temporary tracheal occlusion causes catch-up lung maturation in a fetal model of diaphragmatic...
TRANSCRIPT

Temporary Tracheal Occlusion Causes Catch-Up Lung Maturation in a Fetal Model of Diaphragmatic Hernia
By Konstantinos Papadakis, Monique E. De Paepe, Leslie D. Tackett, George J. Piasecki,
and FranCois I. Luks
Providence, Rhode Island
Background: The lungs of infants born with diaphragmatic hernia are hypoplastic, immature, and surfactant-deficient. Tracheal occlusion in utero, which is being proposed as antenatal treatment of diaphragmatic hernia by promoting compensatory lung growth, decreases surfactant production as well, through loss of type II pneumocytes. The authors studied whether temporary tracheal occlusion might cause ‘catch-up’ lung growth and maturation, without negative effects of prolonged tracheal occlusion on the surfactant system.
Methods: Diaphragmatic hernia was created in time-dated fetal lambs (65 to 75 days). At 108 days, the trachea was occluded with an embolectomy catheter (DH + TO, n = 6). After day 14, the balloon was deflated. Six congenital diaphag- matic hernia (CDH) fetuses were left unobstructed (DH). For comparison, a group of fetuses without diaphragmatic hernia were subjected to prolonged tracheal ligation (TL; 4-week tracheal ligation, n = 3). Unoperated littermates (t-r = 8) were used as controls (CTR). All were killed near term. Lung tissue was processed for light and electron microscopy (computer- ized stereologic morphometry). Type II pneumocytes were identified with antisurfactant protein B antibody.
Results: Four animals in DH + TO and four in DH survived to term. Lung fluid volume (LFV) at 108 days was 5.2 i 4.4 mL in DH and 24.6 i 6.8 mL in controls (P < .05, Student t test). In DH + TO, LFV increased ninefold (to 48.3 IT 13.3 mL) by 1 week postocclusion, suggesting accelerated lung growth. At
term, lung weightto body weight ratio (LW/BW) was higher in TL (9.85% ? 1.81%) than in CTR (3.55% i 0.56%; P< .05, analysis of variance); LW/BW and parenchymal volume tended to be greater in DH + TO than in DH, and air-exchanging parenchymal volume in DH + TO was similarto CTR (va 50% reduction in DH), indicating some degree of hyperplasia after temporary occlusion. Pneumocyte II numerical density was decreased more than IO-fold in TL (60 + 22 v 826 ? 324 in CTR, P < .OOl; it was slightly lower in DH + TO than in CTR, but individual type II pneumocyte cell volume was greater in the latter, and they appeared more mature than in DH (increased granulation by light microscopy, fewer glycogen granules, and abundant lamellar bodies by electron micros- copy). Surfactant was also seen in the air spaces in DH + TO and CTR; it was absent in unobstructed CDH and in TL.
Concfusions:Temporary tracheal occlusion in utero does not cause the dramatic decrease in type II pneumocytes seen after prolonged occlusion. Although only minimal increase in lung volume is seen in CDH, catch-up parenchymal growth and maturation occur, most notably in the surfactant- producing system. J Pediatr Surg 33:1030-1037. Copyright o 1998 by W.B. Saunders Company.
INDEX WORDS: Fetal surgery, congenital diaphragmatic her- nia, pulmonary hypoplasia, type II pneumocyte, surfactant, sheep.
I NFANTS born with congenital diaphragmatic hernia (CDH) continue to have a high perinatal mortality
rate.‘s2 This mortality rate has been attributed to pulmo- nary hypoplasia,3*4 quantifiable as low lung weight to body weight ratio or low lung DNA content.5 There is mounting evidence, however, that the lungs of the fetus
From the Division of Pediatric Surgery, Department of Surger?/ and the Department of Pathology, Brown University School of Medicine, Providence, RI.
Presented at the 1997 Annual Meeting of the Section on Surgery of the American Academy of Pediatrics, New Orleans, Louisiana, October 31-November 2, 1997.
Address reprint requests to FranGois 1. Luks, MD, Hasbro Children 5 Hospital, Division of Pediatric Surgery 2 Dudley St, Suite 180, Providence, RI 02905.
