still another procedure (20).
Preparation A. Synthesis from /3-Ketoadipic Acid-Ethyl @-keto-a-car- bethoxyadipate was prepared according to the method ,of Riegel and Lilien- feld (21) by the condensation of the ethylmagnesiomalonate synthesized (22) from 8 gm. of diethyl malonate with 9.3 gm. of /?-carbethoxypropionyl chloride synthesized from succinic anhydride. The triester, purified by distillation, was hydrolyzed to fi-ketoadipic acid with concentrated hydro-
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

620 SUCCINATE-GLYCINE CYCLE. I
chloric acid for 18 hours at room temperature, according to the method of Eisner, Elvidge, and Linstead (23). The crude crystalline ,&ketoadipic acid was suspended in 50 ml. of glacial acetic acid, and an equimolar amount of ethyl nitrite (24) was passed at 15-20” into the suspension over a period of about 20 minutes. After standing at room temperature for 1 hour, dur- ing which time all the crystalline material dissolved, the solution was taken to dryness in vacua at 30-35” and the oily residue added dropwise to an ice-
TABLE IV
Comparison of Cl4 Activities of Hemin Synthesized from &Aminolevulinic Acid-6-04 (0.06 mc. per mmole), from G12/cine-%Cr4 (0.06 mc. per mmole), and from Succinic Acid-d-C’4 (0.06 mc. per mmole) in Di$erent Duck Red Blood Cell Preparations*
Experi- ment No. ‘2’4 substrate Red cell preparation
Glycine (0.05 mmole)t Hemolyzed ‘I (0.05 “ It Homogenized
Succinate (0.05 mmole)$ Hemolyzed ‘I (0.05 “ I$ Homogenized
&Aminolevulinic acid (0.006 mmole) Hemolyzed ‘I “ (0.006 I‘ “ (0.00!3
1: ) Homogenized
“ (‘ (0.009 “ i Superiatant (12 X 1Oa X g) ‘I I‘ “ “
‘I (0.009 “ ) I‘ (47 x 103 x “) “ (0.009 “ ) “ (100 x 103 x “: “ (0.009 “ ) “ (12 x 108 x “1 “ (0.009 “ ) Lyophilized preparation
(
-
,
I
I
I
74 activ- ity in hemin sample
395 2
565 1
3900 4500 2200 1600 1600 1500 2000 1500
* Each preparation represented 25 ml. of duck blood prepared as previously de- scribed (4, 14) and contained 1 mg. of iron (ferric).
t Plus 0.1 mmole of non-radioactive cu-ketoglutaric acid. 2 Plus 0.33 mmole of non-radioactive glycine.
cold solution of 25 gm. of SnC12.2HzO in 25 ml. of concentrated hydro- chloric acid. The resulting solution, after standing for 48 hours in the refrigerator, was diluted to 500 ml. with water and treated with hydrogen sulfide. The tin sulfide was filtered off and washed with water. The filtrate and washings were taken to dryness in vacua, and the residue was dried in a desiccator over NaOH. The crystalline residue was dissolved in a minimum of ethanol and recrystallized by the addition of ethyl acetate. The white crystalline material was further purified from alcohol-ethyl acetate; yield 2.0 to 2.2 gm.
The 6-aminolevulinic acid hydrochloride, m.p. 149-151”, gave a positive ninhydrin test, reduced Benedict’s solution in the cold, and gave one spot
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

