terrestrial environment
TRANSCRIPT

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 1/28
ENVIRONMENTAL MICROBIOLOGY
Terrestrial Environment
Neeru Narula and Manjula VasudevaDepartment of Microbiology
CCS Haryana Agricultural University
Hisar – 125 004
20-Apr-2006 (Revised 06-Mar-2007)
CONTENTS
Introduction
Rhizosphere
Phyllosphere
Brief account of microbial interactions
Competition
Rumen microbiology
Biofertilizers
Biological N2 fixation
Nitrogenase enzyme
The nif genes
Symbiotic N2 fixation Rhizobium
Frankia
Non-symbiotic N2 fixation
Azotobacter and Azospirillum
Mycorrhizae
Keywords
Terrestrial Environment, Rhizosphere, Phyllosphere, Brief account of microbial interactions, Competition,
Rumen microbiology, Biofertilizers, Biological N2 fixation, Nitrogenase enzyme, The nif genes, Symbiotic N2 fixation, Rhizobium, Frankia, Non-symbiotic N2 fixation, Azotobacter and Azospirilum, Mycorrhizae.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 2/28
2
Introduction
Terrestrial environment is the process occurring within the soil and on or near the plants that
influence the functioning of the ecosystem. The process of soil development involves
complex interactions among the parent material (rock, sand, glacial drift etc.), topography,
climate and living organisms.
The term soil refers to the outer, loose material of the earth’s surface, a layer distinctly
different from the underlying bedrock. Agriculturally, it is the region supporting plant life
and from which plants obtain their mechanical support and many of their nutrients.
Chemically, the soil contains a multitude of organic substances not present in the underlying
strata. For microbiologists, the soil environment is unique in many ways. It contains a vast
population of bacteria, actinomycetes, fungi, algae and protozoa. It is one of the most
dynamic sites of biological interactions in nature; and it is the region where occur many of
the biochemical reactions concerned in the destruction of organic matter, in the weathering of
rocks and in the nutrition of agricultural crops. The physical and chemical characteristics of
soil determine the nature of the environment in which microorganisms are found. These
environmental characteristics in turn affect the composition of the microscopic population both qualitatively and quantitatively. It is from the soil that the water, air and the inorganic
and organic nutrients are obtained. The soil serves as a buffer to the drastic changes that
occur above the ground.
The organisms like algae, lichens or mosses remain dormant on the dry rock and grow when
moisture is present. They are phototrophic and produce organic matter which supports the
growth of chemoorganotrophic bacteria and fungi. The number of chemoorganotrophs
increase directly with the degree of plant cover of the rocks. CO2 produced during respiration
by chemoorganotrophs is converted to carbonic acid (CO2 + H2O ------ H2CO3), which is
involved in the dissolution of lime stone rocks. Many chemoorganotrophs excrete organic
acids which further promote dissolution of rocks into smaller particles. Freezing and thawing
also cause cracks in the rocks. In these crevices, raw soil forms and pioneering higher plants
can develop. Plant roots penetrate into crevices and increase the fragmentation of the rock
and hence develop rhizosphere (soil that surrounds plant roots) microflora. When the plants
die, their remains are added to the soil and become nutrients for microbial development.
Minerals are further rendered soluble and as water percolates, it carries some of these
chemical substances deeper. As weathering proceeds, the soil increases in depth, thus
permitting the development of large plants and trees. Soil animals play an important role in
keeping the upper layers of the soil mixed and aerated. The movement of materials
downwards results in the formation of many layers known as the soil profile which is
dependent on climatic and other factors and takes hundreds of years to be formed.
The soil is a complex habitat with numerous microenvironments and niches. Microorganisms
are present in the soil primarily attached to soil particles. The most important factor
influencing microbial activity in surface soil is the availability of water, whereas in deep soil
nutrient availability plays a major role.
The organic fraction of soil, often termed humus, contains the organic carbon and nitrogen
which is needed for microbial development, is the dominant food reservoir. The greatest
microbial activity is in the organic-rich surface layers, especially in and around the
rhizosphere. The numbers and activity of soil microorganisms depend to a great extent on the
balance of nutrients present. In some soils carbon is not the limiting nutrient, but instead theavailability of inorganic nutrients such as phosphorous and nitrogen limit microbial
productivity.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 3/28
3
The deep soil surface, which can extend for several hundred meters below the soil surface, is
not a biological wasteland. A variety of microorganisms, primarily bacteria, are present in
most deep underground soils. In samples collected aseptically from bore holes drilled down
to 300 m, a diverse array of bacteria have been found including anaerobes such as sulphate
reducing bacteria, methanogens and homoacetogens and various aerobes and facultativeaerobes. Microorganisms in the deep subsurface presumably have access to nutrients because
groundwater flows through their habitats, but activity measurements suggest that metabolic
rates of these bacteria are rather low in their natural habitats. Compared to microorganisms in
the upper layers of soil, the biogeochemical significance of deep subsurface microorganisms
may thus be minimal. However, there is evidence that the metabolic activities of these buried
microorganisms may over very long periods be responsible for some mineralization of
organic compounds and release of products into the ground water (Fig. 1).