Copyright o I998 by WB. Saunders Company 0022-3468/98/3307-0013$03.00/O
with CDH are not only smaller but immature as well. This has been observed in animal model&* and in human autopsy cases, 3,9 and is believed to be at the basis of the surfactant deficiency found in newborns with CDH.l”-12
In utero tracheal obstruction, a recently proposed treatment modality for CDH, was found to cause acceler- ated lung growth.5,13-*8 We have demonstrated that it produces accelerated maturation as well. After tracheal obstruction, the lungs of fetal rabbits progressed through successive stages of development more rapidly than their control littermates.lg This accelerated maturation could help explain the major drawback of tracheal ligation in utero: its dramatically detrimental effect on the type II pneumocyte population. Progressively longer periods of tracheal ligation in a fetal lamb model resulted in a gradual decrease in the number of type II pneumocytes; by 6 weeks after tracheal ligation, there was a virtual
1030 JournalofPediafric Surgery,Vol33, No7 (July),1998:pp 1030-1037

TEMPORARY TRACHEAL OCCLUSION IN CDH 1031
depletion of type II pneumocytes.‘O Similarly, others have found a dramatic surfactant deficiency after prolonged tracheal ligation.” We have therefore postulated that these alterations are a result of excess lung maturation, and that the few remaining type II pneumocytes, after 4 weeks of tracheal ligation in utero, are “postmature” cells incapable of releasing surfactant.20
When, in an effort to compensate for their inherent pulmonary immaturity and hypoplasia, fetal lambs with surgically created CDH were subjected to 4 weeks of tracheal ligation, O’Toole et alz2 again found severe surfactant deficiency and paucity of type II pneumocytes. We hypothesize that, even in CDH-associated pulmonary hypoplasia and immaturity, this prolonged tracheal ob- struction leads to pulmonary overmaturation. If this is true, shortening the time of tracheal obstruction in a CDH model would allow compensation of lung hypoplasia and immaturity but avoid maturation “overshoot.” Figure 1 schematically illustrates the rationale of causing catch-up maturation in CDH. To test this hypothesis, an early fetal lamb model of diaphragmatic hernia, known to produce severe pulmonary hypoplasia’0.23J4 was subjected to temporary tracheal occlusion.
MATERIALS AND METHODS
Experimental Design
Twenty-five fetal lambs were divided in four groups. A first group (DH + TO. n = 7) had a diaphragmatic hernia created at 65 to 75 days’ gestation (term, 145 d). At 108 days, the trachea was occluded with a balloon catheter. After 14 days the balloon was deflated, relieving the obstruction. During the 2 weeks of occlusion the lung fluid was aspirated, measured, and reinfused every other day. A second group consisted of fetuses with CDH, without subsequent tracheal occlusion (DH, n = 7). These two groups were compared with positive controls, consisting of 4-week tracheal ligation without underlying CDH (TL, n = 3)20 and negative controls (unoperated littermates, CTR, n = 8).
OVER- MATURATION
,-‘~CCDH+4WkTO
Fig 1. Schematic representation of working hypothesis: If diaphrag- matic hernia (CDH) lungs are immature and 4 weeks of tracheal obstruction (TO) causes pulmonary “over maturation” in normal and in CDH fetuses, temporary tracheal occlusion (CDH + 2 wk TO) might cause catch-up maturation (= normal lungs) without “overshoot.”
Creation of Diaphvagmutic Hernia The model described by Pringle et al?? was used with some
modifications. Time-dated pregnant ewes (65 to 75 days) underwent midline laparotomy under general halothane anesthesia (0.5% to 2.0% in 100% 02) after intravenous induction with ketamine (1 g). A small hysterotomy was made transversely near the uterine horn with exterior- ization of the fetal hind limbs. Care was taken not to occlude the umbilical vessels as the fetal rump was stabilized outside the uterus. A left-sided thoracotomy was performed between the lowermost ribs. The diaphragm was identified and partially excised. Stomach and viscera were then pulled into the thoracic cavity. The thoracotomy incision was closed in one layer with interrupted 4-O polypropylene perichondrial sutures (Prolene, Ethicon; Johnson & Johnson, Somerville, NJ). Fetal exposure time was kept to a minimum and remained under 8 minutes in all fetuses. The fetus was then returned to the uterus. Antibiotics (ampicillin, 200 mg and chloramphenicol, 250 mg) were added to the amniotic fluid. The hysterotomy was closed using a continuous two-layer closure.
Tracheal Occlusion
At 108 days DH + TO fetuses underwent reoperation. Under the same anesthetic conditions, a hysterotomy was performed, and the fetal head and neck were exteriorized. A midline cervical incision was used to expose the trachea. A tracheotomy was made immediately distal to the cricoid cartilage and a balloon catheter (no. 6 Fogarty arterial embolectomy catheter; Baxter Carp; Vascular systems division, CA) was inserted caudaily and inflated with 1.5 mL of sterile saline. A second tracheotomy was made distal to the balloon and a 19-gauge (inner diameter) flanged tygon catheter was inserted into the distal trachea. Both were secured using 4-O polypropylene sutures. The lung fluid was aspirated, quantified, and reinfused. Fetal head and neck were returned to the uterus. A 17-gauge amniotic catheter was placed in the uterus, antibiotics (ampicillin, 200 mg and chloramphenicol, 250 mg) were added to the amniotic fluid, and the hysterotomy was closed. The catheters were “Witzeled,” and the abdominal wall was closed after ensming adequate slack on the catheters, which were then exteriorized into a pouch on the ewe’s flank.
Lung fluid was aspirated, quantified. and reinfused every other day during the 14-day tracheal occlusion period. Every time, antibiotics were added to the amniotic fluid via the amniotic catheter after each intervention. After 14 days of occlusion (122 days’ gestation) the embolectomy balloon was deflated. The fetus continued to receive intraamniotic antibiotics every other day for the remainder of the experiment.
The DH animals underwent a sham operation at 108 days, with aspiration and quantification of lung fluid volume. The three TL animals underwent tracheal ligation at 108 days, as previously described.?O Near term (136 d) the ewe underwent a cesarean section after induction with ketamine (1 g). The animal was killed by intravenous injection of a euthanasia solution (10 mL of Beuthanasia-D Special; Shering-Plough Animal Health Corp, Kenilworth, NJ).
