tap29: an trichosanthes kirilowii - proceedings of the national
TRANSCRIPT

Proc. Nadl. Acad. Sci. USAVol. 88, pp. 6570-6574, August 1991Biochemistry
TAP 29: An anti-human immunodeficiency virus protein fromTrichosanthes kirilowii that is nontoxic to intact cells
(antivira agent/AIDS/plant protein)
SYLVIA LEE-HUANG*t, PHILIP L. HUANG4§, HSIANG-FU KUNG¶, BAO-QUN LIII, PAUL L. HUANG*§,PETER HUANGt, HENRY I. HUANGt, AND HAO-CHIA CHEN***Department of Biochemistry, New York University School of Medicine, New York, NY 10016; *American Biosciences, New York, NY 10021; ILaboratoryof Biochemical Physiology, National Cancer Institute-Frederick Cancer Research and Development Center, Frederick, MD 21701; I'Biological CarcinogenicDevelopment Program, Program Resources, Inc., National Cancer Institute-Frederick Cancer Research and Development Center, Frederick, MD 21701;and **Endocrinology and Reproduction Research Branch, National Institute of Child Health and Human Development, Bethesda, MD 20892
Communicated by Bernhard Witkop, May 1, 1991 (received for review January 25, 1991)
ABSTRACT An anti-human immunodeficiency virus (anti-HIV) protein capable of inhibiting HIV-1 infection and repli-cation has been isolated and purified to homogeneity fromTrichosanthes krilowU. This protein, TAP 29 (Trichosanthesanti-HIV protein, 29 kDa), is distinct from trichosanthin [alsoknown as GLQ 223 (26 kDa)] in size, N-terminal amino acidsequence, and cytotoxicity. In addition to three conservativesubstitutions-namely, Arg-29 to Lys, Ile-37 to Val, and Pro-42to Ser-a total difference of residues 12-16 was found. TAP 29yielded -Lys-Lys-Lys-Val-Tyr-, whereas trichosanthin has-Ser-Ser-Tyr-Gly-Val-. Although the two proteins exhibit sim-ilar anti-HIV activity, as measured by syncytium formation,p24 expression, and HIV reverse transcriptase activity, theydiffer signlflcantiy in cytotoxicity, as measured by their effectson cellular DNA and protein syntheses. At the dose level of thebioassays, 0.34-340 nM, trichosanthin demonstrates a dose-dependent toxic effect on host cells. TAP 29 displays no toxiceffect, even at 100x ID50, whereas trichosanthin demonstrates38% and 44% inhibition on cellular DNA and protein synthe-sis, respectively. These results indicate that the therapeuticindex ofTAP 29 is at least two orders ofmagnitude higher thanthat of trichosanthin. Thus TAP 29 may offer a broader safedose range in the treatment of AIDS.
Trichosanthin, or GLQ 223, from Trichosanthes kirilowii hasbeen reported as an anti-human immunodeficiency virus(anti-HIV) agent (1). This protein is an active principle of thetraditional medicinal plant Trichosanthes (1-4). It has beenused for centuries in China for abortion and recently for thetreatment of trophoblastic tumors (2-4). The cytotoxic sideeffects of this compound have been well documented (5-7),raising concerns regarding the clinical safety of this drug (7).In this study, we report the isolation of an anti-HIV protein,TAP 29 (Trichosanthes anti-HIV protein, 29 kDa), fromTrichosanthes. Similar to the trichosanthin, TAP 29 is asingle-chain ribosome-inactivating protein (SCRIP). It inhib-its HIV-1 in vitro infection and replication. Distinct from thetrichosanthin, TAP 29 is not toxic-to intact host cells at itseffective doses. Here we report the physical, chemical, andbiological characterization of this anti-HIV agent, TAP 29.
MATERIALS AND METHODST. kirilowii. Root tuber of T. kirilowii was the source
material for the preparation of TAP 29 and trichosanthin.Fresh and dried T. kirilowii root tubers were supplied byAmerican Biosciences, New York.
Chemicals. Chromatographic media were obtained fromPharmacia LKB. [3H]Thymidine, [3H]leucine, and a rabbitreticulocyte lysate translation system were purchased fromDuPont/NEN.
Characterization of TAP 29. The homogeneity, size, andsubunit structure of TAP 29 and trichosanthin were deter-mined by SDS/PAGE (8) in the presence and absence of2-mercaptoethanol. The chemical composition of these com-pounds was determined by N-terminal amino acid sequenc-ing. Amino acid sequencing was carried out by Edmandegradation using an Applied Biosystems model 470A proteinsequencer with on-line phenylthiohydantoin analyzer.
