symbiotic nitrogen fixation by nifa deletion mutant …utilized bythe n2-fixingbacteria are...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, May 1993, p. 2662-2673 Vol. 175, No. 90021-9193/93/092662-12$02.00/0Copyright © 1993, American Society for Microbiology
Symbiotic Nitrogen Fixation by a nifA Deletion Mutant ofRhizobium meliloti: the Role of an Unusual ntrC AlleleMONIKA LABES,1 VIPIN RASTOGI,2 ROBERT WATSON,2 AND TURLOUGH M. FINANl*
McMaster University, Department ofBiology, 1280 Main Street West, Hamilton, Ontario, L8S 4KI, 1 andPlant Research Centre, Agriculture Canada, Ottawa, Ontario, KJA 0C6,2 Canada
Received 30 October 1992/Accepted 1 March 1993
In the N2-fixing alfalfa symbiont Rhizobium meliloti, the three r,4 (NTRA)-dependent positively actingregulatory proteins NIFA, NTRC, and DCTD are required for activation of promoters involved in N2 fixation(pnifHDKE and pfixABCX), nitrogen assimilation (pglnII), and C4-dicarboxylate transport (pdct4), respec-tively. Here, we describe an allele of ntrC which results in the constitutive activation of the above NTRC-,NIFA-, and DCTD-regulated promoters. The expression and activation of wild-type NTRC occur in responseto nitrogen availability, whereas in cells carrying the ntrC283 allele, the NTRC283 protein appearsconstitutively active and is constitutively expressed. The ntrC283 allele was shown to carry a single mutationresulting in the replacement of an Asp by a Tyr residue in the helix-turn-helix motif of ntrC283. Introductionof the ntrC283 allele into a nifA deletion mutant restores the N2-fixation ability to 70 to 80%o of the wild-typelevel. Thus, the nifA gene is dispensable for symbiotic N2 fixation.
Nitrogen-fixing root nodules develop through an intimateinteraction between bacteria (rhizobia) and plant cells. Inone of the early steps in this interaction, flavonoids producedby the plant induce the expression of a set of nodulation(nod) genes in the bacteria (31, 40, 43). In Rhizobium melilotiand Rhizobium leguminosarum, which induce root noduleson alfalfa and pea plants, respectively, the nod gene prod-ucts function in the synthesis of a lipooligosaccharide signalmolecule which triggers root nodule formation (62). Thebacteria enter the root via tube-like structures, or infectionthreads, and are released into the plant cortical cells via anendocytosis-like process in which they are surrounded bythe host plasma membrane. Both the bacteria and plant cellsthen undergo many biochemical changes, one of which is theexpression of the plant globin genes whose products areinvolved in oxygen transport in the nodule. In the bacteria,the nitrogenase (nif) and other genes required for N2 fixationare induced. Expression of many nif genes and associatedgenes in nodules is regulated positively by the nifA geneproduct. Thus, nifA null mutants show no symbiotic N2-fixation activity. Expression of nifA is, in turn, regulated inresponse to oxygen concentration by a two-componentenvironmental sensor-regulator pair, fixL and fixi, and byfixK (4, 11, 12). Recent data have shown that, within anodule, the expression of the nifgenes in the bacteria and theglobin genes in the plant cells occurs synchronously within afew layers and thus shows a high level of control (10, 69).The N2-fixing bacteria within nodules (called bacteroids)
reduce N2 gas to ammonia which is then supplied to theplant. Because the bacteroids are within the plant cells, theyare totally dependent on the plant for a source of energy-reduced carbon, which is necessary for the high-energy-requiring N2-fixation reaction. The supply of a carbon sourcethus potentially represents a key point in control of N2fixation and bacterial proliferation in root nodules. Geneticand biochemical data suggest that the major carbon sourcesutilized by the N2-fixing bacteria are C4-dicarboxylates suchas succinate and malate (1, 7, 20, 47, 66).
* Corresponding author.
In R. meliloti and R. leguminosarum, transport of C4-dicarboxylates appears to occur via a single carrier proteinencoded by the dctA gene (16, 45, 70). Expression of dctA ispositively regulated by a two-component sensor-regulatorgene pair, dctB and dctD, which are constitutively expressedat a low level (45, 46, 65). The dctB and dctD genes showhomology to members of the ntrB and ntrC gene families (38,46, 49), and a model for regulation of dctA expression hasbeen proposed in which the membrane-located DCTB pro-tein senses C4-dicarboxylates and then modifies the cyto-plasmically located dctD gene product (DCTD) to render itsactive form. By analogy to NTRC, activation of DCTDwould occur by phosphorylation (DCTD-P) (46). DCTDupstream activator sequences (UAS) have been locatedbetween bp -96 and -156 upstream of the dctA transcrip-tional start site (22, 30). In addition, NTRC (23, 25, 38) andDCTD (48) belong to the family of transcriptional activatorswhose members require the alternative sigma factor cr54(NTRA) encoded by ntrA (29, 59). Such genes, including theNIFA-regulated nif and fix genes required for free-living andsymbiotic N2 fixation, respectively, are characterized by apromoter consensus sequence of GG-N1O-GC located 11 to14 bp in front of the transcriptional initiation site (8, 35, 36).Detailed studies on the activation of the major glutaminesynthetase promoter (glnA) in enteric bacteria have shownthat NTRC binds to enhancer-like sites located >100 bpfrom the transcriptional start site (44). The formation of aDNA loop allows the regulator to interact with a or54-RNA-polymerase complex at the promoter site (8, 41, 55, 67).NTRC-P stimulates transcription by catalyzing the ATP-dependent isomerization (2, 14, 68) of closed promotercomplexes to open ones, i.e., to a form in which the DNA islocally denatured around the transcriptional start site (41,52).Whereas dctA expression in free-living cells of R. meliloti
and R. leguminosarum is dependent on dctB and dctD, inroot nodules dctA is expressed in the absence of dctB anddctD (15, 45, 70). The existence of a symbiosis-specificactivator has thus been postulated (45, 46, 70). In search ofsuch an alternative activator of dctA transcription, wescreened Dct' revertants obtained from a dctD::TnS mutant
2662
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

ACTIVATION OF ,54-DEPENDENT PROMOTERS 2663
TABLE 1. Bacterial strains, plasmids, and phages
Strain, plasmid,or phage Relevant genotype and characteristic(s) Source or reference
F- end41 hsdRl7 (rk- Mk-) supE44 thi-1 recAl gyrA96 relAl A(argF-lacZYA) U169 080dlacZ AM15 X-
recA::Tn7, integrated RP4 derivative
SU47 str-21nifA239::TnSntrC::TnSntrA75::TnSRCR2011 AnifA, ca. 500 bp of the central portion of nifA were deletedby Bal 31 treatment
dctDl6::TnSntrA74::TnS-233dctDl6::TnS ntrC283dctDl6::TnS ntrC283 fI5199::TnS-233dctDl6::TnS-233ntrC283 il 5199::TnS-233dctDl6::Tn5-Tp ntrC283RCR2011 AnifA ntrC283 fI5199::TnS-233RCR2011 AnifA dctDl6::TnS ntrC283 05199::TnS-233dctDl6::TnS ntrC283::fISp, homogenotedctDl6::TnS ntrC283 fISp downstream of ntrC283, homogenote
Bethesda ResearchLaboratory
54
1758571928
7070This workThis workThis workThis workThis workThis workThis workThis workThis work
PlasmidspSUP202pJel2a5pPH45pRKM1pRKP9pMB211pRS21-1pRD578pRKRD578pJJ5pHLRLAZ-FpHLRLAZ-TpHLRLAZ-VpRKRMAZ+UASpRKRMAZ-UASpTH98pTH99pTH100pVR60pVR61
Phage, M12
Mobilizable derivative of pBR325 (Tcr)3.8-kb EcoRI fragment in pACYC184 containing ntrCRmpBR322 fISprpRK290, 'lacZpRK290, nifIHR-::'lacZpRK290, fixARm::'lacZpRK290, glnIIRm::'lacZpBR322, xyFPp::'lacZCointegrate of pRD578 and pRK290, fusion in EcoRIpRK290, fixKRm::'lacZpRK290, pdctARl::'lacZpRK290, pdctARl::'lacZpRK290, A-derivative of pdctARI::'lacZpRK290, pdctAR::'lacZpRK290, pdCtARm::'lacZpSUP202, PstI fragment carrying 'ntrC::lSprpSUP202, PstI fragment carrying 'ntrC, flSpr downstream of ntrCpSUP202, PstI fragment carrying 'ntrC283 flSpr downstream of ntrC283pUC19, 2.3-kb EcoRI-BglII fragment carrying ntrC from RmF121pUC19, 2.3-kb EcoRI-BglII fragment carrying ntrC283
Transducing R. meliloti phage
for second site mutations which allowed activation of detA,i.e., grew on minimal medium containing succinate as a solecarbon source. In this report, we describe the identificationof an unusual allele of ntrC, designated ntrC283, whichresults in the constitutive activation of the dctA and otheru54-dependent promoters. In addition, we report thatntrC283 can functionally replace the requirement of nifA forsymbiotic N2 fixation.