Estimation of Pulmonary Hypoplasia
The severity of the diaphragmatic hernia (ie, the degree of pulmonary hypoplasia) was estimated at 108 days’ gestation by quantification of total lung fluid volume. An earlier pilot experiment, based on eight fetuses with normal lungs, and various degrees of pulmonary hypopla- sia. had shown excellent con-elation between lung fluid volume at 108 days and lung weight at term (unpublished data). Data from the present study were superimposed on the pilot graph to estimate the degree of pulmonary hypoplasia after creation of a-diaphragmatic hernia and to predict lung weight at term.

PAPADAKIS ET AL
Tissue Processing
The fetus’s body weight was recorded, and the lungs and trachea were removed en bloc. Wet lung weight was determined, small samples from the right upper lobe were fixed for electron microscopy (Karnovsky’s fixative: 4% paraformaldehyde, 2.5% glutaraldehyde, 0.1 mol/L sodium cacodylate buffer, pH 7.2) and the remainder of the trachea-lung block was fixed with 10% buffered formaldehyde at 25 cm Hz0 for at least 1 week. Lung volume was then estimated by the volume displacement method. The lungs were cut in standardized sections (3 to 5 mm thick), random tissue blocks were taken from the center and periphery of each lobe and embedded in paraflin. H&E-stained slides (4 u thick) were prepared from each paraffin block (four per animal).
Stereologic Morphometric Analysis of Lung Growth
Tissue sections from both lungs, stained with H&E, were analyzed morphometrically using a computerized image analysis system (micro- scope interfaced via a charged-coupled device (CCD) video camera (KP-161; Hitachi, Woodbury, NY) to a Power Macintosh 7100/8OAV (Apple Cot-p, Cupertino, CA) equipped with Image NIH 1.59 for Macintosh (National Institutes of Health, Bethesda, MD) image analy- sis software.
The critical data set and hierarchical equations, obtained by examin- ing the lungs at increasing levels of magnification, have been described in detail elsewhere.“O Briefly, the total volume of fixed lungs (V&J) was determined according to the Archimedes principle. Subsequent steps in the structural hierarchy involved point counting methods based on computer-assisted image analysis. The parenchymal area1 density (An[pdZu]) was estimated by dividing the number of points falling on parenchyma (ie, excluding large-sized bronchi and blood vessels) by the number of points falling on the entire lung (magnification X 10). The parenchymal volume (V[pa]) was determined by multiplying V(lu) by AA(pa/lu). The areal density of air exchanging parenchyma (AJae/pa]) was estimated by studying random fields of peripheral lung parenchyma and dividing the number of points falling on air exchanging parenchyma (ie, peripheral lung parenchyma but not airspace) by the number of points falling on the entire field (tissue and airspace; magnification X100). A,(ae/pa) represents the tissue fraction of the lung, and as such is complementary to the airspace fraction (ASF), a traditional measure of fetal lung maturity (ASF = 1 - A,[ae/pa]). The total volume of air exchanging parenchyma (V[ue]) was calculated by multiplying its volume density by V(pa).
As an index of alveolar proliferation and architectural maturity, the radial alveolar count @AC), as originally described by Emery and Mithalzs was calculated. H&E-stained sections were examined at magnification X 40 and perpendicular lines were drawn from the lumen of respiratory bronchioles to the nearest connective tissue septum. The number of alveolar septa intercepted by this line (RAC) was counted as an index of the development of the terminal respiratory unit.
Stereologic Morphometric Analysis of Type I1 Pneumocyte Population
Two sections per lung (central upper lobe, peripheral lower lobe) were processed for avidin-biotin-immunoperoxidase staining using a polyclonal rabbit antibody against surfactant protein B (SP-B). The polyclonal antibovine SP-B antiserum was kindly provided by Dr J.A. Whitsett.26 After treatment with 3,3’-diaminobenzidine tetrachloride (DAB; Sigma, St Louis, MO), sections were lightly counterstained with hematoxylin, cleared, and mounted. Controls for specificity consisted of omission of the primary antibody.
The areal density of the SP-B-immunoreactive compartment (A,[pdZ/ nej) was evaluated automatically, based on the fact that the immunohis-
tochemical staining of type II pneumocytes produces a higher gray level than that of the background (magnification X200). The total type II pneumocyte volume (V[pnIl]) was calculated by multiplying its area1 density by V(ae). To determine the individual type II cell profile area, surfactant-positive cells were manually traced at a final magnification of X600 and their profile area and radius (r-) determined. At least 50 cells per animal were analyzed from five different microscope fields. The individual pneumocyte II volume (V[pnIIcell]) was calculated by the following formula, based on the observation that the shape of the fetal type II pneumocyte approaches a sphere: V(pnIIcel1) = 4/3 ara. The total number of type II pneumocytes per lung (N[p&rj) was calculated by dividing total surfactant-positive volume by individual type II cell volume (V[pnII]N[pnIIcell]). Finally, the numerical density of type II pneumocytes (NM[pnII/ue]), representing the number of type II pneumo- cytes per unit of air exchanging parenchymal volume, was determined by dividing N(pnI1) by V(ae).