Bioassays. The anti-HIV activity ofTAP 29 was measuredby microtiter syncytium formation in infectious cell centerassay (9), by viral core protein p24 expression (10), and byviral-associated reverse transcriptase (RT) activity (11).The CEM-ss (syncytium sensitive, Leu-3 positive) cell line
was used as the indicator cells for the microtiter syncytial-forming assay. The H9 cell line was used for p24 expressionand viral-associated RT activity assays in suspension cul-tures. HIV-1 virus was prepared and stocked as described(9). The cell lines were maintained in RPMI-1640 completemedium that contains 100 units ofpenicillin/streptomycin perml and 10% heat-inactivated fetal calf serum.
Microtiter Syncytium Formation Assay. Freshly preparedindicator cells in complete medium at 5 x 104 in 100 1.I weretreated with 100 p.l of TAP 29 or trichosanthin at variousconcentrations (Table 1) for 90 min. At the end of this time,100 ,ul of frozen pretitrated HIV stock from HxB3/H9 cells,corresponding to 100-300 syncytial-forming units, was addedfor 60 min. The supernatant containing anti-HIV protein andvirus was then removed and the cells were washed withcomplete medium to remove residual free drug and virus. Thecells were then plated onto poly(L-lysine)-coated microtiterwells with 200 ;LI of complete medium containing drug at thesame original concentration. The plates were incubated at370C in a humidified incubator with 5% CO2 in air. Focalsyncytium formation representing single infectious virionunits was scored at day 5 under an inverted microscope.
Viral Core Protein p24 and HIV-RT Assays. The effect ofTAP 29 on the in vitro replication and transmission of HIV-1was measured by p24 expression and HIV-RT activity. H9cells were inoculated with viral stock at a multiplicity ofinfection of 5 x 10-3. Cells at 5 x 107 per ml were incubatedwith the inoculum at 370C for 60 min to allow viral absorption.Unbound virus was removed by washing with medium. The
Abbreviations: HIV, human immunodeficiency virus; RT, reversetranscriptase; TCA, trichloroacetic acid; SCRIP, single-chain ribo-some-inactivating protein.tTo whom reprint requests should be addressed.§Present address: Harvard Medical School and Massachusetts Gen-eral Hospital, Boston, MA 02114.
6570
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 88 (1991) 6571
cells were then resuspended in complete medium and platedat 1 x i0s per ml in the presence or absence ofTAP 29 for theduration of the experiment. Under the assay conditions, viralproduction peaks at day 4. Cells were thus harvested at thistime, and cell-free supernatants were collected for the deter-mination of p24 production and HIV-RT activity. p24 wasassayed by RIA and expressed in ng/ml (10). HIV-RTactivity was measured by the incorporation of [3H]thymidinewith poly(rA)*p(dT)12-18 as primer-template, and the resultsare expressed in cpm/ml.
Cytotoxicity and Cell Viability. The cytotoxicity ofTAP 29and trichosanthin was measured by their effects on cellularsyntheses ofDNA and protein. Cells were pulse-labeled with1 ,uCi (1 Ci = 37 GBq) of [3H]thymidine or [3H]leucine 8 hrprior to harvesting at day 4. Cellular incorporation of labeledprecursor into trichloroacetic acid (TCA)-insoluble productwas determined by scintillation spectroscopy. Cell viabilitywas determined by trypan blue dye exclusion.
Ribosome-Inactivating Activity. Ribosome-inactivating ac-tivity was measured by in vitro translation using a rabbitreticulocyte lysate system (12) with globin message. The25-1.d reaction mixture contained 2 mM MgAc2, 80 mM KAc,2.5 mM spermidine, 34.5 mg of creatine phosphate per ml, 26mg of GTP per ml, 250 mM Hepes buffer, 1 ,uCi of [3H]leu-cine, and 1 ug of globin mRNA. Incubation was carried outat 37°C for 30 min. The product was separated from thereaction mixture by TLC in a solvent system of methanol/water/TCA/glacial acetic acid, 30:50:10:10 (vol/vol/wt/vol). The incorporation of [3H]leucine into TCA-precipitableproduct was measured by scintillation spectroscopy.
RESULTSPurification and Characterization. Step A: initial extrac-
tion. For routine preparation, 60 g of root tubers from T.kirilowii was ground into fine powder. This powder was thenextracted with 200 ml of a phosphate-buffered saline (PBS: 10mM sodium phosphate, pH 7.6/0.15 M NaCl) by stirringgently at 4°C overnight. The extract was first filtered throughtwo layers of cheesecloth and then cleared by centrifugationat 12,000 x g for 30 min. The supernatant was collected,dialyzed thoroughly against water, and finally dialyzedagainst 50 mM sodium phosphate buffer at pH 6.3 (buffer A).
