temporal and spatio-temporal attention to tactile stimuli in extinction patients
TRANSCRIPT

INTRODUCTION
Extinction is a neurological disorder particularlyfrequent after cortical and subcortical right sidedbrain lesions (Barbieri and De Renzi, 1989; Vallaret al., 1994). First described by Anton (1899), thephenomenon of extinction implies that patientsdetect accurately a single contralesional stimulusbut fail to detect it when presented simultaneouslywith a similar ipsilesional stimulus. Although acontralesional sensory impairment may co-occur, itis widely agreed that extinction reflects a higher-order deficit of spatial selective attention ratherthan a simple sensory deficit.
Neglect and extinction disorders have beenclassically interpreted as the consequence of a spatialattention deficit. However, recent studies suggest thepresence of temporal deficits in patients with neglectand extinction. Temporal processing deficit seems tooccur independently of spatial processingimpairment. These results suggest that purely spatialtheories of neglect and extinction are no longersufficient to interpret the deficits. Several recentneuropsychological evidence suggest that disordersof spatial attention like extinction and neglect alsoinvolve deficits of spatio-temporal processing(Rorden et al., 1997; di Pellegrino et al., 1997, 1998;Husain et al., 1997; Karnath et al., 2002; Cate andBehrmann, 2002; Guerrini et al., 2003). Spatio-temporal interactions had been studied in patients
with left-sided visual (di Pellegrino et al., 1997;Baylis et al., 2002) and tactile extinction (Guerrini etal., 2003). In all experiments single stimuli werepresented to the left or to the right hemifield whiledouble stimuli were presented bilaterally,simultaneously or sequentially at different StimulusOnset Asynchronies (SOAs). In all experiments,extinction was maximal when double stimulioccurred simultaneously. Performance for thecontralateral stimuli improved with longer SOAs.These results not only show that a stimulus presentedin the ipsilesional field interferes with the processingof a stimulus presented contralesionally, but also thatthe interference is independent from the sequence ofthe stimuli presentation. Finally, the interference ismaximal at SOA 0 msec and decreases with SOAincreasing. In the last few years, several studiesfocused on purely temporal aspects of attentionmechanisms in healthy subjects as well as in patientswith neglect and extinction (di Pellegrino et al., 1997,1998; Duncan et al., 1994; Husain et al., 1997; Coulland Nobre 1998; Miniussi et al., 1999; Coull et al.,2000; Nobre, 2001; Griffin et al., 2002; Guerrini etal., 2003). There is converging evidence to indicatethat competition among stimuli may depend merelyon the timing of their respective occurrences and beat least partly independent from spatial variables. Theclassical paradigm employed to study temporalattentional mechanisms independently from spatialprocessing demands is the Rapid Serial Visual
Cortex, (2006) 42, 17-27
RESEARCH REPORT
TEMPORAL AND SPATIO-TEMPORAL ATTENTION TO TACTILE STIMULIIN EXTINCTION PATIENTS
Chiara Guerrini1 and Salvatore M. Aglioti2
(1Clinical Neuroscience Centre and Department of Psychology, University of Hull, Hull UK;2Department of Psychology, University “La Sapienza” and Centre of Neuropsychology Research, IRCCS,
Foundation S.ta Lucia, Rome, Italy)
ABSTRACT
The temporal deployment of attention to tactile stimuli delivered to the same or to a symmetrical position of the bodywas assessed in 6 Right-Brain Damaged (RBD) patients with left tactile extinction and 6 healthy controls. Two differenttasks called Temporal (T) and Spatio-Temporal (ST) extinction were used. In the T task single or double electro tactilestimuli were delivered to the same point to the left or, in separate blocks, to the right index finger. Double stimuli wereseparated by different Stimulus Onset Asynchronies (SOAs). In the ST task, stimuli could be single (left or right) or double(left and right). Double stimuli were delivered to the index finger of both hands simultaneously or sequentially. In bothtasks subjects were asked to report the number of the stimuli they perceived. In the ST task, subjects were also requestedto report the stimuli location. Results show that in both tasks RBD patients’ detection of left sided stimuli was significantlylower than of right sided stimuli detection, mainly at shortest SOAs. Moreover, detection of left sided stimuli was higherwhen two stimuli were delivered in sequence and in symmetrical body areas and in different sides of the space than whenstimuli were delivered in sequence in the same body area. Results suggest that the interaction between spatial and temporalvariables enhances the ability of tactile extinction patients to detect left sided stimuli.
