supplementary figure 1 b 32 p-par-4 75 ck2 + rat gst-par-4 ck2 ck2 - + + - ck2 rat gst-par-4 35...
TRANSCRIPT
Supplementary Figure 1Supplementary Figure 1
B
32P-Par-4 75
CK2 + Rat GST-Par-4
CK2CK2
-+
+-
CK2Rat GST-Par-4
35S-CK2
35S-CK2
GST
A IP:GST
32P-CK2 Autophosphorylation37
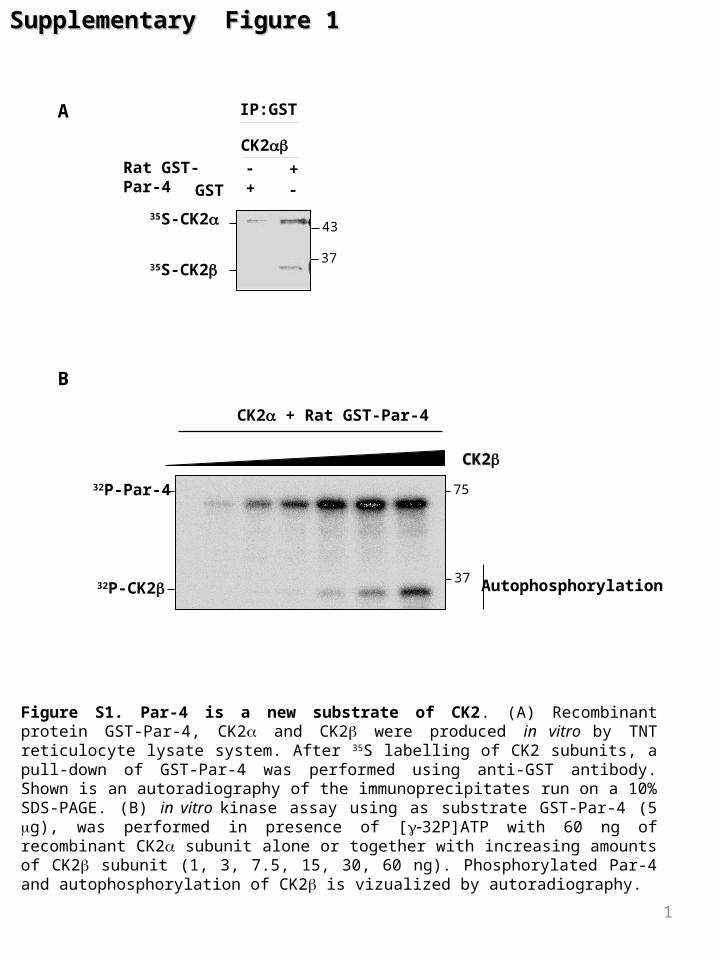
Figure S1. Par-4 is a new substrate of CK2. (A) Recombinant protein GST-Par-4, CK2 and CK2 were produced in vitro by TNT reticulocyte lysate system. After 35S labelling of CK2 subunits, a pull-down of GST-Par-4 was performed using anti-GST antibody. Shown is an autoradiography of the immunoprecipitates run on a 10% SDS-PAGE. (B) in vitro kinase assay using as substrate GST-Par-4 (5 g), was performed in presence of [32P]ATP with 60 ng of recombinant CK2 subunit alone or together with increasing amounts of CK2 subunit (1, 3, 7.5, 15, 30, 60 ng). Phosphorylated Par-4 and autophosphorylation of CK2 is vizualized by autoradiography.
