systems biology lecture series cell cycle€¦ · systems biology lecture series cell cycle pawan...
TRANSCRIPT

Systems Biology lecture seriesCell cycle
Pawan DharBII
Feb 06, 2004

What is similar between cells andwashing machines!

CELLS COME FROM OTHER CELLS
• Questions:• How are they duplicated?• How are they partitioned?• How are these processes coordinated ?• How are mistakes overlooked / rectified?

Interesting facts
• 25,000,000 cells divide per minute in our bodies• Every 8 seconds DNA suffers a break at least once!• Some cells continuously divide (skin)• Some cell types are quiescent, unless provoked (blood)• Some cells never divide (muscle, neurons)• Some cells are dead but still part of our body (hair, nails)
• Division leads to death, eventually !• Control of cell division is control of cancer.• Our goal is to understand cell cycle controls and discover
new ones

The cell cycle is an ordered series ofevents leading to the replication of cells

• Typically human cell takes 24 h to divide,• G1=9h -> S 10h ->G2 4.5h ->M 0.5h• Cells stay in G1 till growth factors, size factors
signal progression• A prolonged stay in G1 is called G0 (example)• M phase - P+M+A+T

The stages of mitosis andcytokinesis in an animal cell

Centrosome duplication precedesand is required for mitosis

The mitotic apparatus is a machinefor separating chromosomes

The mitotic apparatus in a metaphase mammalian cell
Note 3 types of microtubules

Capture of chromosomes is alsoimportant for spindle assembly

Dynamic instability of microtubulesincreases during mitosis

Current concept: Bipolar spindle determines cell division plane
New concept: Cell Division is independent of spindle polarity
Pawan Dhar. The Cell equator - more than poles apart. Trends In Biotechnology (April 2004 issue)

Major concerns:
1. ordered process - activate cellular machines onlyat correct time
2. Ensure completion before progression - replication before mitosis - mitosis before cytokinesis
- growth before replication 3. Must be subject to external controls e.g, divide
only when more cells needed

The original cyclin based model of cell cycle

The Pioneers
Hartwell et al (1999): Nature 402, C47-C52
“We need to develop simplifying, higher levelmodels and find general principles that willallow us to grasp and manipulate the functionof biochemical networks”

Hartwell - Nobel lecture 2001
Cdc - Cell division cycle

Checkpoints in cell cycle regulation

What is a cell cycle?G1-S-G2-MS <=> M (cohesin)
Q: How to ensure the orderlyprogression of events - nuclearcycle vs. cytoplasmic cycle ?
Cell cycle is also achromosomal cycle

Evidence for Cell cycle regulation
Key experiment 1
Rao and JohnstonS+G1: S phase starts in G1nucleiS+G2: No S phase in G2 nucleiG1+G2 : no S or M phaseM phase + Interphase: Mitosis
Conclusions
1. Diffusable factors promote S or M phase
2. S phase promoting factor only works in G1 nuclei.
3. M phase promoter works on everything

Key experiment 2
Xenopus extracts (Tim Hunt)Group 1: Oocytes blocked at M phaseGroup 2: Oocytes at post M phase
Q: Is it possible to induce # 1 to enter M phaseA: Yes.
MPF was discovered !

Key experiment 3
Material: Sea Urchin Embryos , synchronously dividing
Key observation: Of all the proteins, some regularly go up andsome go down with the cell cycle
Cyclins were discovered
Key experiment 4
Material: Yeast cells
Q: Is it possible to isolate yeast mutants that can grow but cannotdivide their nuclei. Use these mutant combinations to identify cellcycle controls ?

Breakthroughs in genetics
Why yeast ? - Budding, fission yeast
Task: Look for conditional mutants
Q: How to determine the stage atwhich the cell is blocked ?
Key finding: cdc mutants discovered(cdc = cell division cycle)
Task: Remove a key componentQ: Does it affect the pattern / timing of regulation ?

ModelSlow down fission yeast cycle - long cellsSpeed up fission yeast cycle - short cells
Task: Isolate short or “wee” mutants< defective in timing or regulation >
Q: How many genes are mutated i.e., for allthe mutants that arrest cells in G2, how manydifferent genes do they represent ?
Task: Combine mutants together.Q: Is it possible to determine the pathwaysbetween them ?

Case study: Fission yeast
Mutants Phenotypecdc25 longwee1 shortcdc2-L longcdc2-w shortOP cdc25 shortOPwee1 longcdc13 long
Mutants Phenotypecdc2-L wee1 longcdc2-L OP cdc25 longcdc2-w cdc25 shortcdc2-w OP wee1 shortcdc25 wee1 normal
Conclusion1. Wee1 cdc25 act upstream of cdc22. Wee1 inhibits cdc2 (absence ofwee1 speeds cell cycle)3. cdc25 activates cdc2, because ifcdc25 is missing, things slow.4. cdc25 and wee1 antagonize eachother, because together everythinglooks okay
`
Model
wee1 cdc25
cdc2 + cdc13
G2 M
Protein kinase Phosphatase
Proteinkinase
cyclin

cyclin
Cyclin and kinase
cdc28cdc28
Budding: Cdc28/Cln1 and Cdc28/cln2
START: Cdc28/Cln3
Mitosis: Cdc28/Clb1, Cdc28/Clb2
DNA replication: Cdc28/Clb5, Cdc28/Clb6
Budding yeast
How are CDK/Cyclindimers regulated ?
cyclin