Key words: human, tactile, extinction, spatial attention, temporal attention

Presentation (RSVP) (Broadbent and Broadbent,1987; Shapiro et al., 1994; Raymond et al., 1992;Duncan et al., 1994; Chun and Potter, 1995; Husainet al., 1997; Rizzo et al., 2001; di Pellegrino et al.,1998). This task may be viewed as an attentionalprocessing task operating in the temporal rather thanin the spatial domain and may be used to investigatethe temporal characteristics of perceptual andattentional processes. In the single-target RSVPparadigm stimuli (usually letters) are presentedbriefly in the same location and in rapid succession.Typically the target item is embedded in a stream ofstimuli and is differentiated in some way (forexample it has a different colour). The subjects’ taskis to identify the target. Results from single-targetRSVP procedures show that target identificationrequires the conjoining of the target definingcharacteristic (for example, the colour) with the to bereported feature (e.g., letter identity). Featureconjunction in this type of task requires attention andtakes approximately 100 msec. One may ask whetherafter the processing of a first target the attentionalsystem is free and is available to process thesubsequent stimulus. Broadbent and Broadbent(1987) employing a visual task argued that althoughsingle target results suggest that visual processing ofthe target is completed by 100 msec after itsoccurrence, the results from dual-target conditionsindicate that target processing or some process thatoccur as a result of target processing extends wellbeyond this time. Multiple-task RSVP procedure, inwhich subjects’ task is to identify two or more targetspresented in sequence, responds to this question.Basically, the idea is that sensorial systems havelimited capacity of processing and this task has beenused to probe the processing limits of the visualsystem, measuring the time course of attentionalprocessing. Employing this procedure it wasdemonstrated not only that identifying a targetrequires attention but also that this process lasts intime. In fact, the identification of a first stimulusgenerates a deficit in the identification of the stimuluspresented after the target. Studies in normal subjectsshow that employing both an identification or adetection task in a RSVP procedure, the processingof the first visual stimulus occupies the attentionalcapacity and consequently continues to interfere witha second stimulus presented shortly thereafter forseveral hundreds of milliseconds (300 msec)(Shapiro et al., 1994; Raymond et al., 1992; Duncanet al., 1994). This phenomenon is called AttentionalBlink or Attentional Dwell Time and may beconsidered a measure of the attentional capacities intemporal domain independently of the spatial domainin normal subjects (Chun and Potter, 1995). Thislimited capacity of the attentional system is aphysiological restriction, some studies suggest thatafter right parietal damage this restriction is evenincreased. Consequently, the identification process ofa target in patients with right parietal damage andextinction or neglect requires more time than in
18 Chiara Guerrini and Salvatore M. Aglioti
healthy control subjects. In a study with a right braindamage extinction patient di Pellegrino et al. (1998)for example, found that the identification of twoletters displayed in rapid succession and at differentSOAs, in the ipsilesional hemifield or in thecontralesional hemifield, is a process which has anabnormal duration in the contralesional hemifieldbeing at least twice as long as in the ipsilesionalhemifield. Similarly, Husain et al. (1997), employinga RSVP task showed that the duration of theAttentional Blink in patients with neglect is threetimes as long as in subjects without neglect. Theirdata suggest that neglect as well as extinction may beconsidered not purely a spatial deficit but a disorderthat involves also the temporal dimension of theattentional processing. A similar phenomenon, namedRepetition Blindness (RB), was firstly reported byKanwisher (1987). The RB phenomenon is observedin normal subjects and is measured with the RSVPprocedure. Typically, a rapid sequence of lettersforming a word or a sentence is presented in the sameplace of the screen. Subject’s task is to report theword or the sentence. In half of the trials an item isrepeated. When two items of the sequence have thesame identity, the second item is missed. Kanwisher(1987) suggested that when two identical eventshappen too close in time, the subjects fail todifferentiate the events as distinct instances. In otherwords, the second occurrence of a repeated itemactivates the same type node that is, the same abstractconceptual representation in memory. However, theinstances fail to be encoded as a distinct event or as aseparate token in episodic memory because of aspecific difficulty in rapid constructing of multipletokens of the same type. In other words, repetition atshort lags impairs token individuation. Kanwisher’stheory support the idea that information processingmay be compromised by simultaneity of differentevents while subjects’ performance may be improvedat increasing asynchronies intervals between twoevents. From this point of view, extinction may beconsidered as a pathological accentuation of the RBeffect. An interesting parallelism is proposed byBaylis et al. (1993) between the RB phenomenon andextinction. In their experiment, five patients withvisual extinction were presented with coloured letterssimultaneously in both visual fields or in sequence ineach visual field. The patient’s task was to reportcolours at each location or shapes at each location.Results showed that extinction was increased whenthe two stimuli were identical on the relevantdimension for the task (for example, red letters in thereport colour task) while it was unaffected by therepetition on the irrelevant dimension (for example,two letter A in the report colour task). The authorspropose an intriguing explanation of their resultsappealing to Kanwisher (1987) and Volpe et al.’s(1979) explanation of RB effect, suggesting thatwhile type recognition is preserved in thecontralesional field, token individuation iscompromised (the episodic representation of a

distinct perceptual object or event). When a singlestimulus is presented contralesionally, it can beprocessed moderately well, but when a simultaneousstimulus of the same type is presented ipsilesionally,token individuation process fails. Following theauthors, specific neural factors support thisinterpretation. Indeed, the information in thecontralesional field is processed by both dorsal andventral pathways. Consequently both identity andlocation are processed. However, the presence of aneurological deficit means that the two pieces ofinformation may not be bound together generating afailure in stimuli detection.
While RSVP paradigm has been frequently usedto study interference in identification of the stimuliin the visual modality, to our knowledge no previousstudies have been carried out to investigate howattention to tactile information in a restricted area ofthe body is distributed over time; moreover, to ourknowledge no previous studies have investigated therelationship between temporal and spatio-temporalvariables in extinction phenomenon. As well as in thevisual modality, we suppose that in patients with rightbrain damage and tactile extinction there is analteration of the temporal elaboration of the stimulibut we ignore the relationship between temporal andspatio-temporal variables. The present study reportsa tactile detection experiment in which two differentrandomised tasks were employed. In the Temporal(T) task, aimed at assessing temporal aspects oftactile attention, single or sequential double electricalstimuli were delivered to the same point of the indexfinger of only one hand. Left and right hands weretested in separate blocks. Double stimuli weresequential and separated by 7 different SOAs. In theSpatio-Temporal (ST) task, aimed at investigatingthe interplay between spatial and temporal variablesinfluencing tactile attention, single or double stimuliwere delivered to the left, the right, or to homologouspoints on left and right index fingers. Double stimuliwere simultaneous or separated by the same timeintervals used in the T task. In both tasks, stimuliwere presented by a computer to control the stimulusduration, the stimulus intensity and the SOA(Guerrini et al., 2003). Since the quality of electricaltactile stimuli cannot be differentiated in any way weused tactile detection rather than a discriminationtask.
METHOD
Participants
Six right-handed Right-Brain Damaged (RBD)patients (1 woman and 5 men) with clinical andradiological evidence for an ischemic orhemorrhagic lesion of the right hemisphereparticipated in the study. Reconstruction of thepatients’ lesion sites and extent is shown in Figure1. Lesions were reconstructed using MRIcro
Space and Time and Tactile Extinction 19
software package (www.MRIcro.com; Rorden andBrett, 2000).
Six (5 right-handed and one ambidextrous)neurologically intact subjects (2 men and 4 women)served as control group. All patients and normalsubjects gave their informed consent to participatein the study. The protocol was approved by theinstitutional review board. Patients’ mean age andmean education was respectively 61.3 years (SD9.4, range 48-75) and 8.3 years of schooling (SD3.7, range 5-13 years). Clinical information andscreening results are shown in Table I.