1
43
37
- + 200 100 µgRecPP1
Par-4Ph-Par-4
CalA
Supplementary Figure 2Supplementary Figure 2
Ph-Par-4
OA
Par-4
CalA
A
B
CIP
:Par
-4
WB: Par-4
WB:PP1
IP Ig
GInputs
37
Par-4 Par-4
CalA 0 30’ 60’ 90’0 30’ 60’ 90’
Hsp90
Ph-Par-4 Ph-Par-4
90
D
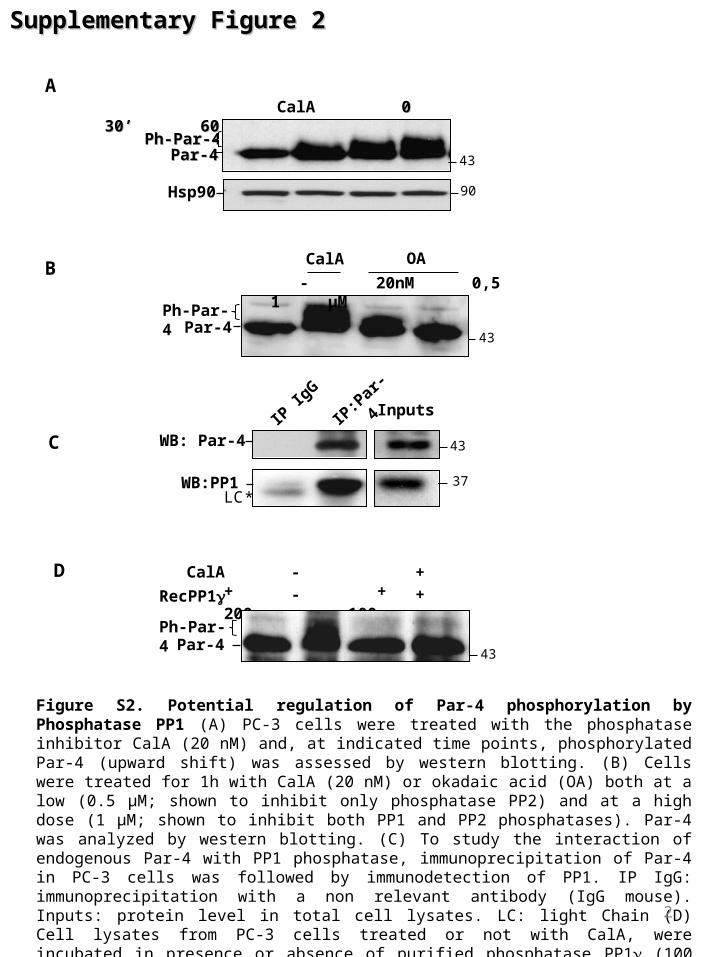
Figure S2. Potential regulation of Par-4 phosphorylation by Phosphatase PP1 (A) PC-3 cells were treated with the phosphatase inhibitor CalA (20 nM) and, at indicated time points, phosphorylated Par-4 (upward shift) was assessed by western blotting. (B) Cells were treated for 1h with CalA (20 nM) or okadaic acid (OA) both at a low (0.5 µM; shown to inhibit only phosphatase PP2) and at a high dose (1 µM; shown to inhibit both PP1 and PP2 phosphatases). Par-4 was analyzed by western blotting. (C) To study the interaction of endogenous Par-4 with PP1 phosphatase, immunoprecipitation of Par-4 in PC-3 cells was followed by immunodetection of PP1. IP IgG: immunoprecipitation with a non relevant antibody (IgG mouse). Inputs: protein level in total cell lysates. LC: light Chain (D) Cell lysates from PC-3 cells treated or not with CalA, were incubated in presence or absence of purified phosphatase PP1 (100 or 200µg) 1,2, 45 min at 30°C. Par-4 dephosphorylation was evaluated by immunoblotting.