Fig. 1: Profile of mature soil (Biology of Microorganisms)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 4/28
4
Rhizosphere
Rhizosphere is the region immediately outside the root. It is a zone where microbial activity
is usually high. Hiltner in 1904 observed the zone of intense microbial activity around the
roots and named it as rhizosphere. The influence of the root on soil microorganisms starts
immediately after seed germination which increases as the plant grows and reaches a
maximum when plants have reached the peak of their vegetative growth. The bacterial count
is almost always higher in the rhizosphere than it is in region of the soil devoid of roots,
often many times higher. This is because roots excrete significant amounts of sugars, amino
acids, hormones and vitamins, which promote such an extensive growth of bacteria and fungi
that these organisms often form microcolonies on the root surface. Roots initially have little
or nomicrobial colonization but as the plants grow in the soil, the root exudates composed of
a mixture of nearly 18 amino acids, 10 sugars, 10 organic acids, mucilage and other
substances together with sloughed-off root cap and other cells and exerts influence on
microbial colonization. These nutrients allow the dormant spores to germinate. The
rhizosphere microorganisms influence plant growth by controlling the availability and uptake
of nutrients.
Phyllosphere
Phyllosphere is the surface of the plant leaf, and under conditions of high humidity, as in wet
forests in tropical and temperate zones, the microbial flora of leaves may be quite high. Leaf
surface carries a heterogenous population of microbes, which grow, reproduce and multiply
on leaves in dynamic equilibrium with the existing micro- and macro-environment. Many of
the bacteria on leaves fix nitrogen and nitrogen fixation presumably aids these organisms in
growing with the predominantly carbohydrate nutrients provided by leaves. The leaf surface
microbes are important in several ways. For instance, some of them are known to fix
atmospheric nitrogen for the benefit of higher plants, have antagonistic action against fungal
parasites, degrade plant surface waxes and cuticles, produce plant hormones, decomposes
plant material after leaf fall, activate plants to produce phytoalexins, have toxic effects on
cattle, act as a source of allerginic air-borne spores and influence the growth behaviour and
root exudation of plants.
The age and position of a leaf on the plant is an important factor for microbial colonization of
its surface. The physiological and biochemical status of the leaf greatly influences the
population and composition of the microflora. The earliest colonizers on newly formed leaves
have to face almost no competition as they are devoid of any microbes, and in fact, they
receive a potential supply of surface nutrients. But as they get established, they face arelatively hostile environment because of widely fluctuating temperatures and the incidence
of UV radiations. They may immediately grow utilizing the fresh supply of substrates present
on the leaf surface or lie dormant and inactive until the leaf becomes senescent.
The leaf surface medium comprises exudates, chemical compounds resulting from biological
activity of various microbes including nitrogen fixers and components resulting from
atmospheric pollution. The structure and the chemistry of the leaf surface influences the
occurrence of the plant surface colonizers apart from physical factors like temperature,
relative humidity, light and wind velocity which also interact with the leaf surface community
in various ways. Leaves at the seedling stage of plants usually harbour the least number of
microbes which increases as the plants age, reaching the maximum population only on leaveswhich start yellowing at maturity.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 5/28
5
Brief account of Microbial Interactions
Plants are exposed to very large numbers of microorganisms that are present in the soil and
are deposited on leaves and stems. Plants are the prime source of nutrients for
microorganisms because they are the main source of organic matter in the environment. They
provide nutrients indirectly from plant exudates, the shedding of leaves, pollen, etc. and also
from dead plant matter. In some cases, nutrients are provided directly to microorganisms that
form close associations with plants. Associations with plants can range from those that are
extremely detrimental to the plant, such as those with virulent pathogens, through interactions
which do not appear to influence plant growth, to beneficial ones such as those formed with
mycorrhizal fungi or nitrogen fixing bacteria. For most microorganisms, interactions with
growing plants extend no further than the colonization of the surfaces of stems, leaves and
roots because these are regions where exudates are available. Symbionts have developed
methods that permit them to enter the host and obtain direct access to nutrients.
For symbionts, such as Rhizobium and mycorrhizal fungi, the plant benefits is usually
positively related to the amount of root that becomes invaded and fairly high levels of infection being most beneficial.
A number of possible interactions may occur between two species. Odum has proposed the
following relations:
a) Neutralism in which the two microorganisms behave entirely independently
b) Symbiosis, the two symbionts relying upon one another and both benefiting the
relationship
c) Protocooperation, an association of mutual benefit to the two species but without the
cooperation being obligatory for their existence or for their performance of some
reaction
d) Commensalisms, in which only one species derives benefit while the other isunaffected
e) Competition, a condition in which there is a suppression of one organism as the two
species struggle for limiting quantities of nutrients, O2, space, or other common
requirements
f) Amensalism, in which one species is suppressed while the second is not affected,
often the result of toxin production
g) Parasitism and Predation, the direct attack of one organism upon another;
h) Synergism, in field situations is the possible synergistic effect in the plant between
inducing virus and other non related viruses which could be brought to those plants
from outside sources e.g. TMV and cucumber mosaic virus together cause a more
severe disease than either of them alone. It would thus seem unwise to introduceTMV into field grown tomatoes where the aphid-borne cucumber mosaic virus might
be present in surrounding areas and eventually be transmitted to the tomato plants.
Symbiosis is the result of interaction of the two partners and both partners are modified in
some way to achieve this. This must arise by the interchange of molecules between the
partners, which may act as signals which cause the partner to modify itself or which may
themselves cause modifications.
In the symbiosis, modifications to the host enable the partners to form the symbiosis, and the
structure and physiology of both host and microorganism are adapted for the aerobic fixation
of nitrogen. Two members are required for the association, a plant and microorganism. The
classical example of such a symbiosis is between leguminous plants and bacteria of the genus

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 6/28
6
Rhizobium. The seat of the symbiosis is within the nodules that appear on the plant roots.