Healthy controls’ mean age and education wasrespectively 59.3 years (SD 10.7, range 42-73) and11.8 years (SD 4.6 years, range 5-18 years). Twoseparate two-sample T-Tests showed the absence ofstatistically significant difference between the twogroups in age or education [Age: t(10) = .3, p = .74;Education: t(10) = – 1.4, p = .18]. All the patientswere submitted to an assessment session to evaluatethe presence of extinction and any other disordersusually associated with right brain damage. Severityof motor impairment, anosognosia, visual, tactile andauditory extinction, personal and extrapersonalneglect were assessed and scored according to astandard neurologic examination (see Bisiach et al.,1986; Aglioti et al., 1998, 1999; Guerrini et al., 2003,
Fig. 1 – Lesion reconstruction for RBD patients, usingMRIcro software (www.MRIcro.com). The figure shows the siteand the size of the lesion (coloured in black) for each extinctionpatient.
for more details). Tactile extinction was alsoassessed employing a computer controlled extinctiontest. The presence of extinction in a StandardisedClinical Test and in a Computerised Extinction Test(Aglioti et al., 1998, 1999; Guerrini et al., 2003) wasthe main criterion of inclusion in our study. Patientswere considered to have extinction when theyomitted at least 30% of the stimuli upon doublestimulation and detected at least 70% of singlestimuli (Table II).
In the Computerised Extinction Test (Figure2B) patients received a fixed random sequence of20 single left stimuli, 20 single right, 20 bilateralsimultaneous and 4 catch trials. All subjectsperformed at least 2 blocks of trials. Stimuliconsisted of noiseless and non-noxious (1 msec)electric stimuli delivered by means of punctiformelectrodes, 1 mm in diameter, positioned on the leftand right index fingers. The electrodes wereactivated by Single Phase Current Stimulators(STM 140, High Technology Laboratory, Udine,Italy) under the control of a PC computerprogrammed with the software package MicroExperimental Laboratory (MEL), Version 2(Schneider, 1988). To ensure an approximate inter-manual perceptual equivalence, stimuli intensitywas defined for each subject before starting theexperimental session, separately for the left and theright hand, in relation with their sensitivity. Thecriteria employed to define intermanual subjectiveequivalence were the following: first, stimuli had
20 Chiara Guerrini and Salvatore M. Aglioti
to be perceived clearly; second, stimuli were not tobe perceived as painful; third, stimuli intensity hadto be referred as similar for both left and righthands. The intensity of single stimuli delivered tothe contralesional hand was set at a value thatallowed the detection of 5 stimuli out of 10(threshold level). Then, the intensity was increasedso as to obtain that at least 8 out of 10 singlestimuli were detected. Stimuli with this intensitydelivered to the ipsilesional hand were typicallyperceived as too much intense. Thus, the intensityof the stimuli on the ipsilesional side was reducedto ensure an approximate intermanual perceptualequivalence. Inter-manual perceptual equivalencewas constantly checked through the experiment.
Apparatus and Stimuli
The same apparatus, the same stimuli describedfor the Computerised Extinction Test, and the sameprocedure employed in the assessment session toensure an approximate inter-manual perceptualsubjective equivalence, were used in theexperimental session.
Procedure
The order of T and ST tasks was counterbalancedin the patient group. The control group performedonly the T task. In the T task (Figure 2A) subjectswere seated with one hand resting on a table, aligned
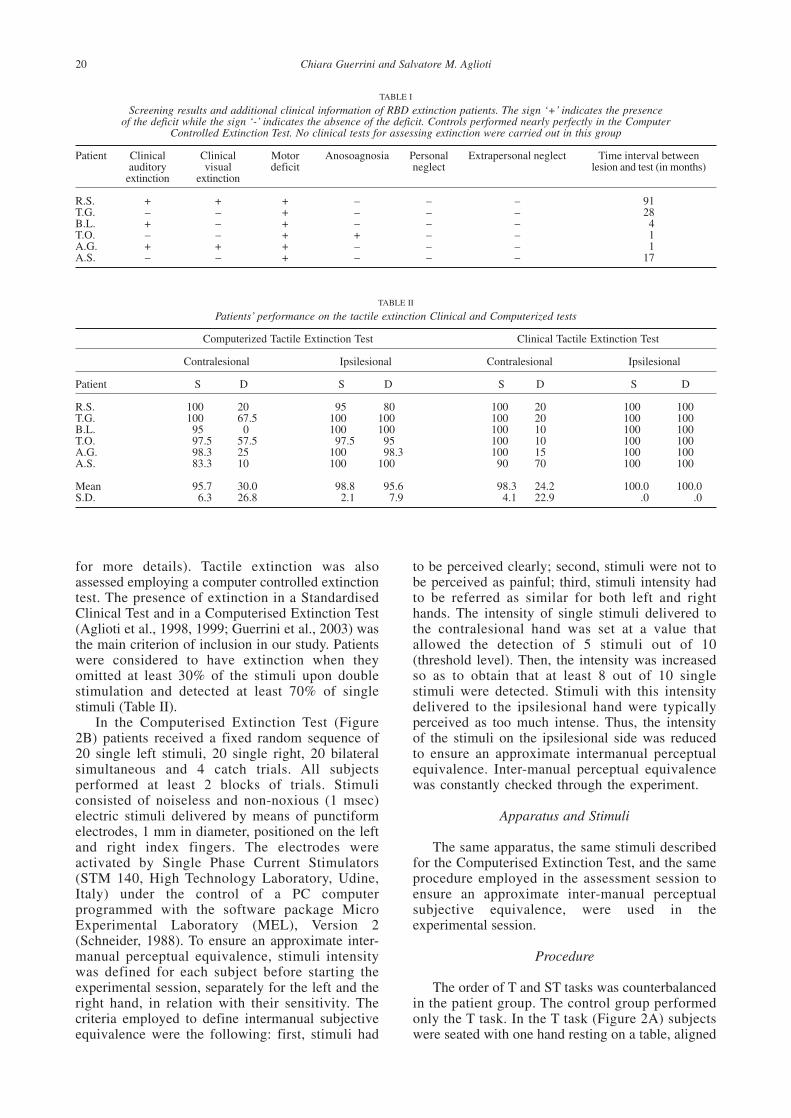
TABLE I
Screening results and additional clinical information of RBD extinction patients. The sign ‘+’ indicates the presenceof the deficit while the sign ‘-’ indicates the absence of the deficit. Controls performed nearly perfectly in the Computer
Controlled Extinction Test. No clinical tests for assessing extinction were carried out in this group
Patient Clinical Clinical Motor Anosoagnosia Personal Extrapersonal neglect Time interval betweenauditory visual deficit neglect lesion and test (in months)
extinction extinction
R.S. + + + – – – 91T.G. – – + – – – 28B.L. + – + – – – 4T.O. – – + + – – 1A.G. + + + – – – 1A.S. – – + – – – 17
TABLE II
Patients’ performance on the tactile extinction Clinical and Computerized tests
Computerized Tactile Extinction Test Clinical Tactile Extinction Test
Contralesional Ipsilesional Contralesional Ipsilesional
Patient S D S D S D S D
R.S. 100 20 95 80 100 20 100 100T.G. 100 67.5 100 100 100 20 100 100B.L. 95 0 100 100 100 10 100 100T.O. 97.5 57.5 97.5 95 100 10 100 100A.G. 98.3 25 100 98.3 100 15 100 100A.S. 83.3 10 100 100 90 70 100 100
Mean 95.7 30.0 98.8 95.6 98.3 24.2 100.0 100.0S.D. 6.3 26.8 2.1 7.9 4.1 22.9 .0 .0

with their sagittal midline. The other hand waspositioned on the ipsilateral thigh. Left and righthands were tested in different randomised blocks oftrials.