Transmission Electron Microscopy
For transmission electron microscopy, lung tissue fragments were fixed in Karnovsky’s fixative, osmicated, dehydrated in ethanol, and embedded in epoxy-resin. Ultrathin sections were stained with uranyl acetate and lead citrate, and examined with a Philips 301 electron microscope (Philips NV, Eindhoven, Netherlands) operated at 60 kV.
Statistical analysis was performed using the Student’s t test or single factor analysis of variance (ANOVA) with posthoc Scheffe test (Statview for Macintosh: Abacus Concepts, Berkeley, CA). Where appropriate, values are expressed as mean i SD. A P value of less than .05 was considered significant. Animals were treated according to current guidelines of animal welfare, and all procedures and protocols were approved by the Brown University Institutional Animal Care and Use Committee.
RESULTS
There was an overall survival rate of 92% after the first procedure. Four DH and four DH + TO animals survived to term. Lung fluid volume at 108 days showed a good exponential correlation (r = 0.967, r2 = 0.936) with abso- lute lung weight at term for CTR and DH animals, confirming the validity of this parameter as an estimation of the degree of pulmonary hypoplasia at term and the severity of the DH. Interestingly, all four DH + TO fetuses had greater lung weights than predicted at 108 days, according to the best fit curve (y = 5.23x + 13.4), suggesting some degree of lung growth after temporary tracheal occlusion. In DH + TO animals, lung fluid volume increased ninefold (from 5.2 -+ 4.4 mL at day 0 of TO [lo8 d] to 48.3 + 13.3 mL by 1 week postocclu- sion), again suggesting cell proliferation and lung growth. Lung and body weight are summarized in Table 1.
Histology
The lungs of CTR fetuses were divided into relatively large alveolar units, well defined by prominent alveolar septa (Fig 2A). The parenchyma was composed of regular thin-walled airspaces, most of which had the appearance of alveoli. The interalveolar septa appeared as thin strips of fibrous tissue containing a single capillary
TEMPORARY TRACHEAL OCCLUSION IN CDH 1033
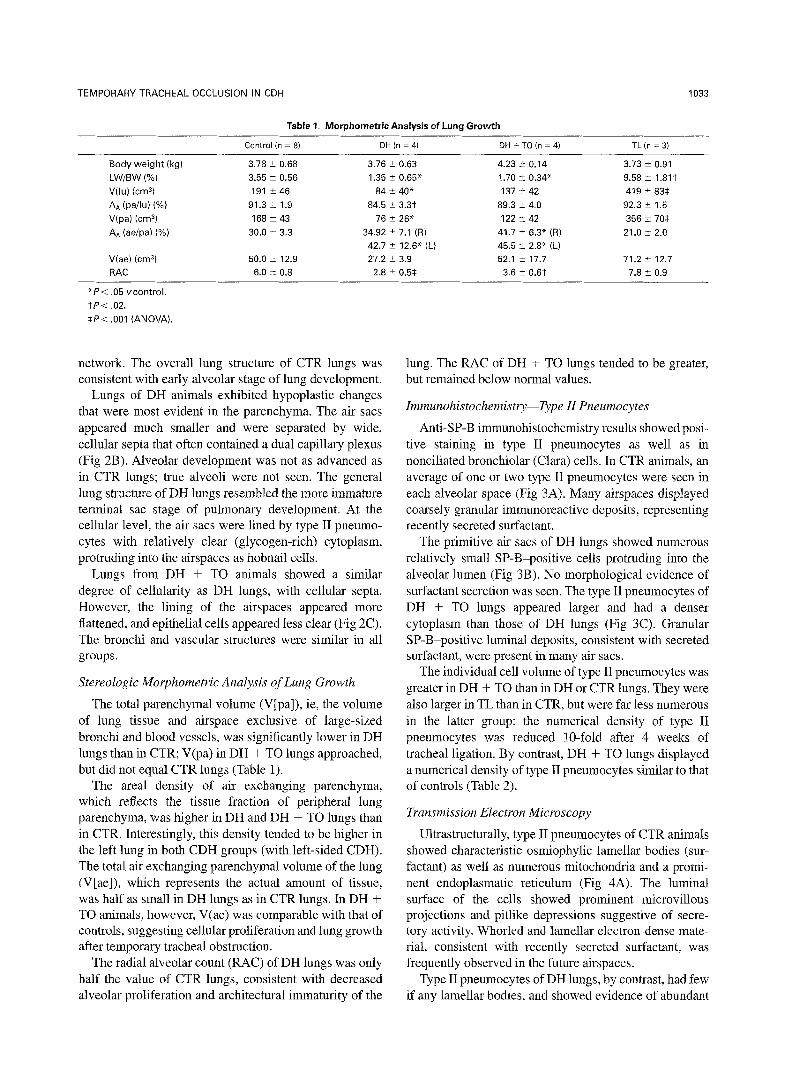
Table 1. Morphometric Analysis of Lung Growth
Control hl = 8) DH (n = 4) DH + TO h = 4) TL (n = 31
Body weight (kg)
LW/BW (%)
V(lu) icm3) AA (pailu) (%)
V(pa) b2n-G)
AA (se/pa) (%)
V(ae) (cm31
RAC
3.78 k 0.68
3.55 2 0.56 191 i 46
91.3 + 1.9
168 2 43 30.0 t 3.3
50.0 2 12.9
6.0 t 0.8
3.76 2 0.63
1.35 k 0.65* 84 t40*
84.5 k 3.3t
76 5 26* 34.92 -t 7.1 (R)
42.7 2 12.6* (L) 27.2 i 3.9
2.8 t 0.5$
4.23 i- 0.14
1.70 t 0.34' 137 t 42
89.3 + 4.0
122 i- 42 41.7 2 6.3" (R)
45.5 t 2.8" (L) 52.1 i 17.7
3.6 2 0.6t
3.73 i 0.91
9.58 ? 1.81t 419 2 83$
92.3 i 1.5
356 I 70s 21.0 t 2.0
71.2 + 12.7
7.8 i 0.9
*Pi .05 vcontrol.
tP< .02.