How is the transition accomplished ?1 .Phosphorylation / Dephosphorylation2. Inhibition (to regulate kinase activity)3. Destruction - of cyclin and inhibitors (sequential and temporal)
Regulated destruction
1. Degradation of cyclins2. CKIs e.g, Sic1, Rum13. Cohesin - molecular glue

Complexregulatory
process

Cell cycle regulation - 1 of 2

Cell cycle controls 2 of 2

http://pingu.salk.edu/~forsburg/cclecture.html
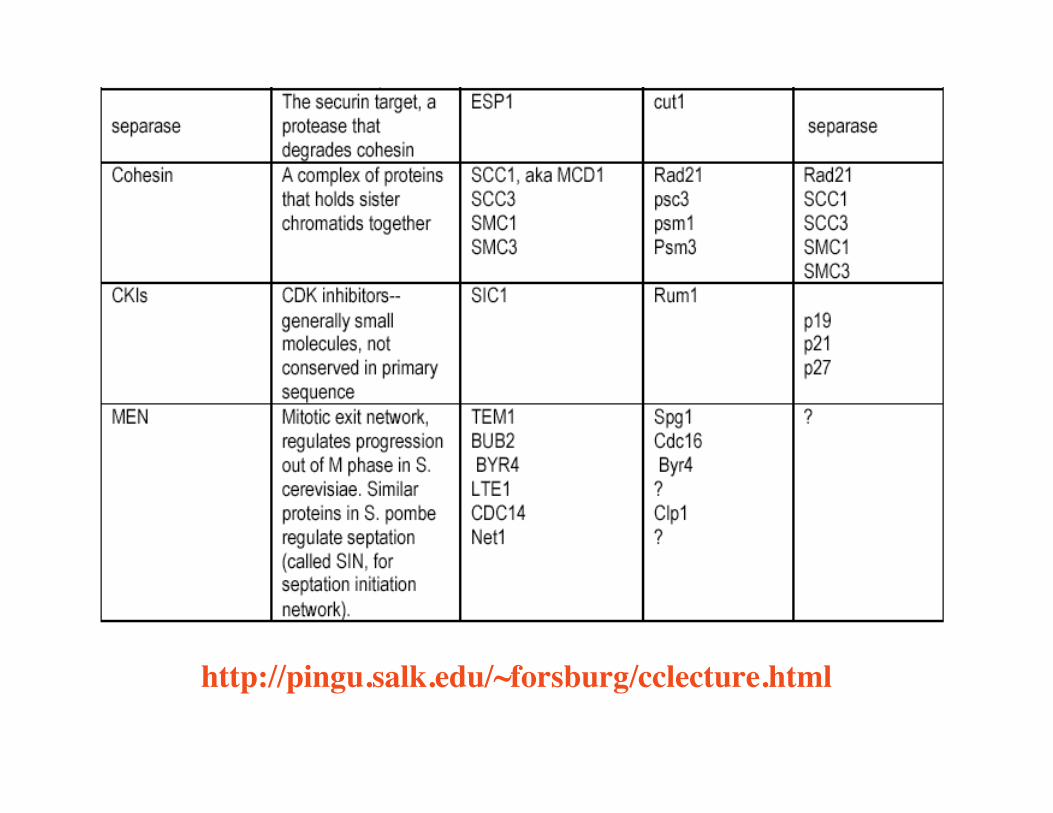
http://pingu.salk.edu/~forsburg/cclecture.html

G1 and G2 arrest in cells with damaged DNAdepends on the p53 tumor suppressor

Modeling cell cycle transactions

• The first mathematical model of cell cycle• Most elaborate analytical model• Correctly predicts linkage between cell size and G1 cyclin (Cln3) gene dosage• Prediction of cyclin and Sic1 abundance in various situation was reasonably accurate
What’s good about Tyson’s model ?
JTB 2001:210, 249-263Mol.Biol.Cell 2000: 11, 369-391Philos.Trans.R.Soc.Lond. 1998: 353, 2063-76

