szpitter et al. 2014 - effect of dionaea muscipula extract and plumbagin on maceration of potato...
DESCRIPTION
EffectTRANSCRIPT

Annals of Applied Biology ISSN 0003-4746
R E S E A R C H A R T I C L E
Effect of Dionaea muscipula extract and plumbagin onmaceration of potato tissue by Pectobacterium atrosepticumA. Szpitter1, M. Narajczyk2, M. Maciag-Dorszynska3, G. Wegrzyn3, E. Lojkowska1 & A. Krolicka1
1 Department of Biotechnology, Intercollegiate Faculty of Biotechnology, University of Gdansk and Medical University of Gdansk, Gdansk, Poland
2 Laboratory of Electron Microscopy, University of Gdansk, Gdansk, Poland
3 Department of Molecular Biology, University of Gdansk, Gdansk, Poland
KeywordsAntimicrobial activity; efflux pumps;
pectinolytic bacteria; plant metabolites.
CorrespondenceA. Krolicka, Department of Biotechnology,
Intercollegiate Faculty of Biotechnology,
University of Gdansk and Medical University of
Gdansk, Kladki 24, 80-822 Gdansk, Poland.
Email: [email protected]
Received: 29 March 2013; revised version
accepted: 5 December 2013; published
online: 28 February 2014.
doi:10.1111/aab.12110
Abstract
Pectobacterium atrosepticum (Pba) is a plant pathogen that causes major croplosses. Dionaea muscipula extracts and their antibacterial constituent, plumbagin,inhibit Pba growth in vitro. However, this effect is reduced when the extractsare added to bacterial cultures present on potato tubers or suspended in potatotuber filtrate (PF). To explain this, we examined the response mechanism ofPba cells to Dionaea extract and plumbagin and compared it with the effect ofa bactericidal peptide – CAMEL. The addition of the extract and plumbagin toa Pba1043 culture in stationary phase increased the extracellular pectate lyase(Pel) activity in the presence of PF. While the addition of the Dionaea extractand plumbagin caused a dramatic reduction in RNA and protein synthesis inPba1043, it did not result in cellular damage. PF alone increased the expressionof Pba genes encoding protein components of cellular efflux pump systems:ompX, acrA and emrA. Application of both PF and plumbagin resulted in asynergistic stimulation of acrA gene expression. Plumbagin added to potatotubers inoculated with a field isolate Pba5A/1/2005 increased extracellularPel activity and reduced tissue maceration but did not affect bacterial countsper gram of tissue. These results show that plumbagin in the presence ofcompounds from potato tuber stimulates Pel production/secretion in Pba cellsand increases the expression of the acrA gene. This may be the molecular basisfor the less pronounced effects of Dionaea extract on Pba in planta relative tothose observed in vitro.
Introduction
Plant pathogenic bacteria are a significant problem in
agriculture. Major losses during the production and
storage of potato, carrot, cabbage and tomato are
caused by pectinolytic Pectobacterium and Dickeya spp.
These bacteria are characterised by high genotypic and
phenotypic diversity and are capable of causing disease
symptoms on a wide range of host plants (including
many vegetables such as potato and ornamental plants)
in different climates (Ma et al., 2007). Additionally,
diseases caused by Pectobacterium and Dickeya species
cannot be controlled by bactericides, and their control
currently depends on the early detection of bacteria and
resistant cultivar plantation (Perombelon and Kelman,
1980; Perombelon, 2002; Zimnoch-Guzowska et al., 2005;Czajkowski et al., 2011).
On the basis of various suppressive effects of plantchemicals on bacterial cells, new antibacterial compoundshave been developed including inhibitors of quorumsensing and of multidrug resistance pumps (Hentzeret al., 2002; Ball et al., 2006). In spite of the chemicaldiversity of plant metabolites so far none of thosecompounds proved to be sufficiently active in orderto be introduced to the market with the intention ofpreventing bacterial diseases in crop plants. This is dueto the fact that bacterial plant pathogens have evolvedefficient mechanisms to avoid plant chemical defenceand to respond to plant antimicrobials as well as plantcell degradation products. Plant phenolics were found to
404 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists

A. Szpitter et al. Pectobacterium response to plant antimicrobial compounds
induce the expression of efflux pumps (Ravirala et al.,2007) and the components of type III secretion system inDickeya dadantii cells (Yang et al., 2008). Bacterial responsemechanisms identified so far also include upregulation ofplant cell degrading enzymes. The presence of ferulicacid, a plant cell wall degradation product, stimulates theproduction of feruloyl esterase and pectate lyases in D.
dadantii cells (Hassan & Hugouvieux-Cotte-Pattat, 2011).Moreover, the expression of proline iminopeptidase genein Xanthomonas campestris pv. campestri was shown tobe upregulated in the presence of an unknown plantmetabolite (Zhang et al., 2007).
Knowledge of the molecular pathways underlyingbacterial response to plant compounds is importantfor understanding complex plant–pathogen interactionsand also for the development of new strategies formanaging bacterial infections in plants. It was previouslyshown that the carnivorous plant Dionaea muscipula
(Droseraceae) possesses strong antibacterial properties(Juniper et al., 1989; Finnie & van Staden, 1993; Krolickaet al., 2008). In this work, we examine the response ofPba to treatment with D. muscipula extract, its majorantimicrobial constituent, plumbagin, and a referenceantimicrobial peptide CAMEL, in order to gain insightinto the mechanism by which bacterial pathogens avoida plant’s chemical defences. Here, we show that thedifference in Pectobacterium atrosepticum (Pba) resistanceto plant antimicrobials observed in in vitro tests andin potato tubers can be at least partially explainedby the stimulation of Pba efflux pump expression andextracellular pectate lyase (Pel) activity by the combinedpresence of metabolites from potato tuber tissue andplumbagin.
Materials and methods
Bacterial strains and media
Pectobacterium atrosepticum (Pba) strain SCRI 1043 anda field isolate Pba 5A/1/2005 were from the collectionof Department of Plant Protection and Biotechnology,University of Gdansk, Poland. In this work, we usedonly one field-isolated strain of Pba as preliminary testsshowed similar susceptibility among different Pba isolatesto tested antimicrobials (data not shown). Bacteria weregrown in standard Luria broth (LB) (Sambrook et al.,1989) and in M63 buffer (Miller, 1972) with 0.2%polygalacturonic acid (Sigma, Munich, Germany) and0.2% glycerol (referred to as MPG medium) at 28°C. TheM63 buffer consisted of 2 g L−1 (NH4)2SO4, 13.6 g L−1
KH2PO4, 0.5 mg L−1 FeSO4·7H2O and 0.247 mg L−1
MgSO4·7H2O. The pH was adjusted to 7.0 with KOH.Where indicated, media were supplemented with 20%
(final concentration) potato tuber tissue filtrate (PF)(the supplemented media are referred to as LB + PFand MPG + PF, respectively). PF was prepared by adding0.02% diethyldithiocarbamic acid (Sigma) to a juice madefrom fresh potato tubers and then centrifuging and filtersterilising the resulting solution. Methanol potato tuberextract (PE) was prepared by adding 50 m L−1 of methanoland 0.01% HCl to 50 g of homogenised fresh potato tubertissue and incubating the mixture on a rotary shaker(100 rpm) for 2 h in 20°C. The extract was then passedthrough Whatman no. 1 filter paper and lyophilised,and the dry residue was dissolved in methanol andagain filtered. The obtained PE was applied to bacterialsuspensions at 100–1500 μg mL−1.