MATERIALS AND METHODS
Bacterial strains, plasmids, and media. The bacterialstrains and plasmids used in this study are listed in Table 1.Complex medium (LBMC) and defined media (M9 andNFDM), antibiotic concentrations, and routine growth con-ditions were as described previously (18, 34, 57).
Genetic techniques. Transductions, plasmid conjugation,and homogenotizations were performed as described previ-ously (17, 54). Dct+ revertants of RmF121 were isolated byspreading washed cells (3 x 108) on M9 medium containingsuccinate. Colonies which grew were single-colony purifiedthree times on succinate prior to analysis. Gene disruption inR. meliloti was performed by first cloning the selected DNAfragment carrying the fkSpr interposon into the mobilizableplasmid pSUP202 in Escherichia coli S17.1. After mating ofthis strain with R. meliloti, Spr transconjugants were se-lected and homogenotes (double crossover) were distin-guished from heterogenotes (single crossover) by screeningthe colonies for tetracycline sensitivity.
Insertion fQ5199 was identified as a Gmr/Spr gentamicin-/spectinomycin-resistant) Dct- Nmr (neomycin-resistant)colony obtained after transduction of random TnS-233 inser-
StrainsE. coliDH5a
S17-1
R. melilotiRmlO21Rm1354Rm5003Rm5422Rm-d8
RmF121RmF250RmF283RmF781RmF863RmG649RmG693RmG711RmG712RmG733RmG855
54T. Nixon (57)425757S5313This work4T. Nixon (30)T. Nixon (30)T. Nixon (30)T. Nixon (21)T. Nixon (21)This workThis workThis workThis workThis work
17
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2664 LABES ET AL.
tions from RmlO21 (Dct') into strain RmF283 (Dct') aspreviously described (70). In subsequent transduction ofrandom Gmr/Spr from this strain into RmF283, 52% of thetransductants failed to grow on succinate (Dct-). Thus, thef15199:TnS-233 insertion is 52% linked to the dctD suppres-sor mutation. One of the above Gmr/Spr Dct+ Nmr transcon-jugants was purified and designated RmF781. This strain wasutilized to transfer the ntrC283 allele into other geneticbackgrounds via its linkage to Gmr/Spr (f15199:Tn5-233).DNA manipulations. Standard methodologies were used
for DNA preparations, agarose gel electrophoresis restric-tion endonuclease digestions, ligations, transformations,random primer labelling, and hybridization (50).RNA biochemistry. Cultures (200 ml) were grown in
NFDM medium with 0.2% (+N) or 0.01% (-N) L-glutamineas a sole nitrogen source, centrifuged at 8,000 rpm for 10 minin a Beckman GA-10 fixed-angle rotor, and resuspended in 3ml of lysis buffer (0.1 M Tris, 5 mM Na-citrate, 1% sarcosyl,0.1 M 3-mercaptoethanol [pH 8.0]). Cells were frozen inliquid nitrogen for 2 min and disrupted by ultrasound. Toextract the RNA, the lysate (mixed with 5 ml of CsCl in lysisbuffer-3 M [final concentration] CsCl) was pipetted onto 3.5ml of 5.7 M CsCl and 0.1 M EDTA (pH 8.0) in BeckmanQuickseal tubes (16 by 76 mm). The tubes were centrifugedfor 14 to 20 h at 30,000 rpm and 19.5°C in an NVT 65 rotor.The RNA pellet was collected by carefully removing thesupernatant and resuspending the pellet in 0.5 ml of 80%ethanol. The RNA was transferred to a microtube andwashed twice in 80% ethanol, and the pellet was dried andresuspended in 100 to 500 ,ul of Tris-EDTA buffer, dependingon the RNA yield. If necessary, the samples were treatedwith RNAse-free DNAse. (The protocol was kindly pro-vided by G. Labes.)
Dot blot analysis. Ten and 5 ,ug of RNA were dissolved in7.4% formamide-6x SSC (lx SSC is 0.15 M NaCl plus 0.015M sodium citrate), denatured for 15 min at 70°C, and vacuumblotted onto a nylon filter (ICN Biotrans). The RNA wasfixed to the filter by incubating it at 80°C for 2 h. Onemicrogram of EcoRI-digested total DNA of RmlO21 and 100ng of R. meliloti ntrC DNA (internal XhoI fragment) werelabelled with [32P]dCTP, and hybridization was carried outfor 24 h at 42°C. After being washed twice for 10 min in 2xSSC-0.1% sodium dodecyl sulfate (SDS) and once for 10 minin 0.2x SSC-0.1% SDS, filters were exposed for 6 to 24 h at-700C.
IP-Galactosidase assays and plant tests. Plant tests werecarried out as described previously (70). 1-Galactosidaseactivities on independently grown triplicate cultures weremeasured. Cultures were grown overnight in 5 ml of appro-priate medium. Cultures (0.5 to 1 ml) were centrifuged, thecells were resuspended in 2 ml of Z buffer (34), and theoptical densities at 600 nm (OD6.) were determined. Twodrops of both 0.1% SDS and chloroform were added, and theculture was vigorously mixed and left at room temperaturefor 5 min. After addition of 1 ml of o-nitrophenyl-13-D-galactopyranoside (0.8 mg/ml) and incubation at 37°C for 5 to45 min (depending on the actual activities), the reaction wasterminated by adding 1 ml of 1 M Na2CO3. The mixture wascentrifuged, and the OD420 was determined. Activities werecalculated according to the following relationship:
,u = (1,000 x 2 x OD420)/(t x OD6M)
in which Ou is the number of ,B-galactosidase units and tis time in minutes. The factor 2 accounts for the dilution
by addition of o-nitrophenyl-,B-D-galactopyranoside and Na2CO3.
Histochemical staining of P-galactosidase in gluteralde-hyde-fixed nodules was done with 5-bromo-4-chloro-3-in-dolyl-o3-D-galactopyranoside as described previously (6).Clearing nodules with hypochlorite was done according tothe method of Truchet et al. (61).DNA sequencing. Nucleotide sequencing was carried out
by the dideoxynucleotide sequencing method (51) with dou-ble-stranded DNA and synthetic primers as described pre-viously (65). Primers were synthesized with a Biosearch8750 DNA synthesizer.
RESULTS
Second site mutation in RmF283 suppresses dctD::TnSmutation in RmF121. R. meliloti RmF121 carries a Tn5insertion (Nmr) in the dctD gene and fails to transportsuccinate and grow on medium containing succinate as a solecarbon source (Dct-) (19, 70). In search of alternativeactivators of dctA transcription, we isolated Dct+ revertantsof strain RmF121 and further characterized those whichretained the Nmr determinant of the dctD::TnS insertion. Inparticular, transduction of Nmr from revertant RmF283 intothe wild-type RmlO21 resulted in 100% cotransduction of theDct- (dtD::TnS) phenotype. Moreover, transductants ob-tained by transducing the ntrA 74::Tn5-233 allele or thedctA::TnS-233 allele into RmF283 failed to grow on succi-nate. Together, these results suggested that RmF283 carrieda second site mutation which suppressed the Dct- pheno-type of the dctDJ6::TnS insertion, apparently by activatingdetA transcription in a or4-dependent manner.
Suppressor allele is linked to ntrC. To manipulate and mapthe suppressor mutation, we isolated a Tn5-233 insertion,fQ5199, which was 52% linked by cotransduction to thesuppressor allele (see Materials and Methods, Genetic tech-niques). In order to determine whether the suppressor mu-tation might be an allele of one of the known as -dependenttwo-component regulatory genes (38), we checked for link-age of the suppressor locus to the ntrB-ntrC gene region inR meliloti (57). After transduction of fQ5199::TnS-233 intothe ntrC::TnS mutant Rm5OO3, we found that 54% of thetransductants were Nms. In addition, transduction of ntrC::Tn5 into an RmF283 derivative in which the dctDl6::TnSinsertion was replaced by a trimethoprim-resistant TnS-Tpderivative (strain RmG693) resulted in 100% loss of theability to grow on succinate. These results suggested a tightlinkage between the suppressor mutation and ntrC. Moredefinite evidence was obtained by generating site-directedntrC: :flSpr (spectinomycin-resistant) RmF283 mutants.These were identified after the construction and transfer ofpTH98 (Fig. 1). Of 12 homogenotes examined (representedby RmG733), all had lost the ability to grow on mediumcontaining succinate as a carbon source and thus reexhibitedthe Dct- phenotype of the dctDl6::Tn5 mutation. We des-ignated the suppressor allele ntrC283.