In order to avoid influences of shifts of visualattention on tactile attention, subjects wererequested to maintain fixation on a central point(Vaishnavi et al., 2001), positioned on the table (ata distance of about 60 cm) and aligned with theirsagittal midline. The experimenter pressed a keyon the computer keyboard to initiate the trial.After a random interval (from 1100 to 2300msec), single or double sequential stimuli weredelivered to the same point of the index finger.Sequential stimuli were separated by one out of 7
Space and Time and Tactile Extinction 21
possible SOAs (50, 100, 200, 300, 500, 700, 900msec). On each trial subjects were asked to reportthe number of the stimuli (one, two, or none forthe catch trials). Subjects’ response modality wasnot constrained, so they could report theirexperience verbally or through gestures.Responses were entered into a computer by theexperimenter. Subjects were previously informedthat stimuli could be single or double, and wereasked to respond only after hearing a beep signal,which was delivered within a random interval of300-800 msec after the second stimulus. The nexttrial was initiated only after the subject declaredhe was ready to restart. All the subjects performedat least 2 blocks of trials (1 block for each hand).
Fig. 2 – Experimental setting: Figure A reports the experimental setting for the T task; Figure B reports the experimental setting forboth the Computerized Extinction Test and the ST task.

Each block consisted of 10 single stimuli, 10double sequential stimuli for each possible SOA,and 10 catch trials. The sequence of single, doubleand catch trials was randomised. SOAs and blocksof trials were also randomised. Comprehension ofthe instructions for each condition was checked bygiving to each subject at least 20 practice trials.Pauses were commensurate to subject’s fatigue.No feedback about response correctness was givenduring testing. In the ST task, all the aspects ofthe method were the same as those for the Tcondition except as noted. In this second conditionsubjects were seated in front of a table with bothhands on it (Figure 2B). Double stimuli weredelivered bilaterally. Bilateral stimuli could beseparated by one out of the 7 possible SOAs usedin the T condition but also simultaneous (SOA 0msec). In the non-simultaneous stimulationcondition, the stimulus to the left hand couldbe preceded (50% of the trials) or followed (50%of the trials) by the stimulus on the right hand.Each subject performed at least four blocks oftrials.
Each block was constituted by 2 single leftstimuli, 2 single right stimuli, 8 simultaneousbilateral stimuli, 2 bilateral stimuli for eachpossible positive SOAs (conventionally defined bythe condition in which the right stimulus precededthe left stimulus) and 2 bilateral stimuli for eachpossible negative SOA (conventionally, thenegative SOA define the condition in which theleft stimulus precedes the right one) yielding atotal of 28 non-simultaneous trials, plus four catchtrials without stimuli. The sequence of single andbilateral stimuli and catch trials was randomised.SOAs and sequence of presentation (left-first orright-first) were also randomised. Subjects wereinformed prior to testing that stimuli could besingle or bilateral, and that bilateral stimuli couldbe synchronous or asynchronous. As in the T task,subjects were requested to report, verbally orthrough gestures, the number of the stimuli (one,two or none).
Because our aim was also to compare thedetection of left sided stimuli in the T versus STtasks, we needed information about the location ofthe stimulus perceived. Thus, in the ST tasksubjects were asked to define also the location ofthe stimuli perceived (left, right or both hands).
Since a previous study (Guerrini et al., 2003)showed that extinction patients performance in theST task was not affected by the differentproportion of double simultaneous stimuli versusdouble asynchronous stimuli, a balanced controlledcondition was not included for this study. However,because the proportion of single versus doublestimuli was not balanced in the T task, oneextinction patient (RS) also performed a modifiedversion of the T task in which the number of singlestimuli was identical to the sum of all the doubleasynchronous stimuli.
22 Chiara Guerrini and Salvatore M. Aglioti
RESULTS
Correct Responses
Accuracy of the healthy control subjects in the Ttask (the only paradigm performed by this group) indetecting single right and left stimuli wasrespectively 91.2% and 93%. Their accuracy indetecting double right and left stimuli wasrespectively 94.5% and 97.5%. The accuracy of the6 extinction patients in detecting single right and leftstimuli in the T task was respectively 82.1% and76.6%. This difference (5.5%), likely due to a deficitof sensitivity of the left hand, was not significantlydifferent [t(5) = 1.1, p = .3]. Mean accuracy fordouble stimuli was 73.5% for the right hand and only31.8% for the left hand (see Figure 3).