Dionaea muscipula extract and chemicals
Dionaea muscipula plants were obtained and grown asdescribed by Krolicka et al. (2008). Whole plants wereextracted with chloroform and a Soxhlet apparatus. Driedextract was dissolved in methanol to the concentration of30 mg dry weight (DW) per mL and filtered. In this work,we have also used plumbagin – a major antibacterialconstituent present in D. muscipula extract as shown byour preliminary studies of antibacterial activity of HighPerformance Liquid Chromatography (HPLC) fractions ofextract (data not shown). Extract and plumbagin (Sigma,Cat. no P7262) were added to the wells of 48-wellplates as methanol solutions, and the organic solventwas allowed to evaporate at room temperature beforemedium was added. Final concentrations of extract andplumbagin were ranged from 0.025 to 400 μg mL−1. Insome experiments, we used the antimicrobial, syntheticpeptide CAMEL (KWKLFKKIGAVLKVL), which is activeagainst Pba strains and was previously shown to be moreeffective against Pectobacterium spp. than streptomycin(Kamysz et al., 2005) to determine whether the effectsof plant extract and plumbagin were specific. CAMELwas synthesised as previously described (Kamysz et al.,2005) and dissolved in distilled water, filter sterilised andapplied at 0.025–16 μg mL−1.
Antibacterial activity of Dionaea muscipula extract,plumbagin and CAMEL
Minimal bactericidal concentrations (MBCs) of D. muscip-
ula extract, plumbagin and CAMEL were determined inLB medium as described (Thornsberry, 1991). For deter-mination of bacterial generation times, Pba cultures weregrown overnight in LB, centrifuged and suspended in LB,LB + PF, MPG or MPG + PF growth media at an OD600 of0.1; the cultures were grown at 28°C with shaking in 24-well plates for 24 h. Chloroform extract from D. muscipula,
Ann Appl Biol 164 (2014) 404–414 405© 2014 Association of Applied Biologists

Pectobacterium response to plant antimicrobial compounds A. Szpitter et al.
plumbagin and CAMEL was added to the cultures at 0.1and 0.25 MBC, and the absorbance was measured at 60-min intervals with a plate reader (Wallac Perkin Elmer,Turku, Finland). Generation times were calculated onthe basis of OD600 values measured during the exponen-tial phase of growth as previously described (Ulanowskaet al., 2007).
Extracellular pectate lyase activity in cell-free culturesupernatants as affected by plumbagin and CAMEL
Pectate lyase activity was measured in cell-free super-natants from cultures treated or untreated with D. mus-cipula extract or plumbagin at 0.1 and 0.25 MBC andwith or without PF. The extract and plumbagin wereadded at the start of culture or at the beginning of thestationary phase of growth. Extracellular Pel activity wasmeasured with a reaction buffer containing polygalactur-onic acid as described previously (Keen et al., 1984). Oneunit (U) of Pel activity was defined as the amount ofenzyme releasing an amount of oligogalacturonides thatincreased Abs235 by 1 unit during a 1-minute reaction.Pel activity was expressed as U g−1 of bacterial DW.
Plant tissue maceration assay and associatedextracellular Pel activity as affected by plumbaginand CAMEL
A maceration assay with potato tubers was performed asdescribed previously (Keen et al., 1984) with some mod-ifications. Briefly, an overnight culture of Pba5A/1/2005in LB was centrifuged and washed with M63 buffer, andthe cells were suspended at an OD600 of 0.075. Potatotubers were washed and surface sterilised with 5% cal-cium hypochlorite. Just before tuber inoculation, thebacterial suspension was mixed 1:1 (v/v) with twofoldconcentrated solutions of plumbagin or CAMEL in M63buffer. Five sterile tips containing 50 μL of bacterial sus-pension with plumbagin or CAMEL (at 1.0, 4.0 or 8.0MBC), suspension without these compounds, or M63buffer alone were inserted into each potato tuber. After48 h at 28°C, the decayed tissue surrounding each tipwas weighed, homogenised and diluted 1:4 (v/v) in M63buffer. Pba colony-forming unit (CFU) were counted aspreviously described (Jett et al., 1997). Extracellular Pelactivity was measured in diluted tissue homogenate asdescribed in the previous section and was expressed asU g−1 of macerated potato tissue.
Inhibition of DNA, RNA and protein synthesisby Dionaea muscipula extract and plumbagin
The influence of plumbagin, methanol and chloroformextracts from D. muscipula, and CAMEL on the synthesis
of DNA, RNA and proteins by Pba1043 was estimatedby measuring the incorporation of radioactive precursors([3H]thymidine, [3H]uridine or [3H]leucine, respectively)according to a previously described procedure (Wegrzynet al., 1991). Briefly, overnight cultures of Pba1043 werediluted 1:100 in fresh LB medium and cultivated to anOD600 of 0.1 at 28°C. [3H]thymidine, [3H]uridine or[3H]leucine was added to the cultures to final concen-trations of 5 μCi mL−1 of [3H]thymidine or [3H]uridineor 2 μCi mL−1 of [3H]leucine together with plumbagin,extract or CAMEL at 0.1 and 0.25 MBC. Control cultureswere supplemented only with radioactive precursors.For measurement of the radioactive compounds, 50 μLsamples of the bacterial suspensions were placed onWhatman no. 3 filter paper after 15, 30, 60 or 90 minof incubation, and were then transferred immediately toan ice-cold 10% trichloroacetic acid (TCA) for 10 min.Following washing once in 5% TCA and then twice in96% ethanol, the filters were dried, and radioactivity wasmeasured in a scintillation counter (Beckman LS3133P,Fullerton, CA, USA) (Ulanowska et al., 2006). Eachtreatment was examined in a separate experiment withthree biological replicates.
