swelling properties of cross-linked dna gels
TRANSCRIPT

Advances in Colloid and Interface Science 158 (2010) 21–31
Contents lists available at ScienceDirect
Advances in Colloid and Interface Science
j ourna l homepage: www.e lsev ie r.com/ locate /c is
Swelling properties of cross-linked DNA gels
Diana Costa a,b,⁎, M. Graça Miguel b, Björn Lindman a,b
a Physical Chemistry 1, Centre for Chemistry and Chemical Engineering, Lund University, Box 124, S-22100 Lund, Swedenb Department of Chemistry, University of Coimbra, Coimbra, Portugal
⁎ Corresponding author. Departamento de Química, U535 Coimbra, Portugal. Fax: +351 239 82 7703.
E-mail address: [email protected] (D. Costa).
0001-8686/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.cis.2009.10.002
a b s t r a c t
a r t i c l e i n f oAvailable online 5 November 2009
This work represents our contribution to the field of physical chemistry of DNA gels, and concerns thesynthesis and study of novel chemically cross-linked DNA gels. The use of covalent DNA gels is a verypromising way to study DNA–cosolute interactions, as well as the dynamic behaviour of DNA and cationiccompacting agents, like lipids, surfactants and polycations. Manipulating DNA in new ways, like DNAnetworks, allows a better understanding and characterization of DNA–cosolute complexes at the molecularlevel, and also allows us to follow the assembly structures of these complexes.The use of responsive polymer gels for targeted delivery of toxic and/or labile drugs has, during the past fewyears, shown to be a promising concept. The features found in the proposed system would find applicationsin a broader field of gel/drug interaction, for the development of controlled release and targeted deliverydevices.niversidade de Coimbra, 3004-
l rights reserved.
© 2009 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 212. Gel formation and characterization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
3. DNA gel deswelling on binding cosolutes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233.1. General aspects of polyelectrolyte gel swelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
3.1.1. Cationic dyes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233.1.2. Simple electrolytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 243.1.3. Polycations and polyamines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263.1.4. Cationic surfactants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
4. Double stranded DNA gels versus single stranded . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 275. Kinetics of surfactant-induced gel deswelling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 286. Skin formation on surfactant binding . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 297. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
1. Introduction
Polymer gels that respond to changes in the surroundingenvironment with a volume transition, often referred to as responsivegels, have attracted much interest in the last few years [1–8]. In thisgroup are the polyelectrolyte gels, which consist of charged polymernetworks, counterions and solvent and are usually synthesized bychemically cross-linking charged or titrating polymers. The environ-
mental conditions include changes in different parameters such as pH[6,9–11], solvent composition [12], ionic strength [13], temperature[14,15], pressure [16], buffer composition [17], surfactants [18–27]and photoelectric stimuli [28]. Because of their significant swellingand syneresis in response to external stimuli, these polymericnetworks are used for a variety of applications such as contact lenses,[29] diapers, wound dressing, membrane materials, pharmaceuticalproducts, [30] monolithic drug delivery systems, [31] chromatogra-phy packing materials and agriculture [32]. Moreover, cross-linkedgels have been investigated for many biomedical uses including tissueculture [33], enzyme activity controlling systems [34], materials forimproved biocompatibility [35], in the design and analysis of artificial

Fig. 1. Scanning electron micrograph of a freeze-dried ds-DNA gel (1% cross-linker),preswollen in 1 mM NaOH solution.
22 D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
muscles and biosensors and in the design of intelligent controlleddrug release devices for site-specific drug delivery. The scientificinterest is due to the complex interplay of counteracting forces, whichare of equal order of magnitude and which determine the swelling ofthe gels [35]. The main contributions are the osmotic pressure orig-inating from the electrostatically confined small ions inside the gel[36], the effective electrostatic interactions between the chargedgroups [37–40] and hydrophobic interactions.
The swelling is restricted by the cross-linking density in thenetwork and the deswelling by the volume occupied by the polymernetwork. The extent of the collapse is affected by molecular param-eters of the network such as the cross-linking density, the hydropho-bic/hydrophilic balance of the polymer network, its content ofcharged groups, its flexibility and its ability to interact with addedsolutes.
Complexes between DNA and cationic surfactants have attractedan increased interest lately due to the possibility of using thesesystems for gene transfection [41]. The nature of both the DNA [42]and the surfactant [43–45] has been found to influence the phaseseparation limits and also the structure of the DNA–surfactantcomplexes. The DNA–surfactant gel systems are expected to followthe behaviour of polyelectrolyte/surfactant gels, in general. Whenpolyelectrolyte gels interact with oppositely charged surfactants,interesting features can occur [46–49]. Swollen polyelectrolyte net-works undergo a collapse after absorbing equimolar amounts ofsurfactant, whereas where there is not enough surfactant available toform complexes involving all polyion chains in the network, a regionin the network remains in a swollen state coexisting with the col-lapsed part. The collapsed region, containing the surfactant aggre-gates, makes up a surface phase (skin) surrounding thewater-swollennetwork (core) [50] This phenomenon has also been observed in gelsabsorbing proteins or polyions of opposite charge to the network[51,52].
During the last 5 years we have paid considerable attention todifferent aspects of covalent DNA gels, concerning their preparation,their characterization, the study of swelling isotherms in the presenceof different cosolutes, such as surfactants, and their light induceddisruption. All this can be used as a basis for the development/manipulation of novel systems to be used in a variety of applications.It is possible to use them to monitor DNA–cosolute interactions bysimply monitoring volume changes. The possibility to use them inbiotechnological applications (separation) is another consideration.With the development of this work, other important challenges arise,namely in the biomedical field, as devices for controlled release andtargeted delivery [49,53–59].
2. Gel formation and characterization
In his pioneering work on covalent gels, which exhibit a discretevolume phase transition in water–acetone mixtures by a solventcomposition change, Tanaka [60] also included the preparation andstudy of covalent DNA hydrogels. Later, Horkay et al. cross-linked DNAand investigated the osmotic properties of these gels in aqueoussolutions containing NaCl and CaCl2 [61]. Recently, another groupsynthesized DNA gels and monitored the gelation reactions of DNAand ethylene glycol diglycidyl ether (EGDE) in aqueous solutions byclassical rheometry using oscillatory deformation tests [62]. In ourgroup we prepared DNA networks, by cross-linking DNA from salmontestes with EGDE [49], which is a bifunctional cross-linker withepoxide structure. As shown by another group [63], epoxides co-valently bind to the guanine bases of the DNA molecule. To thesecompounds, we added N,N,N′,N′,-tetramethylethylenediamine(TEMED) as an initiator of the reaction. Gels with 0.5%, 1%, 3%and 5% cross-linker were prepared. The reaction mechanism of gelformation seems to involve the guanine nitrogen atom at positionseven (N-7), which attacks themore substituted carbon of the epoxide
or the least hindered end of the epoxide. The reaction is of nucleophilicsubstitution type (SN2 reaction) [59]. All cross-linked double andsingle-stranded DNA gels in their swollen state are clear, transparent,porous and with a three-dimensional coral-like structure revealed byscanning electron microscopy (SEM) studies (Fig. 1). Rheologicalanalysis shows that these gels display strong elastic properties andhave a shear-thinning behaviour, as illustrated in Fig. 2 [53].
In neat water, the degree of swelling for single-stranded DNA gelsis 2 orders of magnitude larger compared to that of double strandedones, attributed to the larger chain flexibility. The difference becomessmaller when both gels are immersed in 1 mM NaOH. Moreover, alower cross-linker density was found, as expected, to lead to anincreased swelling.
In addition, we have monitored the light induced disruption of theDNA network by DNA release, gel weight loss and significant changesin both the storage modulus and in the degree of swelling [58]. Gelscross-linked with 0.5% and 1% EGDE presented an increased extent ofswelling with increasing time of disruption, while those cross-linkedwith 3% and 5% EGDE showed only small changes in the swelling ratio,as presented in Fig. 3. Experimental results demonstrate that theprotein release behaviour from these gels was greatly affected by thedegree of network swelling and by variations in the network meshsize [58,59].