The percent of correct detections under doublestimulation condition were entered in to twoseparated repeated-measures analysis of variance(ANOVA), one for patients and the other for thecontrols, with the following within-subject factors:hand (left vs. right) and SOA (50, 100, 200, 300,500, 700, 900 msec). In the healthy control groupthe only main significant factor was SOA [F (6, 30)= 2.9, p < .02]. The significance of this factor is dueto the lack of accuracy (83%) at the shortest SOA,where the subjects commit many errors judging thetwo sequential stimuli as a single one. Meanaccuracy expressed in percentage for each SOA (50,100, 200, 300, 500, 700, 900 msec) wasrespectively: 82.9, 90.9, 98.9, 99.1, 99.3, 99.1,98.75. In the extinction patients’ group significanteffects were obtained for hand [F (1, 5) = 28.8, p <.003], detection accuracy being higher for righthand (73.5%) than for left hand (31.8%) and SOA[F (6, 30) = 8.8, p < .0001]. The interaction hand XSOA was not significant. This means that accuracyincreases with increasing SOAs for both left andright hands. To ensure that the results in the T-Taskwere not due to a bias generated by the differentproportions of single versus double asynchronousstimuli, one extinction patient (RS) performed also amodified version of the T task in which the numberof single stimuli was identical to the number of thedouble stimuli (balanced condition). RS’sperformance in the balanced and unbalancedcondition is shown in Table III. A series of χ2 Testswere performed to compare RS’s performance in thebalanced versus unbalanced condition. Importantly,no comparison showed a significant difference [χ2 (1) < 3.49 for all comparisons].
The main aim of the present study was toinvestigate the relationship between temporal andspatio-temporal variables on extinction of thecontralesional stimulus under double stimulationcondition. For this reason, we decided to comparepatients’ left hand correct detection performance inthe T task with left hand correct detectionperformance in the ST task under doublestimulation. Thus, the subsequent analyses follow

this logic. In the T task, patients’ percentage ofaccuracy for the single left stimulation conditionwas 76.6%; while the percentage of accuracy for thedouble left stimulation condition was 31.8%. In theST task, patients’ percentage of accuracy for thesingle left stimulation condition was 98.3%, whilethe percentage of accuracy for the left doublestimulation condition was 63.4% for the left hand(see Figure 4). A repeated-measures ANOVA wasperformed to compare patient’s correct detection ofleft stimuli in both T and ST tasks (see Figure 4).The within-subject factors were task (T vs. ST) andSOA (Single, 50, 100, 200, 300, 500, 700, 900msec). Among the main factors, significant effectswere obtained for task [F (1, 5) = 18.6, p < .008],detection accuracy being higher for the ST (67.8%)than for the T task (37.4%), and SOA [F (7, 35) =
Space and Time and Tactile Extinction 23
17.4, p < .0001], reflecting a general increasingaccuracy as a function of increasing SOA. Theinsignificance of the interaction task X SOA [F (7,35) < 1] suggests that accuracy increases withincreasing SOAs in both T and ST tasks.
Errors
From a qualitative point of view the followingtypes of errors could have occurred: responses tocatch trials (called false alarms), reports that nostimulus was presented (these errors were calledmisses), reports of two stimuli under singlestimulation conditions (called duplications), reportsof one stimulus under double stimulationconditions (called temporal extinction errors in theT task, and spatio-temporal extinction errors in the
Fig. 3 – RBD extinction patients’ mean accuracy (plus standard error) in reporting left and right sided stimuli under single anddouble stimulation conditions in the T task. The horizontal lines represent accuracy under single left (dotted) and right (continuous)stimulation conditions.
TABLE III
Accuracy of RS in detecting left and right stimuli under single and double stimulation condition in the two versions of the T task.Each cell contains the number of stimuli detected over the number of stimuli administered
SOA
Single 50 100 200 300 500 700 900
Left hand Balanced 202/280 30/40 37/40 36/40 38/40 34/40 26/40 24/40Unbalanced 19/30 24/30 29/30 26/30 28/30 23/30 23/30 20/30
Right hand Balanced 217/280 1/40 3/40 3/40 5/40 5/40 5/40 1/40Unbalanced 25/40 0/40 2/40 4/40 7/40 8/40 0/40 2/40
SOA(msec)

ST task). Percentage of errors is computed withrespect to the total number of stimuli administeredin the specific condition.
T task
False alarms for the right hand amounted to3.5% of the catch trials and to 12% of the catchtrials for the left hand. Although this may indicatea tendency to attribute phantom perceptions to theleft hand, the difference was not significant [T-Test:t(5) = – 1.7, p = .15]. Under single stimulationconditions left hand and right hand duplicationswere respectively 8.7% and 8%. This differencewas not statistically significant [t(5) = – .4, p = .7].Left hand and right hand misses were respectively15.9% and 8.3% (Table IV).
24 Chiara Guerrini and Salvatore M. Aglioti
This difference was statistically significant[t(5) = 3, p = .03]. In the patients group thepercentage of temporal extinction errors and ofmisses under double stimulation conditions servedfor two different ANOVAs with hand (left or right)and SOA (50, 100, 200, 300, 500, 700, 900 msec)as within subjects factors. In the temporalextinction errors analysis, significant differenceswere found for hand [F (1, 5) = 30.03, p < .003]since patients made more temporal extinction errorson the left side stimulation condition (62.1%) thanin the right side stimulation condition (24.7%), andfor SOA [F (6, 30) = 12.02, p < .0001] since thistype of error decreased with increasing SOAs. Inthe misses analysis, significant differences werefound for hand [F (1, 5) = 6.9, p < .05] sincepatients made more misses in left side stimulation
Fig. 4 – Comparison between patients’ left stimulation condition correct detections, in the ST task (continuous) and in the T task(dotted). The horizontal lines represent accuracy under single stimulation condition in T (dotted) and in ST (continuous) tasks.