Step B: CM-Sepharose CL 6B. The dialyzed solution fromstep A was centrifuged at 12,000 x g for 30 min to remove anyprecipitate formed during dialysis. The solution was concen-trated using a Centriprep 10 concentrator (Amicon) to 20 ml.This sample containing 164 mg of protein was loaded onto acolumn of CM-Sepharose CL 6B (1.5 x 36 cm), which waspreviously equilibrated with buffer A. The column waswashed with the same buffer until the baseline of A2,0 wasreached. The majority of the contaminating impurities wasexcluded from the column while TAP 29 and trichosanthinwere retained. They were eluted with a linear gradientconsisting of 250 ml of buffer A and 250 ml of buffer Acontaining 0.3 M NaCl. A typical elution profile is shown inFig. 1. The fractions were assayed for anti-HIV activity andribosome-inactivating activity. TAP 29 was eluted at about0.1-0.13 M NaCl. Trichosanthin was eluted between 0.15 and0.18 M NaCl. Trace amounts of contaminants of smallerproteins of about 16- to 11-kDa were present in these frac-tions. The bulk of the 16- to 11-kDa proteins was elutedbetween 0.22 and 0.28 M. The yields of TAP 29 and tricho-santhin were usually 5-7 mg and 16-24 mg, respectively. Atthis stage, the samples were about 75% pure.
Step C: Sephadex G-75 gelfiltration. The pooled sample ofTAP 29 from step B was dialyzed against 20 mM sodiumphosphate buffer at pH 7.2 (buffer B). The precipitate formedduring dialysis was removed by centrifugation at 12,000 rpmfor 30 min. The sample was then concentrated using a
1.0 I
I-0
0OD
06-
0.4 -
0.2 -
TAP Trip _
I'
06
0.4.a. 4Q-O LP~Pz1
0.2 s-~~~~~~~~~~~~~~~~~~~~~~~O
p0.3
- 0.2
CUz
-0.1
100
.-
0
-50 2
C
-0
0.
-0.50 200 220 240 260
Fraction
FIG. 1. Purification of TAP 29. The dialyzed and concentratedsample from step A, 20 ml containing 164 mg of protein, waschromatographed on a column of CM-Sepharose CL 6B (1.5 x 36cm). The elution gradient is shown as a solid line. Fractions of 6 mlwere collected at a flow rate of 36 ml/hr. Absorbance at 280 nm ofeach fraction was measured. Ribosome-inactivating activity (%inhibition) was assayed with 1 Al of a 1:100 dilution of the fractions.Active fractions were pooled as indicated by the horizontal bars(TAP, TAP 29; Tn, trichosanthin).
Centricon B15 concentrator (Amicon) to about 2 ml andsubjected to gel filtration on Sephadex G-75 superfine. Acolumn of 1.5 x 86 cm in buffer B was used in this case. Theflow rate was adjusted to 2 ml/hr and 1-ml fractions werecollected. Homogeneous TAP 29 was eluted at 0.48 columnvolume, corresponding to a molecular mass of about 29 kDa(Fig. 2). A small shoulder at the leading edge of the peakrepresents aggregates of this protein. The formation of ag-gregates during dialysis and gel filtration reduces the yieldconsiderably. Typically 1-2 mg of pure soluble material isobtained from this step. The small tail peak contains the 16-to 11-kDa proteins, which show neither anti-HIV nor ribo-some-inactivating activity.
Physical Characterization. The size, homogeneity, andsubunit structure of TAP 29 and trichosanthin as determinedby SDS/PAGE are shown in Fig. 3. Single bands of 29 kDaand 26 kDa were obtained for TAP 29 and trichosanthin,
1.0-1
0.8-
0coCY
0.6-
0.4 -
0.2-
0
ilbI'
;I'
I I
1.- -*Io - - - -- .
20 40 60 80 100 120
Fraction
FIG. 2. Gel filtration of TAP 29 on a Sephadex G-75 superfinecolumn. Pooled sample from step B was dialyzed against buffer B andconcentrated to 2 ml. This material containing 5 mg of protein wasapplied to a column of Sephadex G-75 superfine (1.5 x 86 cm)equilibrated with the same buffer. Elution was carried out at 2 ml/hrand 1-ml fractions were collected. Absorbance at 280 nm wasmeasured. TAP 29 was eluted as a homogeneous peak at an elutionvolume of about 0.48 corresponding to a molecular mass of about 29kDa.