Information about the conformational state of the DNA moleculesin the gels was obtained by both fluorescence measurements usingethidium bromide (EtBr), as a fluorescent probe, and fluorescencemicroscopy using acridine orange, as a dye. When EtBr is bound tonucleic acids, a marked enhancement in its fluorescence is observed;however this increase depends on the configuration of the DNA.Double stranded DNA gives a higher increase in ethidium fluorescenceintensity than single stranded DNA [64]. Ethidium bromide is a dyethat binds to double stranded DNA by intercalation between the basepairs. Acridine orange intercalates into double stranded DNA as amonomer, whereas it binds to the single-stranded DNA conforma-tional state as an aggregate. Upon excitation at 470 nm to 490 nm, theacridine orange monomeric form binds to ds-DNA and fluorescesgreen. The aggregated acridine orange on ss-DNA fluoresces red [65].Through the observation of green or red fluorescence, DNA fluores-cencemicroscopy studies with acridine orange have revealed the DNAconformational state in the gels. Information about the DNA con-formational state in the cross-linked gels was obtained mainly byfluorescence measurements using ethidium bromide, as a fluorescentprobe. Ethidium bromide spectral measurements show that thepreparation of the DNA gels is carried out with the conservation of

Fig. 2. Dependence of viscosity on the shear rate demonstrates a shear-thinningbehaviour for chemical DNA gels.
23D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
its native, double stranded conformation. Moreover, the denaturationof covalent ds-DNA gels was induced by heating above the meltingtemperature. The melting temperature was shown to be around 85 ºC(Fig. 4). This process appears to be reversible when a heating/coolingcycle was performed. Also, by fluorescence experiments withethidium bromide, a double stranded conformation of DNAmoleculeswas found for collapsed gels on addition of mono- and divalent salts,different classes of surfactants, chitosan and polyamines. In addition,in the gels collapsed by terbium, DNA fluorescence microscopystudies, using acridine orange as staining, were performed. With theexception for the trivalent lanthanide ions, it appears that the DNAchain always retains a double-helix conformation; with these metalions single-stranded DNA is found.
3. DNA gel deswelling on binding cosolutes
3.1. General aspects of polyelectrolyte gel swelling
In pure water, charged networks swell, due to the osmoticpressure from the counterions [35], which originates from theirtranslational entropy. The counterions are driven to leave the networkin order to increase their entropy, but the requirement of total
Fig. 3. Change of the degree of swelling of DNA gels cross-linked with 0.5%, 1%, 3% and5% (w/v) EGDE as a function of time. The weight of the gels preswollen in the 1 mMNaOH solution was taken as the first point (this swelling ratio given as unity) and,therefore, represents the equilibrium degree of swelling of the “intact” covalently cross-linked DNA gels.
electroneutrality forces the ions to stay inside the gel. The osmoticpressure of the reservoir is zero and the gel swells until thecontribution of the counterions to the osmotic pressure is balancedby the elastic contribution. Gel deswelling can be induced in differentways. Here we consider two effects, one simple electrolyte effectgiving a reduced difference in counterion concentration between thegel inside and the reservoir, and one associating effect, where acosolute associates to the gel network and effectively decreases thepolyelectrolyte charge. For polyelectrolyte gels, which mainly swelldue to the osmotic pressure arising from the confined counterions, thepresence of salt thus leads to a decrease of the gel volume. The saltconcentration outside the gel is larger than inside the gel, and also theincrease of the osmotic pressure due to salt addition is larger in thereservoir than inside the gel. The volume changes of gels areassociated with osmotic effects and not dehydration. A ratherdifferent behaviour is obtained when there is an association of anoppositely charged cosolute, which can be dominated by electrostaticeffects for an oppositely charged polyelectrolyte, or influenced bynon-electrostatic effects, like the binding of dyes to DNA, driven byhydrophobic interactions. Oppositely charged surfactants constitute arather special case in that both a hydrophobic interaction with DNAbases and a polyion–polyion association effect are important. Thuswhen a polyelectrolyte gel is immersed in a solution of oppositelycharged surfactant, the surfactant ions migrate into the network andform micelles at a concentration, cac, which is much lower than thecritical micelle concentration, cmc, of the pure surfactant; themicellesassociate to the gel network. This process is accompanied by ashrinking of the gel and the driving force is all the small counterions,both from the gel and the surfactant molecules, which can be releasedwhen the surfactant binds to the charged gel network. Monitoring thevolume change of a gel, therefore, can give direct indications of thebinding behaviour of a certain surfactant to a polymer; changes in thebinding of the surfactant to the polymer are accompanied by amacroscopic change in the gel volume [7,18,66] The same informationis much more difficult to obtain from solutions of a linear polymer.Moreover, we verified that the gel swelling experiment is a sensitiveand powerful method, from the above discussion and in that the gelpiece can be studied in an infinite bath of surfactant solution, and so,the response of the gel is obtained as a function of the free surfactantconcentration, reflecting the limiting behaviour of the correspondinglinear polymer at infinite dilution.
3.1.1. Cationic dyesDNA interplays with a wide variety of molecules in the cell, from
large proteins to small molecules like water and organic and inorganicions. Some interactions with DNA in the cell are general while othersare highly specific. The features of binding are determined by acomplex interplay between favourable and unfavourable interactionswith DNA. The ligandsmay reside either in one of the grooves or in theinterior of the helix, inserted between adjacent base pairs in asandwich-like complex [67]. The latter type of binding, exhibited byseveral planar, polycyclic aromatic compounds, is termed intercala-tion. To accommodate an intercalator, the DNA structure must changeat and around the binding site; the binding affinity comes partly fromhydrophobic interactions with the base pairs of DNA. Groove bindingtakes place almost exclusively in theminor groove and themajority ofminor groove binders show preferentiality for AT tracts [68]. The dyemethyl green was found to be excluded from binding to triplex DNAand a major groove binding was proposed for the methyl green-DNAcomplex [69]. To probe into the molecular specificity of the DNAnetworks, the swelling isotherms of DNA gels have been investigatedin the presence of four cationic dyes, which are different in their DNAbinding mode and valence. The gel induced volume changes by 4′-6-diamidino-2-phenyl-indole (DAPI-a minor groove binder, divalent),methyl green (a major groove binder, divalent), ethidium bromideand acridine orange (both intercalators and monovalent and divalent,

Fig. 4. Fluorescence spectra of ethidium bromide in the absence and presence of a long ds-DNA gel (1% cross-linker density) after heating until 95 °C (A) and 85 °C (B) and in thepresence of reference solutions of ds-DNA 1% and ss-DNA 1% (C), at 25 °C.
24 D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
respectively) were studied. Fig. 5 shows swelling isotherms for DNAgels in the presence of the dyes. Their features are quite different. Atlow concentrations of DAPI, a small plateau is observed followed by astrong collapse of the gels in a narrow concentration region and amore progressive deswelling thereafter. In the presence of methyl
Fig. 5. Swelling isotherms (V/V0) for DNA gels (1% cross-linker) immersed inDAPI,methylgreen, acridine orange and ethidium bromide solutions. Temperature: 25 °C, pH 9.
green, the plateau, indicating no gel volume changes, is very large andthe deswelling process starts only at a concentration of 6 mM. Fromthis concentration, there is a shrinking of the hydrogel, but it does notentirely collapse, even in the saturated solution. This is in contrast toDAPI, which induces a pronounced degree of gel collapse. Ethidiumbromide and acridine orange behave similarly. At low concentrations,the existence of a small plateau is verified. As the concentration isincreased, we can observe a gradual deswelling of DNA hydrogelsuntil they reach the collapse equilibrium, with gels in the presence ofethidium bromide showing a slightly higher deswelling. Theseswelling studies using different dyes revealed that the gel couldchange its own volume when binding with specific molecules andsuch changes in gel size strongly depend on the binding mode of thedye, more so than on the valence.