TABLE IV
Temporal extinction errors and omissions errors recorded in the patients’ group in the T task. Percentages are computed with respectto the total number of stimuli administered in the specific condition
Temporal SOA
Extinction errors 50 100 200 300 500 700 900 Single
Left hand 89.2 75.8 68.3 65.7 45.1 39.4 51.2 8.7Right hand
60.8 35.3 19.2 10.8 16.7 12.2 17.8 8MissesLeft hand .8 2.5 6.9 3.8 7.4 14.4 6.8 15.9Right hand 2.2 2.2 2.5 .5 2.2 2.7 .5 8.3
SOA(msec)

condition (6.1%) than in the right side stimulationcondition (1.8%), for SOA [F (6, 30) = 3.8, p <.006] and for hand X SOA interaction [F (6, 30) =5.1, p < .001].
ST task
False alarms and misses occurred very rarely(less than 3% and 1%) and therefore were notfurther analyzed. Under single stimulationconditions, duplication errors were comparable forleft and right hand (11.9% vs. 16.7% respectively;T-Test, p = .4). Under double stimulation conditions,spatio-temporal extinction errors were the mostcommon type of errors amounting to 32.8% of thetrials while the percentage of omissions amountedto .5%.
DISCUSSION
Recent neuropsychological studies carried outin the visual modality suggest a deficit in temporalprocessing in extinction (Husain et al., 1997; diPellegrino et al., 1997, 1998; Cate and Behrmann,2002), a disorder classically interpreted as a purelyspatial phenomenon (Posner et al., 1984; Posnerand Petersen, 1990). The current investigation wasmotivated by the scarcity of studies on the role oftemporal variables in tactile extinction. The aim ofthe present work was to analyse the role ofattention distributed over time but focused in arestricted area of the body and to elucidate therelationship between temporal variables and spatio-temporal variables on tactile extinctionphenomenon. In the present study non-noxious andnoiseless electrical computer controlled stimuliwere used. This design has two major benefits overclassical methods for measuring extinction (e.g.,where the stimulus is a tap delivered by theexperimenter). First, experimenter’s subjectivevariability was eliminated as intensity and durationof the stimuli were computer controlled. Second,patients’ strategic behaviours were reduced as novisual or auditory information was given during thecomputerized stimulation.
Our results show that patients’ left handperformance in the T task is significantly anddramatically lower than right hand stimuli detectionat all SOAs, except at 50 msec SOA. In thiscondition, although left hand performance is worsethan right hand performance, patients find itdifficult to discriminate two stimuli in sequenceindependently of the side of stimulation. As SOAincreases, patients’ left hand performance alsoincreases but never reaches right hand performance(baseline). These results are similar with previousdata found in the visual modality (Husain et al.,1997; di Pellegrino et al., 1998). Husain et al.(1997) showed that the ability of neglect patients toidentify a visual object presented in the centre of
Space and Time and Tactile Extinction 25
the screen increases at increasing SOAs and isimpaired if a second stimulus appears within 1440msec of a first stimulus. In this paradigm, althoughlonger SOAs were used than in the present study(from 180 msec to 1800 msec), patients’ accuracynever reached the baseline performance (singletarget identification). di Pellegrino et al. (1998)employed a similar task to study PB’s performance,a left extinction patient, who was asked to identifyletters displayed in rapid succession to the left orto the right hemifield. Results show thatinterference duration of the first stimulus on thesecond is twice longer in the contralesional than inthe ipsilesional field. But contrary to our resultsand Husain et al.’s results (1997), the ability of PBto identify a visual object reaches the baselineaccuracy (identification of a single stimulus) if itappears within 800 msec of a first stimulus. Ourresults appear to be attentionally based and due tothe interaction that the two stimuli presented insequence produces on the attentional demands.Memory limitations in fact, are unlikely to play arole due to the fact that the task requires a simpletactile detection of the stimulus. Also, it is unlikelythat the phenomenon is due to sensory limitations.In fact, in single stimulation condition the loss ofthe left sided stimulus is strongly reduced and thecomparison between the T task and the ST taskseems to confirm this result. The percentage ofaccuracy of the left stimulus detection is higherwhen two stimuli are delivered in sequence insymmetric parts of the body located in symmetricspace areas than when two stimuli are delivered insequence in the same body area aligned with thesagittal midline. Surprisingly, the spatial variableconsidered as the main cause of extinction, seemsto play an important role in the detection of thetactile stimuli delivered contralesionally. Twoexplanations can be offered for interpreting ourresults. Studies of the visual attentional blinkphenomenon in neurologically intact subjectssuggest that the processing of two sequentialstimuli is influenced by the interval between thetwo. Indeed, if the second stimulus appears whenanalysis of features characterizing the first stimulusis still on, interference occurs. It follows that, thetime-course of the interference reflects how longthe processing of a first stimulus capturesattentional resources. Together with di Pellegrino etal.’s results (1997, 1998) and Husain et al.’s results(1997), the present data suggest that in patientswith neglect and extinction the identification of acontralesional stimulus makes more long lastingdemands on attention than an ipsilesional stimulus.On the other hand, according to Baylis et al.(2001), the information is processed by bothventral and dorsal pathways such that both identityand location are processed. In patients with damageto attention-related brain structures, the two typesof information can not be bound together and failto reach awareness. This statement might be valid

also in the tactile modality since new evidenceshow that the neuro-anatomical and functionalsegregation characterize not only the visual systembut also the other sensorial systems (Alain et al.,2001; Rauschecker et al., 1995; Romanski et al.,1999). As Baylis et al. (1993) suggested when asingle stimulus is presented contralesionally, it canbe processed moderately well but when two stimuliof the same type are presented in rapid sequence,in different hemifields, the creation of a distinctperceptual event (token) may fail. Our data confirmthis interpretation but seem to add new informationto previous reports.
