terrestrial ecology zoological part 2004. who is who? koos boomsma michael poulsen daniel kronauer
Post on 21-Dec-2015
213 views
TRANSCRIPT

Terrestrial Ecology
Zoological Part
2004

Who is who?
• Koos Boomsma
• Michael Poulsen
• Daniel Kronauer

What’s up for these two weeks?
• Exiting Evolutionary Ecology• A further confrontation with the hardship of
science• Straightforward textbook chapters versus…• ….recent (mostly) case studies of varying
complexity• Hot issues: Ageing, natural (social) conflicts,
infectious diseases (AIDS), conservation

The Issues
• Life Histories and Phenotypic Plasticity
• Conflict and Cooperation
• Parasites and Diseases
• Metapopulations and Conservation

Life Histories and Phenotypic Plasticity
Investments in and Timing of Growth and Reproduction

Broad Scale Life-history CorrelationsPregnancy Duration versus Body Size
May & Rubenstein, 1984 Clutton-Brock, 1991
Offspring carried
Offspringindependent
Offspring keptIn the nest

Broad Scale Life-history CorrelationsMaximal Life Span versus Body Size
Stearns, 1992
Not the slopes but the level is interesting
Prothero & Jürgens, 1992

Life Span is Tremendously Variable
274 Species of Invertebrates 170 Mammal Species in Zoos
Stearns, 1992
Comfort, 1979
Stearns, 1992
Eisenberg, 1981

Broad Scale Life-history CorrelationsEgg Volume versus Body Size
Residuals Contain the Important Information
.
Blueweiss et al., 1978 Clutton-Brock, 1991

The Comparative Method
The Statistical Analysis of Comparative (Across Taxa) Life History Data
But now ........ To the Explanations (Life History Theory)

Trade-off curves
Convex
ConcaveActual fitness contours
Option Sets
Iteroparity Annual Semelparity
14.10

Plots May also be the Other Way Around
Survival instead of Growth Trade-off Curves May also be Complex
Stearns, 1976, 1992

Cole’s Paradox – Why is Iteroparity so Common?
• Let Ba = # offspring Annual
• Let Bp = # offspring Perennial (Iteroparous)
• Annuals: Nt+1 = erNt = BaNt lnBa = r
• Perennials: Nt+1 = erNt = BpNt + Nt =
(Bp + 1)Nt ln(Bp + 1) = r
• The fitness of these two reproductive types is equal when: Ba = Bp + 1.
• ????? Annuals need to reproduce only marginally more to be selected for

Cole’s Paradox – Why is Iteroparity so Common?
• The Paradox was solved by including age-specific survival rates: pjuv (juveniles) and pad (adults)
• Now the fitness of these two reproductive types is equal when:
ada p
juv
pB B
p
• Conclusion: Because pad >> pjuv in many populations, it is often best to be iteroparous
• See Compendium for Details

The Cost of ReproductionTrade-off clear
unclear
Offspring Size versus Offspring #
14.11
14.17
High CRLobelia’s on Mt. Kenia

Problems in the Measurement of Trade-offs
Stearns, 1992
Survival
Reproduction
Fraction to R
A = R + S Var A >> Var B
Var A << Var B
Trade-offs (genetic correlations) may be invisible in the field

Clutch Size Optimisation
Assume a single optimal egg size Lack’s optimal clutch size
Iteroparous organisms need reserves to buffer the cost of reproduction and to minimise the temporal variation in reproductive performance

Clutch Size Optimisation
Geometric mean fitness is often a better measure than arithmetic mean fitness √Y1.Y2.Y3.Y4.....Yn
n
Boyce & Perrins, 1987 Cockburn, 1991
Large SD meanslarge Temporalvariation in Fitness

Clutch Size Optimisation
Other factors also play a decisive role: Laying date
Clutch size is a phenotypically plastic life-history traitDaan et al., 1990 Krebs & Davies, 1991
Model Predictions Match Observations in the Field
14.24

Size and Age at Maturity
Reznick & Endler, 1982 Cockburn, 1991
3 Streams with Different Predation Risk
C = High Adult Pr.R = Moderate Juv. Pr.A = Low Predation
% Female Biomass Reproduction
A transplantation experiment reproduced these patterns in 11 years (30-60 generations)
R: Size & Age at Maturity
C: ReproductiveEffort
R&C: Body Size ↓
Table 14.1