- + + +
- 20nM 0,5 1 µM
*LC
2
43
43
43
43
Supplementary Figure 3Supplementary Figure 3
B
A
Cycle N° 1 2 3 4 5 6 7 88 9 10
Disc
P1
CK2rec
P2
117GDEEEPDpSAPEK128[MH]+ = 1382,53
117GDEEEPDpSAPEKGR130[MH]+ = 1595,64
220STIpSAPEEEILNRYPR235[MH]+ = 1954,95
P1 P1’ P2
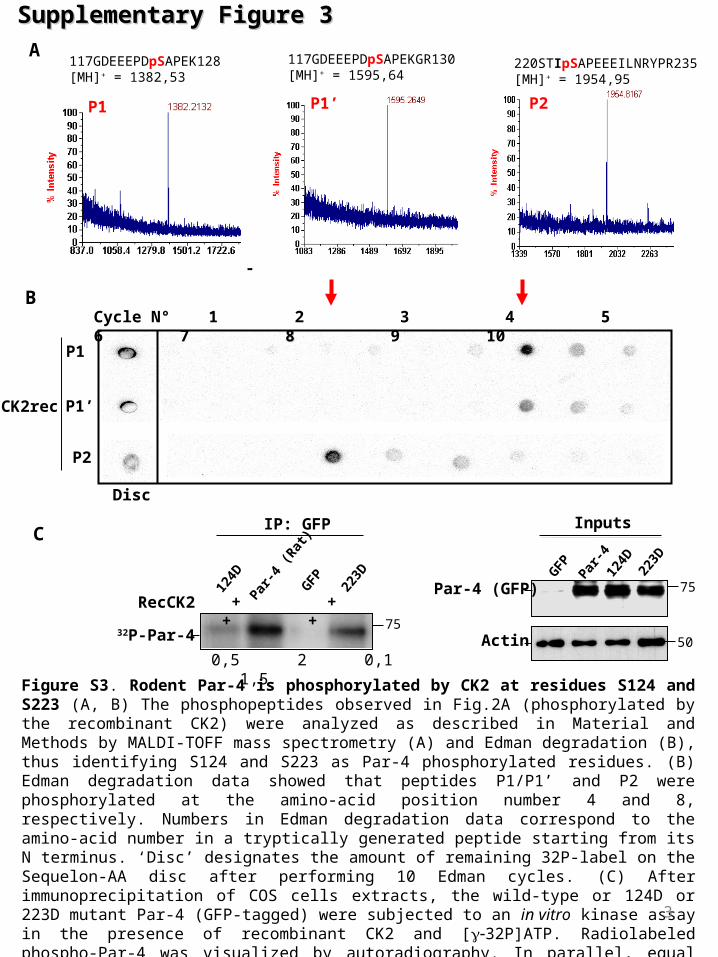
Figure S3. Rodent Par-4 is phosphorylated by CK2 at residues S124 and S223 (A, B) The phosphopeptides observed in Fig.2A (phosphorylated by the recombinant CK2) were analyzed as described in Material and Methods by MALDI-TOFF mass spectrometry (A) and Edman degradation (B), thus identifying S124 and S223 as Par-4 phosphorylated residues. (B) Edman degradation data showed that peptides P1/P1’ and P2 were phosphorylated at the amino-acid position number 4 and 8, respectively. Numbers in Edman degradation data correspond to the amino-acid number in a tryptically generated peptide starting from its N terminus. ‘Disc’ designates the amount of remaining 32P-label on the Sequelon-AA disc after performing 10 Edman cycles. (C) After immunoprecipitation of COS cells extracts, the wild-type or 124D or 223D mutant Par-4 (GFP-tagged) were subjected to an in vitro kinase assay in the presence of recombinant CK2 and [32P]ATP. Radiolabeled phospho-Par-4 was visualized by autoradiography. In parallel, equal expression levels of GFP-Par-4 and GFP-124D, 223D were assessed (aliquot of the immunoprecipitates) by western blot (Inputs, right panel). Actin is the loading control.
124D
Par-4
(Rat
)
223D
GFP
32P-Par-475
RecCK2
IP: GFP
Actin
Inputs
75
50
Par-4 (GFP)12
4D
Par-4
223D
GFP
C
-
+ + + +
0,5 2 0,1 1,5
P1’
3

Supplementary Figure 4Supplementary Figure 4
Figure S4. Effect of Par-4 mutants on NF-B activity Luciferase reporter assay using an NF-B luciferase promoter was performed on COS cells co-transfected either with an empty vector (Mock), wild-type Par-4, 124A223A or 124D223D Par-4 mutants. Prior analysis, cells were treated or not by TPA (50 nM) for 1h to stimulate NF-B activity3 . Inputs: protein level in total cell lysates. Bars represent the mean ±S.D. of at least six independent experiments. Mock, cell transfected with an empty vector. * p<0,05
*
Lu
cif
era
se
in
ten
sit
y (
a.u
)
Par-4 (rat)
Mock
124A223A124D223D
Par-4 (GFP)
Hsp90Inputs
CTLNF-B promLuc +TPA
4

Supplementary Figure 5Supplementary Figure 5
Species P-Site -4 -3 -2 -1 0 1 2 3 4
Human G132 E E E P D G V P E
Bonobo G132 E E E P D G L P E
Macaque G132 E E E Q D G V P E
Mouse S125 E E E P D S A R E
Rat S124 E E E P D S A P E