Biochemistry: Lee-Huang et al.
-1=

6572 Biochemistry: Lee-Huang et al.
A B1 2 3 1 2 3
kDa92-66in_
45 -
31 - _
21 _
u
mm
14 _ _
FIo. 3. SDS/PAGE ofTAP 29. Electrophoresis was carried outin the presence (A) and absence (B) of the reducing agent 2-mercap-toethanol in 10%o acrylamide at a constant voltage of 90 V for 5 hr,until the bromophenol blue tracking dye reached 1 cm from the loweredge of the gel. The gels were stained with silver stain. (A) Samplestreated with 2-mercaptoethanol. Lane 1, molecular mass standards,2 ,ug each; lanes 2 and 3, TAP 29 and trichosanthin, 3 jLg each. (B)Samples without 2-mercaptoethanol treatment. Lane 1, TAP 29, 1.5,ug; lane 2, molecular mass standards, 2 ,ug each; lane 3, trichosan-thin, 1.5,g.
respectively, in the presence and absence of the reducingagent, indicating that these proteins are single-chain poly-peptides.N-Terminal Amino Acid Sequence. The sequence ofthe first
44 amino acids from the N terminus of TAP 29 is shown inFig. 4. It is different from the corresponding sequence oftrichosanthinj as determined by us and as reported by others(13, 15-17). In addition to three conservative substitutions atposition 29 (Arg to Lys), position 37 (Ile to Val), and position42 (Pro to Ser), a stretch of 5 amino acids from position 12 toposition 16 is distinct in TAP 29. This sequence is -Lys-Lys-Lys-Val-Tyr-, whereas the corresponding sequence in tri-chosanthin is -Ser-Ser-Tyr-Gly-Val-. This difference in se-quence results in three potential tryptic sites in TAP 29 notfound in trichosanthin. These three tryptic sites would givetwo meaningful tryptic fragments and a free lysine.
Inhibition of Syncytium Formation. The effects of TAP 29on the infectivity of HIV-1 were measured by the microtitersyncytium formation infectivity assay (9). This assay is basedon the interaction between fusigenic virus-infected cellsexpressing the HIV envelope gene products and uninfectedadjacent cells bearing CD4 molecules. It quantitates acutecell-free HIV-1 infection. Focal syncytium formation repre-senting single infectious virion units was scored at day 5 byexamination under an inverted microscope. The results oftwo independent experiments are summarized in Table 1. A90-min preincubation of the indicator cells with TAP 29resulted in a dose-dependent inhibition of HIV-1 infection
Table 1. Comparison of the effects of TAP 29 and trichosanthin(Tri) on HIV infection as measured by syncytium formation ininfectious cell center (ICC) assay
Syncytia, no.
Conc., per well % ICC CytotoxicitynM TAP 29 Tri TAP 29 Tri TAP 29 Trio 260 260 100 100 - -0.344 128 205 49 79 - -1.724 90 NS 35 NS - +3.44 65 NS 25 NS - +
17.24 38 NS 15 NS - +34.4 0 NS 0 NS - +
Values are averages of triplicates from two independent experi-ments. Triplicate wells of indicator cells containing TAP 29 ortrichosanthin at each concentration without virus were also includedfor the determination of the cytotoxicity of these proteins. The %ICC is expressed in terms of the average number of syncytia indrug-treated samples/the average number of syncytia in untreatedsamples. NS, no score of syncytia because of cytotoxicity.
and replication. At 0.344 and 34.4 nM, TAP 29 caused 51%and 100%o inhibition of syncytial formation, respectively. AnID50 of 0.34 nM was obtained from these results. No cyto-toxicity of TAP 29 was observed at these concentrations.Trichosanthin, on the other hand, exhibited significant toxiceffects on the indicator cells at concentrations above 1.5 nM.At 0.34 nM, the ID" of TAP 29, trichosanthin caused 21%inhibition of syncytium formation.
Inhibition of Viral Core Protein p24 Expression. The effectof TAP 29 on viral core protein p24 expression is shown inFig. 5. TAP 29 exhibited dose-dependent inhibition of HIV-1p24 expression; 47% and 97% inhibition were observed at0.34 and 34 nM, respectively. An ID50 of 0.37 nM wasobtained from these results. The reduced production of p24was not due to cytotoxic or cytostatic effects, and nodecrease in cellular DNA or protein synthesis was observedat these TAP 29 concentrations. At 340 nM, or 100Ox ID50,<30%o inhibition ofcellular incorporation of [3H]thymidine or[3HJleucine was obtained. A similar ID50 was found fortrichosanthin. However, as shown in Fig. 6, trichosanthinexhibited dose-dependent cytotoxicity to the host cells at theconcentration used in this assay.