D. SFIEMIN, C. S. RUSSELL, AND T. ABRAMSKY 621
on an ascending paper chromatogram with an Rp of about 0.42 to 0.44 in a phenol-water mixture.
CsH90aN-HCl. Calculated. C 35.8, H 6.0, N 8.4, Cl 21.2 Found. “ 35.6, “ 6.7, “ 8.3, I‘ 21.0
For the synthesis of &aminolevulinic acid-5-CY4, diethyl malonate-2-Cl4 was used, and for the synthesis of N15-labeled &aminolevulinic acid, ethyl nitrite prepared from KN1502 was used to form the isonitroso derivative. The KN1602 was prepared from KN1603.
Preparation B. By Gabriel Synthesis-To an ethereal solution of 1 gm. of /3-carbomethoxypropionyl chloride was added an ethereal solution of diazomethane obtained from 2 gm. of N-nitroso-N-methylurea, and the mixture was stirred for 2 hours at room temperature. After this time either dry HCl gas was bubbled through the solution or an ethereal solution of HCl was added dropwise until the evolution of Nz ceased. The solvent was removed by passing a dry stream of nitrogen through the solution, and the residue was dried in vacua over NaOH. The oily residue was treated with 4 ml. of dimethylformamide and 1.24 gm. of potassium phthalimide (labeled with 34 atom per cent excess N16). The resulting orange sus- pension was stirred vigorously for 1 hour at room temperature and then dissolved in 25 ml. of chloroform. The chloroform solution was washed with 50 ml. of water and the washings shaken with 2 ml. portions of chloro- form. The combined chloroform solutions were then washed twice with 50 ml. portions of water, once with 25 ml. of 0.2 N sodium hydroxide, and finally four times with water. The chloroform solution was dried by filtration and taken to dryness in vacua. The crude methyl i-phthalimi- dolevulinate was purified by crystallization from methanol, m.p. 96-97”. The phthalimido derivative was then hydrolyzed by boiling for 7 to 10 hours with 6 N hydrochloric acid. After removal of the phthalic acid, the crude &aminolevulinic acid hydrochloride was purified as in Preparation A. Neuberger and Scott recently independently synthesized this aminoketone by a similar procedure (25).
The material of this synthesis had the same characteristic properties as those described in the above preparation, and its melting point was not depressed when mixed with a sample obtained in Preparation A.
Preparation C. From Urocanic Acid6-2 gm. of urocanic acid in aqueous ethanol were reduced to imidazolepropionic acid with palladium and hy- drogen. The acid was converted to its methyl ester with dry methanolic hydrochloric acid and then exhaustively benzoylated by the method of
5 We wish to thank Dr. H. Tabor of the National Institutes of Health, Bethesda, for his generous gift of urocanic acid.
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

622 SUCCINATE-GLYCINE CYCLE. I
Ashley and Harington (26) as modified by Tesar and Rittenberg (27). The dibenzoyl derivative obtained from the benzene layer was purified by recrystallization from methanol-water. 800 mg. of material, m.p. 136- 142’, were obtained.
500 mg. were hydrolyzed by boiling with 20 ml. of dry 10 per cent meth- anolic HCl for 1.5 hours. The residue obtained after distillation in vucuo was then refluxed with 20 ml. of 6 N hydrochloric acid for another 1.5 hours. The crystals which were formed on cooling were removed by filtration, and the filtrate was taken to dryness and purified by crystallization from al- cohol-ethyl acetate solution. 60 mg. of &aminolevulinic acid hydrochlo- ride were obtained, m.p. 149-151”, having the same RF as samples prepared by the foregoing methods and the same chemical characteristics.
No depression in melting point was observed when the compound was mixed with a sample from Preparation A.
Periodate Treatment-In order to secure further support of the structure assigned to the compounds obtained from the three independent syntheses, samples of &aminolevulinic acid were treated with periodate. Above pH 7, 1 mole of periodate was taken up, and from the reaction products a 95 per cent yield of formaldehyde, isolated as the dimedon derivative, m.p. 192”, and a 90 per cent yield of succinic acid, m.p. 190-191”, were obtained. Mixed melting points with authentic samples gave no depression in the melting point.
Hemolyzed Duck Red Blood Cells-These preparations were made as de- scribed previously, and the same additions were made (4).
Preparation of Extract-The cold hemolyzed preparations were homog- enized in a cold Waring blendor for 10 minutes, and the mixture was then centrifuged for 30 minutes to 4 hours in a Spinco preparative centrifuge at gravitational forces indicated in Table IV. This extract was used, without further additions, in the experiments.
Incubation Experiments-The hemolyzed preparations and the extracts were shaken at room temperature, usually for 16 hours. The additions and radioactivities of the substrates are specified in Tables I to IV.
Degradation of Labeled Protoporphyrin-The radioactive hemin isolated was degraded to methylethylmaleimide and hematinic acid by procedures previously described (5, 7).
Measurement of Radioactivity-The methods employed were published previously, and the use of the term “molar activity” is as defined earlier
(4, 5).
DISCUSSION
The results obtained in this study of the conversion of d-aminolevulinic acid into protoporphyrin are in complete agreement with the conclusion
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