Taurus C216 E E E R D C A P E
Zebrafish S117 E S P T H S K S K
Caspase cleavage site
CK2 site
A
B
Figure S5. Par-4 Sequence conservation (A) Sequence alignment of Human Par-4 vs. Rat Par-4 (B) Alignment between species of the sequence containing the conserved caspase targeted site. In red, the caspase cleavage site and in blue, the CK2 site conserved specifically in rodent (S124 in rat). 5

Supplementary Figure 6Supplementary Figure 6
A
Par-4 (GFP)
D123A
Cleaved Par-4
Par-4 (R
at)
+ - +TRAIL75
50
Hsp90 90
hPar-4 (Myc)
D131A
Cleaved Par-4
- + - +TRAIL
34
Hsp90 90
hPar-4
Moc
kPar
-4 (r
at)
D123A
TRAIL
PARP
Cleaved PARP
B
95
% A
po
pto
sis
*
Moc
kPar
-4 (r
at)
D123A
Hsp90
28
90
Par-4 (GFP)
GFPG
FP
Par-4
(Rat
)
D123A
GFP
Par-4
(Rat
)
D123A
TRAIL
Casp-8
Cleaved Casp-8
p41/43
p18
D
Rec.CK2 + - + Rec.Casp3 - + +
hPar-4 (GFP)
Cleaved Par-4
C
75
*
6
43
55
75
26
50
26

Supplementary Figure 6Supplementary Figure 6
Figure S6. Caspases mediated cleavage of rodent Par-4, at residue D123, is required for its full proapoptotic activities. (A) HCT116 cells transfected either with the GFP-tagged constructs rat Par-4 (wild type, D123A mutant, left panel) or with the Myc-tagged constructs human Par-4 (wild type, D131A mutant, right panel) were treated with TRAIL (150 ng/ml, 3h) and Par-4 cleavage was evaluated by Western-blotting using GFP or Myc antibody. (B) As already shown for the human D131A mutant of Par-424, the caspase resistant D123A mutant of rat Par-4 impaired apoptosis observed by PARP (left panel)/caspase-8 (right panel) cleavage (Western-blot). ) Hsp90 was used as a loading control. (C) Caspase assay using recombinant caspase-3 (Rec.Casp3) was performed on pre-phosphorylated human GFP-Par-4 (in vitro kinase assay using recombinant CK2 (Rec.CK2) in the presence of ATP as described in Fig.2) and cleaved Par-4 was detected by immunoblotting using GFP antibody. *non-specific band
7
GFP
PARP
Cleaved Casp8
A
B
% A
po
pto
sis
Mock Par-4 124A 124D 223A 223D
TRAIL
Cleaved PARP
Supplementary Supplementary Figure 7Figure 7
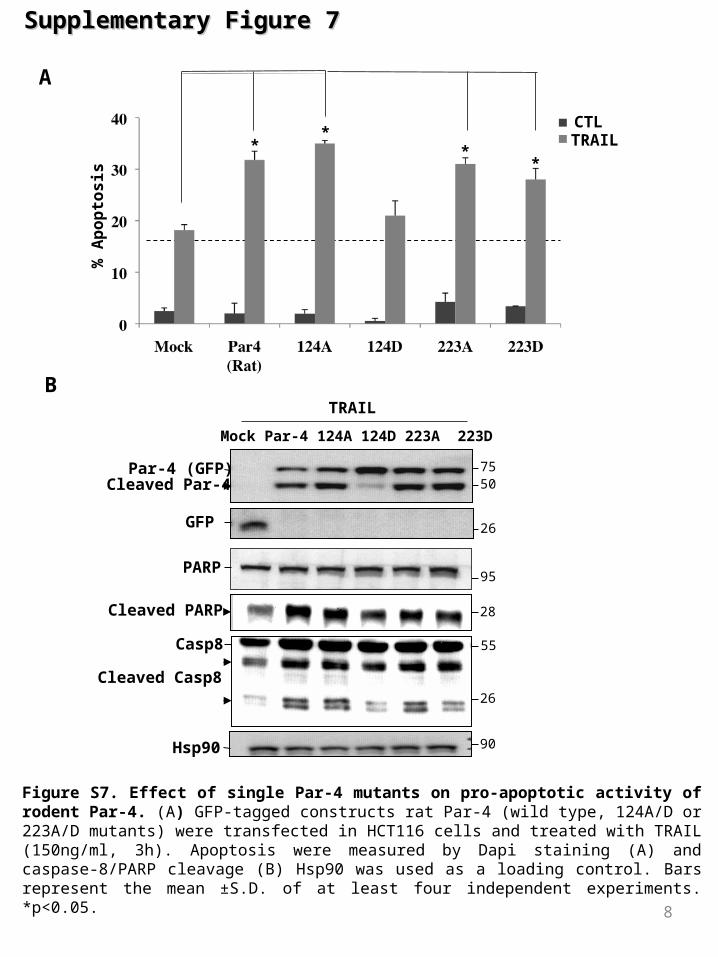
Figure S7. Effect of single Par-4 mutants on pro-apoptotic activity of rodent Par-4. (A) GFP-tagged constructs rat Par-4 (wild type, 124A/D or 223A/D mutants) were transfected in HCT116 cells and treated with TRAIL (150ng/ml, 3h). Apoptosis were measured by Dapi staining (A) and caspase-8/PARP cleavage (B) Hsp90 was used as a loading control. Bars represent the mean ±S.D. of at least four independent experiments. *p<0.05.