Inhibition of Viral-Associated RT Activity. The effects ofTAP 29 on the activity of viral-associated RT are also shownin Fig. 5. In this assay poly(rA)-p(dT)12.18 was used astemplate-primer and [3H~dTTP was used as the substrate.Results are expressed as the polynucleotide incorporation of3H label in terms of cpm/ml. Here again, TAP 29 yieldeddose-dependent inhibition ofHIV-RT activity with an ID50 of0.46 nM. Trichosanthin, on the other hand, exhibited alimited effective concentration range. Although a similar ID50was observed, significant cytotoxic effects were apparent.
1 5 10TAP 29 Asp-Val-Ser-Phe-Arg-Leu-Ser-Gly-Ala-Thr-Ser-Lys-Lys-Lys-Tri Asp-Val-Ser-Phe-Arg-Leu-Ser-Gly-Ala-Thr-Ser-8er-8er-Tyr-
15 20 25TAP 29 Val-Tyr-Phe-Ile-Ser-Asn-Leu-Arg-Lys-Ala-Leu-Pro-Asn-Glu-Tri Gly-Val-Phe-Ile-Ser-Asn-Leu-Arg-Lys-Ala-Leu-Pro-Asn-Glu-
30 35 40TAP 29 Lys-Lys-Leu-Tyr-Asp-Ile-Pro-Leu-Val-Arg-Ser-Ser-xxx-ser-Tri Arg-Lys-Leu-Tyr-Asp-11e-Pro-Leu-Ile-Arg-Ser-Ser-Leu-Pro-
45TAP 29 Gly-Ser-Lys-Tri Gly-Ser-
FIG. 4. N-terminal amino acid sequence of TAP 29 and its comparison with trichosanthin (Tri). Boldface residues are unique amino acidsbetween these two proteins. Residue 34 was Leu in trichosanthin 1 reported by Zhang and Wang (13) and cited previously (14). Our analysisof trichosanthin yielding Ile at this position is consistent with that of a-trichosanthin determined by Chow et al. (15). xxx, Unknown amino acid.
Proc. Natl. Acad. Sci. USA 88 (1991)
Proc. Natl. Acad. Sci. USA 88 (1991) 6573
100 r
80
c
0
:.0RT
dThd
0.34 3.4 34 340
nM
FIG. 5. Effect ofTAP 29 on HIV-1 replication. The effect ofTAP29 on the replication ofHIV-1 was measured by viral core antigen p24expression and HIV-RT activity. p24 production was determined byRIA and is expressed in ng/ml. Viral RT activity was determined bythe incorporation of [3H]dTTP into TCA-precipitable products and isexpressed in terms of cpm x 103 per ml. These values in controlculture (without the addition ofTAP 29) were 2094 ng/ml and 7% x103 cpm/ml, respectively, as determined in triplicates in two inde-pendent experiments. Results are normalized to values obtained incontrol cultures. Standard deviations are indicated by error bars.dThd, thymidine.
Assays on Cytotoxicity. The cytotoxicity of the two anti-HIV proteins from Trichosanthes was compared by measur-ing their effects on cellular DNA and protein synthesis inuninfected H9 cells. The incorporation of [3H]thymidine or[3H]leucine into TCA-precipitable DNA or protein was ex-
c0.
-C._
._1
100
80
60
pressed as cpm/ml. A 103-fold concentration range wastested and the results are summarized in Fig. 6. From 0.34 to34 nM, TAP 29 caused no detectable effect on cellularincorporation of labeled thymidine or leucine, whereas p24production and HIV-RT activity were inhibited >90%o. At 340nM, or 1000x ID50, TAP 29 resulted in 22% and 30%reductions in cellular DNA or protein synthesis, respec-tively. Thus TAP 29 has a therapeutic index of at least 1000.Trichosanthin, on the other hand, showed dose-dependentinhibition of cellular DNA and protein synthesis. At 3.4 nM(10x ID50) trichosanthin caused about 26% and 31% inhibi-tion of the incorporation of [3H]thymidine and [3H]leucine.These results suggest that the therapeutic index ofTAP 29 isat least two orders of magnitude higher than that of tricho-santhin.