Morphology of Pectobacterium atrosepticum cellsas affected by plumbagin and CAMEL
In order to examine morphology of Pba1043 cells treatedwith antimicrobials bacteria were grown overnight inLB in 28°C with shaking before CAMEL at 1.0 MBC orplumbagin at 2.0 MBC was added. After 1 h at 28°Cwith shaking, bacteria were adsorbed onto carbon-coated grids (Sigma), stained with 1.5% uranyl acetate(Laboratory Reagent BDH, Chemicals LTD, Netherfield,UK), and immediately examined with a Philips CM100electron microscope (EM, FEI Company, Eindhoven, theNetherlands).
Construction of Pectobacterium atrosepticum reporterplasmids
Sequences (about 500 bp long) preceding transcriptionstart sites of acrA, ompX and emrA genes from Pba1043(Bell et al., 2004) were amplified using primers listedin Table 1. Each amplified sequence contained one ofthe following restriction sites: BamHI, HindIII, SalI orEcoRI. The obtained fragments and a pPROBE-GT Gmr
pVS1/p15a plasmid containing promoterless gfp genepreceded by a multicloning site (Miller et al., 2000) weredigested with appropriate endonucleases (Fermentas) andligated using T4 ligase (Fermentas, GMBH, St. Leon-Rot, Germany). The resulting plasmids (pPROBE-GTPacrA, pPROBE-GT PemrA and pPROBE-GT PompX) were
406 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists

A. Szpitter et al. Pectobacterium response to plant antimicrobial compounds
Table 1 Primers used in this studya
Primer name Primer sequence 5′ → 3′ Restriction site Product length (bp)
PacrA_F GTGAGTGTCGACAACGGTTAAACG SalI 542
PacrA_R ATTAAGCCGCCAGAATTCATCAGA EcoRI
PemrA_F ACGCGAAAGCTTCAGTGACAGGCG HindIII 522
PemrA_R TGCGGCGTCTGTGGATCCTGATTT BamHI
PompX_F GATAGAAAGCTTCGAGATAATGAT HindIII 520
PompX_R ACCAGCAAATGCGGATCCTGCGCC BamHI
aRestriction sites are underlined, and non-complementary bases are shown in bold.
electroporated into Pba 5A/1/2005 cells using Gene PulserXcell (BioRad, Hercules, CA, USA), and bacterial clonescarrying plasmids were selected on LB medium withgentamycin (50 μg mL−1).
Efflux pump expression as affected by testedcompounds
Plumbagin, CAMEL, PF and PE were added to the wellsof 96-well plates. For methanol solutions of plumbaginand PE, the solvent was evaporated. The liquid in eachwell was then increased to 100 μL by addition of LB.Final concentrations of compounds and extracts in thewells were: for plumbagin 0.025–0.5 μg mL−1, CAMEL0.025–1 μg mL−1, PE 100–1500 μg mL−1 and PF 2–20%.Cultures of Pba 5A/1/2005 containing different reporterplasmids were grown overnight at 28°C on LB medium
supplemented with 50 μg mL−1 gentamycin. The cultureswere diluted 1:100 in the same medium and grown to anOD600 of 0.5. Cells were washed once with LB and resus-pended in the same medium to an OD600 of 0.5. A 100 μLvolume of suspension was added to the plates containingthe tested compounds and extracts. Fluorescence ofthe suspension in each well was measured immediately(t = 0) and after 4 h at 28°C (t = 4) with a plate readerat λex = 405 nm and λem = 535 nm. The change influorescence is a measure of transcriptional activity of acorresponding promoter: PacrA, PemrA or PompX.
Statistical analysis
All data were analysed by a Student’s t-test. Differenceswere considered significant for P < 0.05. All experimentswere performed at least twice with similar results and
Table 2 In vitro growth of two Pba strains as affected by Dionaea muscipula extract, plumbagin and CAMEL.
Generation time (min)bin four mediac
Bacterial strain Compound addeda LB LB + PF MPG MPG + PF
Pba1043 Control 78 ± 2 79 ± 7 117 ± 9 67 ± 1Extract 0.1 MBC 307 ± 23 (4)d 128 ± 7 (1.6) 558 ± 261 (5) 126 ± 7 (2)Extract 0.25 MBC 1562 ± 442 (20) 303 ± 34 (4) No growth 351 ± 26 (5)Plumbagin 0.1 MBC 197 ± 41 (3) 109 ± 9 (1.4) 752 ± 90 (6) 129 ± 3 (2)Plumbagin 0.25 MBC 2187 ± 442 (28) 341 ± 23 (4) No growth 316 ± 1 (5)CAMEL 0.1 MBC 87 ± 2 (1.1) 78 ± 2 119 ± 3 66 ± 0.1CAMEL 0.25 MBC 81 ± 2 76 ± 3 115 ± 3 69 ± 2
Pba 5A/1/2005 Control 94 ± 3 73 ± 2 115 ± 2 68 ± 2Extract 0.1 MBC 147 ± 13 (1.6) 91 ± 5 (1.3) 131 ± 13 83 ± 6 (1.2)Extract 0.25 MBC 479 ± 85 (5) 168 ± 37 (2) 475 ± 36 (4) 197 ± 28 (3)Plumbagin 0.1 MBC 282 ± 4 (3) 91 ± 5 (1.3) 247 ± 39 (2) 106 ± 1 (1.6)Plumbagin 0.25 MBC 1127 ± 235 (12) 240 ± 10 (3) 1563 ± 442 (14) 262 ± 7 (4)CAMEL 0.1 MBC 96 ± 6 75 ± 4 119 ± 5 69 ± 2CAMEL 0.25 MBC 99 ± 0.3 71 ± 3 123 ± 2 (1.1) 68 ± 1
MBC, minimal bactericidal concentration; Pba, Pectobacterium atrosepticum; PF, potato filtrate.aThe MBC were 50 μg mL−1 for D. muscipula chloroform extract (Extract), 50 μg mL−1 for plumbagin and 8 μg mL−1 for CAMEL, for both Pba strains.bGeneration times were calculated on the basis of the trend in culture growth recorded for 24 h. Values are means ± SD of three culture replicates.cLB, Luria broth medium; MPG, M63 buffer with 0.2% polygalacturonic acid and 0.2% glycerol; LB + PF/MPG + PF, media supplemented with 20% potato
filtrate.dGiven in the brackets is the fold increase in generation time in relation to the respective control, presented only for generation times significantly
(P < 0.05) different from the control.
Ann Appl Biol 164 (2014) 404–414 407© 2014 Association of Applied Biologists

Pectobacterium response to plant antimicrobial compounds A. Szpitter et al.