3.1.2. Simple electrolytesIn Fig. 6 the volume changes of the DNA gels in the presence of
monovalent, divalent and trivalent counterions, are represented as afunction of salt concentration [54]. When gels, preswollen in 1 mMNaOH, are placed into salt solutions at different concentrations, theyshrink due to the screening effect of the salt and mainly due to thedecreased concentration difference of mobile ions inside the gel andthe external solution governed by the Donnan equilibrium. A generalelectrostatic effect will be modulated by any specific interactions thatmight occur. For simple ions, the size of the counterion will play a role,the appropriate size being dependent on the degree of dehydrationthat occurs on binding. For binding of dehydrated ions thus we expect

Fig. 6. Swelling isotherms (V/V0) for DNA gels (1% cross-linker) immersed in solutions of monovalent, divalent and trivalent ions. Temperature: 25 °C, pH 9.
25D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
deswelling to decrease with the atomic number, while the oppositewould apply if the ions would retain their hydration layer on binding.As will be seen, our results indicate a more complex behaviour, and amixed picture, with some counterions keeping largely their state ofhydration on association with DNA, while others are extensivelydehydrated.
In general the nature of the monovalent counterion has only amoderate effect on the deswelling of DNA gels. The degree of collapsefollows the order Na+>K+>Li+>Rb+>Cs+, with the sodium ionshowing the highest ability in collapsing DNA gels. Except for lithium,this dependence follows the Hofmeister series (lyotropic series),according to which the observed deswelling effect would increasewith decreasing ionic radius of the naked ion. The order observedsuggests thatmonovalent cationswith a smaller atomic number, exceptfor Li interactmore stronglywith DNA gels [54].With its smaller atomicnumber and its higher charge density, lithium demonstrates anunexpected behaviour in this set of monovalent salts. Recently, Horkayet al. showed that the deswelling degree of polyacrylate gels in purewater follows the series: Li+>Na+>K+> Cs+ [13]. Unchargedpolyvinylpyrrolidone gels show the same cationic specificity [70].Moreover, in a study developed by Liu et al., dimethylacrylamide/2-acrylamide-2-methylpropyl sulfonate copolymer gels with differentcharge densities were immersed in salt solutions with differentcombinations of monovalent and divalent ions. The swelling degreewas shown to depend on the counterion and to follow the Hofmeisterseries [71]. ThedifferenceobservedbetweenK+ andNa+was explainedby different counterion condensation according to the Manning theory[71]. Firestone and Siegel showed that the swelling degree for cationic
methylmethacrylate/dimethylaminoethylmethacrylate (MMA/DMAEMA) (70/30) gels, immersed in salt solutions, was also dependenton the counterion and the swelling degree follows theHofmeister seriesfor anions [72]. The effect of lithium thus seems to be rather specific forcovalent DNA gels. The general trend seems to be that the swellingdegree of polyelectrolyte gels with different counterions increases withdecreasing counterion binding according to the Hofmeister series (foranions typically:highbinding/lowgel swellingand lowbinding/highgelswelling). However, the differences are typically quite small. Thedeswellingon addition of thedivalent salts CaCl2,MgCl2 andSrCl2 occursat considerably lower salt concentrations and the deswelling is morepronounced, as comparedwith themonovalent ions. The collapse ratiosfor these gels show a trend in the order Ca2+>Mg2+>Sr2+, withcalcium being the most effective ion on collapsing DNA gels. Ca2+ andother divalent ions such as Zn2+ and Mn2+, are suggested to bind bothto DNAphosphate groups and baseswhileMg2+ ions only to phosphategroups ofDNA [73]. Thismay explain theobserveddifference in theDNAgel collapse ability between calcium and magnesium ions [54].However, we note also the analogue sequences for alkali and alkalineearth ions.
As for DNA condensation in solution, the effect ofmono and divalentions is less pronounced in the gel collapse than that of trivalentlanthanide ions, showing a very pronounced deswelling at low con-centrations. As in the presence of divalent metal ions, the deswelling ofgels on addition of trivalent ions seems to follow a two-step process,suggested to involve an initial compaction of individual DNA chains.Clear differences in the efficiency of gel collapse were found betweenthe trivalent ions, with terbium being the most efficient. Considerably

Fig. 8. Swelling isotherms (V/V0) for ds-DNA gels (1% cross-linker) immersed insolutions of chitosan and then immersed in solutions of NaBr. Temperature: 25 °C, pH 9.Chitosan concentrations are presented in mM on a charge basis.
26 D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
lower concentrations of terbiumare required to induce a decrease in thegel volume,when compared to ceriumandeuropium,with thedegreeofcollapse being in the order Tb3+>Ce3+>Eu3+ [54].
From the results for simple electrolytes we can conclude that themain points affecting the gel collapse are the ion valency and ionspecific effects; the valency is the dominating effect indicating thedominant role of general electrostatic effects.
3.1.3. Polycations and polyaminesThe swelling behaviour of DNA gels was studied in the presence of
several poly-cationic species, which include chitosan, spermine(Spm), spermidine (Spd), lysozyme, poly-L-lysine (PL) and poly-L-arginine (PA) (Fig. 7) [54]. Chitosan has the highest potential toinduce collapse of DNA gels in the range of concentrations studied.The deswelling process starts already at very low concentrations ofchitosan, 0.1 mM. In similar studies, using other cations, higher con-centrations of the cosolute are needed, like in the case of the poly-amines. Moreover, the cationic protein lysozyme has the capability tocollapse DNA gels, but to a smaller extent than the agents mentioned.Lysozyme has a lower charge density than the other cosolutes inves-tigated; it has both cationic and anionic groups resulting in adecreased net charge. This leads to a lower degree of binding. Theaddition of the polypeptides, poly-L-arginine and poly-L-lysine, leadsto gel deswelling but at higher concentrations. No significant changesin the volume of the gels were observed until 0.5 mM. Poly-L-lysine isthe weakest agent in the collapse of DNA gels. The swelling behaviourappears to be reversible; thus when the gels, collapsed by addition ofchitosan, polyamines and charged proteins, were immersed insolutions of NaBr they reswell, due to the screening effect of salt onthe association of the polycations to the DNA network. (Fig. 8).
3.1.4. Cationic surfactantsIn the bulk phase and at low concentrations, the surfactant
molecules are dissolved as unimers, whereas at higher surfactantconcentrations a self-assembly into aggregates occurs. For the single-chain surfactants studied here, the aggregates formed in this self-assembly are commonly spherical micelles with micelle formationstarting at a well-defined concentration, the critical micelle concen-tration (cmc). The cmc depends on the chemical structure of thesurfactant, surfactants with longer hydrophobic tails having lowercmc values. In the presence of an oppositely charged polyelectrolyte,the micelle formation of an ionic surfactant is strongly facilitatedleading to a major lowering of the cmc; the cmc in the presence of apolymer is often referred to as the critical association concentration,cac. The stabilisation of micelles due to an oppositely charged
Fig. 7. Swelling isotherms (V/V0) for DNA gels (1% cross-linker) immersed in solutionsof chitosan, spermine, spermidine, lysozyme, poly-L-lysine and poly-L-arginine.Temperature: 25 °C, pH 6.5. Concentrations are presented in mM on a charge basis.
polyelectrolyte is mainly an entropic effect, due to a release of coun-terions. Therefore, the critical aggregation concentration, for thesurfactant in the presence of DNA, cac, is lower than the criticalmicellization concentration, cmc. The fact that the cationic surfactantbinding occurs preferentially to anionic polyelectrolytes of highcharge density further enhances this behaviour [74,75].