CTLTRAIL
Casp8
Hsp90
Par-4 (GFP)
**
**
Cleaved Par-4
8
26
90
26
28
95
7550
55
Mock
Par-4 (124-332)
Par-4 (Rat)
= 18% apoptosis
= 40% apoptosis
= 22% apoptosis
24h
Par-4
(124
-332
)
Par-4
(124
-332
)
Cell lysate
ConditionedMedia
Par-4
(Rat
)
Mock
Par-4
(Rat
)
Mock
75
26
Par-4 (GFP)
GFP
Par-4(124-332)
Hsp110 100
A
B
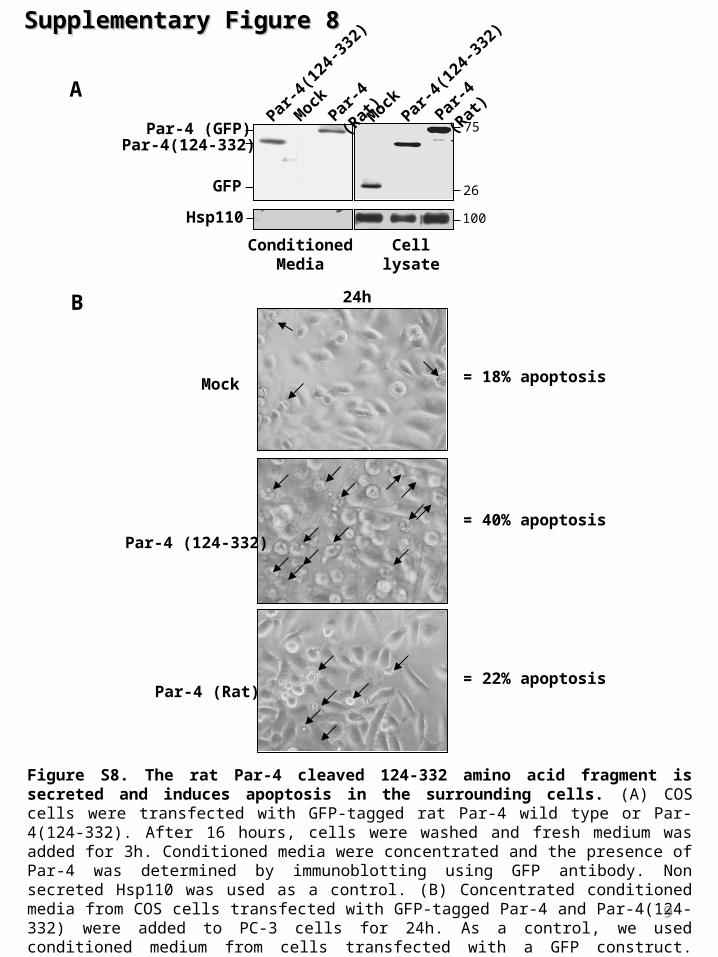
Supplementary Supplementary Figure 8Figure 8
Figure S8. The rat Par-4 cleaved 124-332 amino acid fragment is secreted and induces apoptosis in the surrounding cells. (A) COS cells were transfected with GFP-tagged rat Par-4 wild type or Par-4(124-332). After 16 hours, cells were washed and fresh medium was added for 3h. Conditioned media were concentrated and the presence of Par-4 was determined by immunoblotting using GFP antibody. Non secreted Hsp110 was used as a control. (B) Concentrated conditioned media from COS cells transfected with GFP-tagged Par-4 and