Activation of dcti4 by the suppressor is constitutive. In orderto compare the levels of activation of the dctA promoter bythe wild-type DCTD protein with those achieved by thesuppressor molecule, lac fusions of R. meliloti (lines 1 and 2in Table 2) and R. leguminosarum dctA promoters (lines 3and 4 in Table 2) were transferred into strains RmlO21(dctD+), RmF283 (dctD mutant ntrC283), and RmF121(dctD mutant) and corresponding ,-galactosidase activitieswere determined (Table 2). In contrast to RmlO21, in thesuppressor strain both the complete R. leguminosarum and
J. BACTERIOL.
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

ACTIVATION OF o54-DEPENDENT PROMOTERS 2665
1 kb
'ntrBpJel2a5
ntrC
. .II II V I I I
E X P XEVI I
Bg X P
pACYC184
E
pTH98 -K----- pSUP2O2
Sm Sp/Sm Sm
FIG. 1. Inactivation of the ntrC gene by insertion of an Li fragment. The fl fragment LiSpr (spectinomycin resistance) from pPH45 was
cloned as a SmaI fragment into the EcoRV site of ntrC in pJel2a5. Subsequently, the internal PstI fragment marked by Spr was ligated intothe mobilizable E. coli plasmid pSUP202, rendering pTH98. This plasmid was used to generate homogenotes by transferring it into RmF283and screening for loss of the plasmid-encoded tetracycline resistance. E, EcoRI; X, XhoI; P, PstI; EV, EcoRV; Bg, BglII; Sm, SmaI.
the completeR meliloti fusions were induced in succinate as
well as in glucose (Table 2, rows 1 and 3). RmF283 activatesthese fusions in glucose to about 70% of the fully inducedlevel. Thus, in contrast to the wild type, activation of dctAtranscription in the suppressor strain RmF283 was constitu-tive. Both of these promoters showed relatively high back-ground activities in the wild type when grown in glucose. Nosignificant activities in the dctD::TnS background (RmF121)were measured because both promoters require DC-D foractivation.
Transcription of ntrC283 is constitutive. Evidence thatntrC283 is expressed constitutively in RmF283 was obtainedfrom RNA dot blot analysis (Fig. 2). RNA was extractedfrom RmlO21 and RmF283 grown in NFDM medium con-taining 0.2% (+N) and 0.01% (-N) L-glutamine, respec-tively. Ten and 5 pg ofRNA were probed with a 32P-labelledinternal DNA fragment (XhoI) of ntrC and, as a positivecontrol, EcoRI-digested and 32P-labelled RmlO21 totalDNA. Transcription of wild-type ntrC was only detected incells grown under nitrogen-limiting conditions, as previouslyreported (57). In contrast, in RmF283, ntrC283 transcriptionwas readily detected under nitrogen-limiting as well asnitrogen-excess conditions (Fig. 2). The constitutive expres-sion is consistent with a previous suggestion (57) that ntrC isautoregulated in R meliloti.
Activation ofginII expression by NTRC283 is not regulatedby nitrogen availability. To determine whether or not activa-tion by NTRC283 was still regulated by nitrogen availability,a glnII::lac fusion, which is normally regulated by ntrC (53),was introduced into the wild-type strain RmlO21, the dctDsuppressor strain RmF283 (dctDl6::TnS ntrC283), and thedctD and ntrC mutant strains RmF121 and Rm5003, respec-tively. The strains were cultured in NFDM medium with0.01% (N limitation) and 0.2% (N excess) L-glutamine andassayed for ,3-galactosidase activity (Table 3). The resultsshowed that ginII expression in the wild type was regulatedby nitrogen availability; however, expression in RmF283was constitutive. Because ginII expression in the dctD::TnSmutant strain RmF121 shows the same pattern of inductionas that in the wild-type Rm1021, we conclude that theconstitutive expression observed in RmF283 results from thentrC283 mutation.NTRC283 activates or"-dependent promoters. To investi-
gate the specificity of NTRC283-mediated promoter activa-tion, plasmids bearing translational lacZ fusions to severalu54-dependent promoters were transferred into RmlO21,RmF283, RmF121, Rm5003, RmG733, and Rm5422 (Table4). Strain RmG733 is a site-directed ntrC283::ftSpr insertionmutant of RmF283, and strain Rm5422 is an rpoN (ntrA::TnS) insertion mutant. Of the promoters tested, pnijHtm
TABLE 2. Activation of dctA::lacZ fusions by DCID and the suppressor moleculea
Activation (Ou + SD) in:
Plasmid Promoter RmlO21 RmF283 RmF121
Glucose Succinate Glucose Succinate Glucose
pRKRMAZ+UAS pdct,,Rm 1,008 + 130 16,104 ± 263 8,194 ± 194 11,677 ± 1,282 110 ± 14PRKRMAZ-UAS pdctARm 76 + 13 217 ± 6 1,356 ± 92 1,177 + 162 75 ± 10pHLRLAZ-T pdctAX 1,258 ± 265 17,829 + 3,499 10,118 ± 969 8,976 ± 2,649 198 ± 50pHLRLAZ-V pdctAR, 203 ± 28 1,721 -+ 304 2,308 ± 557 2,210 + 223 244 ± 24pRKM1 None 68 22 72 ± 10 260 ± 127 94 16 60 ± 11No plasmid 72 12 140 ± 57 98 ± 21 124 39 47 ± 16
a ,3-Galactosidase activities of IacZ fusions to the dct4 promoters ofR meliloti (line 1, with complete UAS; line 2, without UAS) andR leguminosarum (line3, with complete UAS; line 4, without UAS) in different genetic backgrounds (Rm1021, dctD+ ntrC+; RmF283, dctD::TnS ntrC283 [suppressor+]; RmF121,dctD::TnS) and under different growth conditions are shown. Values are mean f-galactosidase units (13u) from three independent cultures ± the standarddeviation.
r- v
IIr
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2666 LABES ET AL.
fi_ j_D G_09 S
>U: i:
CX a, 70F-00 0f ,lS, C:s,t- 7f'vX wwS -tr:= _ ^p .:?- Li_
IOg 5sg lOyg 5AgFIG. 2. Transcription of ntrC in RmlO21 (wild type) and RmF283
(dctDJ6::TnS ntrC283). Ten and 5 ,ug of RNA extracted fromRmlO21 and RmF283 grown under N-excess (+N) and N-limiting(-N) conditions were vacuum blotted onto a nylon filter. Lanes 1and 2 show RNA probed with 32P-labelled ntrC DNA (internal XhoIfragment); lanes 3 and 4 show RNA dots probed with 32P-labelledRmlO21 total DNA (EcoRI digested).
(pl) and PfiX4Rm (p2) are symbiotically regulated by NIFAand are not normally activated under free-living conditions(5; but see Discussion); pglnIIRm and the pdctARm promot-ers are regulated by NTRC and DCTD, respectively (30, 53).The Pseudomonas putida pxyL4 (pxyLpp) and the R. legu-minosarum pdctA (pdctARm) promoters are normally acti-vated by XYLR and DCTD, respectively (13, 30). ThepfixKRm promoter, which does not require e4 for activation,and the promoterless lacZ plasmid pRKM1 served as con-
trols. The 3-galactosidase activities were determined and are
listed in Table 4. The pnifH and pfix4 fusions were activated26- and 20-fold, respectively, in the suppressor (RmF283)background. The glnHI and the xylA promoters showed 10-and 2-fold higher expression in RmF283 than in RmlO21,respectively. Activation of pdctARm, pdctAR1, pnifHfRm, andPfixARm promoters in the suppressor background was re-
duced to background levels upon introduction of ntrC283::flSpr (strain RmG733). Together, these data suggested thateither NTRC283 itself or an NTRC283-activated moleculeled to the constitutive activation at diverse c54-dependentpromoters.The DCTD UAS are not essential for activation by
NTRC283. The detA promoter of both R. meliloti and R.leguminosarum contains two DC1LD binding sites locatedbetween bp -96 and -156 from the transcriptional start site(30). Because these sites have been shown to play a majorrole in the activation of the dctA promoter by DCTD, we
wished to determine whether these UAS were required foractivation by NTRC283 (30). Plasmids carrying translationallacZ fusions to either the intact dctA promoter or deletionderivatives in which the UAS were removed were trans-ferred into RmlO21 (wild type), RmF283 (dctDl6: :Tn5ntrC283), and RmF121 (dctDJ6::TnS). The wild-type R.meliloti promoter region extended from bp +115 to -177,whereas the UAS deletion extended from bp +115 to -81(21). The wild-type R. leguminosarum promoter regionextended from bp +113 to -239, and the UAS deletionextended from bp +113 to -37 (30). The activities of thesefusions (Table 2, lines 1 to 4) showed that the promoterfusions lacking the UAS (Table 2, lines 2 and 4) wereexpressed 10-fold higher in glucose-grown RmF283 cellsthan in glucose-grown RmlO21 cells. The UAS were thus notrequired for activation of transcription. However, the pres-ence of the UAS resulted in a further approximately 7- to10-fold activation (Table 2, lines 1 and 3).NTRC283 restored the symbiotic nitrogen-fixation ability of
a nifA deletion mutant. Because the NTRC283 mutation ledto high-level expression of the normally NIFA-regulatednifIH and fixA4 promoters (Table 4), we investigated whetherntrC283 could suppress the symbiotic Fix- phenotype ofnifA mutants. Because nifA transposon insertions are polaron nifB and fdxN transcription (28), we tested for suppres-sion of a nifA deletion mutation in strain Rm-d8 (28). Thiswell-characterized deletion removes ca. 500 bp from thecentral region of nifA without affecting nifB and fdxNexpression. The ntrC283 allele was transduced into Rm-d8via the linked insertion Q5199::TnS-233. Transductantscarrying ntrC283 were identified by their ability to growon succinate minimal medium after the transfer of thedctDJ6::TnS insertion. The symbiotic phenotypes of theresulting strains are shown in Table 5. As expected, the nifAdeletion mutant Rm-d8 showed no N2-fixing activity. How-ever, constructs carrying the nifA deletion and ntrC283, withand without dctDJ6::TnS (strains RmG711 and RmG712,respectively), showed N2-fixing activity 70 to 90% of that ofthe wild-type RmlO21. Bacteria isolated from surface-steril-ized nodules of plants inoculated with RmG711 and RmG712had the expected antibiotic markers. We therefore concludethat ntrC283 can functionally replace nifA in alfalfa rootnodules. To address the question of whether NTRC couldserve as the symbiotic activator of dctA, we constructeddctD ntrC double mutants. We found that these strains hadN2-fixing abilities similar to that of the dctD mutant alone(data not shown). We thus conclude that NTRC does not actas a symbiotic activator of dctA.NtrC283 alters the temporal and spatial expression of nijj
and fixA in nodules. The symbiotic effect of the ntrC283mutation was further investigated by determining the in situ
TABLE 3. Effect of N availability on the activation of glnII::lacZ and dctA::lacZ fusions in different genetic backgrounds
Activation (1u + SD) in:
Strain Genotype glnII::lacZ dctA::lacZ
+N -N +N -N
Rm1021 Wild type 640 ± 156 6,120 ± 1,972 867 ± 75 963 ± 83RmF283 dctD::TnS ntrC283 10,307 ± 1,356 8,373 - 1,216 8,273 ± 1,929 8,658 ± 750Rm5003 ntrC::TnS 210 ± 2 264 ± 41 223 ± 21 253 ± 65RmF121 dctD::TnS 812 ± 125 6,262 - 1,897 748 ± 36 975 ± 139
a Nitrogen-excess (+N) and nitrogen-limiting (-N) conditions (57) were as previously described. Values are the mean P-galactosidase units (13u) from threeindependent cultures + the standard deviation.