The effect of an oppositely charged surfactant on the swelling of apolyelectrolyte gel is, because of this self-assembly, expected to bemore complex than for the other cosolutes considered. At pre-cacconcentrations a cationic is thus expected to behave as a simple 1:1electrolyte, with some additional effect due to the hydrophobic inter-actions between surfactant unimers and the DNA bases. From the cac,highly charged surfactant self-assemblies are present; these act asother polycations and are expected to be very efficient in the DNA geldeswelling. As we illustrate below cationic surfactants do indeedshow an insignificant deswelling below the cac, while they are strongdeswelling agents above the cac. Both the onset of deswelling, and thedegree of deswelling, correlate nicely with the surfactant alkyl chainlength.
The DNA gels are as mentioned highly swollen due to the osmoticpressure arising from the counterions, which are confined to the gel.After the immersion of the swollen double stranded DNA gel in asolution of an oppositely charged surfactant with long alkyl chainlength, like cetyltrimethylammonium bromide (C16TAB), the surfac-tant ions migrate into the network and replace the network counter-ions which are released. This process is extremely favourable from thepoint of view of translational entropy of the counterions [76]. Adsorp-tion of a considerable amount of surfactant ions leads to a transition ofthe swollen network to the collapsed state. The main reason for thistransition is as indicated the aggregation of surfactant ions within theDNA gel due to hydrophobic interactions between their hydrocarbonchains; the surfactant self-assemblies are effectively bound to theDNA chains, thus reducing the charge density of the gel network. As aconsequence of this, the mobile counterion concentration in thenetwork decreases, leading to a significant decrease in the internalosmotic pressure in the gel. Furthermore, the surfactant aggregateswill act as multivalent counterions and by ion correlation effectscontribute to the contraction of the gel [77,78]. From Fig. 9 it can beinferred that there is no volume change for C16TAB concentrationsbelow 0.02 mM [49]. Above this concentration, there is a dramaticdecrease in the size of the gels in a very narrow concentration range.Above the collapse concentration, which we interpret as the criticalaggregation concentration (cac) of the surfactant, the gels have ahomogeneous composition, but the degree of swelling decreasesgradually with increasing concentration in the solution. The

Fig. 9. Swelling isotherms (V/V0) for ds-DNA gels (1% cross-linker) immersed insolutions of the cationic surfactants C16TAB, C14TAB, C12TAB and C8TAB. Theconcentration range from up to 0.2 mM is also represented in detail (see inset).Temperature: 25 °C, pH 9.
27D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
aggregation of surfactant in the DNA network starts at a concentrationthat is much lower than the cmc of the surfactant in water (0.9 mM)[79]. This is so because normal micellization involves counterionbinding to neutralize the charge of the micelle, while binding ofsurfactant to the DNA network involves the release of the smallcounterions. In accordance with this, a cac of 0.02 mMwas found andthis is in excellent agreement with cac values for CTAB in solutionswith DNA, as obtained by other methods [80–82].
Again the gel swelling appears to be reversible, since when DNAgels collapsed by the addition of CTAB were immersed in SDSsolutions the relative volume (V/V0) returned to between 90 and100% of the initial value, as shown in Fig. 10. The underlying mech-anism of the deswelling is the strong interaction between the anionicand cationic surfactant, decreasing the cationic surfactant activity,thus effectively removing the cationic surfactant from the DNA chains.
To study the effect of surfactant tail length on gel deswelling,different surfactants were used. Fig. 9 shows swelling isotherms forDNA gels on addition of a number of cationic surfactants. As can beseen, the surfactants have no effect at lower concentrations, but there
Fig. 10. Swelling isotherms (V/V0) for ds-DNA gels (1% cross-linker) immersed insolutions of C16TAB, and thereafter in solutions of sodium dodecyl sulphate (SDS).
is amarked deswelling at higher concentrations, which becomesmoreimportant the longer the surfactant alkyl chain. We note that theconcentration of onset of deswelling varies by orders of magnitudebetween different surfactants. We also note that the plateau valueobtained at high surfactant concentrations is lower the longer thealkyl chain length. The pronounced chain length dependences directlysuggest a dominant role of surfactant self-assembly. In particular,these results for the different alkyl chain lengths confirm that thedeswelling occurs below the normal critical micelle concentration ofthe surfactant. The cmc values for C16TAB, C14TAB, C12TAB and C8TABare 0.9 mM, 3.5 mM, 15 mM and 144 mM, respectively. We found thatthe surfactants induce the volume transition starting at a certainrather well-defined concentration ~0.02 mM for C16TAB [49],~0.05 mM for C14TAB, ~0.1 mM for C12TAB and ~1 mM for C8TAB[54]; these values are somewhat higher than the observed cac valuesin bulk solution [83]. The reason for this is probably the constraintsexperienced by the polyelectrolyte in the gel network; the polymer–surfactant association is weaker for polymer chains with lowerconformational freedom. The initial plateau, indicating no volumetransition of the gels, in the concentration range under study is thusvery narrow for C16TAB but increases for C14TAB and C12TAB, whileC8TAB shows a very large plateau. This effect was also shown forethylhydroxiethylcellulose (EHEC) gels in solutions with alkyltri-methylammonium bromides [66] and for hydroxyethyl cellulose(HEC) gels in solutions with sodium alkyl sulfates [7]. The gel collapsedisappears when the hydrophobic tail is too short.
4. Double stranded DNA gels versus single stranded
The effect of DNA molecular weight and conformation on theswelling behaviour was investigated using the same cosolutes as inthe previous study [55]. For this purpose, the following well-definedDNA samples have been used to prepare cross-linked DNA gels: longss-DNA (heat-denatured 2000 base pairs), short ds-DNA (146 basepairs) and short ss-DNA (580 and 831 base pair fragments). Whenstudying the effect of mono- and divalent metal ions, and polyaminesand polycations, on the swelling behaviour of double and single-stranded DNA gels, we found that the extent of swelling depends onboth DNA molecular weight and conformation. The DNA molecularweight has only a slight effect on the deswelling curves; short DNAgels show a slightly larger deswelling. The DNA conformation shows amore pronounced effect on the swelling degree. For all systemsmentioned above, single-stranded DNA gels collapse more efficientlythan double stranded DNA gels. This difference is more pronouncedwith surfactants than with the other additives mentioned above [55].It was found that the surfactants induce the volume transition startingat a certain rather well-defined concentration (critical aggregationconcentration). For gels of both long and short ds-DNA cac ~0.02 mMfor C16TAB, ~0.05 mM for C14TAB, ~0.1 mM for C12TAB and ~1 mM forC8TAB; for gels of both long and short ss-DNA cac ~0.015 mM forC16TAB, ~0.045 mM for C14TAB, ~0.08 mM for C12TAB and ~1 mM forC8TAB [55]. Except for C8TAB, also the decreased cac values displayedby CnTAB/ss-DNA gel systems demonstrate that, with the single-stranded conformation the association starts at lower surfactantconcentrations. An analogous result was found in solution studies,where the cac value for the ss-DNA/CTAB system is about half of thecac of ds-DNA/CTAB [83]. In order to explain this observation, thedifferences between double and single-stranded DNA molecules areconsidered. Firstly, ss-DNA has rather high flexibility (small persis-tence length), while ds-DNA is quite rigid and characterized by a largepersistence length (~500 Å [84]). Furthermore, ss-DNA has a lowerlinear charge density and a larger hydrophobicity as compared to ds-DNA. From a simple electrostatic mechanism, ds-DNA should interactmore strongly with oppositely charged polyelectrolytes or surfactantmicelles. On the other hand, a flexible chain interacts more stronglywith an oppositely charged macroion than a rigid one [85]. When a
28 D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
polyelectrolyte contains some hydrophobic groups, the associationwith oppositely charged surfactants is much strengthened because ofthe combination of electrostatic and hydrophobic attraction. Thehydrophobic character varies with DNA conformation: in ds-DNA, thehydrophobic groups are largely hidden, while in ss-DNA they areexposed to the solution. Thus, the hydrophobic interactions should bemore significant for the latter DNA conformation. All these factorsinfluence the extent of gel collapse. The low chain stiffness and thehigher hydrophobicity of ss-DNA lead to an increased deswelling ofgels in the presence of added cosolutes such asmetal ions, polyaminesand polycations. A cationic surfactant should associate with DNAmainly by electrostatic interactions and both linear charge densityand DNA flexibility will have important influence. The cac decreaseswith increasing hydrophobicity of the network as seen here except forthe surfactant with the shortest alkyl chain. Due to the shorthydrophobic tail of C8, the effect of increasing the hydrophobicattractive force with ss-DNA becomes less important. Other authorshave shown that when the hydrophobicity of a polymer gel increasesthe cac decreases [86]. Recently, it was suggested that there is aminimum hydrophobicity threshold, which must be overcome inorder for an association to occur [87].