SP< ,001 (ANOVA).
network. The overall lung structure of CTR lungs was consistent with early alveolar stage of lung development.
Lungs of DH animals exhibited hypoplastic changes that were most evident in the parenchyma. The air sacs appeared much smaller and were separated by wide, cellular septa that often contained a dual capillary plexus (Fig 2B). Alveolar development was not as advanced as in CTR lungs; true alveoli were not seen. The general lung structure of DH lungs resembled the more immature terminal sac stage of pulmonary development. At the cellular level, the air sacs were lined by type II pneumo- cytes with relatively clear (glycogen-rich) cytoplasm, protruding into the airspaces as hobnail cells.
Lungs from DH + TO animals showed a similar degree of cellularity as DH lungs, with cellular septa. However, the lining of the airspaces appeared more flattened, and epithelial cells appeared less clear (Fig 2C). The bronchi and vascular structures were similar in all groups.
Stereologic Morphometric AnaLysis of Lung Growth
The total parenchymal volume (V[pa]), ie, the volume of lung tissue and airspace exclusive of large-sized bronchi and blood vessels, was significantly lower in DH lungs than in CTR; V(pa) in DH + TO lungs approached, but did not equal CTR lungs (Table 1).
The area1 density of air exchanging parenchyma, which reflects the tissue fraction of peripheral lung parenchyma, was higher in DH and DH + TO lungs than in CTR. Interestingly, this density tended to be higher in the left lung in both CDH groups (with left-sided CDH). The total air exchanging parenchymal volume of the lung (V[ae]), which represents the actual amount of tissue, was half as small in DH lungs as in CTR lungs. In DH + TO animals, however, V(ae) was comparable with that of controls, suggesting cellular proliferation and lung growth after temporary tracheal obstruction.
The radial alveolar count (RAC) of DH lungs was only half the value of CTR lungs, consistent with decreased alveolar proliferation and architectural immaturity of the
lung. The RAC of DH + TO lungs tended to be greater, but remained below normal values.
Immunohistochemist~-Type II Pneumocytes
Anti-SP-B immunohistochemistry results showed posi- tive staining in type II pneumocytes as well as in nonciliated bronchiolar (Clara) cells. In CTR animals, an average of one or two type II pneumocytes were seen in each alveolar space (Fig 3A). Many airspaces displayed coarsely granular immunoreactive deposits, representing recently secreted surfactant.
The primitive air sacs of DH lungs showed numerous relatively small SP-B-positive cells protruding into the alveolar lumen (Fig 3B). No morphological evidence of surfactant secretion was seen. The type II pneumocytes of DH + TO lungs appeared larger and had a denser cytoplasm than those of DH lungs (Fig 3C). Granular SP-B-positive luminal deposits, consistent with secreted surfactant, were present in many air sacs.
The individual cell volume of type II pneumocytes was greater in DH + TO than in DH or CTR lungs. They were also larger in TL than in CTR, but were far less numerous in the latter group: the numerical density of type II pneumocytes was reduced lo-fold after 4 weeks of tracheal ligation. By contrast, DH + TO lungs displayed a numerical density of type II pneumocytes similar to that of controls (Table 2).
Transmission Electron Microscopy
Ultrastructurally, type II pneumocytes of CTR animals showed characteristic osmiophylic lamellar bodies (sur- factant) as well as numerous mitochondria and a promi- nent endoplasmatic reticulum (Fig 4A). The luminal surface of the cells showed prominent microvillous projections and pitlike depressions suggestive of secre- tory activity. Whorled and lamellar electron-dense mate- rial, consistent with recently secreted surfactant, was frequently observed in the future airspaces.
Type II pneumocytes of DH lungs, by contrast, had few if any lamellar bodies, and showed evidence of abundant

1034 PAPADAKIS ET AL
Fig 2. Parenchymal lung tissue at term. (A) normal lungs KTRI. Note complex alveolar architecture and thin alveolar lining. IB) DH-associated pulmonary hypoplasia and immaturity. Note thick, cellular air space (terminal sac) walls. (C) DH + TO. Note alveolar pattern, thinner septa than in B. (H&E, original magnification x100.)
pneumocytes in TL lungs showed cytoplasmic degenera- tive changes, with marked mitochondrial swelling, vacu- olization of the cytoplasm, and poorly preserved lamellar inclusions. No surfactant was seen in the airspaces.
glycogen granules (washed out by fixation, and appearing as whited-out areas, Fig 4B). No surfactant was seen in the lumen. In DH + TO, however, large type II pneumo- cytes exhibited signs of secretory activity, a dense concentration of lamellar bodies, and a very prominent endoplasmatic reticulum. Here, as in CTR, surfactant was
Fig 3. lmmunoperoxidasestain for type II pneumocytes, which appear as dark, round cells. (Al normal lungs. Note thin alveolar lining, one to two type II pneumocytes per alveolus, and surfac- tant secreted in lumen (arrows). (B) DH lungs. Numerous, but smaller type II pneumocytes, lining air sacs in-“hobnail” fashion. No surfactant in lumen. (Cl DH + TO. Alveolar wall thicker than in CTR but flattened compared with DH. Note surfactant secretion in alveolar spaces (arrows; ABC immunoperoxidase stain for antisur-
seen in the luminal spaces (Fig 4C). The sparse type II factant-B protein, original magnification x400).