D. SHEMIN, C. S. RUSSELL, AND T. ABRAMSKY 623
that “active” succinate and glycine condense to form a-amino-/3-ketoadipic acid, and the latter keto acid then decarboxylates to form d-aminolevulinic acid. The distribution of the &carbon atom of this aminoketone among the carbon atoms of the porphyrin supports the suggested mechanism for the formation of the monopyrrole. The mechanism pictured is the con- densation of 2 moles of b-aminolevulinic acid by a Knorr type of condensa- tion to form the precursor pyrrole (Fig. 2) ; this has subsequently been sup- ported by the finding that the molar radioactivity of porphobilinogen enzymatically synthesized from d-aminolevulinic acid-5-Cl4 is twice that of the latter compound (11). It is of interest that the structure of the pre- cursor pyrrole arrived at from these theoretical considerations is the same as that determined for porphobilinogen (28, 29).
All the previous data were consistent with the suggestion that a pre- cursor pyrrole is first formed and subsequently utilized for synthesis of the tetrapyrrole compound. The previous considerations and the findings in this communication, coupled with those dealing with the structure of porphobilinogen, and the experiments demonstrating the formation of porphobilinogen from 6-aminolevulinic acid and its conversion to porphyrins in biological systems (11, 12, 17-19) adequately demonstrate that the mono- pyrrole porphobilinogen is indeed an intermediate in porphyrin synthesis.
Although no evidence has yet been obtained concerning the biological mechanism of conversion of the monopyrrole to the tetrapyrrole structure, several suggestions have been advanced (17, 30). We should like to sug- gest still another possibility which may explain the distribution of the a-carbon atom of glycine or the a-carbon atom of d-aminolevulinic acid in the porphyrin molecule of the I and III series. It is based on the synthetic mechanism of dipyrrole and tetrapyrrole formation demonstrated by Cor- win et al. (31,32) and is similar to that postulated by Bogorad and Granick
(17). Condensation of 3 moles of the precursor pyrrole (porphobilinogen), or
of a closely related derivative, by a mechanism of Corwin et al. (31, 32) would lead to a tripyrrylmethane compound, as schematically represented in Fig. 4. The tripyrrylmethane then breaks down into a dipyrrylmethane and a monopyrrole. The structure of the dipyrrylmethane is dependent on the place of splitting. An A split would give rise to dipyrrylmethane A, and a B split would give rise to dipyrrylmethane B. Condensation of 2 moles of dipyrrylmethane A would give rise to a porphyrin of the I series, while condensation of a mole of A and a mole of B would give rise to a por- phyrin of the III series. In the formation of the porphyrin of the III series it can be seen from Fig. 4 that it is necessary to lose a l-carbon com- pound, since there are three aminomethyl side chains and only two are required to condense the two dipyrroles to the porphyrin structure. If the
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

624 SUCCINATE-GLYCINE CYCLE. I
mechanism similar to that outlined in Fig. 4 is concerned with porphyrin synthesis, it would appear that this l-carbon compound given off could well be formaldehyde. Consistent with this idea is our finding6 that on the conversion of porphobilinogen to porphyrins, either by heating under acid conditions (27) or by enzymatic conversion in cell-free extracts (2), formaldehyde was formed. This was established by heating or incubating porphobilinogen labeled with Cl4 in the aminomethyl group (11) and isolat- ing radioactive formaldehyde as the dimedon derivative. It is difficult at present to speculate as to a uniform metihanism concerning the format,ion of the tetrapyrrole structures, but we should like to suggest that in theinter- mediate stages in the formation of protoporphyrin at least some of the bridge atoms are methylene groups and that it is in such intermediates that
FIG. 4. A mechanism of porphyrin formation from the monopyrrole. AC = acetic acid side chain; P = propionic acid side chain; l = a-carbon atom of gly- tine and &carbon atom of S-aminolevulinic acid.
decarboxylation occurs. This is consistent with our finding that copro- porphyrin III is not converted to protoporphyrin in a duck red blood hemolysate or extract.’
SUMMARY
The biological mechanism concerned with the utilization of ‘(active” succinate and glycine for porphyrin formation was investigated. It was concluded that the succinate condenses on the a-carbon atom of glycine to form ar-amino-fi-ketoadipic acid. This keto acid undergoes decarboxyla- tion to yield b-aminolevulinic acid. Condensation of 2 moles of the amino- ketone forms the precursor monopyrrole. This series of reactions provides
B Shemin, D., and Schmid, R., unpublished results. 7 We have synthesized C14-labeled coproporphyrin III, and in all our experiments
we have been unable to demonstrate its conversion to protoporphyrin (Russell, C. S., and Shemin, D., unpublished results).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