5. Kinetics of surfactant-induced gel deswelling
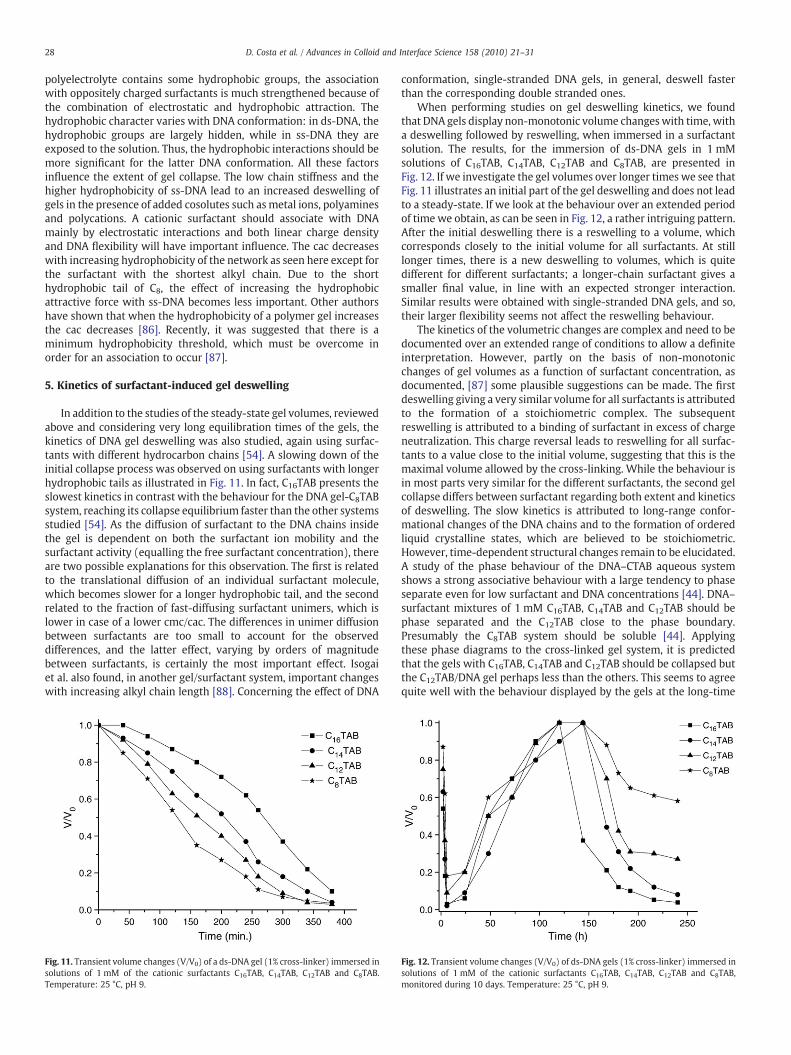
In addition to the studies of the steady-state gel volumes, reviewedabove and considering very long equilibration times of the gels, thekinetics of DNA gel deswelling was also studied, again using surfac-tants with different hydrocarbon chains [54]. A slowing down of theinitial collapse process was observed on using surfactants with longerhydrophobic tails as illustrated in Fig. 11. In fact, C16TAB presents theslowest kinetics in contrast with the behaviour for the DNA gel-C8TABsystem, reaching its collapse equilibrium faster than the other systemsstudied [54]. As the diffusion of surfactant to the DNA chains insidethe gel is dependent on both the surfactant ion mobility and thesurfactant activity (equalling the free surfactant concentration), thereare two possible explanations for this observation. The first is relatedto the translational diffusion of an individual surfactant molecule,which becomes slower for a longer hydrophobic tail, and the secondrelated to the fraction of fast-diffusing surfactant unimers, which islower in case of a lower cmc/cac. The differences in unimer diffusionbetween surfactants are too small to account for the observeddifferences, and the latter effect, varying by orders of magnitudebetween surfactants, is certainly the most important effect. Isogaiet al. also found, in another gel/surfactant system, important changeswith increasing alkyl chain length [88]. Concerning the effect of DNA
Fig. 11. Transient volume changes (V/V0) of a ds-DNA gel (1% cross-linker) immersed insolutions of 1 mM of the cationic surfactants C16TAB, C14TAB, C12TAB and C8TAB.Temperature: 25 °C, pH 9.
conformation, single-stranded DNA gels, in general, deswell fasterthan the corresponding double stranded ones.
When performing studies on gel deswelling kinetics, we foundthat DNA gels display non-monotonic volume changeswith time, witha deswelling followed by reswelling, when immersed in a surfactantsolution. The results, for the immersion of ds-DNA gels in 1 mMsolutions of C16TAB, C14TAB, C12TAB and C8TAB, are presented inFig. 12. If we investigate the gel volumes over longer times we see thatFig. 11 illustrates an initial part of the gel deswelling and does not leadto a steady-state. If we look at the behaviour over an extended periodof time we obtain, as can be seen in Fig. 12, a rather intriguing pattern.After the initial deswelling there is a reswelling to a volume, whichcorresponds closely to the initial volume for all surfactants. At stilllonger times, there is a new deswelling to volumes, which is quitedifferent for different surfactants; a longer-chain surfactant gives asmaller final value, in line with an expected stronger interaction.Similar results were obtained with single-stranded DNA gels, and so,their larger flexibility seems not affect the reswelling behaviour.
The kinetics of the volumetric changes are complex and need to bedocumented over an extended range of conditions to allow a definiteinterpretation. However, partly on the basis of non-monotonicchanges of gel volumes as a function of surfactant concentration, asdocumented, [87] some plausible suggestions can be made. The firstdeswelling giving a very similar volume for all surfactants is attributedto the formation of a stoichiometric complex. The subsequentreswelling is attributed to a binding of surfactant in excess of chargeneutralization. This charge reversal leads to reswelling for all surfac-tants to a value close to the initial volume, suggesting that this is themaximal volume allowed by the cross-linking. While the behaviour isin most parts very similar for the different surfactants, the second gelcollapse differs between surfactant regarding both extent and kineticsof deswelling. The slow kinetics is attributed to long-range confor-mational changes of the DNA chains and to the formation of orderedliquid crystalline states, which are believed to be stoichiometric.However, time-dependent structural changes remain to be elucidated.A study of the phase behaviour of the DNA–CTAB aqueous systemshows a strong associative behaviour with a large tendency to phaseseparate even for low surfactant and DNA concentrations [44]. DNA–surfactant mixtures of 1 mM C16TAB, C14TAB and C12TAB should bephase separated and the C12TAB close to the phase boundary.Presumably the C8TAB system should be soluble [44]. Applyingthese phase diagrams to the cross-linked gel system, it is predictedthat the gels with C16TAB, C14TAB and C12TAB should be collapsed butthe C12TAB/DNA gel perhaps less than the others. This seems to agreequite well with the behaviour displayed by the gels at the long-time
Fig. 12. Transient volume changes (V/V0) of ds-DNA gels (1% cross-linker) immersed insolutions of 1 mM of the cationic surfactants C16TAB, C14TAB, C12TAB and C8TAB,monitored during 10 days. Temperature: 25 °C, pH 9.