The present results suggest that the spatialvariable in some way contributes to create, atappropriate SOAs, different tokens. Consequently,the contralesional stimulus may be detected moreaccurately under bilateral asynchronous stimulationthan under double unilateral asynchronousstimulation. When two stimuli of the same type arepresented in rapid sequence and in the same place,they cannot be distinguished in any way andconsequently the token individuation process fails.When the spatial information is absent, longer timeis needed so that the two stimuli are perceived asseparated. The temporal processing deficit presentin our extinction patients group seems to increasethe interval necessary to perceive the two stimuli asseparated. When the token individuation processfails subjects appear confused and consequentlytend to commit many errors. On the basis ofpatients’ reports, it is possible to suppose that whenthe token individuation fails, patients might havedifferent subjective experience. In the singlestimulation condition and in the catch trials patientsseem not to be sure they perceived a stimulus ornot. Consequently, they make missing errors orfalse alarms. Under double stimulation condition,patients experience some unclear perception butthey can not define the characteristics of theperception. Consequently, they make manyextinction errors and sometimes, if they are indoubt, they state that they perceive nothing. Ourdata suggest that spatial and temporal variables maywork independently but create similar phenomena.Both spatial and temporal dimensions areresponsible for the failure in detection of left sidedstimuli under double stimulation. These results arecorroborated by a research by Shapiro et al. (2002)and in Rizzo et al. (2001) who suggested thatabnormal temporal attention dynamics are notneglect specific or extinction specific but ratherthey may follow different brain lesions, in bothhemispheres. Rizzo et al. (2001) studied 13 subjects(10 without neglect and 3 with neglect) withchronic focal brain lesions and 9 neurologicallyintact controls in a RVSP task. They found that anabnormal Attentional Blink may occur withdifferent focal brain lesions, in either hemisphereand in the absence of neglect. In a similar vein,Shapiro et al. (2002) studied a group of patients
26 Chiara Guerrini and Salvatore M. Aglioti
with focal lesions with or without spatial neglectemploying a visual RSVP task. Their results showthat regardless of the presence of neglect, damageto the inferior parietal lobe and to superior temporalgyrus leads to a more prolonged deployment oftemporal attention. In a recent work, Husain andRorden (2003) proposed that both spatiallylateralised and non-spatially lateralised mechanismsare involved in neglect. Interestingly, some researchsuggests that spatially lateralised and non-lateralised functions are anatomically segregated,supporting this hypothesis. Functional imagingstudies show that lateralised attentional mechanismsappear linked to the Superior Parietal Lobe (SPL)(Vanderberghe et al., 2001; Corbetta et al., 1993),and non-lateralised mechanisms seem to be relatedto the Inferior Parietal Lobe (IPL) (Husain et al.,1997). Moreover, regions between the IPL and SPL,probably sub-serve both lateralised and non-lateralised functions (Coull and Frith, 1998;Wojciulik and Kanwisher, 1999). According toHusain and Rorden (2003) the combination ofspatially lateralised and non-lateralised deficitsleads to a persistent neglect and multiplecomponents interact to exacerbate neglect. Indeed,the interaction of the two variables at a specifictime and spatial window seems to play animportant role, enhancing the ability of the tactileextinction patients to detect left sided stimuli.
Acknowledgements. This research was supported bygrants from the HFSP (RG 0161/1999-301) and MIUR,Italy to Salvatore M. Aglioti. We wish to thank Mr. EneaF. Pavone (Dipartimento di Scienze Neurologiche e dellaVisione, Sezione Fisiologia Umana, Università degli Studidi Verona, Italia) for his expert advice on reconstructingthe lesions with MRIcro; Miss Nicoletta Finco, Miss DariaGiacon (Ospedale Chiarenzi, Zevio, Verona, Italia) and Dr.Feliciana Cortese (Ospedale S. Bortolo, Vicenza, Italia)for their help in patients recruitment.
REFERENCES
AGLIOTI SM, SMANIA N, MORO V and PERU A. Tactile salienceinfluences extinction. Neurology, 50: 1010-1014, 1998.
AGLIOTI SM, SMANIA N and PERU A. Frames of reference formapping tactile stimuli in brain-damaged patients. Journal ofCognitive Neuroscience, 11: 67-79, 1999.
ALAIN C, ARNOTT S, HEVENOR S, GRAHAM S and GRADY C.“What” and “where” in the human auditory system. PNAS,98: 12301-12306, 2001.
ANTON G. Ueber die Selbstwahrnehmung der Herderkrankungendes Gehirns durch den Kranben bei Rindenblindheit undRindentaubheit. Archiv Psychiatrie und Nervenkrankheiten,32: 86-111, 1899.
BARBIERI C and DE RENZI E. Patterns of neglect dissociation.Behavioural Neurology, 2: 13-24, 1989.
BAYLIS GC, DRIVER J and RAFAL R. Visual extinction and stimulusrepetition. Journal of Cognitive Neuroscience, 5: 453-466,1993.
BAYLIS GC, GORE C, RODRIGUEZ P and SHISLER R. Visualextinction: The importance of binding dorsal and ventralvisual pathways. Visual Cognition, 8: 359-379, 2001.
BAYLIS GC, SIMON SL, BAYLIS LL and RORDEN C. Visualextinction with double simultaneous stimulation: What issimultaneous? Neuropsychologia, 40: 1027-1034, 2002.
BISIACH E, VALLAR G, PERANI D, PAPAGNO C and BERTI A.Unawareness of disease following lesions of the righthemisphere: Anosognosia for hemiplegia and anosognosia forhemianopia. Neuropsychologia, 24: 471-482, 1986.

Space and Time and Tactile Extinction 27
BROADBENT DE and BROADBENT MH. From detection toidentification: Response to multiple targets in rapid serialvisual presentation. Perception and Psychophysics, 42: 105-113, 1987.
CATE A and BEHRMANN M. Spatial and temporal influences onextinction in parietal patients. Neuropsychologia, 40: 2206-2225, 2002.
CHUN MM and POTTER MC. A two-stage model for multiple targetdetection in rapid serial visual presentation. Journal ofExperimental Psychology: Human Perception andPerformance, 21: 109-127, 1995.
CORBETTA M, MIEZIN FM, SHULMAN GL and PETERSEN SE. A PETstudy of visuospatial attention. Journal of Neuroscience, 13:1202-1226, 1993.