€
Equation governing cyclin - dependent kinases :
1. ddt
Cln2[ ] = ks, n 2' + ks, n 2
' ' SBF[ ]( ) ⋅mass− kd, n 2 Cln2[ ]
2. ddt
Clb2[ ]T = ks, b 2' + ks, b 2
' ' Mcm1[ ]( ) ⋅mass− Vd, b 2⋅ Clb2[ ]T
3. Vd, b 2 = kd, b 2' 1− Hct1[ ]( ) + kd, b 2
' ' Hct1[ ]+ kd, b 2' ' ' Cdc20[ ]A
4. ddt
Clb5[ ]T = ks, b 5' + ks, b 5
' ' MBF[ ]( ) ⋅mass− Vd, b 5⋅ Clb5[ ]T
5. Vd, b 5 = kd, b 5' ' ' + Esp1[ ] ⋅ kd, b 5
' ' + kd, b 5' Cdc20[ ]A( ) + kd, b 5 Cdc20[ ]A
6. Cln3[ ] = k c ln 3⋅mass7. Clb2[ ]T = Clb2[ ]+ Clb2 Sic1[ ]8. Clb5[ ]T = Clb5[ ]+ Clb5 Sic1[ ]9. Sic1[ ]T = Sic1[ ]+ Clb2 Sic1[ ]+ Clb5 Sic1[ ]+ Sic1P[ ]
Equation governing the inhibitor of Clb - dependent kinases :
10. ddt
Sic1[ ]T = ks, c 1' + ks, c 1
' ' Swi5[ ]− kd, c 1 Sic1[ ]P
11. ddt
Sic1[ ]P = ?k 4 Sic1[ ]T − Sic1[ ]P( ) − kpp, sic Cdc14[ ]+ kd, sic( ) ⋅ Sic1[ ]P
12. ?k 4 = k4' + k4
?cdk, sic
km 2+ Sic1[ ]T13. ?cdk, sic = ?c 3⋅ ?c ln 3+ 0.1 ⋅ X[ ] ⋅ mass[ ]( ) +?c 2⋅ Cln2[ ]+?c 5⋅ Clb5[ ]+?c b2⋅ Clb2[ ]
14. ?c ln 3 =max c ln 3⋅ Cln3[ ]Cln3[ ]+ kf, s 3
15. ddt
Clb2 Sic1[ ] = − kir2 Clb2 Sic1[ ]+ ki Clb2[ ] ⋅ Sic1[ ]− kd 2+?k 4( ) ⋅ Clb2 Sic1[ ]
16. ddt
Clb5 Sic1[ ] = − kir5 Clb5 Sic1[ ]+ ki Clb5[ ] ⋅ Sic1[ ]− kd 5+?k 4( ) ⋅ Clb5 Sic1[ ]
17. ddt
Swi5[ ]T = ks, swi 5' + ks, swi 5 Mcm1[ ]− kd, swi 5 Swi5[ ]T
Tyson’s equations

Work at BII
Forward modelingMandar Chitre (NUS)
Meng How (NTU)Mother Daughter
BudScars
The Budding Yeast Saccharomyces cerevisiae
Reverse engineeringDr.Sun Yan (BII)
Courtesy: Uttam Surana, IMCB

Our Cell cycle Model
• 37 entities - its growing !• 51 reactions• 78 parameters• 17 initial values• Takes 30 min on a
1.6 GHz Pentium IV torun a 120 min simulation

Which experimentalresults do weintend to confirm ?

* Dr.Uttam Surana, IMCB
G1 S G2 G1metaphase telophase
Mitosis
Clb 1,2 levels
APC Hct1OldModel
APC Cdc20
APC Hct1
NewModel
Pattern of Destruction*

Cell Cycle Pathway Modelin Saccharomyces cerevisiaeby Mandar Chitre, Mar 2003
CLB2Mcm1
Inactive
Mcm1
APC/Cdc20
Hct1
Hct1-P
Cdc14
RENT/Cdc14
CLN2
SBF
Inactive
SBF
CLB5
Cdc28/Cln2
Cdc28/Clb2
Cdc28/Clb5
MBF
InactiveMBF
SIC1
Sic1
Swi5
InactiveSwi5
Sic1-P
Cdc28/
Clb5/Sic1
Cln3
Cln3
via Pds1, Esp1
APC/
Hct1
CDC20
via MCM
Cdc20
MAD/Cdc20
MAD
APC
Cdc28/Clb2/Sic1
Used to triggercell cycle
Forwardmodeling

SGD Expression Data
• Possible constraints:– Strong constraints
• Relative concentration of Cyclins at constant time step– Weak constraints
• Minimum/maximum relative concentrations of Cyclins• Cell cycle time
• Should constraints include non-Cyclin concentrations ?• Which synchronization method is most similar to
an undisturbed budding yeast cell ?

Training Constraints• Automated tuning can be based on observed
constraints derived from SGD data• However, SGD uses 4 different synchronization
methods:– Alpha factor– CDC15– Cell size– CDC28
• Each method leads to different cell cycleparameters – amplitude, cycle time, etc.

Problems with numerical model
Mandar 2003

Multi-state logic model
Qualitativemodel onMEN

Reverse EngineeringDr.Sun Yan’s work
1. Wavematch method v/s Spellman method2. Identified NEW genetic regulators of cell
cycle from genomic expression profiles

Reading material
Dr Forsburg's all-purpose Cell Cycle Lecture Noteshttp://pingu.salk.edu/~forsburg/cclecture.html
http://mips.gsf.de/Yeast: http://mips.gsf.de/genre/proj/yeast/index.jsp