Fig. 13. Schematic representation of the two possible paths for the gel deswellingprocess. (A) In the regular deswelling, as the surfactant binds to the gel network, asurface phase forms. As more surfactant is absorbed, this surface phase grows at theexpense of the core, until the entire gel has been collapsed. (B) In the irregulardeswelling, the collapsed part makes up a surface phase (skin) rich in surfactantaggregates, surrounding a water-swollen network (liquid-filled core), and this is thefinal state of the gel.
29D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
plateau. This intriguing behaviour suggests interesting applicationsbut is far from captured in our current understanding, even more sooutside the capability of conventional modelling approaches. It is clearthat further progress is only possible after a better understanding ofmicrostructures has been obtained.
Some previous studies reported in the literature treated thereswelling behaviour on immersion of a gel in a solvent. Ohbu et al.showed that sodium dodecyl sulphate (SDS) binds to linear cationichydroxyethyl cellulose (cat-HEC) in two steps, with a resolubilizationof the complex in the second binding step [89]. Dialyldimethylam-monium chloride (DADMACl) gels were shown to reswell in solutionsof SDS [19,90]. Moreover, a study found that gels based on cat-HECfirst collapse at low SDS concentrations and later reswell at SDSconcentrations somewhat lower than the cmc of SDS. It was shownthat the first binding step indicates a normal cac in an oppositelycharged polyelectrolyte–surfactant system, while the second bindingstep, involving the reswelling, is due to a second cac [18]. Recently, theinteraction between the cationic surfactant hexadecyltrimethylam-monium (C16TA+) and a series of anionic alkylacrylamide gels ofincreasing hydrophobicity and charge density was studied [91].Depending on these properties, there was either a significantreswelling at the cac (2) (second cooperative binding step), or nosuch drastic reswelling. No reswelling occurred for sodium-2-acrylamido N, N-dimethylacrylamide (NaA:DAM) gels in hexadecyl-trimethylammonium bromide (C16TABr) solutions, while reswellingwas observed for sodium-2-acrylamido N-isopropylacrylamide (NaA:NIPA) and sodium-2-acrylamido N-tert-butylacrylamide N-isopropy-lacrylamide (NaA:BAM:NIPA) gels [91]. Sodium-2-acrylamido-2-methyl propane sulfonate (NaAMPS) copolymer gels with 94 mol%acrylamide (AAm)monomers were also reported to have a reswelling[92]. There are also examples on “equilibrium overshoots”, wherecharged gels first overswell slightly and then deswell to theequilibrium volumes [93,94].
6. Skin formation on surfactant binding
As mentioned above, when a cross-linked polyelectrolyte gel isimmersed in a solution of an oppositely charged surfactant, thesurfactant unimers will diffuse into the gel and there form micellesthat will be neutralized by the polyion, leading to the release of thecondensed counterions. The micelles form with a local collapse of thepolymer network [95].
The surfactant micelles initially induce a local collapse in thesurface of the gel, forming a micelle-rich surface phase around aswollen core, without micelles, both regions being part of the samepolymer network [48,96]. Due to the large deformation energyrequired to compress the entire network, the surface is the onlypossible position of the collapsed phase [97,98]. The formed surfacephase will be anisotropically deformed, extended more in the lateraldirection, and compressed in the transversal direction. This originatesfrom a gradual compression of the swollen gel core, induced by thepresence of the surface phase. Asmore surfactant binds to the polymergel, the surface phase grows at the expense of the core, furthercompressing it, until the entire network is in a collapsed state(Fig. 13A). This regular deswelling occurs when swollen polyelectro-lyte networks undergo a collapse after absorbing equimolar amountsof surfactant. Moreover, the volume changes depend linearly on thedegree of surfactant binding. On the other hand, if there is not enoughsurfactant available to form complexes involving all polyion chains inthe network, the gel core remains in a swollen state coexistingwith thecollapsed surface phase. The “skin”, containing the surfactant micelles,retains its outer dimensions and surrounds a liquid-filled void in themiddle of the gel, thewater-swollen core, as illustrated in Fig. 13B. Thistype of phase separation in gels was discovered by Zezin and Kabanov[50] and lately by others [48,95,99,100]. The same phenomenon hasbeen observed in gels absorbing proteins or short polyions of opposite
charge to the network [51,96]. A schematic representation of the twopossible paths, regular and irregular shrinking, is shown in Fig. 13. Theinitial part of the deswelling process is similar in the two cases, but forthe irregular case, at some point, the deswelling stops and no furtherchange in gel size is detected (Fig. 13B).
When both ds-DNA and ss-DNA gels came into contact with acationic surfactant solution, a cloudy layer formed on the surface of thegel, and water was expelled to the bulk solution. The boundarybetween the collapsed surface layer and the highly swollen interior ofthe gel is sharp. However, the final state depended both on theconcentration and the total amount of surfactant available in thesolution and the conformation of DNA [55]. Additionally, we observedthat ds-DNA and ss-DNA gels shrink in an irregular fashion with CTABwhen the surfactant concentration is higher than 0.02 mM and0.015 mM, respectively. Observations by the naked eye allow us toverify the differences concerning the formation of a surfactant skinlayer between different DNA conformations. A strongly collapsed skinis formed for ss-DNA gels at surfactant/DNA charge ratios, ß, in the gelabove 0.5 while for ds-DNA gels no macroscopic separation ofcollapsed and swollen regions was observed at low or intermediatedegrees of binding, suggesting that CTAB aggregates are evenlydistributed in the gels. There is thus a large difference in the conditionsof skin formation between different DNA conformations, ss-DNAgiving rise to skin over a much broader range of surfactant binding[55]. Skins, from both double and single-stranded DNA gels, are stableand remain unchanged for several months.
In Fig. 14, the V/V0 parameter is represented as a function of thedegree of binding, β. For ds-DNA gels, no macroscopic separation ofcollapsed and swollen regions was observed at intermediate degreesof binding. Only the ds-DNA gel with the largest ß appeared to have acore/shell structure (1mM CTAB; β=0.84; charge ratio=1.0). As theDNA molecular size does not affect substantially the swelling degreeof gels, it is assumed that short ds-DNA gels present the same kind ofbehaviour. A rather different situation is thus verified for ss-DNA gelson binding CTAB. In the case of ss-DNA gels, a surfactant rich surfacephase (skin) is found to surround a swollen core network for gels withß>0.5. Also, it is observed that the ss-DNA gel volume is smaller thanthe volume of ds-DNA gels. The amount of surfactant is not enough tocollapse the entire network. As pointed out elsewhere [48], thenetwork in a collapsed surface phase is non-uniformly deformed.Thus, in the direction parallel to the gel surface it is stretched out tothe same extent as the swollen core network, while in the directionperpendicular to that it is compressed to an extent depending on howcollapsed the phase is. One explanation, for the absence of a surface

Fig. 14. Dependence of relative volume (V/V0) of ds-DNA and ss-DNA gels (1% cross-linker density) on the degree of surfactant binding (β). All gels were equilibrated in0.50 mM CTAB solutions of different volume.
30 D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
phase at low and intermediate ß in ds-DNA gels, can thus be that theformation of a strongly collapsed skin is associated with too largedeformation energy for the compression of the very stiff DNA chainsin the direction perpendicular to the gel surface. The formation of avery concentrated phase may not be critical for the interactionbetween DNA and CTAB. However, an even distribution of surfactantin the gel is a better alternative for the system as a whole than nosurfactant binding. As mentioned above, the intrinsic characteristicsof ss-DNA, such as the greater flexibility and hydrophobicty and lowcharge density, lead to a more pronounced shrinking of gels in thepresence of cationic surfactants. In the formation of the skin layer, thelarger flexibility of ss-DNA chains plays a determinant role indecreasing the deformation energy for the compression of the DNAchains in the direction perpendicular to the gel surface.