D. SHEMIN, C. S. RUSSELL, AND T. ABRAMSKY 625
the mechanism by which the carboxyl group of glycine is detached from its a-carbon atom and provides the mechanism for pyrrole formation. These conclusions were reached from studies in which the utilization of b-amino- levulinic acid for porphyrin formation was investigated. It was found that unlabeled d-aminolevulinic acid markedly lowered the Cl4 activity of protoporphyrin synthesized by duck red blood cell hemolysates from either CY4-labeled succinate or U4-labeled glycine. Also &aminolevulinic acid-s- Cl4 gave rise to labeled protoporphyrin whose Cl4 distribution pattern was the same as that found in protoporphyrin synthesized from glycine-2-CY4. This is in agreement with the postulated reactions, since the S-carbon atom of d-aminolevulinic acid should arise from the a-carbon atom of glycine.
It was also found that, whereas a homogenate of duck red blood cells is incapable of forming porphyrin from succinate and glycine, it can synthe- size protoporphyrin from &aminolevulinic acid. Further, a particle-free extract of duck red blood cells can synthesize porphyrin from S-aminolevu- linic acid, even after lyophilization. It would appear that those enzymes which can convert &aminolevulinic acid to protoporphyrin are soluble and that homogenization destroys the functional activity of only those enzymes concerned with the condensation of “active” succinate and glycine.
The formation of porphyrin is discussed, and consistent with the theory is the finding that on conversion of porphobilinogen to porphyrin formal- dehyde is formed from the aminomethyl group of the porphobilinogen.
BIBLIOGRAPHY
1. Shemin, D., and Russell, C. S., J. Am. Chem. Sot., 76, 4873 (1953). 2. Shemin, D., Abramsky, T., and Russell, C. S., J. Am. Chem. SOL, 76, 1204 (1954). 3. Shemin, D., inMcElroy, W. D., and Glass, B., Amino acidmetabolism, Baltimore,
4, 727 (1954). 4. Shemin, D., and Kumin, S., J. Biol. Chem., 198, 827 (1952). 5. Shemin, D., and Wittenberg, J., J. Biol. Chem., 192, 315 (1951). 6. Radin, N. S., Rittenberg, D., and Shemin, D., J. Biol. Chem., 184, 745 (1950). 7. Wittenberg, J., and Shemin, D., J. Biol. Chem., 186, 103 (1950). 8. Muir, H. M., and Neuberger, A., Biochem. J., 47, 97 (1950). 9. Wriston, J. C., Jr., Lack, L., and Shemin, D., J. Biol. Chem., 216, 603 (1955).
10. Grinstein, M., Kamen, M. D., and Moore, C. V., J. BioZ. Chem., 174, 767 (1948). 11. Schmid, R., and Shemin, D., J. Am. Chem. SOL, 77, 506 (1955). 12. Neuberger, A., and Scott, J. J., Nature, 173, 1093 (1953). 13. Dresel, E. I. B., and Falk, J. E., Nature, 172, 1185 (1953). 14. Shemin, D., London, I. M., and Rittenberg, D., J. BioZ. Chem., 183, 757 (1950). 15. London, I. M., and Yamasaki, M., Federation Proc., 11, 250 (1952). 16. Dresel, E. I. B., and Falk, J. E., Biochem. J., 66, 156 (1954). 17. Bogorad, L., and Granick, S., Proc. Nat. Acad. SC., 39, 1176 (1953). 18. Falk, J. E., Dresel, E. I. B., and Rimington, C., Nature, 172, 292 (1953). 19. Gibson, K. D., Neuberger, A., and Scott, J. J., Biochem. J., 68, p. xii (1954). 20. Wynn, R. W., and Corwin, A. H., J. Org. Chem., 16, 203 (1950).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

626 SUCCINATE-GLYCINE CYCLE. I
21. Riegel, B., and Lilienfeld, W. M., J. Am. Chem. Sot., 67, 1273 (1945). 22. Lund, H., and Voigt, A., Org. Syntheses, ~011.2, 594 (1943). 23. Eianer, U., Elvidge, J. A., and Linstead, R. P., J. Chem. Sot., 2223 (1950). 24. Semon, W. L., and Damerell, V. R., Org. Syntheses, ~011.2, 204 (1943). 25. Neuberger, A., and Scott, J. J., J. Chem. Sot., 1820 (1954). 26. Ashley, J. N., and Harington, C. R., J. Chem. Sot., 2586 (1930). 27. Tesar, C., and Rittenberg, D., J. Biol. Chem., 170, 35 (1947). 28. Westall, R. G., Nature, 170, 614 (1952). 29. Cookson, G. H., and Rimington, C., Nature, 171,875 (1953). 30. Cookson, G. H., and Rimington, C., Biochem. J., 67,476 (1954). 31. Corwin, A. H., and Andrews, J. S., J. Am. Chem. Sot., 69,1973 (1937). 32. Andrews, J. S., Corwin, A. H., and Sharp, A. G., J. Am. Chem. Sot., 73,491 (1950).
by guest on June 25, 2018http://w
ww
.jbc.org/D
ownloaded from

AbramskyDavid Shemin, Charlotte S. Russell and Tessa
SYNTHESISTHE MECHANISM OF PYRROLE
THE SUCCINATE-GLYCINE CYCLE: I.
1955, 215:613-626.J. Biol. Chem.
http://www.jbc.org/content/215/2/613.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/215/2/613.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on June 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from