Par-4(124-332) were added to PC-3 cells for 24h. As a control, we used conditioned medium from cells transfected with a GFP construct. Apoptosis was visualized by microscopy and percentages of cell death evaluated by FACS analysis. 9
Supplementary Figure 9Supplementary Figure 9
Ph231-hPar-4
Hsp90
hPar-4
- + - +
- + + +
TBB
CalA
Hsp90
hPar-4
A
B
43
90
43
90
43
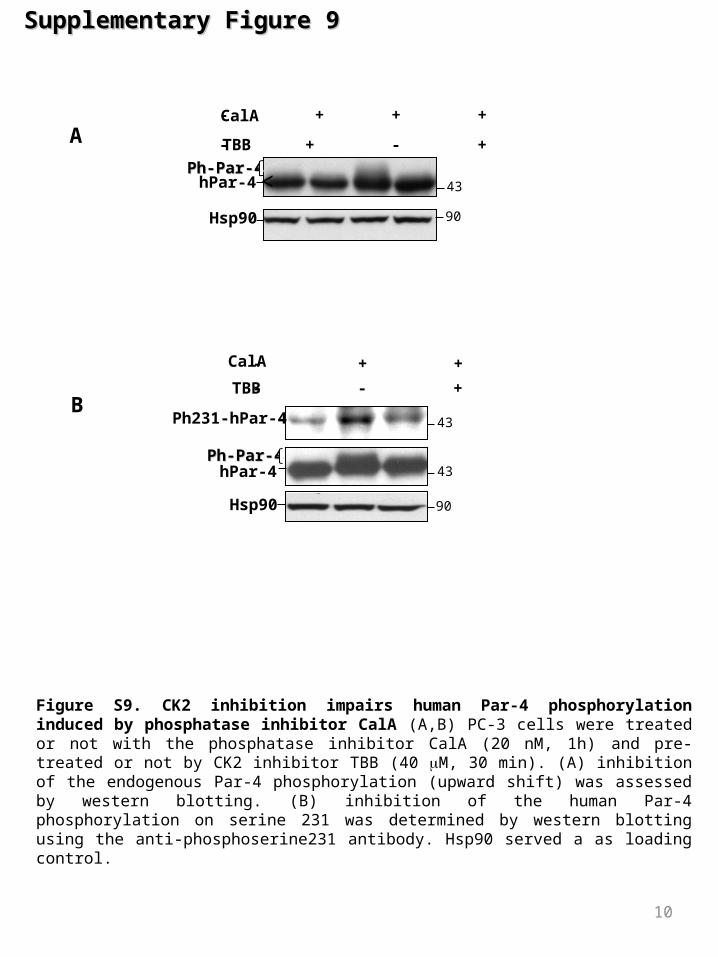
Figure S9. CK2 inhibition impairs human Par-4 phosphorylation induced by phosphatase inhibitor CalA (A,B) PC-3 cells were treated or not with the phosphatase inhibitor CalA (20 nM, 1h) and pre-treated or not by CK2 inhibitor TBB (40 M, 30 min). (A) inhibition of the endogenous Par-4 phosphorylation (upward shift) was assessed by western blotting. (B) inhibition of the human Par-4 phosphorylation on serine 231 was determined by western blotting using the anti-phosphoserine231 antibody. Hsp90 served a as loading control.