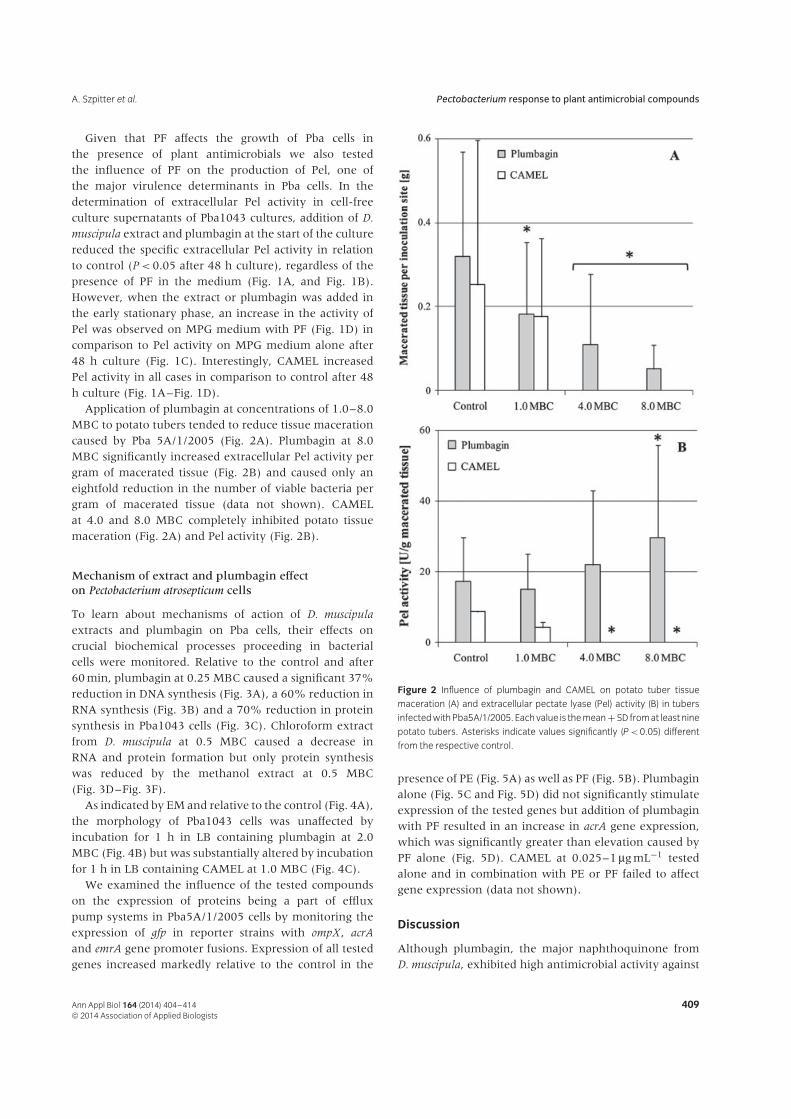
Figure 1 Specific extracellular pectate lyase (Pel) activity in Pba1043 culture supernatants as affected by Dionaea muscipula extract, plumbagin and
CAMEL. Bacteria were grown in MPG without potato filtrate (PF) (A, C) or with PF (B, D). A chloroform extract from D. muscipula (Extract), plumbagin and
CAMEL was added at the beginning of the culture (t = 0 h) (A, B) or in the early stationary phase (t = 10 h) (C, D) at 0.1 and 0.25 minimal bactericidal
concentration (MBC) for the extract and plumbagin and at 0.25 MBC for CAMEL. The results are means ± SD of three culture replicates. Pel activities for
control cultures grown for 12, 24 and 48 h on MPG medium were 3.8, 4.3 and 3.8 U g−1 dry weight (DW) of bacteria, respectively, and for those grown on
MPG + PF were 1.5, 1.7 and 1.2 U g−1 DW of bacteria, respectively.
only representative results from individual experimentsare presented. Standard deviations of the ratios (valuesrelative to the control) were estimated using the totaldifferential method.
Results
Effect of Dionaea muscipula extract, plumbaginand CAMEL on viability and virulence of Pectobacteriumatrosepticum
To assess the effects of D. muscipula extracts and plumbaginon viability of Pba, we have determined MBCs of them. Asa positive control, CAMEL – a known antimicrobial pep-tide – was used. The MBC values against both tested Pba
strains were 100 μg mL−1 for the D. muscipula chloroformextract, 50 μg mL−1 for the naphthoquinone plumbaginand 8 μg mL−1 for CAMEL. At 0.1 and 0.25 MBC, theextract and plumbagin increased Pba generation times by1.6- to 28-fold in LB medium and by 2- to 14-fold in MPGmedium (Table 2). However, for cultures supplementedwith extract and plumbagin at 0.25 MBC the addition ofpotato filtrate (PF) to the medium significantly reducedthe observed growth inhibition in comparison to control.The field isolate Pba 5A/1/2005 was more resistant thanPba1043 to D. muscipula extract and plumbagin at 0.25MBC in all media except for LB + PF (Table 2). CAMELpeptide at 0.1 and 0.25 MBC did not cause significantgrowth inhibition in the majority of Pba cultures (Table 2).
408 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists
A. Szpitter et al. Pectobacterium response to plant antimicrobial compounds
Given that PF affects the growth of Pba cells inthe presence of plant antimicrobials we also testedthe influence of PF on the production of Pel, one ofthe major virulence determinants in Pba cells. In thedetermination of extracellular Pel activity in cell-freeculture supernatants of Pba1043 cultures, addition of D.
muscipula extract and plumbagin at the start of the culturereduced the specific extracellular Pel activity in relationto control (P < 0.05 after 48 h culture), regardless of thepresence of PF in the medium (Fig. 1A, and Fig. 1B).However, when the extract or plumbagin was added inthe early stationary phase, an increase in the activity ofPel was observed on MPG medium with PF (Fig. 1D) incomparison to Pel activity on MPG medium alone after48 h culture (Fig. 1C). Interestingly, CAMEL increasedPel activity in all cases in comparison to control after 48h culture (Fig. 1A–Fig. 1D).
Application of plumbagin at concentrations of 1.0–8.0MBC to potato tubers tended to reduce tissue macerationcaused by Pba 5A/1/2005 (Fig. 2A). Plumbagin at 8.0MBC significantly increased extracellular Pel activity pergram of macerated tissue (Fig. 2B) and caused only aneightfold reduction in the number of viable bacteria pergram of macerated tissue (data not shown). CAMELat 4.0 and 8.0 MBC completely inhibited potato tissuemaceration (Fig. 2A) and Pel activity (Fig. 2B).