32P RmlO213 4
32P ntrC1 2
RmlO2l +N
Rm1O21 - N
RmF283 +N
RmF283 - N
J. BACTERIOL.
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
ACTIVATION OF a54-DEPENDENT PROMOTERS 2667
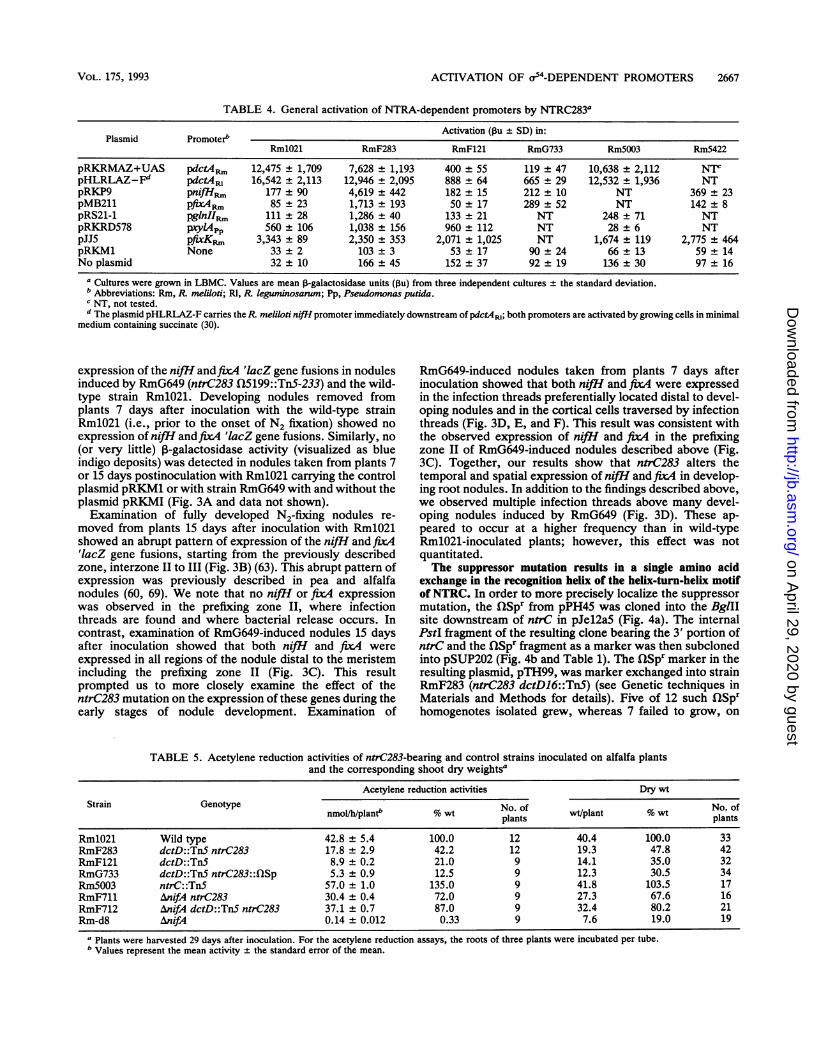
TABLE 4. General activation of NTRA-dependent promoters by NTRC2831
Plasmid PromoterbActivation (13u ± SD) in:
RmlO21 RmF283 RmF121 RmG733 Rm5003 Rm5422
pRKRMAZ+UAS pdctAR. 12,475 ± 1,709 7,628 ± 1,193 400 ± 55 119 ± 47 10,638 ± 2,112 NTCpHLRLAZ-Fd pdctA4, 16,542 ± 2,113 12,946 ± 2,095 888 ± 64 665 ± 29 12,532 ± 1,936 NTpRKP9 pnifHRI 177 ± 90 4,619 ± 442 182 ± 15 212 ± 10 NT 369 ± 23pMB211 pfix4Rm 85 ± 23 1,713 ± 193 50 ± 17 289 ± 52 NT 142 ± 8pRS21-1 pglnIIm, 111 ± 28 1,286 ± 40 133 ± 21 NT 248 ± 71 NTpRKRD578 pyL4PP 560 ± 106 1,038 ± 156 960 ± 112 NT 28 ± 6 NTpJJ5 pfixKp 3,343 ± 89 2,350 ± 353 2,071 ± 1,025 NT 1,674 ± 119 2,775 ± 464pRKM1 None 33 ± 2 103 ± 3 53 ± 17 90 ± 24 66 ± 13 59 ± 14No plasmid 32 ± 10 166 ± 45 152 ± 37 92 ± 19 136 ± 30 97 ± 16
a Cultures were grown in LBMC. Values are mean 13-galactosidase units (Pu) from three independent cultures ± the standard deviation.b Abbreviations: Rm, R. meliloti; RI, R leguminosarum; Pp. Pseudomonas putida.C NT, not tested.d The plasmid pHLRLAZ-F carries theR melioti nifI promoter immediately downstream of pdctA4R; both promoters are activated by growing cells in minimal
medium containing succinate (30).