Legumes, the most important plant group concerned in symbiotic N2 fixation, are
dicotyledonous plants of the family Lesuminosae, having species such as Trifolium,
Melilotus, Medicago, Lotus, Phaseolus, Dalea, Crotalaria, Vicia, Vigna, Pisum and Lathyrus.
Rhozibia grow readily in culture media containing a carbon source such as mannitol or
glucose, ammonium or nitrate to supply the required nitrogen and several inorganic salts. None of the bacteria in culture solution utilize N2; the fixation reaction is thus the result of a
true symbiosis as neither symbiont can carry out the process alone. Of particular importance
to the development of the symbiotic relationship is the presence of a large population of
rhizobia.
Symbiosis is the living together in close physical association of two or more different
organisms. There are three types of symbiotic relationships:
Commensalism: ( Latin com, together and mensa table) is a relationship in which one
symbiont, the commensal, benefits while the other (sometimes called the host) is neither
harmed or helped. Often both the host and the commensal “eat at the same table”. The spatial proximity of the two partners permits the commensal to feed on substances captured or
ingested by the host. The commensal also obtains shelter by living either on or in the host.
The commensal is not directly dependent on the host metabolically and causes it no particular
harm. When the commensal is separated from its host experimentally, it can survive without
being provided some factor or factors of host origin e.g. the common, nonpathogenic strain of
E. coli lives in the human clone. E. coli flourishes in the colon, but also grows quite well
outside the host and this is a typical commensal. These relationships can be very complex.
When O2 is used up by the facultative anaerobic E. coli, obligate anerobes such as Bacteroids
are able to grow in the colon. The anaerobes benefit from their association with the host and
E. coli but E. coli derives no obvious benefit from the anaerobes. In this case, the commensal
E. coli contributes to the welfare of other symbionts.
Mutualism: ( Latin mutus, borrowed or reciprocal) defines the relationship in which some
reciprocal benefit accrues to both partners. In this relationship the mutualist and the host are
metabolically dependent on each other.
Symbiosis involves intimate interactions based on mutual benefit, which is a good definition
of mutualism. Under nitrogen-limiting conditions legumes nodulated with active N2-fixing
strains of Rhizobium benefit from the interaction and the growth of the legume plants
stimulates the growth of rhizobia and other microorganisms in the soil. However, strains of
Rhizobium exist that fix N2 inefficiently, or not at all and such strains are either of little benefit to plant growth or are detrimental because they are utilizing the plant’s energy
without providing reciprocal benefit. The interaction between rhizobia and leguminous plants
has been studied in great detail for many years. Rhizobia are able to nodulate only a small
proportion of the very large number of species in the family Leguminosae and one non
legume, Parasponia. Within this range of host plants, specificity in the ability of particular
plant and Rhizobium species to form effective symbioses is observed e.g. R. trifolii nodulates
clovers (Trifolium spp.) which in turn are not usually nodulated by other rhizobia.
Symbioses involving Rhizobium are only one example of the type of interaction between
plants and N2 fixing microorganisms than can occur. The actinomycetes Frankia nodulates a
range of dicotyledonous plants, as does the cyanobacterium Nostoc on the cycad Macrozamia. In each case there is a degree of specificity, which implies that there are mutual
recognition systems. The range of plants with which cyanobacteria can form a symbiosis is

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 7/28
7
very wide indeed. It encompasses diatoms, fungi, mosses, liverworts, ferns, cycads and
angiosperms.
The proportion of heterocysts to vegetative cells is much higher in the symbiotic form than in
the free-living cyanobacteria. Studies on the differentiation of heterocysts in the filaments has
shown that this is determined by the nitrogen status. N2 is fixed by the heterocysts and diffuses into the vegetative cells. In the symbiosis where the products of N2 fixation are
excreted for use by the host, combined nitrogenlevels along the filament become depleted
sooner than in free-fixing forms, thus resulting in a greater proportion of heterocysts in
symbiotic systems. As with other symbioses, there are morphological adaptations of the host
and also physiological adaptations to cater for the special demands of N2 fixation.
Lichens are symbioses of fungi, ascomycetes and basidiomycetes with algae. By far the
highest proportion of lichen species are associations of fungi with green algae, but about 25
genera have cyanobacteria as the ‘algal’ symbiont. In the lichens, Azolla and most cycad
systems, the cyanobacteria exist outside the host’s cells, but in one species of cycad
Macrozamia communis, the cyanobacteria have been found inside the cells. This is similar tothe symbiosis with Gunnera. The cyanobacterial symbiont is Nostoc, and this symbiosis is of
interest as it is the only known cyanobacterial association with an angiosperm. In Azolla the
cyanobacteria ( Anabaena) exist in pockets within the leaf, and in Gunnera, Nostoc is
contained in glands at the base of leaves. Both are examples of containment that has resulted
from the development of a complex interaction. The cyanobacterial symbioses have a wide
range, and from a consideration of some of the associations it can be seen that there is a great
diversity of interactions, ranging from the lichens where the cyanobacterial partner
photosynthesizes and provides the host with both carbon and nitrogen, through the
association with Azolla where photosynthesis takes place but carbon is supplemented by the
host, to the situation in cycads and in Gunnera where the endophyte is dependent wholly
upon the host for carbon. N2 has to be given up to the host if the symbiosis is to be
mutualistic and the high proportion of heterocysts in all these associations show that it is.