Mechanism of extract and plumbagin effecton Pectobacterium atrosepticum cells
To learn about mechanisms of action of D. muscipula
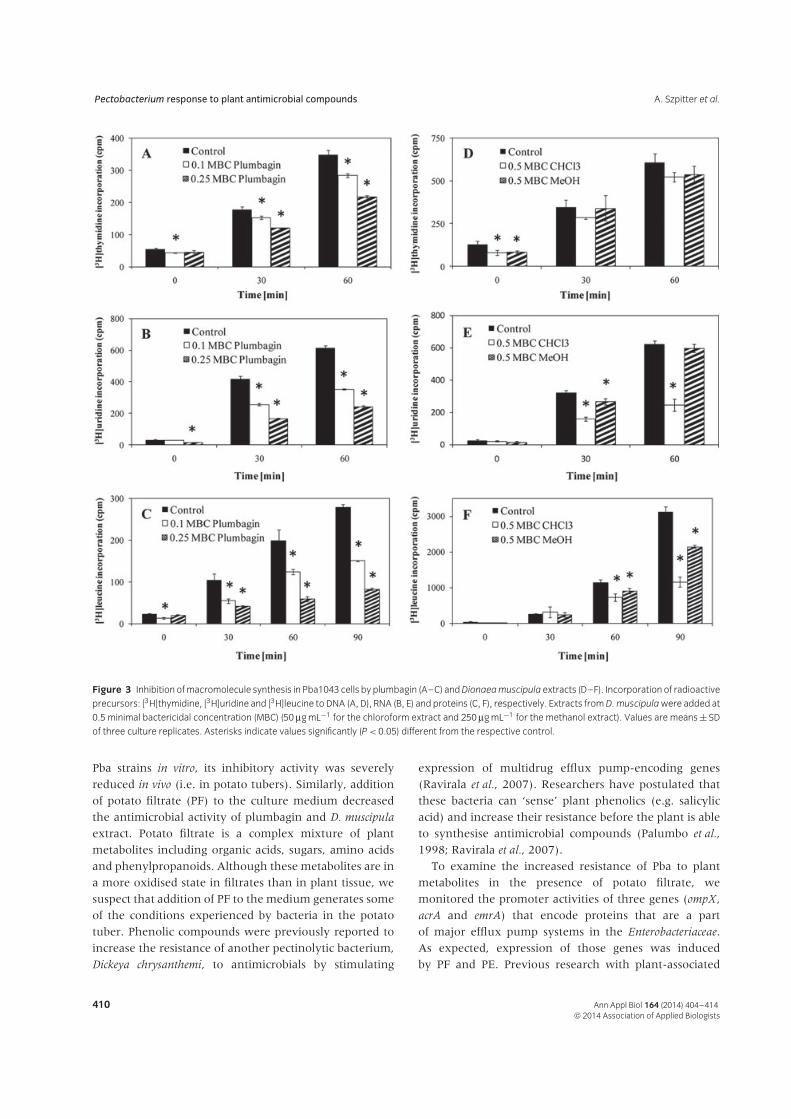
extracts and plumbagin on Pba cells, their effects oncrucial biochemical processes proceeding in bacterialcells were monitored. Relative to the control and after60 min, plumbagin at 0.25 MBC caused a significant 37%reduction in DNA synthesis (Fig. 3A), a 60% reduction inRNA synthesis (Fig. 3B) and a 70% reduction in proteinsynthesis in Pba1043 cells (Fig. 3C). Chloroform extractfrom D. muscipula at 0.5 MBC caused a decrease inRNA and protein formation but only protein synthesiswas reduced by the methanol extract at 0.5 MBC(Fig. 3D–Fig. 3F).
As indicated by EM and relative to the control (Fig. 4A),the morphology of Pba1043 cells was unaffected byincubation for 1 h in LB containing plumbagin at 2.0MBC (Fig. 4B) but was substantially altered by incubationfor 1 h in LB containing CAMEL at 1.0 MBC (Fig. 4C).
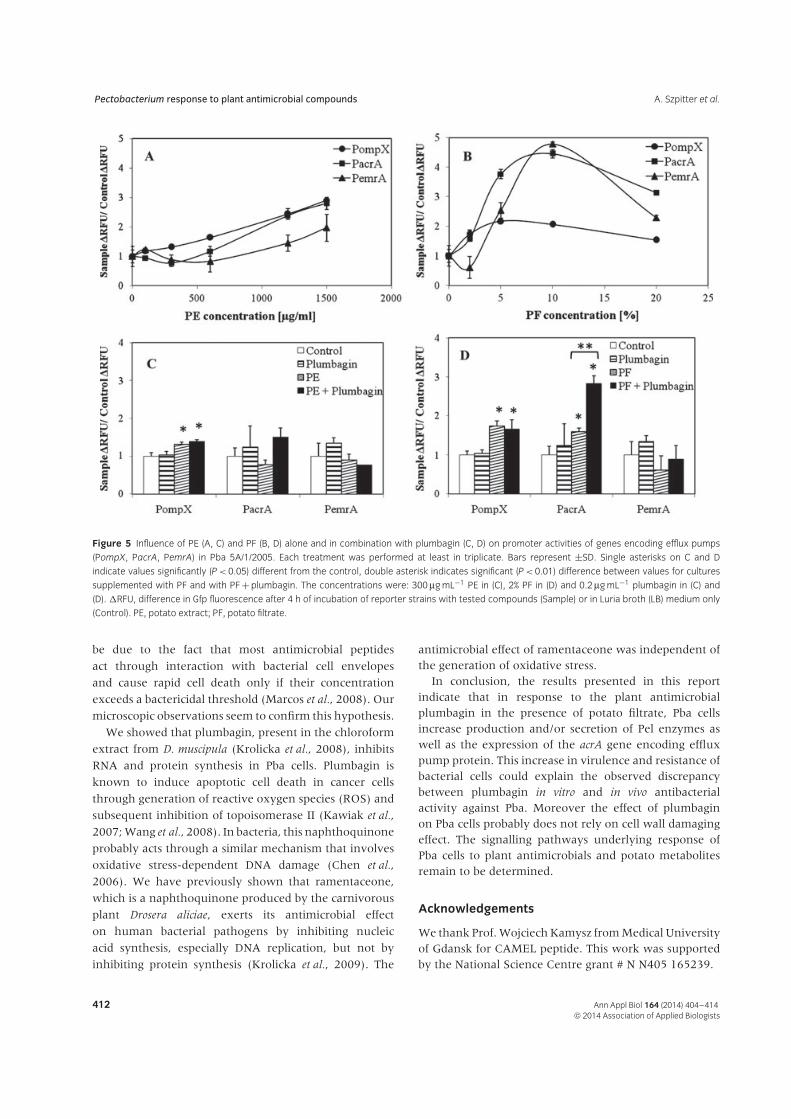
We examined the influence of the tested compoundson the expression of proteins being a part of effluxpump systems in Pba5A/1/2005 cells by monitoring theexpression of gfp in reporter strains with ompX, acrA
and emrA gene promoter fusions. Expression of all testedgenes increased markedly relative to the control in the
Figure 2 Influence of plumbagin and CAMEL on potato tuber tissue
maceration (A) and extracellular pectate lyase (Pel) activity (B) in tubers
infected with Pba5A/1/2005. Each value is the mean + SD from at least nine
potato tubers. Asterisks indicate values significantly (P < 0.05) different
from the respective control.
presence of PE (Fig. 5A) as well as PF (Fig. 5B). Plumbaginalone (Fig. 5C and Fig. 5D) did not significantly stimulateexpression of the tested genes but addition of plumbaginwith PF resulted in an increase in acrA gene expression,which was significantly greater than elevation caused byPF alone (Fig. 5D). CAMEL at 0.025–1 μg mL−1 testedalone and in combination with PE or PF failed to affectgene expression (data not shown).