- - +
- + +
TBB
CalA
Ph-Par-4 Ph-Par-4 <
Ph-Par-4 Ph-Par-4
10

90
28
90
43
43
55
26
Supplementary Figure 10Supplementary Figure 10
Par-4
CK2
PARP
Par-4
sc/
Scr
Par-4
sc/C
K2AB
ScrCK2A
B/Scr
Cleaved PARP
Hsp90
TRAIL
Figure S10. Anti-apoptotic role of CK2 is dependent on Par-4 in the human resistant prostate cancer cells. PC-3 cells were transfected with different CK2 siRNA (siCK2T from ThermoFischer or siCK2AB from Ambion) together with scrambled siRNA fluorescently labeled with FAM (Scr siRNA) or Par-4 siRNA (siPar-4sc from Santa-cruz) for 48h hours. Then, cells were treated or not with TRAIL (3h, 500ng/ml). Apoptosis were measured by PARP/caspase-8 cleavage. The downregulation of Par-4 and CK2α proteins was confirmed by immunoblot using the corresponding antibodies. Hsp90 was used as a loading control. Bar graphs show semi-quantified densitometry from Caspase-8 and PARP Western blot analysis of the Figure 7B (A) and panel (B).
Cleaved Casp8
Casp8
11
A
B
PARP cleavage
Casp8 cleavage
PARP cleavage Casp8 cleavage
Nor
mal
ized
rat
io C
leav
ed P
AR
P/ P
AR
P (
a.u)
Nor
mal
ized
rat
io C
leav
ed C
8/C
8(a.
u)
Nor
mal
ized
rat
io C
leav
ed P
AR
P/ P
AR
P (
a.u)
Nor
mal
ized
rat
io C
leav
ed C
8/C
8(a.
u)
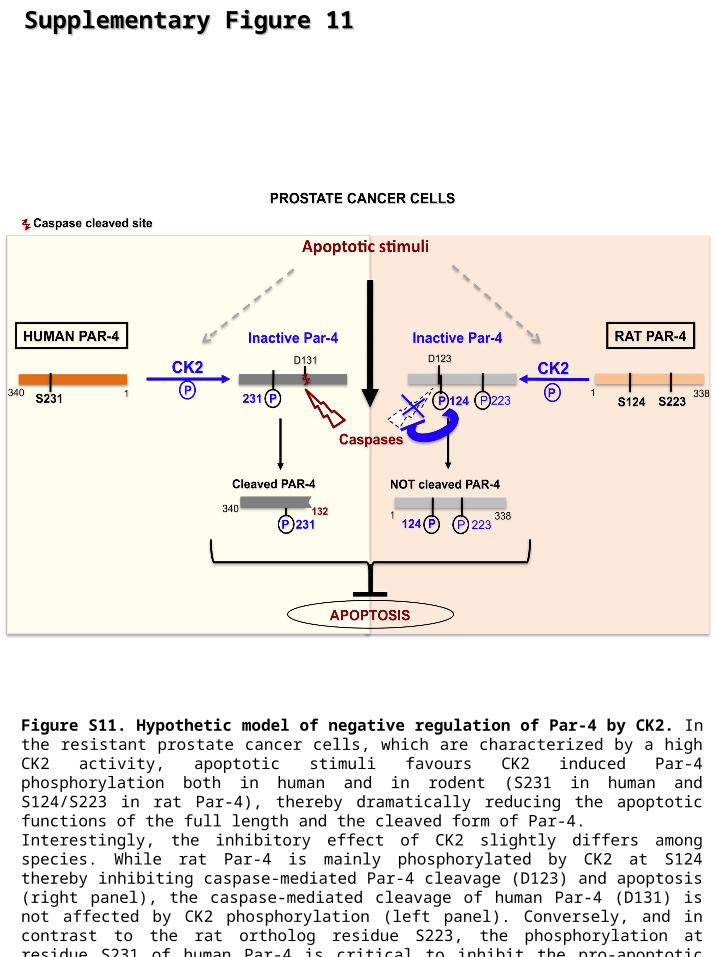
Figure S11. Hypothetic model of negative regulation of Par-4 by CK2. In the resistant prostate cancer cells, which are characterized by a high CK2 activity, apoptotic stimuli favours CK2 induced Par-4 phosphorylation both in human and in rodent (S231 in human and S124/S223 in rat Par-4), thereby dramatically reducing the apoptotic functions of the full length and the cleaved form of Par-4. Interestingly, the inhibitory effect of CK2 slightly differs among species. While rat Par-4 is mainly phosphorylated by CK2 at S124 thereby inhibiting caspase-mediated Par-4 cleavage (D123) and apoptosis (right panel), the caspase-mediated cleavage of human Par-4 (D131) is not affected by CK2 phosphorylation (left panel). Conversely, and in contrast to the rat ortholog residue S223, the phosphorylation at residue S231 of human Par-4 is critical to inhibit the pro-apoptotic functions of Par-4 (both full length and cleaved form).