Size and Age at Maturity
Comparative data corrected for body size
14.27

Reproductive ValuePhlox drummondii
Age at maturity Life Span
Age at Maturity= Constant
Cockburn, 1991 Charnov & Berrigan, 1991
But only within taxa
14.16
cf. 14.4

Sex ratio• Should be measured in terms of investment
• Is often but far from always 50:50 at the end of parental investment
• The equilibrium ESS sex ratio is independent of an XX/XY sex chromosome system
• Adult sex ratios may be very skewed owing to sex specific mortality or mating success
• Is often skewed in haplo-diploid parasitoids and social insects (ants, bees, wasps)
• See Compendium for Details

Sex ratio and Cost of Reproduction
Only females in their prime age can reproduce each year
Male calfs are usually more ”expensive”Clutton-Brock, 1984, 1991 C
lutt
on
-Bro
ck, 1
98
1, 1
99
1

Sex ratio and Cost of Reproduction
a: daughters are more expensive
A paper on human twins of different sex
Clutton-Brock et al., 1982
Sons
Daughters
b,c,d: sons are more expensive

Why does almost every multicellular organism senesce?
• Germ-line and Soma are separated
• Soma is disposable if that serves the fitness of the germ-line
• Selection does not remove deleterious mutations expressed late in life
• Selection favors mutations that are beneficial early in life, even if they are bad later in life

The Optimal Repair Model
3 papers this afternoonKirkwood, 1985 Stearns., 1992
Excess Repair is not Favoured by Selection
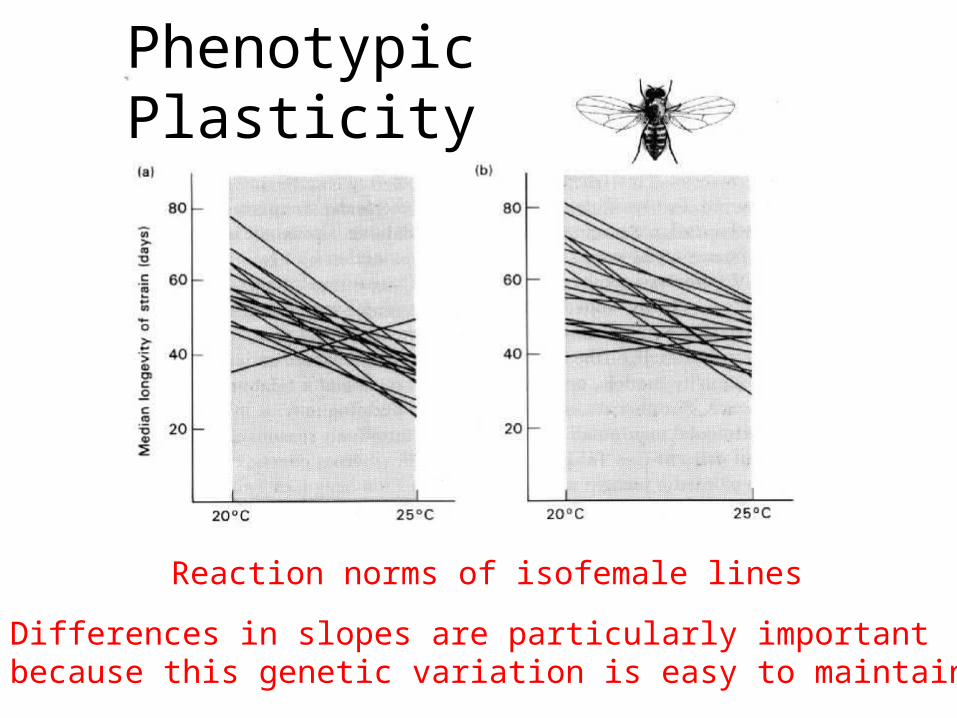
Phenotypic Plasticity
Reaction norms of isofemale lines
Differences in slopes are particularly importantbecause this genetic variation is easy to maintain

Reaction Norm Theory
Size and Age at Maturity Reproductive Effort versus Survival
Stearns, 1989, 1992

Practical Examples
Drosophila mercatorum Human females
Gebhardt & Stearns, 1988; Stearns, 1992 Stearns & Koella, 1986; Stearns, 1992
14.22
Good Nutrition
Bad Nutrition

Summary
• Life-history traits are heritable, but usually in a phenotypically plastic way
• Many key aspects of life are determined by selection on life-history traits
• Reproduction is costly and has a carefully balanced, but context dependent, economy
• In plants, animals, (microorganisms), and humans• 3 papers on ageing and 1 on early growth effects