Discussion
Although plumbagin, the major naphthoquinone fromD. muscipula, exhibited high antimicrobial activity against
Ann Appl Biol 164 (2014) 404–414 409© 2014 Association of Applied Biologists
Pectobacterium response to plant antimicrobial compounds A. Szpitter et al.
Figure 3 Inhibition of macromolecule synthesis in Pba1043 cells by plumbagin (A–C) and Dionaea muscipula extracts (D–F). Incorporation of radioactive
precursors: [3H]thymidine, [3H]uridine and [3H]leucine to DNA (A, D), RNA (B, E) and proteins (C, F), respectively. Extracts from D. muscipula were added at
0.5 minimal bactericidal concentration (MBC) (50 μg mL−1 for the chloroform extract and 250 μg mL−1 for the methanol extract). Values are means ± SD
of three culture replicates. Asterisks indicate values significantly (P < 0.05) different from the respective control.
Pba strains in vitro, its inhibitory activity was severely
reduced in vivo (i.e. in potato tubers). Similarly, addition
of potato filtrate (PF) to the culture medium decreased
the antimicrobial activity of plumbagin and D. muscipula
extract. Potato filtrate is a complex mixture of plant
metabolites including organic acids, sugars, amino acids
and phenylpropanoids. Although these metabolites are in
a more oxidised state in filtrates than in plant tissue, we
suspect that addition of PF to the medium generates some
of the conditions experienced by bacteria in the potato
tuber. Phenolic compounds were previously reported to
increase the resistance of another pectinolytic bacterium,
Dickeya chrysanthemi, to antimicrobials by stimulating
expression of multidrug efflux pump-encoding genes
(Ravirala et al., 2007). Researchers have postulated that
these bacteria can ‘sense’ plant phenolics (e.g. salicylic
acid) and increase their resistance before the plant is able
to synthesise antimicrobial compounds (Palumbo et al.,
1998; Ravirala et al., 2007).
To examine the increased resistance of Pba to plant
metabolites in the presence of potato filtrate, we
monitored the promoter activities of three genes (ompX,
acrA and emrA) that encode proteins that are a part
of major efflux pump systems in the Enterobacteriaceae.
As expected, expression of those genes was induced
by PF and PE. Previous research with plant-associated
410 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists

A. Szpitter et al. Pectobacterium response to plant antimicrobial compounds
Figure 4 Influence of plumbagin and CAMEL on Pba cell morphology as
indicated by electron microscopy. (A) Control culture (no addition); (B)
plumbagin at 2.0 minimal bactericidal concentration (MBC); (C) CAMEL
at 1.0 MBC. Pba1043 cells were incubated for 1 h in Luria broth (LB)
medium with the indicated compound. In (C), arrows indicate the abnormal
morphology caused by CAMEL. Bars at the lower right corner are 2 μm.
The experiment was performed twice and the results obtained in both
analyses were similar.
bacteria demonstrated that efflux pumps are importantfor establishing a successful plant–bacterium interactionand that they can be induced by plant compounds(Palumbo et al., 1998; Kang and Gross, 2005; Valecilloset al., 2006; Ravirala et al., 2007). The outer membraneprotein TolC, which is a part of the multidrug effluxsystem AcrAB-TolC, was crucial for colonisation of plantsby D. chrysanthemi, and tolC mutants were found tobe susceptible to a wide range of plant antimicrobials(Barabote et al., 2003). In addition, plant phytoalexinsincreased expression of the acrAB operon in anotherbacterial plant pathogen, Erwinia amylovora (Burse et al.,2004).
The present work documented a synergistic effect ofplumbagin and potato filtrate in the activation of acrAtranscription in Pba cells. We hypothesise that this effectcould result from the stimulation of the Mar regula-tory network governing bacterial cell resistance to stress,which has been identified in many species of Enter-obacteriaceae. In Escherichia coli, acrAB and tolC promoterscontain a marbox sequence recognised by MarA, SoxS andRob proteins. Expression of marA and soxS is stimulatedby phenolic compounds and oxidative stress, respec-tively (Martin et al., 1999; Ravirala et al., 2007). Othersignalling pathways and mechanisms were reported to beinvolved in the response of bacterial pathogens to plantmetabolites – GacA/S – RsmA/B (Yang et al., 2008) andregulators of the LuxR family (Zhang et al., 2007).
Although plumbagin inhibited potato tissue macerationcaused by Pba 5A/1/2005, it also stimulated productionand/or secretion of Pel enzymes in potato tissues. Inthe presence of PF, moreover, addition of D. muscipulachloroform extract and plumbagin to Pba1043 culturesin the stationary phase increased the extracellular Pelactivity in culture supernatants in relation to control(Fig. 1). In soft-rotting bacteria, the production ofextracellular enzymes, as well as proteins that constitutethe type II secretion system taking part in the exportof those enzymes out of the bacterial cell, is regulatedby a multitude of signalling pathways in response toenvironmental stimuli such as plant growth regulators orphenolic acids (Liu et al., 2008). The effect of plumbaginseems to be unspecific because CAMEL, an antimicrobialpeptide with a completely different mode of action, couldalso induce production and/or secretion of Pel enzymesin Pba cells. The 15-residue hybrid peptide CAMELwas previously reported to be an effective antimicrobialagainst a range of Pectobacterium and Dickeya species(Kamysz et al., 2005). Although it has strong bactericidalproperties against tested Pba strains, it did not inhibitgrowth in this study (Table 2) and had no influenceon macromolecule synthesis in Pba1043 cells at theconcentrations tested (results not shown). This could
Ann Appl Biol 164 (2014) 404–414 411© 2014 Association of Applied Biologists
Pectobacterium response to plant antimicrobial compounds A. Szpitter et al.