The interactions of polyelectrolytes with oppositely chargedsurfactants are strong and can lead to complex formationwith orderedstructures. To confirm the existence of ordered structures, the skins ofboth double and single-stranded DNA gels were checked for opticalbirefringence by positioning them between crossed polarizers. Thesamples appeared birefringent (anisotropic) indicating the existenceof ordered structures. Fluorescence decay curves from ds-DNA/C16TABgels containing small amounts of a fluorescent probe (pyrene) and aquencher (cetylpyridinium chloride, CPC) were recorded with thesingle photon counting technique. The surfactant self-assembliesshowed to be, depending on the surfactant concentration, either smallglobular micelles or long rod-like structures. At low surfactant con-centrations small discrete micelles with aggregation numbers, N,160<N<210 are found, whereas large micelles (N>500) form atsurfactant concentrations of 1 mM [49]. We studied the structure ofboth types of DNA/CTAB complexes by means of small-angle X-raydiffraction (SAXS). No characteristic ordered structure, such as rod-like micelles in a hexagonal packing or a lamellar phase, was revealed.It was shown [101] that gels with higher charge densities exhibitmoredefined crystalline structures. Also, the ss-DNA/CTAB complexstructure in aqueous medium was found to be more disorganizedthan the corresponding ds-DNA/CTAB complex [102]. From this, wecan predict that skins from ds-DNA gels displayed a more defined andorganized structure when compared to the structure of ss-DNA skins.
7. Conclusions
Both single-stranded and double stranded DNA have been cross-linked at different cross-linking densities and their swelling behaviourinvestigated in particular in the presence of different cosolutes. Ageneral observation is that single-stranded DNA in the absence of any
electrolyte swells much more than double stranded DNA, which canbe understood from the higher flexibility of the polymer chains. In linewith this, is also themuch stronger collapse of single-stranded DNA inthe presence of a wide range of cationic cosolutes. In general, cationiccosolutes give a progressively larger deswelling as concentrationincreases, the degree of collapse increasing with the number ofcharges. Surfactants are different in that the deswelling is minimalbefore the critical association concentration but dramatic afterwards;thus unimeric surfactant does not give any deswelling while self-assembled surfactant has a very large effect. Therefore, the collapseincreases strongly with surfactant chain length and can be attributedto differences in the concentration of the free surfactant, which can actas a screening electrolyte. The degree of binding is also higher for asurfactant with a longer hydrophobic tail, because of larger micelles.The surfactant-induced deswelling is larger for ss-DNA, than for ds-DNA gels, demonstrating the role of hydrophobic interactions. Thekinetics of surfactant-induced collapse shows a complex pattern withan initial deswelling and subsequently a reswelling back to theoriginal volume. This behaviour is interpreted in terms of chargereversal and long-time changes in the liquid crystalline structure ofthe DNA–surfactant aggregates. Surfactant-induced deswelling of theDNA gels under some conditions appears to be quite homogeneous,while in others there is a separation into a collapsed region in theouter parts of the gel sample and an inside swollen part. Such a “skin”formation is quite different for ss- and ds-DNA, ss-DNA giving a morepronounced skin formation over a wider range of binding ratio, β.
Acknowledgments
We are grateful to Fundação para a Ciência e a Tecnologia (FCT)(SFRH/BD/16736/2004), the EU Research Training Network, CIPSNAC(contract no. MRTN-CT-2003-504932), FEDER-POCTI/QUI and to theSwedish Science Research Council (VR) for financial support. Theauthors also thank Håkan Wennerström and Per Hansson for helpfuldiscussions. Particular thanks are due to Lennart Piculell for manyimportant comments on this work and for helpful suggestions.
References
[1] Kokufuta E, Suzuki H, Yoshida R, Yamada K, Hirata M, Kaneko F. Langmuir1998;14:788–95.
[2] Lee W, Hsu CJ. App Polym Sci 1999;74:3242–53.[3] Eichenbaum GM, Kiser PF, Shah D, Meuer WP, Needham D, Simon SA.
Macromolecules 2000;33:4087–93.[4] Annaka M, Ogata Y, Nakahira TJ. Phys Chem B 2000;104:6755–60.[5] Zhang J, Peppas NA. Macromolecules 2000;33:102–7.[6] Zhao B, Moore JS. Langmuir 2001;17:4758–63.[7] Sjöström J, Piculell L. Langmuir 2001;17:3836–43.[8] Sayil C, Okay OJ. App Polym Sci 2002;83:1228–32.[9] Kiser PF, Wilson G, Needham D. Nature 1998;394:459–62.[10] Akala EO, Kopeckova P, Kopecek J. Biomaterial 1998;19:1037–47.[11] Torres-Lugo M, Peppas NA. Macromolecules 1999;32:6646–51.[12] Tanaka, Hydrogels T. In: Mark HF, Kroschwitz JI, editors. Encyclopedia of polymer
science and technology. New York: Wiley; 1985. p. 514–31.[13] Horkay F, Tasaki I, Basser PJ. Biomacrom 2000;1:84–90.[14] Yodhida R, Uchida K, Kaneko Y, Sakai K, Kikuchi A, Sakurai Y, Okano T. Nature
1995;374:240–2.[15] Gotoh T, Nakatani Y, Sakohara SJJ. Appl Polym Sci 1998;69:895–906.[16] Lee KK, Cussler EL, Marchetti M. Chem Eng Sci 1990;45:766–7.[17] Tanaka T. Phys Rev Lett 1978;40:820–3.[18] Sjöström J, Piculell L. Colloids Surf A 2001;429:183–5.[19] Mironov AV, Starodoubtsev SG, Khokhlov AR. Macromolecules 1998;31:7698–705.[20] Khokhlov AR, Kramarenko EYu, Makhaeva EE, Starodoubtsev SG. Macromol
Chem Theory Simul 1992;1:105–18.[21] Philippova OE, Hourdet D, Audebert R, Khokhlov AR. Macromolecules
1996;29:2822–30.[22] Starodoubtsev SG, Churochkina NA, Khokhlov AR. Langmuir 2000;16:1529–34.[23] Bronstein LM, Platonova OA, Yakunin AN, Yanovskaya IM, Valetsky PM. Langmuir
1998;14:252–9.[24] Khokhlov AR, Kramarenko EYu, Makhaeva EE, Starodoubtsev SG. Macromole-
cules 1992;25:4779–83.[25] Philippova OE, Starodoubtsev SGJ. Polym Sci B: Polymer Physics 1993;31:1471–6.[26] Philippova OE, Chtcheglova LA, Karybiants NS, Khokhlov AR. Polymer Gels and
Networks 1998;6:409–21.

31D. Costa et al. / Advances in Colloid and Interface Science 158 (2010) 21–31
[27] Starodoubtsev SG, Dembo AT, Dembo KA. Langmuir 2004;20:6599–604.[28] Irie M. Adv Polym Sci 1990;94:28–67.[29] Gehrke SH, Lee PI. Hydrogels for drug delivery systems. In: Tyle P, editor.
Specialized drug delivery systems. New York: Marcel Dekker; 1990. p. 333–92.[30] Eichenbaum GM, Kiser PF, Dobrynin AV, Simon SA, Needham D. Macromolecules
1999;32:4867–78.[31] Kazanskii KS, Dubrovskii SA. Adv Polym Sci 1992;97:104–9.[32] Takezawa T, Mori Y, Yoshizato K. Biotechnology 1990;8:854–6.[33] Liu F, Tao GL, Zhuo RX. Polym J 1993;25:561–9.[34] Okano T, Kikuchi A, Sakurai Y, Takei Y, Ogata NJ. Controlled Release 1995;36:125–233.[35] Khokhlov AR, Starodubtzev SG, Vasilevskaya VV. Conformational transitions in
polymer gels: theory and experiment. In: Dušek K, editor. Responsive gels:volume transitions 1, volume 109 of Adv. Polym. Sci. New York: Springer-Verlag;1993. p. 123–72.