TEMPORARY TRACHEAL OCCLUSION IN CDH
Table 2. Morphometric Analysis of Type II Pneumocyte Population
Control DH DH + TO TL (n = 81 (n = 41 (I- = 4) (n = 3)
AA (pnlliae) (%) 10.0 t 5.9 7.0 f 2.1 10.3 t 2.6 1.7 k 0.6 V (pnllcell) (p3) 136 t 42 173 +- 12 354 i 42* 248 2 91
Nv (pnlliae) (lOs/cms) 826 ? 324 404 f 107 294 2 74 60 2 22t
‘P< .05 vcontrol.
tP < ,001. Note pneumocyte II numerical density (Nv [pnll/ael) one order of magnitude smaller in TLthan in all other groups.
DISCUSSION
The pathophysiology of congenital diaphragmatic her- nia and its concomitant pulmonary pathology is still not understood fully. Although it has been known for many years that CDH is associated with pulmonary hypopla- sia,3,7 it is now becoming apparent that this is accompa- nied by a delay in pulmonary maturation as we11.6~9JoJ2~27 Lung immaturity may be measured by two components: the structure of the lung and its cellular makeup. Architec- tural immaturity of the hypoplastic lung can manifest itself by a reduction in the number of generations of bronchiolar branching, a lesser degree of alveolar com- plexity as evidenced by a low radial alveolar count (RAC), and by thickened alveolar septa, all suggestive of earlier developmental stages.6,9,23.28 The cellular immatu- rity of these lungs is characterized by the presence of numerous, but immature, type-II pneumocytes unable to produce surfactant. Ultrastructurally, these cells are filled with glycogen but contain few lamellar bodies,24,27 which is consistent with a developmental arrest in the canalicu- lar stage.5 It has been suggested that lung immaturity and
A
1035
surfactant deficiency are at least partially responsible for the respiratory failure seen in infants with CDH.1°J2
Tracheal occlusion, however, causes accelerated lung growth, which is able to compensate for the CDH- associated lung hypoplasia14.29 and has been applied through open fetal surgery” or via minimal access techniques.16x30,31 However, the improvement in pulmo- nary hypoplasia comes at a cost. It is associated with a surfactant deficiency that is even more profound than the one present in CDH.21,22
Thus, both CDH and (prolonged) tracheal obstruction are associated with surfactant deficiency, albeit through different mechanisms. Whereas the absence of alveolar surfactant in CDH appears to be the result of a develop- mental arrest of the lungs and their type II pneumocyte population, the surfactant deficiency after tracheal occlu- sion is secondary to a virtual disappearance of surfactant- producing type II pneumocytes; the sparse cells that remain are enlarged and irregular and appear to be engorged with surfactant that they cannot secrete.20 Although the cause of this decline in pneumocyte II cell population is unknown, it is likely the result of one or more of the following mechanisms: decreased prolifera- tion, increased cell death (apoptosis or other), or in- creased differentiation into type I pneumocytes, their normal progeny.3’ Because tracheal occlusion results in pulmonary hyperplasia, mostly of alveolar-lining type I pneumocytes, it is tempting to ascribe the vanishing of type II pneumocytes to accelerated terminal differentia- tion into type I cells. This, and the finding that tracheal
B
Fig 4. Transmission electron microscopy of type II pneumocytes. IAj CTR. Numerous mitochondria, abundant lamellar bodies; pneumocyte in lower right corner displays “pitlike” depression of cell membrane, indicating recent secretory activity. (Bj DH. Immature type II pneumocyte, projecting more prominently into the air space, with washed-out glycogen granules fgj and only sparse lamellar bodies (arrow). (C) DH + TO. Mature type II pneumocytes with overabundant lamellar bodies fsurfactantj. (Original magnification ~10,155.)

1036
occlusion accelerates fetal lung development,lg could suggest that prolonged tracheal occlusion results in an exaggerated, and ultimately detrimental maturation pro- cess.
If this exaggerated response is responsible for the observed surfactant deficiency after four weeks of tra- cheal ligation in the fetal lamb with diaphragmatic hernia,22 shorter periods of tracheal occlusion may cause catch-up maturation without “overshoot.” The current experiment, whereby tracheal occlusion was maintained for only 2 weeks, seems to achieve just this. After occlusion, net lung fluid production promptly rose, suggesting cellular proliferation (as previously demon- strated in a non-CDH model of tracheal ligation33). At term, this 2-week occlusion did result in moderate hyperplasia compared with unobstructed CDH fetuses.