Mycorrhizae represent particularly interesting plant-microbe interactions because they are so
wide spread and grown on lettuce can infect maize, grasses, beans, citrus and almost any
other plant species that can form mycorrhizal associations of the VA type. Mycorrhizae are
symbiotic because the plant provides the fungus with organic nutrients and in return fungus
facilitates the uptake of mineral nutrients and in particular phosphate. Mycorrhizal fungi can
be so important that some species of host plant are almost dependent upon them to be able to
grow in soils low in PO4; citrus and cassava are particularly good examples. Mycorrhizae are
thus something of an enigma. They are wide spread and show little sign of specificity, yetthey involve complex interactions, and possibly most plants are partially dependent upon
them for their PO4 nutrition. These symbioses have evolved to enable the fungus to invade
roots and routinely colonize from 10 to 90% of the root length with no obvious harmful effect
on the plant.
In soil, many microorganisms occur in close proximity and they interact in a unique way that
is in marked contrast to the behaviour of pure cultures studied by the microbiologists in the
laboratory. Members of the microflora rely upon one another for certain growth substances,
but at the same time they exert detrimental influences so that both beneficial and harmful
effects are evident.
The three beneficial relationships, symbiosis, protocooperation and commensalism are found
to operate among the soil inhibitor. Microorganisms in time develop certain relations that are

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 8/28
8
beneficial and others that are detrimental. Sometimes the benefit is mutual, but commensal
relationships are quite frequent. One of the more important beneficial associations is that
involving two species, one of which can attack a substrate not available to the second
organism, but the decomposition results in the formation of products utilized by the second.
This type of commensalism is not infrequent in nature and it is the way many polysaccharides
are transformed to nutrients supporting non-specialized microorganisms; e.g. cellulolyticfungi produce from cellulose a number of organic acids that serve as carbon sources for non-
cellulolytic bacteria and fungi. A second beneficial association arises from the need of many
microorganisms for accessory growth substances. These growth factors are synthesized by
certain microorganisms, and their excretion permits the proliferation of nutritionally complex
soil inhabitants. The microbial decomposition of biologically produced inhibitors that prevent
the proliferation of sensitive species is another instance of a beneficial relationship. Aerobes
may permit the growth of obligate anaerobes by consuming the O2 in the environment. In
addition to these instances of commensalism and protocooperation, several well documented
example of true symbiosis are in evidence, particularly those concerned with N2 fixation.
Competition
Microbial Competition
The categories of deleterious interactions are summarized by the terms competition,
amensalism, parasitism and predation, i.e. (a) the rivalry for limiting nutrients, (b) the release
by one species of products toxic to its neighbours and (c) the direct feeding of one organism
upon a second. Because the supply of nutrients in soil is perennially inadequate, competition
for carbon, minerals or oxygen is quite common. Alteration of the environment to the
detriment of certain microbial species may occur through the synthesis of metabolic products
that are bacterostatic or bactericidal by the utilization of oxygen which leads to the
suppression of obligate aerobes, or by the autotrophic formation of nitric and sulphuric acid
which affects the proliferation of acid-sensitive microorganisms. Predatory and parasitic
activities likewise are not rare. Predation and parasitism are observed in the feeding upon
bacteria by protozoa and myxobacteria, the attack on nematodes by predacious fungi, the
digestion of fungal hyphae by bacteria and the lysis of bacteria and actinomycetes by
bacteriophages. In mixed cultures of several microorganisms in laboratory media or in
partially sterilized soil, some species are suppressed while others survive, multiply and
assume dominance. The usual cause of this phenomenon is the competition for nutrients,
space or oxygen. In competition, certain microorganisms dominate through their capacity to
make most effective use of the limiting factors in the environment. Therefore, when large
populations of alien bacteria are added to soil, the invaders do not establish and soon die out.The habitat is foreign and the invader fails to find a niche. The disappearance itself is
probably the result of competitive effects since specific toxic substances active against the
alien bacteria are difficult to demonstrate. Microbiological competition for available carbon,
however, is probably one of the more important interactions between organisms. It is likely
that the role of an element in modifying the biological equilibrium is determined by the
demand of the microflora and the supply in the soil. As a first approximation, the ability of an
organism to compete is probably governed by its capacity to utilize the carbonaceous
substrates found in soil, its growth rate, and its nutritional complexity. A simple nutrition
could be advantageous, but the presence in soil of growth factors suggests the effective
competitors need not be nutritionally independent, as they can develop at the expense of
growth factors obtained from the environment.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 9/28
9
Amensalism
It is the suppression of the growth of one organism by the products of growth of a second
organism. This may result from a situation as simple as the alteration of the soil pH or the
production of the growth-inhibiting or lethal biological product. Thiobacillus spp. commonly
reduces soil pH through the oxidation of sulphide to sulfate. Because the pH may reach
values as low as 2, the growth of any pH sensitive microbes is inhibited. Two major types of
biological inhibitors or toxins are produced by soil microbes: those effective at high
concentrations (organic acids, chelators) and those that are effective at low concentrations
(antibiotics). The growth-controlling impact of the former compounds in soil has been
reasonably well accepted, because the substances can be quantified easily in soil samples and
their interactions with soil microbial populations can be shown. The role of antibiotics within
the soil ecosystem is more problematic. Another group of biologically synthesized
compounds that appear to be useful in reducing plant disease through antagonism of
pathogens are siderophores. These substances appear to be active at higher concentrations
than is characteristic of antibiotics, but when they result in suppression of microbial growth at
low concentrations, they can be classified as antibiotics. Siderophores are extra cellular, low-molecular weight (500 to 1000 dalltons) iron-transporting compounds synthesized by a
variety of microorganisms growing under low iron conditions. These substances selectively
complex ferric ion with a high affinity, thereby reducing iron availability to competing
organisms. Most commonly studied siderophore-synthesizing microbes from the view of
controlling plant pathogens are members of the fluorescent pseudomonad group.