[36] Barrat JL, Joanny JF. In: Prigogine I, Rice SA, editors. Advances in chemical physics.New York: John Wiley; 1997. 1, 94.
[37] Hill TL. An introduction to statistical thermodynamics. New York: Dover; 1986.[38] Wilder J, Vilgis TA. Phys Rev E 1998;57:6865–74.[39] Vilgis TA, Wilder J. Comp Theor Polym Sci 1998;8:61–73.[40] Frusawa H, Hayakawa R. Phys Rev E 1998;58:6145–54.[41] Somia N, Verma IM. Nature 1997;389:239–42.[42] Shirahama K, Takashima K, Takisawa N. Bull Chem Soc Jpn 1987;60:43–7.[43] Gorelov AV, McLoughlin DM, Jacquier JC, Dawson KA. Nuovo Cimento
1998;20:2553–7.[44] Dias R, Mel`nikov S, Lindman B, Miguel M. Langmuir 2000;16:9577–83.[45] Koltover I, Salditt T, Rädler JO, Safinya CR. Science 1998;281:78–81.[46] Kokufuta E, Suzuki H, Yoshida R, Kaneko F, Yamada K, Hirata M. Colloids Surf A
1999;147:179–87.[47] Hansson P, Schneider S, Lindman B. Prog Colloid Polym Sci 2000;115:342–6.[48] Hansson P, Schneider S, Lindman BJ. Phys Chem B 2002;106:9777–93.[49] Costa D, Hansson P, Schneider S, Miguel M, Lindman B. Biomacr 2006;7:1090–5.[50] Khandurina YV, Rogacheva VB, Zezin AB, Kabanov VA. Polym Sci 1994;36:184–9.[51] Karabanova VB, Rogacheva VB, Zezin AB, Kabanov VA. Polym Sci 1995;37:1138–44.[52] Zezin AB, Rogacheva VB, Kabanov VA. Macromol Symp 1997;126:123–8.[53] Costa D, Santos S, Antunes F, Miguel M, Lindman B. Arkivoc 2006, (iv), 161–172.[54] Costa D, Miguel MG, Lindman BJ. Phys Chem B 2007;111:8444–52.[55] Costa D, Miguel MG, Lindman B. Phys Chem B 2007;111:10886–96.[56] Lindman B, Móran C, Costa D, Dias R, Miguel M. Polym 2007;56(1):49–50.[57] Costa D, Reischl M, Kuzma B, Brumen M, Zerovnik J, Ribitsch V, Miguel M,
Lindman BJ. Disp Sci Tech 2009;30:16.[58] Costa D, Valente A, Pais A, Miguel M, Lindman B. submitted.[59] Costa D. Covalent DNA Gels. Synthesis, Swelling and Release Properties. Doctoral
Thesis. Coimbra University, 2008.[60] Amiya T, Tanaka T. Macromolecules 1987;20:1162–5.[61] Horkay F, Basser P. Biomacr 2004;5:232–7.[62] Topuz F, Okay O. Macromolecules 2008;41:8847–54.[63] Eriksson M, Kim SK, Sen S, Gräslund A, Jernström B, Nordén BJ. Am Chem Soc
1993;115:1639–41.[64] Beach L, Schweitzer C, Scaiano JC. Org Biomol Chem 2003;1:450–1.
[65] Ichimura S, Zama M, Fujita H. Biochim Biophys Acta 1971;240:485–95.[66] Rosén O, Sjöström J, Piculell L. Langmuir 1998;14:5795–9.[67] Nielsen PE, Jeppesen C, Buchardt O. FEBS Lett 1988;235:122–8.[68] Lown JW. In: Pullman B, Jortner J, editors. Molecular basis of specificity in nucleic
acid–drug interactions. Dordrecht: Kluwer Academic Publishers; 1990.[69] Kim S, Nordén B. FEBS Lett 1993;315:61–4.[70] Takano M, Ogata K, Kawauchi S, Satoh M, Komiyama J. Polym Gels Networks
1998;6:217–21.[71] Liu X, Tong Z, Hu O. Macromol 1995;28:3813–7.[72] Firestone BA, Siegel RAJ. Biomater Sci Polymer Edn 1994;5:433–8.[73] Hackl EV, Kornilova SV, Blagoi YP. Int J Biol Macrom 2005;35:175–91.[74] Stake I, Takahashi T, Hayakawa K, Maeda T, Aoyagi M. Bull Chem Soc Jpn
1990;63:926–8.[75] Hansson P, Almgren MJ. Phys Chem 1995;99:9201–9.[76] Thalberg K, Lindman B, Karlström GJ. Phys Chem 1990;94:4289–95.[77] Khokhlov AR, Kramarenko EYu. Macromolecules 1996;29:681–5.[78] Schneider S, Linse P. Macromolecules 2004;37:3850–6.[79] Holmberg K, Jönsson B, Kronberg B, Lindman B. Surfactants and polymers in
aqueous solution2nd Edition. West Sussex, U.K: John Wiley & Sons Ltd; 2003.[80] Nishiyama Y, Satoh MJ. Polym Sci B 2000;38:2791–800.[81] Hiroki A, Maekawa Y, Iwai Y, Arai Y. Colloid Polym Sci 1994;272:1313–6.[82] Gouda JH, Povodator K, Warren TC, Prins WJ. Polym Sci B 1970;98:225–30.[83] Dias R, Magno L, Valente A, Das P, Maiti S, Miguel M, Lindman BJ. Phys Chem B
2008;112:8824–31 29.[84] Lu Y, Weers B, Stellwagen NC. Biopolym 2002;61:261–75.[85] Jonsson M, Linse PJ. Chem Phys 2001;115:10975–85.[86] Sasaki S, Yamazoe Y, Maeda H. Langmuir 2000;16:7126–32.[87] Lynch I, Sjöström J, Piculell LJ. Phys Chem B 2005;109:4252–7.[88] Isogai N, Narita T, Chen L, Hirata M, Gong JP, Osada Y. Colloids and Surfaces A
1999;147:189–201.[89] Ohbu K, Hiraishi O, Kashiwa IJ. Am Oil Chem Soc 1982;59:108–13.[90] Sokolov EL, Yeh F, Khokhlov A, Chu B. Langmuir 1996;12:6229–34.[91] Lynch I, Sjöström J, Piculell LJ. Phys Chem B 2005;109:4258–62.[92] Travas-Sejdic J, Easteal AJ. Polym 2000;41:7451–5.[93] Kabra B, Gehrke S, Hwang S, Ritschel WJ. Appl Polym Sci 1991;42:2409–13.[94] English AE, Mafé S, Manzanares JA, Yu X, Yu Grosberg A, Tanaka TJ. Chem Phys
1996;104:8713–7.[95] Hansson P. Langmuir 1998;14:4059–64.[96] Kabanov VA, Zezin B, Rogacheva V, Khandurina Y, Novoskoltseva O. Macromol
Symp 1997;126:79–94.[97] Sekimoto K. Phys Rev Lett 1993;70:4154–7.[98] Tomari T, Doi M. Macromol 1995;28:8334–43.[99] Mironov AV, Starodoubtsev SG, Khokhlov AR, Dembo AT, Yakunin AN. Colloids
and Surfaces A 1999;147:213–20.[100] Zhou S, Yeh F, Burger C, Hu H, Liu T, Cho B. Polym Adr Technol 2000;11:235–41.[101] Dembo AT, Yakunin AN, Zaitsev VS, Mironov AV, Starodubtsev SG, Khokhlov AR,
Chu BJ. Polym Sci Phys 1996;34:2893–902.[102] Cardenas M, Braem A, Nylander T, Lindman B. Langmuir 2003;19:7712–8.