It had, however, a marked effect on pulmonary matu- rity. Whereas in fetuses with diaphragmatic hernia alone, lungs were clearly arrested in an earlier stage (by histological and ultrastructural criteria), the lungs of fetuses with CDH and tracheal occlusion showed architec- tural maturity similar to that of controls, with flattened alveolar epithelial lining and an increased radial alveolar count. The most striking findings were the presence of
PAPADAKIS ET AL
mature type II pneumocytes by electron microscopy. At the light microscopic level, this resulted in the presence, in the alveolar spaces, of surfactant protein. In contrast, unobstructed DH lungs did not show any surface-active substance in the air spaces. As expected, prolonged tracheal occlusion (4 weeks) resulted in a virtual absence of type II pneumocytes and no demonstrable surfactant in the alveolar spaces.
Temporary tracheal occlusion allows catch-up pulmo- nary maturation to such a degree that surfactant is now produced and secreted into the air spaces. Timely release of the obstruction helps avoid the nefarious effect of this procedure on the type II pneumocyte population, thereby avoiding surfactant deficiency by exaggerated maturation and growth.
It remains to be determined whether it is better, from a clinical stand point at birth, to produce large lungs, even if they lack surfactant, or to preferentially obtain catch-up lung maturation at the expense of a less-impressive lung growth. In addition, the long-term outcome and eventual regeneration of type II pneumocytes has not been studied yet. It might well be that tracheal obstruction produces hyperplastic lungs that are only temporarily devoid of surfactant-producing cells.
REFERENCES
1. Adzick NS, Vacanti JP, Lillehei CW, et al: Fetal diaphragmatic hernia: Ultrasound diagnosis and clinical outcome in 38 cases. J Pediatr Surg 24:654-658, 1989
2. Harrison M, Adzick NS, Estes JM, et al: A prospective study of the outcome for fetuses with diaphragmatic hernia. JAMA 271:382-384, 1994
3. Areechon W, Reid L: Hypoplasia of lung with congenital diaphrag- matic hernia. Br Med J 1:230-233, 1963
4. Nguyen L, Guttman F, De Chadaverian JP, et al: The mortality of congenital diaphragmatic hernia. Is total pulmonary mass inadequate, no matter what?Ann Surg 198:766-770, 1983
5. Jobe AH: Fetal lung development, tests for maturation, induction of maturation, and treatment, in Creasy RK, Resnick R (eds): Matemal- fetal medicine. Philadelphia, PA, Saunders, 1994, pp 423-441
6. Alfonso LF, Vilanova J, Aldazabal P, et al: Lung growth and maturation in the rat model of experimentally induced congenital diaphragmatic hernia. Eur J Pediatr Surg 3:6-11, 1993
7. Kitagawa M, Hislop A, Boyden EA, et al: Lung hypoplasia in congenital diaphragmatic hernia. A quantitative study of airway, artery and alveolar development. Br J Surg 58:342-346, 1971
8. Schnitzer JJ, Hedrick HL, Pacheco BA, et al: Prenatal glucocort- coid therapy reverses pulmonary immaturity in congenital diaphrag- matic hernia in fetal sheep. Ann Surg 224:430-439, 1996
9. Askenazi SS, Perlman M: Pulmonary hypoplasia: Lung weight and radial alveolar count as criteria of diagnosis. Arch Dis Child 54:614-618, 1979
10. Glick PL, Stannard VA, Leach CL, et al: Pathophysiology of congenital diaphragmatic hernia II: The fetal CDH lamb model is surfactant deficient. J Pediatr Surg 27:382-388, 1992
11. Suen HC, Catlin EA, Ryan DP, et al: Biochemical immaturity of lungs in congenital diaphragmatic hernia. J Pedian Surg 28:471-477, 1993
12. Moya FR, Thomas VL, Romaguera J, et al: Fetal lung maturation
in congenital diaphragmatic hernia. Am J Obstet Gynecol 173:1401- 1405,1995
13. Carmel JA, Friedman F, Addams FH: Fetal tracheal ligation and lung development. Am J Dis Child 109:452-456,1965
14. Nardo L, Hooper SB, Harding R: Lung hypoplasia can be reversed by short term obstruction of the trachea in fetal sheep. Pediatr Res 38:690-696, 1995
15. DiFiore JW, Fauza DO, Slavin R, et al: Experimental fetal tracheal ligation reverses the structural and physiological effects of pulmonary hypoplasia in congenital diaphragmatic hernia. J Pediatr Surg 29:248-255, 1994
16. Flageole H, Evrard VA, Vandenberghe K, et al: Tracheoscopic tracheal occlusion in the ovine model. J Pediatr Surg 32:1328-1331, 1997
17. Skarsgard ED, Meuli M, VanderWall KJ, et al: Fetal endoscopic tracheal occlusion (‘Fetendo-PLUG’) for congenital diaphragmatic hernia. J Pediatr Surg 31:1335-1338, 1996
18. Luks FI, Gilchrist BF, Jackson BT, et al: Endoscopic tracheal obstruction with an expanding device in a fetal lamb model. Preliminary considerations. Fetal Diagn Ther 11:67-71, 1996
19. De Paepe ME, Papadakis K, Johnson BD, et al: Time-course study of accelerated lung growth in a fetal rabbit model. Am J Path01 152:179-190, 1998
20. De Paepe ME, Papadakis K, Johnson BD, et al: Fate of the type II pneumocyte following tracheal occlusion in utero: A time-course study in fetal sheep. Virchows Arch 432:7-16, 1998