Parasitism and Predation
Predators and parasites, organisms that feed upon living biomass, play a key role in the soil
ecosystem. Parasites and predators maintain the soil bacterial and fungal populations in an
active state and enhance nutrient movement between soil reservoirs through consumption of
microbial biomass. The feeding activity of predators and infectivity of parasites maintains a
younger, more active, soil microbial population. Essentially all types of predators or parasites
are present in the soil ecosystems. Bacteria, which prey on other bacteria, bolellovibrios,
bacteriophages, protozoa, as well as nematodes are all active in soil ecosystems. These
organisms may ingest their nutrients by consuming intact cells (holozoic feeding), as is
commonly described for protozoa, or extra cellular enzymes that lyse other bacteria, fungi or
algae may be produced. In the predator prey relationship between protozoa and bacteria, a
change in either group will bring about a qualitative and quantitative change in the other. The
presence of a nutrient supply in the form of bacteria is essential for the development of soil
protozoa and large numbers of bacteria must be ingested for one protozoan cell division. Inwell-mannered fields, the daily increases in bacteria and protozoa seem to be inversely
related, one group increases as the other decreases. Protozoa, therefore are undoubtedly a key
factor in limiting the size of the bacterial population, probably reducing the abundance of
edible cells and serving as a biological antagonist in maintaining the equilibrium.
Myxobacteria and myxomycetes also affect the true bacteria by feeding directly upon them.
Fungi are capable of parasitizing one another, and the parasitized species is thereby often
eliminated. The parasitism may entail a penetration into the host’s mycelium or a coiling
around the host’s hyphae. The virulence of individual fungi varies greatly even in a single
species. Certain fungi are predacious, capturing and consuming nematodes or amoebae, and
the study of the nematode-trapping fungi may prove of practical value in the control of plant
diseases caused by nematodes. A key consideration in evaluating predator or parasite behaviour in any ecosystem parasitic relates to the observation that both the host and
parasites or prey and predators coexist in the same ecosystem.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 10/28
10
Rumen Microbiology
Ruminants are the herbivorous mammals. Their digestive system includes four
compartments, the large pouch, small honey comb like, Omasum and abomasums (the
stomach). Domestic animals such as cows, sheep, goat, buffalo, camel etc. and the wild
animals such as deer, giraffe are the ruminants. Rumen is a special organ within which the
digestion of cellulose and other plant polysaccharides occurs through the activity of special
microbial populations. Mammals lack the enzymes needed to digest cellulose, so the rumen
containing a large number of microorganisms (mainly bacteria and protozoa) which play an
essential role in ruminant nutrition, break down plant material ingested by the host animal
and provide the animal with protein, vitamins and assimilable carbon and energy yielding
substrates. The rumen has a large size (100-150 litres in a cow, 6 litres in a sheep) and its
position in the alimentary tract as the organ where ingested food goes first. The rumen
contents are anaerobic, pH varies with diet but generally it is between 6-6.5. The rumen
temperature is about 39-40°C in the cow due to the (exothermic) microbial fermentation. The
reduction potential in rumen is –30 mV.
Food enters the rumen mixed with saliva containing bicarbonate and is churned in a rotary
motion during which the microbial fermentation occurs. This peristallic action grinds the
cellulose into a fine suspension, which assists in microbial attachment. The food mass then
passes gradually into the reticulum where it is formed into small clumps called cuds, which
are regurgitated into the mouth where they are chewed again. Now finely divided solids, well
mixed with saliva, are swallowed again, but this time the material passes to the omasum,
finally ending in the abomasum, an organ more like a true (acidic) stomach. Here chemical
digestive processes begin that continue in the small and large intestine. (In the suckling
animal, the rumen and reticulum are not fully developed and ingested food passes from the
oesophagus via the oesophageal groove to the omasum and abomasum – thus by passing the
rumen).
The number and type of microorganisms depend upon the nature of animal’s diet and on the
period of time since the last intake of food; the rumen contents contain approximately 1010
cells bacteria ml-1 rumen fluid. Food remains in the rumen for about 9-12 hrs. During this
period, cellulolytic bacteria and protozoa hydrolyze cellulose to the disaccharide cellobiose
and to free glucose units. Released glucose then undergoes bacterial fermentation with the
production of volatile fatty acids (VFAs), primarily acetic, propionic and butyric and the
gases CO2 and methane. The host animal absorbs the fatty acids from the rumen and from the
omasum and abomasums and eliminates the gases by erutation (the ruminant uses fatty acids
rather than glucose as primary sources of energy and carbon). The acidity of the fermentation
products is counteracted by the buffering action of the ruminant’s saliva – which is produced in copious amounts and contains sodium bicarbonate and sodium hydrogen phosphate. Many
microbial cells formed in the rumen are digested in the gastrointestinal tract and serve as a
major source of proteins and vitamins for the animal. Since many of the microorganisms of
the rumen are able to grow on urea as a sole nitrogen source, it is often supplied in cattle feed
in order to promote microbial protein synthesis. The bulk of this protein ends up in the animal
itself. A ruminant is thus nutritionally superior to a non-ruminant when subsisting on foods
that are deficient in protein, such as gases (Fig. 2a & b).