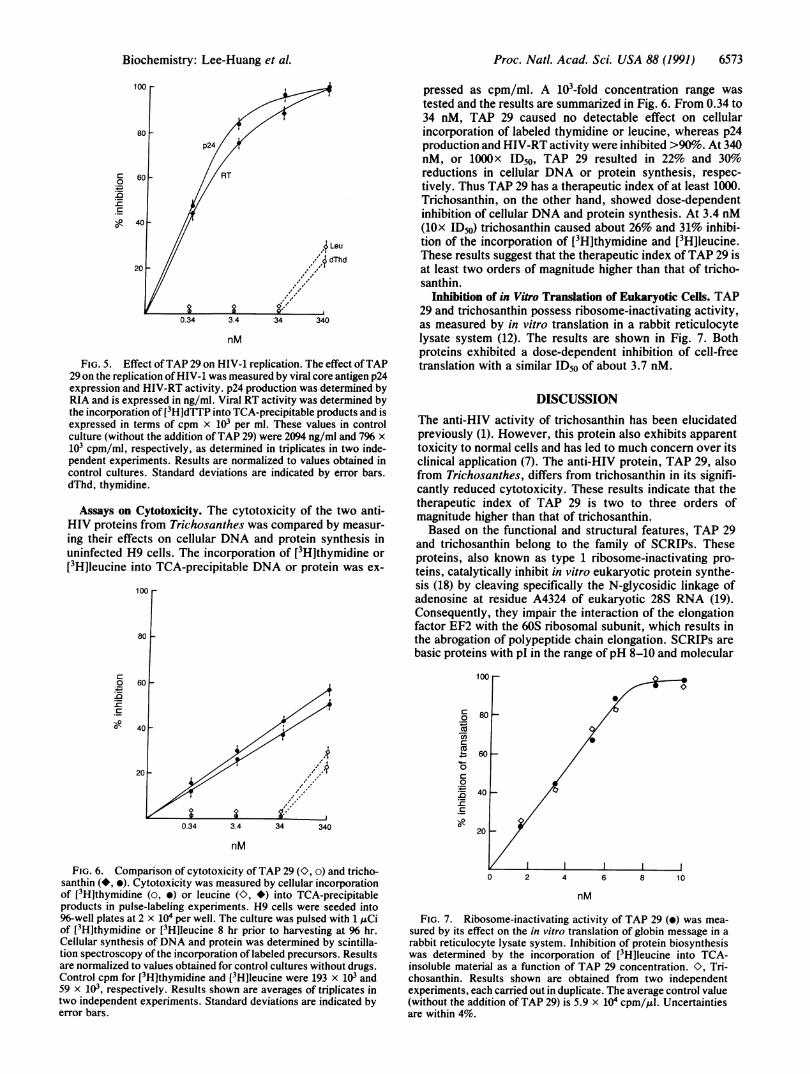
Inhibition of in Vitro Translation of Eukaryotic Cells. TAP29 and trichosanthin possess ribosome-inactivating activity,as measured by in vitro translation in a rabbit reticulocytelysate system (12). The results are shown in Fig. 7. Bothproteins exhibited a dose-dependent inhibition of cell-freetranslation with a similar ID50 of about 3.7 nM.
DISCUSSIONThe anti-HIV activity of trichosanthin has been elucidatedpreviously (1). However, this protein also exhibits apparenttoxicity to normal cells and has led to much concern over itsclinical application (7). The anti-HIV protein, TAP 29, alsofrom Trichosanthes, differs from trichosanthin in its signifi-cantly reduced cytotoxicity. These results indicate that thetherapeutic index of TAP 29 is two to three orders ofmagnitude higher than that of trichosanthin.Based on the functional and structural features, TAP 29
and trichosanthin belong to the family of SCRIPs. Theseproteins, also known as type 1 ribosome-inactivating pro-teins, catalytically inhibit in vitro eukaryotic protein synthe-sis (18) by cleaving specifically the N-glycosidic linkage ofadenosine at residue A4324 of eukaryotic 28S RNA (19).Consequently, they impair the interaction of the elongationfactor EF2 with the 60S ribosomal subunit, which results inthe abrogation of polypeptide chain elongation. SCRIPs arebasic proteins with pI in the range ofpH 8-10 and molecular
CO0
CO
0
0
.0-C
Nool0.34 3.4 34 340
nM
FIG. 6. Comparison of cytotoxicity ofTAP 29 (., o) and tricho-santhin (*, *). Cytotoxicity was measured by cellular incorporationof [3H]thymidine (o, *) or leucine (O, *) into TCA-precipitableproducts in pulse-labeling experiments. H9 cells were seeded into96-well plates at 2 x 104 per well. The culture was pulsed with 1 ,Ciof [3H]thymidine or [3H]leucine 8 hr prior to harvesting at 96 hr.Cellular synthesis of DNA and protein was determined by scintilla-tion spectroscopy of the incorporation of labeled precursors. Resultsare normalized to values obtained for control cultures without drugs.Control cpm for [3H]thymidine and [3H]leucine were 193 x 103 and59 X 103, respectively. Results shown are averages of triplicates intwo independent experiments. Standard deviations are indicated byerror bars.