21. O’Toole SJ, Karamanoukian HL, Irish MS, et al: Tracheal ligation: The dark side of in utero congenital diaphragmatic hernia treatment. J Pediatr Surg 32:407-410, 1997
22. O’Toole SJ, Sharma A, Karamanoukian HL, et al: Tracheal ligation does not correct the surfactant deficiency in congenital diaphrag- matic hernia. J Pediatr Surg 31:546-550, 1996

TEMPORARY TRACHEAL OCCLUSION IN CDH 1037
23. Pringle KC, Turner JW, Schofield JC, et al: Creation and repair of diaphragmatic hernia in the fetal lamb: Lung development and morphol- ogy. .I Pediatr Surg 19:131-140: 1984
24. Hashimoto EG, Pringle KC, Soper RT, et al: The creation and repair of diaphragmatic hernia in fetal lambs: Morphology of the type II alveolar cell. J Pediatr Surg 20:354-356, 1985
25. Cooney TP, Thurlbeck WM: The radial alveolar count method of Emery and Mithal: A reappraisal. 1. Postnatal lung growth. Thorax 37512-519, 1982
26. Voorhout WF, Veenedaal HP, Haagsman TE, et al: Intracellular processing of pulmonary surfactant protein B in an endosomal/ lysosomal compartment. Am J Physiol263:L479-486,1992
27. Asabe K, Tsuji K, Handa N, et al: Immunohistochemical distribution of surfactant apoprotein-A in congenital diaphragmatic hernia. J Pediatr Surg 321667.672, 1997
28. Hashim E, Laberge JM, Chen MF, et al: Reversible tracheal
obstruction in the fetal sheep: Effects on tracheal fluid pressure and lung growth. J Pediatr Surg 30:1172-1177, 1995
29. Wilson JM, Difiore JW, Peters CA: Experimental fetal tracheal ligation prevents the pulmonary hypoplasia associated with fetal nephrectomy: Possible application in congenital diaphragmatic hernia. J Pediatr Surg 28:1433-1440, 1993
30. Papadakis K, Luks FI, Deprest JA, et al: Single-port tracheo- scopic surgery in the fetal lamb. J Pediatr Surg (in press)
31. Luks FI, Deprest JA, Vandenberghe K, et al: A model for fetal surgery through &a-uterine endoscopy. J Pediatr Surg 29:1007-1009,1994
32. Mason RJ, Shannon JM: Alveolar type II cells, in Crystal RG, West JB, Weibel ER? et al, (eds): The Lung: Scientific Foundations. Philadelphia, PA, Lippincott-Raven, 1997, pp 534-555
33. Papadakis K, Luks FI, De Paepe ME, et al: Fetal lung growth after tracheal ligation is not solely a pressure phenomenon. J Pediatr Surg 32:347-351, 1997
Discussion
E. Beierle (Philadelphia, PA): The authors have shown the preservation of the integrity of the type II pneumo- cytes with a short period of tracheal obstruction in a fetal sheep model of congenital diaphragmatic hernia. I have a few questions about your study.
Several investigators, including those at the University of Florida, Boston, and Montreal, have demonstrated a profound increase in lung weight body to weight ratios, lung volumes, and radial alveolar counts in a congenital diaphragmatic hernia model with tracheal ligation. Also, the fetal physiologists Hooper and Harding have shown that two thirds of the increase in pulmonary DNA seen in the fetal sheep with tracheal ligation occurred very early, in fact between 2 to 7 days after initialization of tracheal ligation. Can you explain the absence of a statistically significant increase in the lung to body weight ratio and the radial alveolar count in your study between diaphragmatic hernia and your short-term ligation animals? Did you monitor the integrity of the embolectomy balloon in any manner?
Second, it is known that the fetal lung development is profoundly affected by the hormonal millieu of the fetus, especially the type II pneumocyte development and the hormone cortisol. Could the exclusion of sham-operated control animals from your study and the lack of fetal stressers in that group have affected your results and biased your conclusions?
K. Papadakis (response): Thank you for those com- ments. On your first question, we monitored the patency of the balloon or the effectiveness of it by not having any tracheal fluid leak around the catheter. And at the time when the balloon occlusion was removed, the catheter was again patent. We did not have any balloon failure during this time.
We did not measure directly any cortisol effects or any other prenatal stressors, but DH animals were sham- operated.
T.M. Crombleholme (Philadelphia, PA): I congratulate you on a nice study. I have one question, which is, have you had an opportunity to look at the tracheal ligation group specifically of the type II pneumocytes? Is in- creased tracheal pressure resulting in induction of apopto- sis of type II pneumocytes or is there a reversible arrest in their cell cycles?
K. Papadakis (response): That is also an ongoing topic in our laboratory, whether the deficit of type II pneumo- cytes is caused by apoptosis or increased differentiation into type I pneumocytes. We also agree with Hooper and Harding and show that the initial increase in lung growth occurs quickly after tracheal ligation. But what happens with these type II pneumocytes after a prolonged period is still up for investigation.