Rumen bacteria
Biochemical reactions taking place in rumen are complex and hence involve a large number
of microorganisms where anaerobic bacteria dominate. Several different bacteria hydrolyze

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 11/28
11
cellulose to sugars and ferment sugars to VFAs. Fibrobacter succinogens and Ruminococcus
albus are cellulolytic anaerobes. If a ruminant is gradually switched from cellulose to a diet
of high in starch (grains), then starch digesting bacteria Ruminobacter amylophilus and
Succinomonas amylolytica develop. If an animal is fed legume hay, which is high in pectin,
then pectin digesting bacterium Lachnospira multiparus is in the rumen flora. In fermentation
process, succinate is converted to propionate and CO2 and lactate is fermented to acetic and other acids by Selenomonas and Megaphaera. A number of rumen bacteria produce ethanol
which is fermented to acetate + H2. H2 quickly reduces CO2 to CH4 by methanogens. In the
rumen 65% CO2 and 35% CH4 are present and which leave the rumen by belching. In
addition to prokaryotes, rumen also has protozoal fauna (about 106/ml) which are obligate
anaerobes as well as anaerobic fungi that ferment cellulose to VFAs. Rumen fungi also
degrade plant polysaccharides as well as partially degrades lignin, hemicellulose and pectins.
(a)
(b)Fig.2: (a) Schematic diagram of the rumen and gastrointestinal system of a cow.
(b) Biochemical reactions in the rumen. (Biology of Microorganisms)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 12/28
12
Sometimes changes in the microbial composition of the rumen is fatal i.e. death of the animal
e.g. if a cow is changed abruptly from forage to grain diet, an explosive growth of
Streptococcus bovis from normal growth of 107 cell/ml to 1010 cells/ml takes place. Grains
contain high level of starch whereas grasses contain cellulose. S. bovis is a lactic acid
bacterium, ferments starch to lactate which acidifies the rumen which is called acidosis,killing off normal rumen flora. Such carbohydrates lead to a proliferation of acid producing
bacterium which cause a fall in pH and consequent loss of protozoans and many species of
bacteria. As a result, acidophilic Lactobacillus spp. predominate and cause a further fall in
pH.
Other animals
Buffalo, deer, reindeer, caribon and elk are also ruminants. Beleen whales also have a rumen
like fermentation. They contain multichambered stomach whose fore stomach is similar to
rumen and show abundant volatile fatty acids similar to cattle.
Biofertilizers
Biofertilizers can be defined as a microbial preparation containing N2 fixing or PO4-
solubilizing or celluoylic or such other useful microorganisms which by virtue of special
biochemical processes can increase the availability of a certain important nutrients in the
vicinity of the root system leading to better plant growth and crop productivity. Biofertilizers
are also called microbial inoculants. They are products containing living microorganisms
which have the ability to mobilize nutritionally important elements from nonusable to usable
form through biological process. Normally, the microorganisms are evolved after intensive
researches and are included in certain carriers such as charcoal, lignite or peat. Biofertilizers
have an important role to play in improving the nutrient supplies to crop plants as well as
trees in Indian agriculture as an alternate source of soil fertility building through renewable
sources. These can help in increasing the biologically fixed atmospheric N or increase the
native P availability to crop plants. Among the biofertilizers useful in increasing N supply, N2
fixing bacteria Rhizobium, Azotobacter, Azospirillum, blue green algae and Azolla are
important. Among those associated with increased P availability, different P solubilizing
bacteria and mycorrhizae are of significance. Thus, industrial production of biofertilizers has
come to help farmers to economize on chemical fertilizer inputs.
Biofertilizers can be defined as preparations containing live efficient microbes performing
various functions like nitrogen fixation, phosphorus solubilization or mobilization. They arecheap, economical sources of nutrients and are ecofriendly i.e. nonpolluting in nature.
Biofertilizers are known to make a number of positive contributions in soil thereby improving
soil health in general and crop health in particular by various mechanisms as under:
a) They fix atmospheric nitrogen and provide to various crops.
b) They produce and liberate various plant growth promoting substances, vitamins and
help in better and quick growth.
c) They produce certain antimicrobial agents and suppress the incidence of pathogens.
d) They solubilize or mobilize phosphorus in soil.
e) They improve soil physical, chemical and biological health of soil.
Following are the various groups of biofertilizers for different crops which are being popularly used:

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 13/28
13
Nitrogen fixing biofertilizers
1. Rhizobial Biofertilizers: Fix atmospheric nitrogen in symbiosis with leguminous
crops. Bacteria provide nitrogen to plants and plants in turn provide carbon sources
and other nutrients to bacteria e.g. Rhizobium.
2. Azotobacter Biofertilizers: Fix atmospheric nitrogen a symbiotically in soil. Produce
unspecified plant growth promoting substances thereby induce profuse root and shoot
growth. They also produce certain antimicrobial agents which keep away pathogens.
Azotobacter biofertilizers are used for non leguminous crop like cereals millets and oil
seed.
3. Azospirillum Biofertilizers: Azospirillum form a loose symbiosis with
nonleguminous crops and are known as associative symbionts. These bacteria are
benefited by root exudates of plants and help the plants by fixing atmospheric
nitrogen and producing plant growth promoting substances.
4. Cyanobacterial Biofertilizers or BGA Biofertilizers: These are useful in rice
fields. They fix atmospheric nitrogen and produce plant growth promoting substances
and Vitamins.5. Phosphate solubilizing biofertilizer: Many times phosphate becomes a imiting
factor for plant growth because much of it in the soil is bound as insoluble calcium,
iron or aluminium phosphates. The availability of phosphates therefore depends on
the degree of solubilization of insoluble phosphates by various organic and iorganic
acids produced by the microorganisms thereby solubilize insoluble phosphates and
make it available to the plant. Bacillus mregaterium, B. polymyxa, Predomonas,
Aspergillus, Mycorrhiza are commonly used phosphate solubilizing biofertilizers.