100
80
60
0 2 4 6 8 10
nM
FIG. 7. Ribosome-inactivating activity of TAP 29 (e) was mea-sured by its effect on the in vitro translation of globin message in arabbit reticulocyte lysate system. Inhibition of protein biosynthesiswas determined by the incorporation of [3H]leucine into TCA-insoluble material as a function of TAP 29 concentration. O. Tri-chosanthin. Results shown are obtained from two independentexperiments, each carried out in duplicate. The average control value(without the addition of TAP 29) is 5.9 x 104 cpm/,iul. Uncertaintiesare within 4%.
Biochemistry: Lee-Huang et al.

6574 Biochemistry: Lee-Huang et al.
mass of about 24-33 kDa. In addition to trichosanthin andTAP 29, other examples of SCRIPs include Momordicainhibitors (20, 21), dianthins (22), the pokeweed antiviralproteins (24-26), and MAP 30, a SCRIP that we recentlyreported (14).Among these SCRIPs, trichosanthin (1), pokeweed antivi-
ral proteins (23), MAP 30 (14), and TAP 29, as shown here,are reported to possess anti-HIV activity. It is interesting tonote that of these anti-HIV SCRIPs, trichosanthin appears toexhibit significant cytotoxicity, whereas MAP 30 and TAP 29show negligible cytotoxicity. In recent studies, as will bereported elsewhere, we have found that three other SCRIPs,DAPs 30 and 32 and GAP31, also exhibit anti-HIV activitywith low cytotoxicity. Together, these findings emphasizeremarkable similarities among SCRIPs in antiviral activityand distinct differences in cytotoxicity to host cells. It is mostintriguing that our two highly purified anti-HIV proteins fromTrichosanthes have such a great difference in cytotoxicity.Since these protein preparations are homogeneous, the dif-ference in their cytotoxicity is unlikely to be due to contam-inants.
It is important to understand the structure-function rela-tionships of these molecules in terms of their various bioac-tivities and cytotoxicity. Such studies would provide usefulinsight into the molecular mechanism ofthe anti-HIV activityof these SCRIPs as well as valuable information for thera-peutic development of nontoxic drugs from these molecules.It is likely that unique domains are involved in each of thespecific activities of these molecules. For example, theanti-HIV activity of these SCRIPs may not necessarily func-tion through ribosome inactivation as these proteins do notaffect cellular DNA and protein synthesis at the concentra-tions at which they exert their antiviral activity. Similarly,unique motifs may be involved in cytotoxicity.From our 40 plus N-terminal amino acid sequence analyses
of MAP 30 (14), TAP 29, and trichosanthin, extensive se-quence homology is evident among them. However, thesequence -Lys-Lys-Lys-Val-Tyr-, at positions 12-16 in TAP29, differs from -Ser-Ser-Tyr-Gly-Val- at the correspondingposition in trichosanthin. These distinctive basic residues inTAP 29 favor tryptic-type cleavages, whereas similar sitesare absent in trichosanthin. Interestingly, the correspondingsequence in MAP 30, another nontoxic anti-HIV protein, is-Lys-Thr-Tyr-Thr-Lys- (14), which also has two lysine resi-dues to accommodate tryptic cleavages. Similarly, gelonion,yet another nontoxic anti-HIV SCRIP, also contains a trypticcleavage site near this region of the sequence (unpublisheddata), whereas the A chains of the toxin ricin lack basicresidues in the same segment of the sequence. Whether thepresence of basic residues in the unique sequences plays arole in reducing the cytotoxicity of these anti-HIV proteinsawaits further study. Nevertheless, this feature offers animportant clue for the site-specific mutation and modificationstudies related to the structure-function of these antiviralproteins. Furthermore, the complete elucidation of primarysequence and the understanding of secondary structure ofthese proteins as well as the development of structure-function studies may lead to the rational design in focusing ona safer and highly effective future generation of anti-HIVdrugs.
We express our thanks to Drs. Han Fang and Chang Shai Li fortheir help in growing the Trichosanthes plants. We thank Drs. HenryLee and Margaret Song for their contribution of different varieties ofT. kirilowii. We also thank Drs. C. K. Lee and Cheng Hsuan for theirinterest and helpful discussions. S.L.-H. acknowledges partial sup-port of her work by BRSG S07 RR05399-28, awarded by theBiomedical Research Support Grant Program, Division of ResearchResources, National Institutes of Health.