Supplementary Figure 11Supplementary Figure 11

SUPPLEMENTAL DATA
Par-4 phosphorylation is regulated by phosphatase PP1
Par-4 is a highly phosphorylated protein and the addition of a broad inhibitor of phosphatases
like Calyculin A induced an upward shift which increases in a time dependent manner
(Fig.Suppl. Fig.2A). Knowing that several substrates of CK2 are dephosphorylated by PP1 (ref.46),
we compared the efficacy of CalA with that of Okadaic Acid (OA). The latter inhibitor has the
ability to inhibit PP2A at low doses (<0.5µM) whereas it blocks both PP1 and PP2A at high
doses (>1µM). We observed that OA had no effect on human endogenous Par-4
phosphorylation at low doses, thus suggesting the involvement of PP1 (Fig.Suppl. Fig.2B). In
that respect, we observed that both endogenous human Par-4 and PP1 co-immunoprecipitate
(Fig.Suppl. Fig.2C). To further confirm that dephosphorylation of Par-4 is PP1-dependent, we
added to the lysate of cells treated with CalA, recombinant PP1 enzyme and evaluated Par-4
phosphorylation status. Indeed, the addition of recombinant PP1 that overcome the CalA
inhibitory effect leads to an efficient dephosphorylation of Par-4 (Fig.Suppl. Fig. 2D).
13

SUPPLEMENTAL EXPERIMENTAL PROCEDURES
35S metabollic labeling
In vitro translation of the subunit GST-CK2 or and GST-Par-4 were performed using the
TNT assay (Promega, Charbonnières-les-Bains, France) following the manufacturers instruction:
0,5 µg of each gene was transcribed by phage T7 polymerase and 35S-labeled methionine (10
µCi) was incorporated into the translated proteins. To test the yield, each reaction was submitted
to autoradiography. For immunoprecipitation, the CK2 subunits were incubated with GST-Par-4
or GST alone pre-associated with glutathion beads in PBS, 0,3% triton, BSA 5 mg/ml, 0,01%
tween, protease inhibitors cocktail (Roche diagnostic) for 90 min. After washes,
immunocomplexes were denaturated in Laemmli buffer and resolved in SDS-PAGE 12%. The
gels were stained in Coomassie blue and dried before exposure to autoradiography.
Protein Phosphatase Assay
5x105 cells were lysed in 50 mM Tris-HCl (pH 7.5), 1% NP40, 1mM EDTA, 100 mM NaCl.
80µg of protein were added to a final volume of 100 µl in 20 mM Hepes (pH 7.4), 2 mM MnCl
2,5mM DTT with 100 or 200µg of recombinant phosphatase PP1 (ref.2) and incubated 45 min
30°C.
Secretion analysis
The medium of transfected Cos cells was replaced by fresh free medium and incubated 3 hours.
Conditioned media were harvested and concentrated using microcon 10 or Amicon ultracel-10k
following the manufacturer’s instruction (Millipore). For western-blot, Laemmli Buffer was
added and samples were run on 10% SDS-PAGE. For induction of apoptosis, conditioned media
was added on PC-3 cells for 48h and apoptosis was assessed by FACS.
14

SUPPLEMENTAL REFERENCES1. Eriksson, J. E., Toivola, D. M., Sahlgren, C., Mikhailov, A. and Harmala-Brasken, A. S. Strategies to assess phosphoprotein phosphatase and protein kinase-mediated -mediated regulation of the cytoskeleton. Methods Enzymol., 1998; 298, 542-69.2. Renglin, A., Harmala-Brasken, A. S., Eriksson, J. E. and Onfelt, A. Mitotic aberrations induced by carbaryl reflect tyrosine kinase inhibition with coincident up-regulation of serine/threonine protein phosphatase activity: implications for coordination of karyokinesis and cytokinesis. Mutagenesis, 1999; 14, 327-33.3. Spencer, W., Kwon, H., Crepieux, P., Leclerc, N., Lin, R. and Hiscott, J. Taxol selectively blocks microtubule dependent NF-kappaB activation by phorbol ester via inhibition of IkappaBalpha phosphorylation and degradation. Oncogene,1999; 18, 495-505.
15