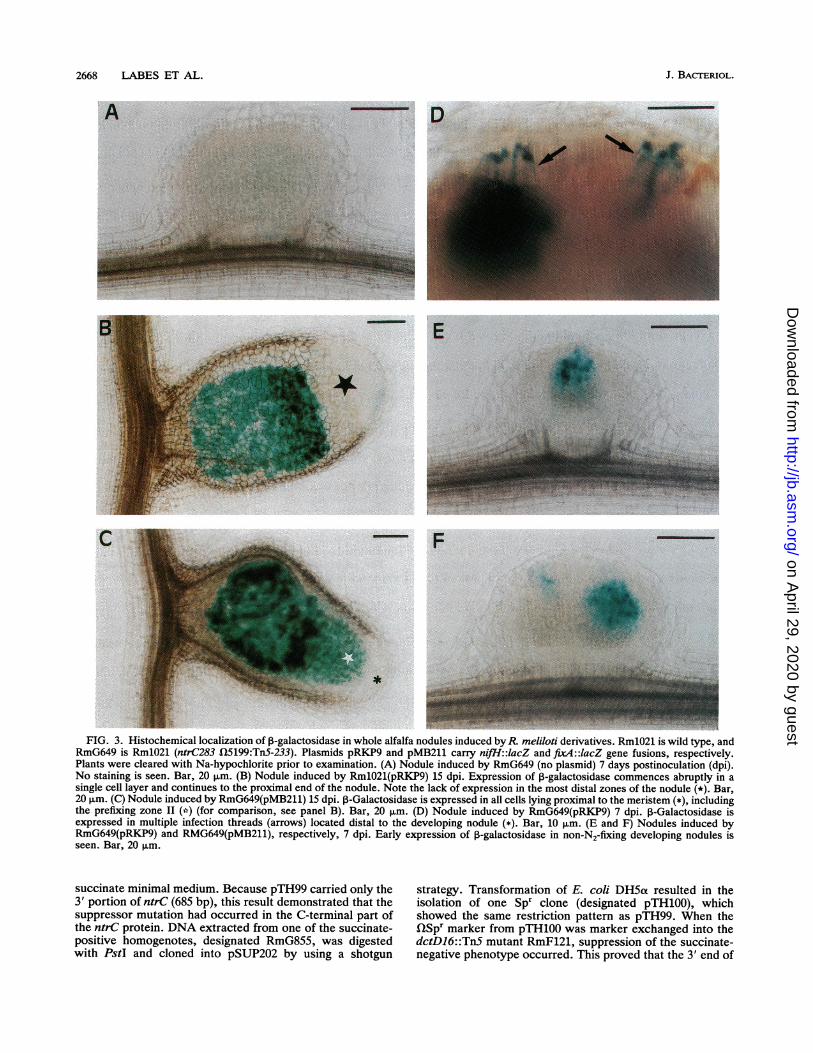
expression of the nifl andfix4 'lacZ gene fusions in nodulesinduced by RmG649 (ntrC283 Q5199::TnS-233) and the wild-type strain RmlO21. Developing nodules removed fromplants 7 days after inoculation with the wild-type strainRmlO21 (i.e., prior to the onset of N2 fixation) showed noexpression of nifH andfix4 'lacZ gene fusions. Similarly, no(or very little) P-galactosidase activity (visualized as blueindigo deposits) was detected in nodules taken from plants 7or 15 days postinoculation with RmlO21 carrying the controlplasmid pRKM1 or with strain RmG649 with and without theplasmid pRKMI (Fig. 3A and data not shown).Examination of fully developed N2-fixing nodules re-
moved from plants 15 days after inoculation with RmlO21showed an abrupt pattern of expression of the nifH and fixA'lacZ gene fusions, starting from the previously describedzone, interzone II to III (Fig. 3B) (63). This abrupt pattern ofexpression was previously described in pea and alfalfanodules (60, 69). We note that no nifH or fixA expressionwas observed in the prefixing zone II, where infectionthreads are found and where bacterial release occurs. Incontrast, examination of RmG649-induced nodules 15 daysafter inoculation showed that both nifH and fix4 wereexpressed in all regions of the nodule distal to the meristemincluding the prefixing zone II (Fig. 3C). This resultprompted us to more closely examine the effect of thentrC283 mutation on the expression of these genes during theearly stages of nodule development. Examination of
RmG649-induced nodules taken from plants 7 days afterinoculation showed that both nifH and fix4 were expressedin the infection threads preferentially located distal to devel-oping nodules and in the cortical cells traversed by infectionthreads (Fig. 3D, E, and F). This result was consistent withthe observed expression of nifHl and fixA in the prefixingzone II of RmG649-induced nodules described above (Fig.3C). Together, our results show that ntrC283 alters thetemporal and spatial expression of nifH and fix4 in develop-ing root nodules. In addition to the findings described above,we observed multiple infection threads above many devel-oping nodules induced by RmG649 (Fig. 3D). These ap-peared to occur at a higher frequency than in wild-typeRmlO21-inoculated plants; however, this effect was notquantitated.The suppressor mutation results in a single amino acid
exchange in the recognition helix of the helix-turn-helix motifof NTRC. In order to more precisely localize the suppressormutation, the flSpr from pPH45 was cloned into the BglIIsite downstream of ntrC in pJel2a5 (Fig. 4a). The internalPstI fragment of the resulting clone bearing the 3' portion ofntrC and the jlSpr fragment as a marker was then subclonedinto pSUP202 (Fig. 4b and Table 1). The flSpr marker in theresulting plasmid, pTH99, was marker exchanged into strainRmF283 (ntrC283 dctDl6::TnS) (see Genetic techniques inMaterials and Methods for details). Five of 12 such fISprhomogenotes isolated grew, whereas 7 failed to grow, on
TABLE 5. Acetylene reduction activities of ntrC283-bearing and control strains inoculated on alfalfa plantsand the corresponding shoot dry weights'
Acetylene reduction activities Dry wt
Strain Genotype nmolplant' % wt No. of w/plant % wt No. ofnmol/h/plant" ~~~~plants wtpat %'t plants
RmlO21 Wild type 42.8 ± 5.4 100.0 12 40.4 100.0 33RmF283 dctD::TnS ntrC283 17.8 ± 2.9 42.2 12 19.3 47.8 42RmF121 dctD::TnS 8.9 ± 0.2 21.0 9 14.1 35.0 32RmG733 dctD::TnS ntrC283::fQSp 5.3 ± 0.9 12.5 9 12.3 30.5 34RmS003 ntrC::TnS 57.0 ± 1.0 135.0 9 41.8 103.5 17RmF711 AnifA ntrC283 30.4 ± 0.4 72.0 9 27.3 67.6 16RmF712 AnifA dctD::TnS ntrC283 37.1 ± 0.7 87.0 9 32.4 80.2 21Rm-d8 AnifA 0.14 ± 0.012 0.33 9 7.6 19.0 19
aPlants were harvested 29 days after inoculation. For the acetylene reduction assays, the roots of three plants were incubated per tube.b Values represent the mean activity ± the standard error of the mean.
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
2668 LABES ET AL.
FIG. 3. Histochemical localization of P-galactosidase in whole alfalfa nodules induced by R. meliloti derivatives. Rm1021 is wild type, andRmG649 is Rm1021 (ntrC283 t15199:TnS-233). Plasmids pRKP9 and pMB211 carry nifH::lacZ and fixA::lacZ gene fusions, respectively.Plants were cleared with Na-hypochlorite prior to examination. (A) Nodule induced by RmG649 (no plasmid) 7 days postinoculation (dpi).No staining is seen. Bar, 20 pm. (B) Nodule induced by RmlO21(pRKP9) 15 dpi. Expression of P-galactosidase commences abruptly in asingle cell layer and continues to the proximal end of the nodule. Note the lack of expression in the most distal zones of the nodule (*). Bar,20 pum. (C) Nodule induced by RmG649(pMB211) 15 dpi. P-Galactosidase is expressed in all cells lying proximal to the meristem (*), includingthe prefixing zone II (*) (for comparison, see panel B). Bar, 20 pum. (D) Nodule induced by RmG649(pRKP9) 7 dpi. P-Galactosidase isexpressed in multiple infection threads (arrows) located distal to the developing nodule (*). Bar, 10 pLm. (E and F) Nodules induced byRmG649(pRKP9) and RMG649(pMB211), respectively, 7 dpi. Early expression of P-galactosidase in non-N2-fixing developing nodules isseen. Bar, 20 ,um.
succinate minimal medium. Because pTH99 carried only the3' portion of ntrC (685 bp), this result demonstrated that thesuppressor mutation had occurred in the C-terminal part ofthe ntrC protein. DNA extracted from one of the succinate-positive homogenotes, designated RmG855, was digestedwith PstI and cloned into pSUP202 by using a shotgun
strategy. Transformation of E. coli DH5a resulted in theisolation of one Spr clone (designated pTH100), whichshowed the same restriction pattern as pTH99. When thefISpr marker from pTH100 was marker exchanged into thedctDl6::TnS mutant RmF121, suppression of the succinate-negative phenotype occurred. This proved that the 3' end of
J. BACTERIOL.
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

ACTIVATION OF &,54-DEPENDENT PROMOTERS 2669
(a)'ntrB ntrC
pJel 2a5
EP HTH I
. p
II rB H Sp/Sm
pACYC184
E
IIHB
'ntrC(b) pTH99 pSUP202
I HTH I
P / B_. I-II
BHr I I
Sp/Sm H B- recombination into RmF283- cloning of the ntrC283 allele from homogenote RmG855
pTH1 00'ntrC283_:- pSUP202HTH
P B PI A _
I
Sp/Smr I I
H BI 11
BHI \I\P H
(d) sequenced regionZ~~~~~~~~~ATQ I K A A D L L G L N R N T L R K K I R E L G V S V Y R S S R S A -
CCAGATCAAGGCGGCCGACCTCCTGGGTCTCAACCGCAATACGC TGCGCAAGAAGATCCGCGAACTCGGGGTCTCGGTGTACCGCAG=CGCGTAGCGCTTGACTG
1610 1620 1630 1640 1650 1660 1670 1680 1690 1700 1710
FIG. 4. Strategy of cloning and sequencing of the ntrC283 allele. flSpr of pPH45 was inserted into the BglII site downstream of ntrC inpJe12a5 (a). The internal PstI fragment including fjSpr was subcloned into pSUP202 (b). The resulting clone, pTH99, was used to recombinefQSpr into the RmF283 genome. From the succinate-positive homogenote RmG855, total DNA was extracted, digested with PstI, and ligatedinto pSUP202. After transformation of recombinant plasmids, an Spr clone bearing the 3' portion of the ntrC283 allele was isolated and wasdesignated pTH100 (c). (d) The region of pTH100 which has been sequenced (PstI-HindIII fragment) and the part of the sequence bearing themutation in the helix-turn-helix (HTH) motif (marked by asterisks). The single nucleotide replacement in the helix-turn-helix motif isunderlined. Nucleotide positions 1692 and 1693, which differ from the original sequence published by Szeto et al. (57), are underlined and inboldface. The shift results in the addition of 2 amino acids to the C terminus of NTRC. E, EcoRI; P, PstI; B, BamHI; Bg, BglII; H, HindIII;Sm, SmaI.
ntrC carried on plasmid pTH100 harbored the ntrC283mutation. The PstI-HindIII fragment (the HindIII site islocated on the fISpr interposon [Fig. 4]) from pTH100 wassubcloned into pUC19, and the nucleotide sequence wasdetermined. In a parallel experiment, partial clone banks ofRmF283 and RmF121 were generated by rescuing fragmentsof EcoRI-BglII-digested total DNA (1.5 to 4 kb in size) ofboth strains from an agarose gel and cloning them intopUC19. Both libraries were probed with 32P-labelled ntrC
DNA. By using this strategy, the ntrC gene of RmF121, thedirect precursor of the suppressor strain, and the ntrC283allele could be isolated.Sequence analysis of both ntrC283 clones revealed that a
single A- T nucleotide substitution in the recognition helix(nucleotide 1642) had occurred, which led to the replacementof the Asn-466 residue in the wild-type NTRC protein by a
Tyr residue in the suppressor protein (Fig. 4d). The Asnresidue, which is located in the recognition helix for binding
(c)
IDI I -.01r,%- I
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2670 LABES ET AL.
to the UAS, has been found to be conserved in ntrC proteinsof several species (27).One C nucleotide, instead of two, was found at position
1692 in the wild-type ntrC sequence, compared with thatpreviously reported (57). The nucleotide difference shiftedthe reading frame and resulted in the addition of 2 aminoacids to the C-terminus of the encoded protein. The revisedsequence is given in Fig. 4d.