Figure 5 Influence of PE (A, C) and PF (B, D) alone and in combination with plumbagin (C, D) on promoter activities of genes encoding efflux pumps
(PompX, PacrA, PemrA) in Pba 5A/1/2005. Each treatment was performed at least in triplicate. Bars represent ±SD. Single asterisks on C and D
indicate values significantly (P < 0.05) different from the control, double asterisk indicates significant (P < 0.01) difference between values for cultures
supplemented with PF and with PF + plumbagin. The concentrations were: 300 μg mL−1 PE in (C), 2% PF in (D) and 0.2 μg mL−1 plumbagin in (C) and
(D). �RFU, difference in Gfp fluorescence after 4 h of incubation of reporter strains with tested compounds (Sample) or in Luria broth (LB) medium only
(Control). PE, potato extract; PF, potato filtrate.
be due to the fact that most antimicrobial peptides
act through interaction with bacterial cell envelopes
and cause rapid cell death only if their concentration
exceeds a bactericidal threshold (Marcos et al., 2008). Our
microscopic observations seem to confirm this hypothesis.
We showed that plumbagin, present in the chloroform
extract from D. muscipula (Krolicka et al., 2008), inhibits
RNA and protein synthesis in Pba cells. Plumbagin is
known to induce apoptotic cell death in cancer cells
through generation of reactive oxygen species (ROS) and
subsequent inhibition of topoisomerase II (Kawiak et al.,
2007; Wang et al., 2008). In bacteria, this naphthoquinone
probably acts through a similar mechanism that involves
oxidative stress-dependent DNA damage (Chen et al.,
2006). We have previously shown that ramentaceone,
which is a naphthoquinone produced by the carnivorous
plant Drosera aliciae, exerts its antimicrobial effect
on human bacterial pathogens by inhibiting nucleic
acid synthesis, especially DNA replication, but not by
inhibiting protein synthesis (Krolicka et al., 2009). The
antimicrobial effect of ramentaceone was independent ofthe generation of oxidative stress.
In conclusion, the results presented in this reportindicate that in response to the plant antimicrobialplumbagin in the presence of potato filtrate, Pba cellsincrease production and/or secretion of Pel enzymes aswell as the expression of the acrA gene encoding effluxpump protein. This increase in virulence and resistance ofbacterial cells could explain the observed discrepancybetween plumbagin in vitro and in vivo antibacterialactivity against Pba. Moreover the effect of plumbaginon Pba cells probably does not rely on cell wall damagingeffect. The signalling pathways underlying response ofPba cells to plant antimicrobials and potato metabolitesremain to be determined.
Acknowledgements
We thank Prof. Wojciech Kamysz from Medical Universityof Gdansk for CAMEL peptide. This work was supportedby the National Science Centre grant # N N405 165239.
412 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists

A. Szpitter et al. Pectobacterium response to plant antimicrobial compounds
References
Ball A.R., Casadei G., Samosorn S., Bremner J.B., Ausubel
F.M., Moy T.I., Lewis K. (2006) Conjugating berberine to
a multidrug resistance pump inhibitor creates an effective
antimicrobial. ACS Chemical Biology, 1, 594–600.
Barabote R.D., Johnson O.L., Zetina E., San Francisco
S.K., Fralick J.A., San Francisco M.J.D. (2003) Erwinia
chrysanthemi tolC is involved in resistance to antimicrobial
plant chemicals and is essential for phytopathogenesis.
Journal of Bacteriology, 185, 5772–5778.
Bell K.S., Sebaihia M., Pritchard L., Holden M.T.G., Hyman
L.J., Holeva M.C., Thomson N.R., Bentley S.D., Churcher
L.J.C., Mungall K., Atkin R., Bason N., Brooks K.,
Chillingworth T., Clark K., Doggett J., Fraser A., Hance Z.,
Hauser H., Jagels K., Moule S., Norbertczak H., Ormond D.,
Price C., Quail M.A., Sanders M., Walker D., Whitehead S.,
Salmond G.P.C., Birch P.R.J., Parkhill J., Toth I.K. (2004)
Genome sequence of the enterobacterial phytopathogen
Erwinia carotovora subsp. atroseptica and characterization
of virulence factors. Proceedings of the National Academy of
Sciences of the United States of America, 101, 11105–11110.
Burse A., Weingart H., Ullrich M.S. (2004) The phytoalexin-
inducible multidrug efflux pump AcrAB contributes to
virulence in the fire blight pathogen, Erwinia amylovora.
Molecular Plant–Microbe Interactions, 17, 43–54.
Chen J.W., Sun C.M., Sheng W.L., Wang Y.C., Syu
W.J. (2006) Expression analysis of up-regulated genes
responding to plumbagin in Escherichia coli. Journal of
Bacteriology, 188, 456–463.
Czajkowski R., Perombelon M.C.M., van Veen J.A., van der
Wolf J.M. (2011) Control of blackleg and tuber soft rot
of potato caused by Pectobacterium and Dickeya species: a
review. Plant Pathology, 60, 999–1013.
Finnie J.F., van Staden J. (1993) XII Drosera spp. (Sundew):
Micropropagation and the in vitro production of plubagin.
In Biotechnology and Agriculture and Forestry 24. Medicinal
and Aromatic Plants V , pp. 164–177. Ed. Y.P.S. Bajaj.
Berlin, Heidelberg, Germany; New York, NY; London, UK;
Paris, France; Tokyo, Japan; Hong Kong, China; Barcelona,
Spain; Budapest, Hungary: Springer-Verlag.
Hassan S., Hugouvieux-Cotte-Pattat N. (2011) Identification
of two feruloyl esterases in Dickeya dadantii 3937
and induction of the major feruloyl esterase and of
pectate lyases by ferulic acid. Journal of Bacteriology, 193,
963–970.
Hentzer M., Riedel K., Rasmussen T.B., Heydorn A.,
Andersen J.B., Parsek M.R., Rice S.A., Eber L., Molin
S., Høiby N., Kjelleberg S., Givskov M. (2002) Inhibition of
quorum sensing in Pseudomonas aeruginosa biofilm bacteria
by a halogenated furanone compound. Microbiology, 148,
87–102.
Jett B.D., Hatter K.L., Huycke M.M., Gilmore M.S. (1997)
Simplified agar plate method for quantifying viable
bacteria. Biotechniques, 23, 648–650.
Juniper B.E., Robins R.J., Joel D.M. (1989) The Carnivorous
Plants. London, UK/San Diego, CA, USA: Academic Press,
Inc. Harcourt Brace Jovanovich Publishers.
Kamysz W., Krolicka A., Bogucka K., Ossowski T., Lukasiak
J., Lojkowska E. (2005) Antibacterial activity of synthetic
peptides against plant pathogenic Pectobacterium species.
Journal of Phytopathology, 153, 313–317.
Kang H., Gross D.C. (2005) Characterization of a resistance-
nodulation-cell division transporter system associated
with the syr-syp genomic island of Pseudomonas syringae
pv. syringae. Applied and Environmental Microbiology, 71,
5056–5065.