1. McGrath, M. S., Hwang, K. M., Caldwell, S. E., Gaston, I.,Luk, K.-C., Wu, P., Ng, V. L., Crowe, S., Daniels, J., Marsh,J., Deunhart, T., Lekas, P. V., Vennari, J. C., Yeung, H.-W.& Lifson, J. D. (1989) Proc. Natl. Acad. Sci. USA 86, 2844-2848.
2. Li, S. C. (15%) Pen Ts'ao Kang Mu (Chinese PharmaceuticalCompendium); reprinted (1977) by People's Medical PublishingHouse, Beijing.
3. Cheng, K. F. (1982) Obstet. Gynecol. (NY) 59, 494-498.4. Chan, W. Y., Tam, P. P. & Yeung, W. H. (1984) Contracep-
tion 29, 91-100.5. Qian, R. Q., Gu, Z. W., Zhang, X. L., Zhu, S. Q., Zhang,
W. Q., Fu, Y. H., Weng, X. Q., Wu, Y. W., Liu, Y. F., Xu,S. Z., Lin, S. W., Jin, S. W., Tian, G. Y. & Wang, Y. (1981)Acta Chim. Sin. (Chinese) 39, 927-931.
6. Gu, Z. W., Qian, R. Q., Jin, S. W., Qian, W. W., Xu, S. Z.,Zhang, L. Q., Zhang, X. L., Yao, Y. Z., Liu, Y. F., Zhu,S. Q., Cao, B. S., Wang, S. F., Wang, Q. H., Zhang, W. J.,Liu, Y. Z., Fu, Y. H. & Wang, Y. (1984) Acta Chim. Sin.(Chinese) 43, 943-945.
7. Palcca, J. (1990) Science 247, 1406.8. Laemmli, U. K. (1969) Nature (London) 227, 680-685.9. Nara, P. L. & Fischinger, P. J. (1988) Nature (London) 332,
469-470.10. Nara, P. L., Hatch, W. C., Dunlop, N. M., Robey, W. G.,
Arthur, L. O., Gonda, M. A. & Fischinger, P. J. (1987) AIDSRes. Hum. Retroviruses 3, 283-302.
11. Hoffman, A. D., Banapour, B. & Levy, J. A. (1985) Virology147, 326-335.
12. Pelham, R. B. & Jackson, R. J. (1976) Eur. J. Biochem. 67,247-256.
13. Zhang, X. & Wang, J. (1986) Nature (London) 321, 477-478.14. Lee-Huang, S., Huang, P. L., Nara, P. L., Chen, H. C., Kung,
H.-F., Huang, P., Huang, H. I. & Huang, P. L. (1990) FEBSLett. 272, 12-18.
15. Chow, T. P., Feldman, R. A., Lovett, M. & Piatak, M. (1990)J. Biol. Chem. 265, 8670-8674.
16. Maraganore, J. M., Joseph, M. & Bailey, M. C. (1987) J. Biol.Chem. 262, 11628-11633.
17. Collins, E. J., Robertus, J. D., LoPresti, M., Stone, K. L.,Williams, K. R., Wu, P., Hwang, K. & Piatak, M. (1990) J.Biol. Chem. 265, 8665-8669.
18. Stripe, F. & Barbieri, L. (1986) FEBS Lett. 195, 1-8.19. Endo, Y. & Tsurugi, K. (1987) J. Biol. Chem. 262, 8128-8130.20. Barbierei, L., Zamboni, M., Lorenaoni, E., Montanaro, L.,
Sperti, S. & Stripe, F. (1980) Biochem. J. 186, 443-452.21. Jimenez, A. & V~zquez, D. (1985) Annu. Rev. Microbiol. 39,
649-672.22. Stirpe, F., Williams, D. G., Onyon, L. J. & Legg, R. F. (1981)
Biochem. J. 195, 399-405.23. Zarling, J. M., Moran, P. A., Haffar, O., Sias, J., Richman,
D. D., Spina, C. A., Myers, D. E., Kuebelbeck, V., Ledbetter,J. & Uckun, F. M. (1990) Nature (London) 347, 92-95.
24. Irvin, J. D. (1975) Arch. Biochem. Biophys. 169, 522-528.25. Irvin, J. D., Kelly, T. & Robertus, J. D. (1980) Arch. Biochem.
Biophys. 200, 418-425.26. Barbieri, L., Aron, G. M., Irvin, J. D. & Stripe, F. (1982)
Biochem. J. 203, 55-59.
Proc. Natl. Acad. Sci. USA 88 (1991)