DISCUSSION
We have presented genetic and physical evidence that anunusual ntrC allele, designated ntrC283, carries a pointmutation in the 3' end of the ntrC gene. The ntrC283mutation results in the constitutive expression of the detA,niflI, fixA, and ginII promoters in free-living cells. More-over, we show that, in symbiosis, ntrC283 can functionallyreplace nifA, which is normally required for N2 fixation inroot nodules. While not directly demonstrated, we believethat the transcription activation observed in the strainscarrying the ntrC283 allele occurs through direct interactionof the NTRC283 protein with the transcriptional apparatus atthe respective activated promoters.The ntrC283 allele was initially identified as a second site
mutation which suppressed the Dct- phenotype of adctD::TnS insertion mutant. Data suggesting that the sup-pressor mutation mapped to ntrC were obtained from trans-duction experiments together with results showing thatsite-directed disruption of the ntrC gene in RmF283 abol-ished suppression. The suppressor mutation was localizedwithin 685 bp of the 3' end of ntrC through the observationthat Dct- recombinants were generated after the transfer ofa DNA fragment containing the wild-type ntrC into RmF283(Fig. 4b). Moreover, in the reciprocal experiment, Dct'recombinants were generated after the transfer of this regionfrom RmF283 (dctDl6::TnS ntrC283) into RmF121 (dctDJ6::TnS). Sequence analysis of the complete 685-bp region ofRmF283 and the parental strain RmF121 revealed a singleA-*T exchange leading to the substitution of the Asp-466residue in wild-type ntrC by Tyr in the ntrC283 mutant allele.
All of the or"-dependent promoters examined to daterequire a transcriptional activator to catalyze open complexformation (29) and transcription. Thus, the nifH, detA, andginII promoters are normally activated by the NIFA, DCTD,and NTRC proteins, respectively (30, 36, 53). However, ithas been observed that regulators can sometimes substitutefor each other, particularly when the regulator concentrationis increased by-overexpression. Thus, overexpressed NIFAcan substitute for NTRC in activating the glnAp2 promoterand in activating transcription of operons for histidine,proline, and arginine utilization in enterobacteria (33). E. coliNTRC can activate the R. meliloti nifH promoter (39, 56).On the other hand, substitution of NIFA by NTRC in theactivation of the nifH or nifLA promoters in Klebsiellaepneumoniae could not be detected (39). Neither wild-typenor mutant forms of NTRC have previously been observedto replace the requirement for NIFA in N2-fixing rootnodules. However, we note that in free-living R. meliloticells growing under N-limiting conditions (i.e., when theNTRC protein is activated), wild-type ntrC can activatetranscription from the nifH and fixA promoters. Interest-ingly, we have found that ntrC283 is constitutively expressedand, moreover, that the NTRC283 protein appears to beconstitutively active, independent of the nitrogen status ofthe cells (Table 3). Thus, the constitutive activation of the
nifH and fix4 promoters is consistent with the data of Szetoet al. (57).How can a single mutation in the helix-turn-helix motif of
NTRC283 lead to constitutive ntrC283 expression and con-stitutive ntrC283 activation? Interestingly, an ntrC allele ofBradyrhizobiumjaponicum which shares the latter two prop-erties of NTRC283 has been described (32). The B. japoni-cum ntrC allele carries an insertion in the 3' end of the ntrCgene. This mutation results in the synthesis of a truncatedNTRC protein. In addition, the truncated NTRC constitu-tively activates transcription from the ginlI promoter. Inwild-type cells, ntrC and ginII transcription was activated inN-limiting conditions. In B. japonicum, as in R. meliloti,ntrB and ntrC seem to form a single transcriptional unit (32,57). Martin et al. (32) suggested that the constitutive ntrCexpression they observed in their ntrC insertion mutantmight be due to loss of repressor activity at the ntrBCpromoter. In E. coli and K pneumoniae, unphosphorylatedNTRC represses transcription of ntrBC by binding to se-quences upstream of the promoter (44). Similarly, we sug-gest that the constitutive ntrC283 expression we observed(Fig. 2) is due, at least in part, to a loss of NTRC repressorbinding at the ntrBC promoter. We would thus expect thatboth NTRB and NTRC283 are overexpressed in RmF283(dctDl6::TnS ntrC283). The activation phosphorylation ofNTRC by NTRB in response to the cellular nitrogen status isnormally regulated via the interaction of the PII protein withNTRB (25, 37). Under conditions of N excess, PII inducesNTRB to dephosphorylate (inactivate) NTRC. Under Nlimitations, PII is uridylated and this frees NTRB to activate(phosphorylate) NTRC. Thus, overexpressed NTRB proteinwill titrate PIT, and this will tend to result in the samephenotype as a PIT-negative mutant, i.e., constitutive acti-vation (phosphorylation) of NTRC. For this reason, in R.meliloti RmF283, the derepression of the ntrBC operonresulting from the ntrC283 mutation is likely to have as asecondary effect the activation (phosphorylation) of theNTRC283 protein. An alternate explanation for the effects ofthe ntrC283 mutation is that the mutation in the DNAbinding region of NTRC283 results in an increase in phos-phorylation or activation of the NTRC283 protein indepen-dent of phosphorylation. These possibilities could be clearlyexamined in a biochemical study of the NTRC283 protein.The Asp-466 residue is located in the second or so-called
"recognition helix" of the postulated DNA binding helix-turn-helix motif of the R. meliloti NTRC protein (57). Theinvolvement of this motif in DNA binding by the NTRCprotein of K pneumoniae was clearly demonstrated byContreras and Drummond (9). How does the Asp--Tyrmutation in NTRC283 lead to high-level activation of theginII, dctA, nifH, and fix4 promoters? In wild-type cells, itappears that activated (phosphorylated) NTRC-P bound toUAS is first brought into contact with the or4-RNA poly-merase complex by DNA bending. NTRC-P-dependent ATPhydrolysis then catalyzes the isomerization of the closedtranscription initiation complex to an open form (2, 3, 35,68). We noted above that the ntrC283 mutation results in theconstitutive synthesis and (probably indirect) constitutiveactivation of the NTRC283 protein. It is possible that theconcentration of NTRC283 may be sufficient to allow it todirectly contact u5r4 and thus catalyze open complex forma-tion at the &154-RNA polymerase transcription initiationcomplex. Binding to UAS would not be required for suchactivation, and hence an unspecific activation of or4 promot-ers would occur. The data in Table 2 showing a 13-foldactivation of the R. meliloti dctA promoter, which lacks the
J. BACTERIOL.
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

ACTIVATION OF a54-DEPENDENT PROMOTERS 2671
helix turn helix
I I I I
Kp NTRC KQEAARLLGWGRNTLTRKLK
Bp NTRC QIRAADLLGLNRNTLRKKIR
Rm NTRC QIKAADLLGLNRNTLRKKIR
Rm NTRC283 QIKAADLLGLNRXTLRKKIR
Rm NIFA QAKAARLLGNTPRQVAYRIQ
Rm DCTD VKETLQALGIPRKTFYDKLQ
FIG. 5. Alignment of the helix-turn-helix motifs of the regulatorproteins NTRC, NTRC283, NIFA, and DCTD (27, 38, 57, 65). Themutation in NTRC283 is underlined. Rm, R. meliloti; Bp, Brady-rhizobium sp. (Parasponia); Kp, K pneumoniae.
two UAS, are consistent with this model. In addition, theconstitutive ntrC allele of Martin et al. (32) lacks the helix-turn-helix motif and yet constitutively activates ginII tran-scription. There are a number of other reports which areconsistent with the suggestion that NTRC283 can activatetranscription apparently without binding to UAS. Better etal. (5) demonstrated that the UAS in front of the R. melilotinifH promoter are not required for full activation by NIFA innodules. More recently, Huala and Ausubel (24) showed thata NIFA derivative in which the C-terminal DNA bindingdomain was removed can still activate the nifH promoter to10% of the level obtained with intact wild-type NIFA pro-tein. Reitzer and Magasanik (44) showed that, in E. coli, theminimal glnAp2 promoter in which all known UAS weredeleted could still be activated by NTRC.The Asn- Tyr substitution giving rise to NTRC283 may
also change the DNA binding specificity so that it can now
recognize activator sequences, perhaps at a low affinity,which are normally bound by DCTD, NTRC, and NIFA.While this would appear unlikely, the more than sixfoldreduction in the NTRC283-dependent dctA transcriptionwhich occurs when the DCTD binding sites are removed(Table 2) suggests that some recognition of these sites byNTRC283 does occur. Comparisons of the amino acid se-
quence of the second helix of NTRC283 with equivalenthelices of DCTD or NIFA revealed little similarity in theirprimary structures (Fig. 5).The nifA gene product regulates the expression of many
genes involved in N2 fixation in symbiotic and free-livingbacteria (59). Thus, nifA R. meliloti, B. japonicum, and R.leguminosarum mutants show no N2-fixing activity (26, 58,59). The nifA-independent activation of nif genes in free-living R. meliloti has already been described (57). However,this is the first report showing that nifA is dispensable fortheir symbiotic activation.