Kawiak A., Piosik J., Stasilojc G., Gwizdek-Wisniewska A.,
Marczak L., Stobiecki M., Bigda J., Lojkowska E. (2007)
Induction of apoptosis by plumbagin through reactive
oxygen species-mediated inhibition of topoisomerase II.
Toxicology and Applied Pharmacology, 15, 267–276.
Keen N.T., Dahlbeck D., Staskawicz B., Belser W. (1984)
Molecular cloning of pectate lyase genes from Erwinia
chrysanthemi and their expression in Escherichia coli. Journal
of Bacteriology, 159, 825–831.
Krolicka A., Szpitter A., Gilgenast E., Romanik G., Kamin-
ski M., Lojkowska E. (2008) Stimulation of antibacterial
naphthoquinones and flavonoids accumulation in carniv-
orous plants grown in vitro by addition of elicitors. Enzyme
and Microbial Technology, 42, 216–221.
Krolicka A., Szpitter A., Maciag M., Biskup E., Gilgenast
E., Romanik G., Kaminski M., Wegrzyn G., Lojkowska
E. (2009) Antibacterial and antioxidant activity of the
secondary metabolites from in vitro cultures of Drosera
aliciae. Biotechnology and Applied Biochemistry, 53, 175–184.
Liu H., Coulthurst S.J., Pritchard L., Hedley P.E., Ravensdale
M., Humphris S., Burr T., Takle G., Brurberg M.B., Birch
P.R.J., Salmond G.P.C., Toth I.K. (2008) Quorum sensing
coordinates brute force and stealth modes of infection
in the plant pathogen Pectobacterium atrosepticum. PLoS
Pathogens, 4, e1000093.
Ma B., Hibbing M.E., Kim H.-S., Reedy R.M., Yedidia I.,
Breuer J., Breuer J., Glasner J., Perna N.T., Kelman A.,
Charkowski A.O. (2007) Host range and molecular phylo-
genies of the soft rot enterobacterial genera Pectobacterium
and Dickeya. Phytopathology, 97, 1150–1163.
Marcos J.F., Munoz A., Perez-Paya E., Misra S., Lopez-
Garcıa B. (2008) Identification and rational design of novel
antimicrobial peptides for plant protection. Annual Review
of Phytopathology, 46, 273–301.
Martin R.G., Gillette W.K., Rhee S., Rosner J.L. (1999)
Structural requirements for marbox function in tran-
scriptional activation of mar/sox/rob regulon promoters
in Escherichia coli: sequence, orientation and spatial rela-
tionship to the core promoter. Molecular Microbiology, 34,
431–441.
Miller J.H. (1972) Experiments in Molecular Genetics. Cold
Spring Harbor, NY: Cold Spring Harbor Laboratory
Press.
Ann Appl Biol 164 (2014) 404–414 413© 2014 Association of Applied Biologists

Pectobacterium response to plant antimicrobial compounds A. Szpitter et al.
Miller W.G., Leveau J.H., Lindow S.E. (2000) Improvedgfp and inaZ broad-host-range promoter-probe vectors.Molecular Plant–Microbe Interactions, 13, 1243–1250.
Palumbo J.D., Kado C.I., Phillips D.A. (1998) Anisoflavonoid-inducible efflux pump in Agrobacterium tume-
faciens is involved in competitive colonization of roots.Journal of Bacteriology, 180, 3107–3113.
Perombelon M.C.M. (2002) Potato diseases caused by softrot Erwinias: an overview of pathogenesis. Plant Pathology,51, 1–12.
Perombelon M.C.M., Kelman A. (1980) Ecology of thesoft rot Erwinias. Annual Review of Phytopathology, 18,361–387.
Ravirala R.S., Barabote R.D., Wheeler D.H., ReverchonS., Tatum O., Malouf J., Lui H., Pritchard L., HedleyP.E., Birch P.R., Toth I.K., Payton P.R.J., San FranciscoM.J.D. (2007) Efflux pump gene expression in Erwinia
chrysanthemi is induced by exposure to phenolic acids.Molecular Plant–Microbe Interactions, 20, 313–320.
Sambrook J., Fritsch E.F., Maniatis T. (1989) Molecular
Cloning: a Laboratory Manual. Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press.
Thornsberry C. (1991) Antimicrobial susceptibility testing:general considerations. In Manual of Clinical Microbiology.5th edn, pp. 1059–1201. Eds A. Balows, W.J. Hausler, K.L.Herrmann, H.D. Isenberg and H.J. Shadomy. WashingtonDC: American Society for Microbiology.
Ulanowska K., Tkaczyk A., Konopa G., Wegrzyn G. (2006)Differential antibacterial activity of genistein arising fromglobal inhibition of DNA, RNA and protein synthesis
in some bacterial strains. Archives of Microbiology, 184,271–278.
Ulanowska K., Majchrzyk A., Moskot M., Jakobkiewicz-Banecka J., Wegrzyn G. (2007) Assessment of antibacterialeffects of flavonoids by estimation of generation times inliquid bacterial cultures. Biologica Bratislava, 2, 132–135.
Valecillos A.M., Palenzuela P.R., Lopez-Solanilla E. (2006)The role of several multidrug resistance systems inErwinia chrysanthemi pathogenesis. Molecular Plant–Microbe
Interactions, 19, 607–613.Wang C.C., Chiang Y.M., Sung S.C., Hsu Y.L., Chang
J.K., Kuo P.L. (2008) Plumbagin induces cell cycle arrestand apoptosis through reactive oxygen species/c-Jun N-terminal kinase pathways in human melanoma A375.S2cells. Cancer Letters, 18, 82–98.
Wegrzyn G., Kwasnik E., Taylor K. (1991) Replication ofλ plasmid in amino acid-starved strains of Escherichia coli.Acta Biochimica Polanski, 38, 181–186.
Yang S., Peng Q., San Francisco M., Wang Y., Zeng Q., YangC.-H. (2008) Type III secretion system genes of Dickeya
dadantii 3937 are induced by plant phenolic acids. PLoS
One, 3, e2973.Zhang L., Jia Y., Wang L., Fang R. (2007) A proline iminopep-
tidase gene upregulated in planta by a LuxR homologueis essential for pathogenicity of Xanthomonas campestris pv.campestris. Molecular Microbiology, 65, 121–136.
Zimnoch-Guzowska E., Lojkowska E., Perombelon M. (2005)Resistance to bacterial pathogens. In Genetic Improvement of
Solanaceous Crops. Volume 1: Potato. Eds M.K. Razdan andA.K. Mattoo. Enfield, NH, USA: Science Publishers, Inc.
414 Ann Appl Biol 164 (2014) 404–414© 2014 Association of Applied Biologists