6. Plant growth promoting rhizobacteria (PGPR): PGPR are also being used a
biofertilizer as they are able to produce various phytohormones like, IAA, Cytokinin
and gibberellins etc which are important for plant growth and productivity. Popularly
used PGPR are Pseudomonas, Bacillus, Agrobacterium, Cellomonas, Arthrobacter, Alcaligenes, Actinoplane
Biological N 2 fixation
The utilization of atmospheric N2 gas as a source of nitrogen is called nitrogen fixation.
Prokaryotes both anaerobic and aerobic fix N2. No eukaryotic organisms fix N2. There are
some bacteria called symbiotic fix N2 only in association with certain plants. Biological N2
fixation is brought about by free-living bacteria or blue-green algae, which make use of N2 by
non-symbiotic means and by symbiotic associations composed of a microorganisms and a
higher plant. N2 fixation, the reduction of N2 to NH3 involves a complex enzyme systemcalled nitrogenase, which consists of dinitrogenase and dinitrogenase reductase, metal-
containing enzyme found only in certain prokaryotic organisms. Most nitrogenase contain
molybdenum or vanadium and iron as metal cofactors and the process of N2 fixation is highly
energy-demanding. Nitrogenase and associated regulatory proteins are encoded by the nif
regulation. Certain artificial substrates that are structurally similar to N2, such as acetylene
and cyanide are also reduced by nitrogenase (Fig. 3).
Nitrogenase is a functional enzyme which reduces N2 to ammonia and depends on energy
source from ATP. The nitrogenase has two components: one containing Mo-Fe, designated as
Mo-Fe protein (nitrogenase) and the other iron protein (nitrogenase reductase). Both the
components are essential for nitrogenase activity.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 14/28
14
ATP
N2 + 3H2 Æ 2 NH3
For this reaction, for every two electron transfer by nitrogenase, four ATP moles are required.
Fig. 3: Pathway of nitrogen fixation (Agricultural Microbiology)
Nitrogenase enzyme
The reduction of N2 to ammonia is catalysed by the enzyme nitrogenase. Reaction has
a high activation energy because molecular N2 is an unreactive gas with a triple bond
between the two N2 atoms. Therefore, N2 reduction requires at least 8 electrons and 16 ATP
moles, 4 ATPs per pair of electrons.
N2 + 8H+ + 8e- + 16 ATP Æ 2NH3 + H2 + 16 ADP + 16 P;
The electrons come from ferredoxin that has been reduced in a variety of ways:
i) by photosynthesis in cyanobacteria.
ii) Respiratory processes in aerobic N2 fixers,
iii) Fermentations in anaerobic bacteria
e.g. Clostridium pasteurianum (an anaerobic bacterium) reduces ferredoxin during pyruvate
oxidation, whereas the aerobic Azotobacter uses electrons from NADPH to reduce
ferredoxin.
Nitrogenase is a complex system consisting of two major protein components a MoFe protein
joined with one or two Fe proteins. The MoFe protein contains 2 atoms of molybdenum and
28 to 32 atoms of iron; the Fe protein has 4 iron atoms. Fe protein is first reduced byferredoxin, then it binds ATP. ATP binding changes the conformation of the Fe protein and
lowers its reduction potential, enabling it to reduce the MoFe protein. ATP is hydrolyzed

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 15/28
15
when this electron transfer occurs. Finally, reduced MOFe protein donates electrons to atomic
nitrogen. Nitrogenase is quite sensitive to O2 and must be protected from O2 inactivation
within the cell. The reduction of N2 to NH3 occurs in 3 steps, each of which requires an
electron pair. Six electron transfers take place and this requires a total 12 ATPs per N2
reduced. The overall process actually requires at least 8 electrons and 16 ATPs because
nitrogenase also reduces protons to H2. The H2 reacts with diimine (HN=NH) to form N2 and H2 (Fig 4).
Fig. 4: Mechanism of Nitrogenase Action (Prescott- Harley-Klein)
Nitrogenase can reduce a variety of molecules containing triple bonds (e.g. acetylene,
cyanide and azide)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 16/28
16
HC = CH + 2H+ + 2e- Æ H2C = CH2
The rate of reduction of acetylene to ethylene is even used to estimate nitrogenase activity.
Once molecular N2 has been reduced to ammonia, the ammonia can be incorporated into
organic compounds.
The nif genes
The genes for nitrogen fixation, called nif genes are found in both symbiotic and free living
systems, but in the Rhizobium-legume system, the nif genes are distinct. The symbiotic
activation of nif -genes in the Rhizobium is dependent on low oxygen concentration, which in
turn is regulated by another set of genes called fix-genes which are common for both
symbiotic and free living nitrogen fixation systems.
The nif -genes have been investigated most thoroughly in Klebsiella. Work with Klebsiella pneumoniae has shown that there are 17 nif -genes. Proteins have been identified and in some
cases functions for these genes are known. If one gene codes for the synthesis of one
polypeptide, then several genes will be necessary to code for the nitrogen-fixing system. The
Fe protein is composed of two sub-units but, as each of these is the same, one gene will code
for this protein. The MoFe protein has two different sub-units, each of which will require one
gene. The molybdenum cofactor will require a gene and further genes will be necessary to
code for any special electron donors in the system. The genes that code for these proteins are
all adjacent to one another on the Klebsiella chromosomes.