Symbiotic regulation of nifA expression during noduledevelopment occurs via the fixLJ and fixK cascade in re-
sponse to low oxygen concentrations (4, 11, 12, 64). We notethat because fixLJ is required for expression of nifA-inde-pendent genes, we would not expect the ntrC283 allele tosuppress the Fix- phenotype of fixL or fixJ mutations. Thiswas verified by inoculating alfalfa plants with fixL ntrC283and fix. ntrC283 double mutants (data not shown). In wild-type pea nodules, both nifA and nifH mRNAs are expressedin the third or fourth cell layers of the late symbiotic zone
(69). A similar abrupt expression of these genes occurs in
alfalfa root nodules (60). Here, we have shown that thepresence of ntrC283 alters the temporal and spatial regula-tion of nifH andfixA expression in root nodules (Fig. 4). Thisresult is consistent with the ability ofntrC283 to suppress theFix- phenotype of the nifA deletion mutant (Table 5). Ourresults suggest that the fine-tuned regulation of gene expres-sion in nodules may represent an evolutionary optimizationof symbiosis and that such regulation is not absolutelyrequired for symbiotic nitrogen fixation.
ACKNOWLEDGMENTS
We thank Michael Kahn and Ray Dixon for providing plasmidscarrying promoter fusions. Special thanks are due to Tracy Nixonfor providing us with the dctA promoter constructs, for communi-cating information on unpublished results, and for discussions. Wethank Trevor Charles for performing the initial transduction exper-iments; Jacques Batut, Pierre Boistard, and Clive Ronson fordiscussions; and Brian Driscoll for critical reading of the manu-script. Pictures showing the in situ localization of 13-galactosidasewere taken by Georges Truchet in his laboratory. The latter exper-iments could not have been done without his assistance and inter-pretive guidance, merci mon ami.
This study was supported by operating and strategic grants fromthe Natural Sciences and Engineering Research Council of Canadaand Agriculture Canada.
REFERENCES1. Arwas, R., I. A. McKay, F. R. P. Rowney, M. J. Dilworth, and
A. R. Glenn. 1985. Properties of organic acid utilization mutantsof Rhizobium leguminosarum strain 300. J. Gen. Microbiol.144:2059-2066.
2. Austin, S., and R. Dixon. 1992. The prokaryotic enhancerbinding protein NTRC has an ATPase activity which is phos-phorylation and DNA dependent. EMBO J. 11:2219-2228.
3. Austin, S., C. Kundrot, and R. Dixon. 1991. Influence of amutation in the putative nucleotide binding site of the nitrogenregulatory protein NTRC on its positive control function. Nu-cleic Acids Res. 19:2281-2287.
4. Batut, J., M.-L. Daveran-Mignot, M. David, A. M. Garnerome,and D. Kahn. 1989. fixK, a gene homologous with fnr and crpfrom Escherichia coli, regulates nitrogen fixation genes bothpositively and negatively in Rhizobium meliloti. EMBO J.8:1279-1286.
5. Better, M., G. Ditta, and D. R. Helinski. 1985. Deletion analysisof Rhizobium meliloti symbiotic promoters. EMBO J. 4:2419-2424.
6. Boivin, C., S. Camut, C. A. Malpica, G. Truchet, and C.Rosenberg. 1990. Rhizobium meliloti genes encoding catabolismof trigonellin are induced under symbiotic conditions. Plant Cell2:1157-1170.
7. Bolton, E., B. Higgisson, A. Harrington, and F. O'Gara. 1986.Dicarboxylic acid transport in Rhizobium meliloti: isolation ofmutants and cloning of dicarboxylate transport genes. Arch.Microbiol. 144:142-146.
8. Buck, M., W. Cannon, and J. Woodcock. 1987. Transcriptionalactivation of the Kiebsiella pneumoniae nitrogenase promotermay involve DNA loop formation. Mol. Microbiol. 1:243-249.
9. Contreras, A., and M. Drummond. 1988. The effect of thefunction of the transcriptional activator NTRC from Klebsiellapneumoniae of mutations in the DNA recognition helix. NucleicAcids Res. 16:4025-4039.
10. de Billy, F., D. G. Barker, P. Gallusci, and G. Truchet. 1991.Leghaemoglobin gene transcription is triggered in a single celllayer in the indeterminate nitrogen-fixing root nodule of alfalfa.Plant J. 1:27-35.
11. de Philip, P., J. Batut, and P. Boistard. 1990. Rhizobium melilotiFIXL is an oxygen sensor and regulates R. meliloti nifA andfixK genes differentially in E. coli. J. Bacteriol. 172:4255-4262.
12. Ditta, G., E. Viris, A. Palomares, and C. H. Kim. 1987. The nifAgene of Rhizobium meliloti is oxygen regulated. J. Bacteriol.169:3217-3223.
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

2672 LABES ET AL.
13. Dixon, R. 1986. The xyIABC promoter from Pseudomonasputida TOL plasmid is activated by nitrogen regulatory genes inEscherichia coli. Mol. Gen. Genet. 203:129-136.
14. Dixon, R., T. Eydmann, N. Henderson, and S. Austin. 1991.Substitutions at a single amino acid residue in the nitrogen-regulated activator protein NTRC differentially influence itsactivity in response to phosphorylation. Mol. Microbiol.5:1657-1667.
15. Engelke, T. H., M. N. Jagadish, and A. Puehler. 1987. Biochem-ical and genetic analysis of Rhizobium meliloti mutants defec-tive in C4-dicarboxylate transport. J. Gen. Microbiol. 133:3019-3029.
16. Engelke, T. H., D. Jording, D. Kapp, and A. Puehler. 1989.Identification and sequence analysis of the Rhizobium melilotidetA gene encoding the C4-dicarboxylate carrier. J. Bacteriol.171:5551-5560.
17. Finan, T. M., E. Hartwieg, K. LeMieux, K. Bergman, G. C.Walker, and E. R. Signer. 1984. General transduction in Rhizo-bium meliloti. J. Bacteriol. 159:120-124.
18. Finan, T. M., B. Kunkel, G. F. De Vos, and E. R. Signer. 1986.Second symbiotic megaplasmid in Rhizobium meliloti carryingexopolysaccharide and thiamine synthesis genes. J. Bacteriol.167:66-72.
19. Finan, T. M., I. Oresnik, and A. Bottacin. 1988. Mutants ofRhizobium meliloti defective in succinate metabolism. J. Bac-teriol. 170:3396-3403.
20. Finan, T. M., J. M. Wood, and D. C. Jordan. 1983. Symbioticproperties of C4-dicarboxylic acid transport mutants of Rhizo-bium leguminosarum. J. Bacteriol. 154:1403-1413.
21. Gu, B., H. Ledebur, and T. Nixon. Personal communication.22. Gu, B., and T. Nixon. 1990. In P. M. Gresshoff, L. E. Roth, G.
Stacey, and W. E. Newton (ed.), Nitrogen fixation: achieve-ments and objectives, p. 538. Chapman and Hall, New York.
23. Hirschmann, J., P.-K. Wong, K. Sei, J. Keener, and S. Kustu.1985. Products of nitrogen regulatory genes ntrA and ntrC ofenteric bacteria activate glnA transcription in vitro: evidencethat the ntrA product is a sigma factor. Proc. Natl. Acad. Sci.USA 82:7525-7529.
24. Huala, E., and F. M. Ausubel. 1989. The central domain ofRhizobium meliloti NIFA is sufficient to activate transcriptionfrom the R. meliloti nifH promoter. J. Bacteriol. 171:3354-3365.
25. Hunt, T. P., and B. Magasanik. 1985. Transcription of ginA bypurified Escherichia coli components: core RNA polymeraseand the products ofglnF, glnG and glnL. Proc. Natl. Acad. Sci.USA 82:8453-8457.
26. Iismaa, S. E., and J. M. Watson. 1989. The nifA gene product ofRhizobium leguminosarum biovar trifolii lacks the N-terminaldomain found in other NIFA proteins. Mol. Microbiol. 3:943-955.
27. Jones, R., and R. Haselkorn. 1989. The DNA sequence of theRhodobacter capsulatus ntrA, ntrB and ntrC gene analoguesrequired for nitrogen fixation. Mol. Gen. Genet. 215:507-516.
28. Klipp, W., H. Reilaender, A. Schlueter, R. Krey, and A. Puehler.1989. The Rhizobium melilotifdxN gene encoding a ferredoxin-like protein is necessary for nitrogen fixation and is cotran-scribed with nifA and nifl. Mol. Gen. Genet. 216:293-302.