COULL JT and FRITH CD. Differential activation of right superiorparietal cortex and intraparietal sulcus by spatial andnonspatial attention. NeuroImage, 8: 176-187, 1998.
COULL JT, FRITH CD, BUCHEL C and NOBRE AC. Orientingattention in time: Behavioural and neuroanatomical distinctionbetween exogenous and endogenous shifts. Neuropsychologia,38: 808-819, 2000.
COULL JT and NOBRE AC. Where and when to pay attention: Theneural systems for directing attention to spatial locations andto time intervals as revealed by both PET and fMRI. Journalof Neuroscience, 18: 7426-7435, 1998.
DI PELLEGRINO G, BASSO G and FRASSINETTI F. Spatial extinctionon double asynchronous stimulation. Neuropsychologia, 35:1215-1223, 1997.
DI PELLEGRINO G, BASSO G and FRASSINETTI F. Visual extinction asa spatio-temporal disorder of selective attention. NeuroReport,9: 835-839, 1998.
DUNCAN J, WARD R and SHAPIRO K. Direct measurement ofattentional dwell time in human vision. Nature, 369: 313-315,1994.
GRIFFIN IC, MINIUSSI C and NOBRE AC. Multiple mechanisms ofselective attention: Differential modulation of stimulusprocessing by attention to space or time. Neuropsychologia,40: 2325-2340, 2002.
GUERRINI C, BERLUCCHI G, BRICOLO E and AGLIOTI SM. Temporalmodulation of spatial tactile extinction in right brain-damagedpatients. Journal of Cognitive Neuroscience, 15: 523-536,2003.
HUSAIN M and RORDEN C. Non-spatially lateralised mechanisms inhemispatial neglect. Nature Reviews Neuroscience, 4: 26-36,2003.
HUSAIN M, SHAPIRO K, MARTIN J and KENNARD C. Abnormaltemporal dynamics of visual attention in spatial neglectpatients. Nature, 385: 154-156, 1997.
KANWISHER NG. Repetition blindness: Type recognition withouttoken individuation. Cognition, 27: 117-143, 1987.
KARNATH HO, ZIMMER V and LEWALD J. Impaired perception oftemporal order in auditory extinction. Neuropsychologia, 40:1977-1982, 2002 .
KEPPEL G. Design and Analysis: A Researcher’s Handbook (2nded.). Englewood Cliffs, NJ: Prentice-Hall, Inc., 1982.
MINIUSSI C, WILDING EL, COULL JT and NOBRE AC. Orientingattention in time: Modulation of brain potentials. Brain, 122:1507-1518, 1999.
NOBRE AC. Orienting attention to instants in time.Neuropsychologia, 39: 1317-1328, 2001.
POSNER MI and PETERSEN SE. The attention system of the humanbrain. Annual Review of Neuroscience, 13: 25-42, 1990.
POSNER MI, WALKER JA, FRIEDERICH FA and RAFAL RD. Effects ofparietal injury on covert orienting of attention. Journal ofNeuroscience, 4: 1863-1874, 1984.
RAUSCHECKER J, TIAN N and HAUSER M. Processing of complexsounds in the macaque nonprimary auditory cortex. Science,268: 111-114, 1995.
RAYMOND JE, SHAPIRO KL and ARNELL KM. Temporarysuppression of visual processing in an RSVP task: Anattentional blink? Journal of Experimental Psychology:Human-Perception and Performance, 18: 849-860, 1992.
RIZZO M, AKUTSU H and DAWSON J. Increased attentional blink after focal cerebral lesions. Neurology, 57: 795-800,2001.
ROMANSKI LM, TIAN B, FRITZ J, MISHKIN M, GOLDMAN-RAKIC PSand RAUSCHEKER J. Dual streams of auditory afferent targetmultiple domains in the primate prefrontal cortex. NatureNeuroscience, 2: 1131-1136, 1999.
RORDEN C and BRETT M. Stereotaxic display of brain lesions.Behavioural Neurology, 12: 191-200, 2000.
RORDEN C, MATTINGLEY JB, KARNATH HO and DRIVER J. Visualextinction and prior entry: Impaired perception of temporalorder with intact motion perception after unilateral parietaldamage. Neuropsychologia, 35: 421-433, 1997.
SCHNEIDER W. Micro experimental laboratory: An integratedsystem for IBM-PC compatibles. Behavior Research Methods,Instrumentation and Computers, 20: 206-217, 1988.
SHAPIRO KL, HILLSTROM AP and HUSAIN M. Control ofvisuotemporal attention by inferior parietal and superiortemporal cortex. Current Biology, 12: 1320-1325, 2002.
SHAPIRO KL, RAYMOND JE and ARNELL KM. Attention to visual pattern information produces the attentional blink inrapid serial visual presentation. Journal of ExperimentalPsychology: Human Perception and Performance, 20: 357-371, 1994.
VAISHNAVI S, CALHOUN J and CHATTERJEE A. Binding personal andperipersonal space: Evidence from tactile extinction. Journalof Cognitive Neuroscience, 13: 181-189, 2001.
VALLAR G, RUSCONI ML, BIGNAMINI L, GEMINIANI G and PERANID. Anatomical correlates of visual and tactile extinction inhumans: A clinical CT scan study. Journal of Neurology,Neurosurgery and Psychiatry, 57: 464-470, 1994.
VANDERBERGHE R, GITELMAN DR, PARRISH TB and MESULAM MM.Functional specificity of superior parietal mediation of spatialshifting. NeuroImage, 14: 661-673, 2001.
VOLPE BT, LEDOUX JE and GAZZANIGA MS. Informationprocessing of visual stimuli in an “extinguished” field. Nature,282: 722-724, 1979.
WOJCIULIK E and KANWISHER N. The generality of parietalinvolvement in visual attention. Neuron, 23: 747-764, 1999.
Chiara Guerrini, Clinical Neuroscience Centre and Department of Psychology,University of Hull, Cottingham Road, Hull HU6 7RX, England, UK.e-mail: [email protected]
(Received 19 December 2003; reviewed 9 March 2004; revised 3 May 2004; accepted 18 May 2004; Action Editor CarloUmiltà)