29. Kustu, S., E. Santero, J. Keener, D. Popham, and D. Weiss.1989. Expression of o54 (ntrA)-dependent genes is probablyunited by a common mechanism. Microbiol. Rev. 53:367-376.
30. Ledebur, H., B. Gu, J. Sojda III, and B. T. Nixon. 1990.Rhizobium meliloti and Rhizobium leguminosarum dctD geneproducts bind to tandem sites in an activation sequence locatedupstream of cr54-dependent dctA promoters. J. Bacteriol. 172:3888-3897.
31. Long, S. R. 1989. Rhizobium-legume nodulation: life together inthe underground. Cell 56:203-214.
32. Martin, G. B., K. A. Chapman, and K. B. Chelm. 1988. Role ofthe Bradyrhizobium japonicum ntrC gene product in differentialregulation of glutamine synthetase II gene (glnII). J. Bacteriol.170:5452-5459.
33. Merrick, M. 1983. Nitrogen control of the nif regulon in Kleb-siella pneumonia: involvement of the ntrA gene and analogiesbetween ntrC and nifA. EMBO J. 2:39-44.
34. Miller, J. H. 1972. Experiments in molecular genetics. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
35. Minchin, S. D., S. Austin, and R. A. Dixon. 1989. Transcrip-tional activation of the Kiebsiella pne moniae nifL4 promoterby NTRC is face-of-the-helix dependent and the activatorstabilizes the interaction 'of or54-RNA polymerase with thepromoter. EMBO J. 8:3491-3499.
36. Morett, E., and M. Buck. 1988. NIFA-dependent in vivo pro-tection demonstrates that the upstream activator sequence of nifpromoters is a protein binding site. Proc. Natl. Acad. Sci. USA85:9401-9405.
37. Ninfa, A. J., and B. Magasanik. 1986. Covalent modification ofthe ginG product, NRI, by the glnL product, NRII, regulatesthe transcription of the glnALG operon in Escherichia coli.Proc. Natl. Acad. Sci. USA 83:5909-5913.
38. Nixon, B. T., C. W. Ronson, and F. M. Ausubel. 1986. Two-component regulatory systems responsive to environmentalstimuli share strongly conserved domains with the nitrogenassimilation regulatory genes ntrB and ntrC. Proc. Natl. Acad.Sci. USA 83:7850-7854.
39. Ow, D. W., and F. M. Ausubel. 1983. Regulation of nitrogenmetabolism genes by nifA gene product. Nature (London)301:307-313.
40. Peters, N. K., J. W. Frost, and S. R. Long. 1986. A plant flavone,luteolin, induces expression of Rhizobium melioti nodulationgenes. Science 233:917-1008.
41. Popham, D. L., D. Szeto, J. Keener, and S. Kustu. 1989.Function of a bacterial activator protein that binds to transcrip-tional enhancers. Science 243:629-635.
42. Prentki, P., and H. M. Krisch. 1984. In vitro insertional muta-genesis with a selectable DNA fragment. Gene 29:303-313.
43. Redmond, J. W., M. Batley, M. A. Djordjevic, R. W. Innes,P. L. Kuempel, and B. G. Rolfe. 1986. Flavones induce expres-sion of nodulation genes in Rhizobium. Nature (London) 323:632-634.
44. Reitzer, L. J., and B. Magasanik. 1986. Transcription ofglnA inE. coli is stimulated by activator bound to sites far from thepromoter. Cell 45:785-792.
45. Ronson, C. W., and P. M. Astwood. 1985. Genes involved in thecarbon metabolism of bacteroids, p. 201-207. In H. J. Evansand P. J. Bottomley (ed.), Nitrogen fixation research progress.Martinus Nijhoff Publishers, Dordrecht, The Netherlands.
46. Ronson, C. W., P. M. Astwood, B. T. Nixon, and F. M. Ausubel.1987. Deduced products of C4-dicarboxylate transport regula-tory genes of Rhizobium leguminosarum are homologous tonitrogen regulatory gene products. Nucleic Acids Res. 15:7921-7934.
47. Ronson, C. W., P. Lyttleton, and J. G. Robertson. 1981. C4-dicarboxylate transport mutants of Rhizobium leguminosarum.Proc. Natl. Acad. Sci. USA 78:4284-4288.
48. Ronson, C. W., B. T. Nixon, L. M. Albright, and F. M. Ausubel.1987. Rhizobium meliloti ntrA (rpoN) gene is required fordiverse metabolic functions. J. Bacteriol. 169:2424-2431.
49. Ronson, C. W., B. T. Nixon, and F. M. Ausubel. 1987. Con-served domains in bacterial regulatory proteins that respond toenvironmental stimuli. Cell 49:579-581.
50. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
51. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
52. Sasse-Dwight, S., and J. D. Gralla. 1990. Role of eucaryotic-typefunctional domains found in the procaryotic enhancer receptorfactor sigma-54. Cell 62:945-954.
53. Shatters, R. G., J. E. Somerville, and M. Kahn. 1989. Regulationof glutamine synthetase II activity in Rhizobium meliloti104A14. J. Bacteriol. 171:5087-5094.
54. Simon, R., U. Priefer, and A. Puehler. 1983. A broad host rangemobilization system for in vivo genetic engineering: transposonmutagenesis in gram negative bacteria. Bio/Technology 1:784-791.
55. Su, W., S. Porter, S. Kustu, and H. Echols. 1990. DNA-looping
J. BA=rRIOL.
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

ACTIVATION OF p54-DEPENDENT PROMOTERS 2673
and enhancer activity: association between DNA-bound NTRCactivator and RNA polymerase at the bacterial glnA promoter.Proc. Nati. Acad. Sci. USA 87:5504-5508.
56. Sundaresan, V., D. W. Ow, and F. M. Ausubel. 1983. Activationof Kiebsiella pneumoniae and Rhizobium meliloti nitrogenasepromoters by gin (ntr) regulatory genes. Proc. Nati. Acad. Sci.USA 80:4030-4034.
57. Szeto, W. W., B. T. Nixon, C. W. Ronson, and F. M. Ausubel.1987. Identification and characterization of the Rhizobium me-liloti ntrC gene: R. meliloti has separate regulatory pathways foractivation of nitrogen fixation genes in free-living and symbioticcells. J. Bacteriol. 169:1423-1432.
58. Szeto, W. W., J. L. Zimmermann, V. Sundaresan, and F. M.Ausubel. 1984. A Rhizobium meliloti regulatory gene. Cell36:1035-1043.
59. Thony, B., and H. Hennecke. 1989. The -24/-12 promotercomes of age. FEMS Microbiol. Rev. 5:341-357.
60. Truchet, G. Personal communication.61. Truchet, G., S. Camut, F. de Billy, R. Odorico, and J. Vasse.
1989. The Rhizobium-legume symbiosis: two methods to dis-criminate between nodules and other root-derived structures.Protoplasma 149:82-88.
62. Truchet, G., P. Roche, P. Lerouge, J. Vasse, S. Camut, F. deBilly, J.-C. Prome, and J. Denarie. 1991. Sulphated lipo-oligo-saccharide signals of Rhizobium meliloti elicit root noduleorganogenesis in alfalfa. Nature (London) 351:670-673.
63. Vasse, J., F. de Billy, S. Camut, and G. Truchet. 1989. Corre-lation between ultrastructural differentiation of bacteroids and
nitrogen fixation in alfalfa plants. J. Bacteriol. 172:4295-4306.64. Virts, E. L., S. W. Stanfield, D. R. Helinski, and G. S. Ditta.
1988. Common regulatory elements control nitrogen fixationand microaerobic induction of nifA in Rhizobium meliloti. Proc.Natl. Acad. Sci. USA 85:3062-3065.
65. Watson, R. 1990. Analysis of the C4-dicarboxylate transportgenes of Rhizobium melioti: nucleotide sequence and deducedproducts of dctA, dctB and dctD. Mol. Plant-Microbe Interact.3:174-181.
66. Watson, R. J., Y. K. Chan, R. Wheatcroft, A. F. Yang, and S.Han. 1988. Rhizobium melioti genes required for C4-dicarbox-ylate transport and symbiotic nitrogen fixation are located on amegaplasmid. J. Bacteriol. 70:927-934.
67. Wedel, A., D. S. Weiss, D. Popham, P. Droge, and S. Kustu.1990. A bacterial enhancer functions to tether a transcriptionalactivator near a promoter. Science 248:486-490.
68. Weiss, D. S., J. Batut, K. E. Klohse, J. Keener, and S. Kustu.1991. The phosphorylated form of the enhancer binding proteinNTRC has an ATPase activity that is essential for activation oftranscription. Cell 67:155-167.
69. Yang, W.-C., B. Horvath, J. Hontelez, A. Van Kammen, and T.Bisseling. 1991. In situ localisation of Rhizobium mRNAs in pearoot nodules: nifA and nifHl localization. Mol. Plant-MicrobeInteract. 5:464-468.
70. Yarosh, 0. K., T. C. Charles, and T. M. Finan. 1989. Analysisof the C4-dicarboxylate transport genes in Rhizobium meliloti.Mol. Microbiol. 3:813-823.
VOL. 175, 1993
on April 29, 2020 by guest
http://jb.asm.org/
Dow
nloaded from