Functions of the nif genes of Klebsiella pneumonia
Gene Function of the gene or gene product
nif H Codes fore the sub-unit of the Fe protein
nif D Codes for the sub-unit of the FeMo protein
nif K Codes for the ß-sub-unit of the FeMo protein
nif M Activation of the Fe protein
nif B Involved in the synthesis and insertion of the iron
molybdenum cofactor, FeMoCo
nif N As for B
nif E As for B
nif V As for B
nif F Codes for a flavodoxin
nif J Codes for a pyruvate: flavodoxin oxidoreductase
nif A Codes for an activator mole for the other nif genes
nif L Codes for a repressor molecule for the other nif genes
nif Q Possibly concerned with molybdenum uptake
nif S Possibly concerned with processing the FeMo protein
nif U As S
nif X Unknown
nif Y Unknown

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 17/28
17
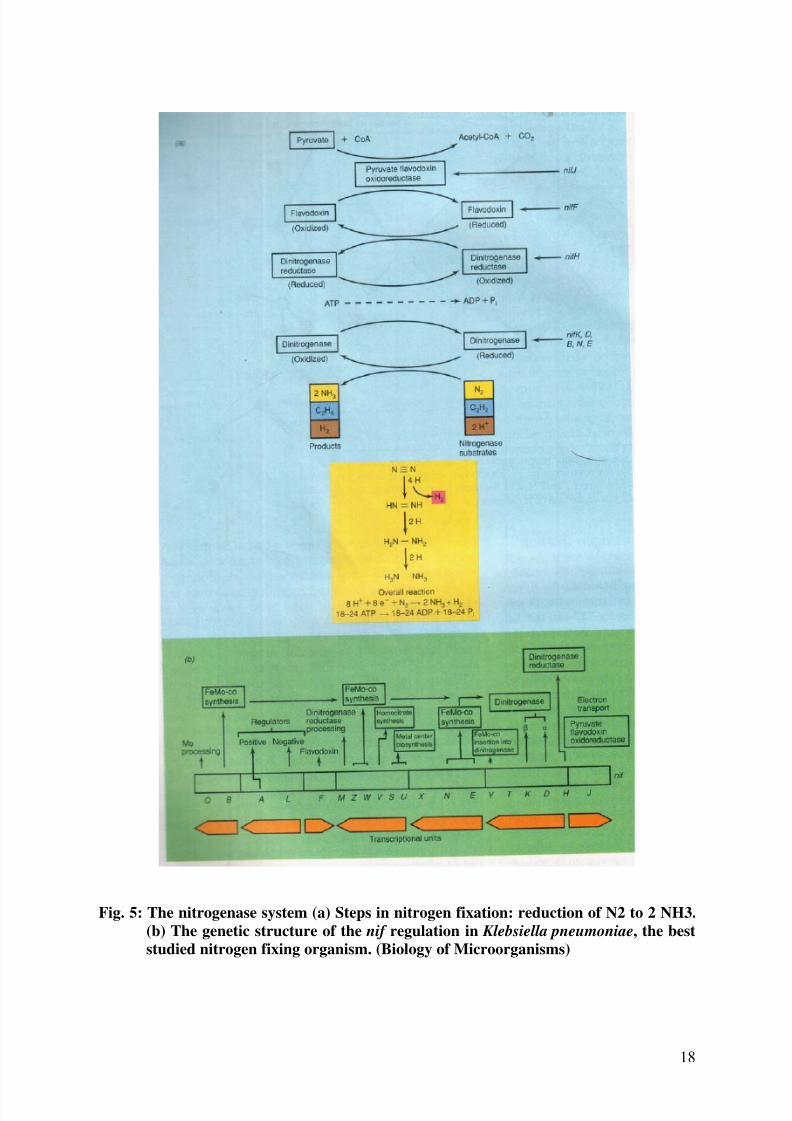
The nif H, D and K genes code for the polypeptides of the Fe and MoFe proteins. These
genes are readily identified by the lack of, or alteration of the particular proteins in mutants.
Mutations in the nif M gene results in an inactive Fe protein, so that the product of the nif M
gene must be involved in modifying the protein in some way, perhaps incorporating the Fe-Scluster. Similarly, mutations in several genes affect the activity of the MoFe protein.
Mutations of nif V give an altered substrate specificity. These mutants are unable to reduce
N2 but can reduce acetylene. Carbon monoxide, which does not inhibit hydrogen evaluation
from normal nitrogenase, inhibits hydrogen evaluation from nif V mutants.
When FeMoCo was obtained from the nif V mutant protein and was combined with protein
of a nif B mutant, a protein from which FeMoCo is absent, the nif V-phenotype was obtained.
However, when FeMoCo from a normal protein was added to the nif B mutant protein a
normal protein resulted. Thus it was concluded that the nif V product modifies FeMoCo in
order to produce effective nitrogenase. From studies of nif V- mutants it has been concluded that FeMoCo contains the binding site for N2 and CO.
Three other genes, nif B, N and E have been identified with the synthesis of FeMoCo and the
nif Q product’s action is thought to be the acquisition of molybdenum. Thus five of the genes
nif Q, B, N, E and V are connected with the synthesis of the molybdenum cofactor. The
products of the genes nif S and U are thought to modify the MoFe protein, although there is
no hard evidence for this as yet. If this is true, then nine genes are needed to produce the
complete active MoFe protein. Two genes are concerned with electron transport to
nitrogenase: nif F and J. Extracts of mutants of both of these genes can fix nitrogen if they are
provided with the artificial electron donor, sodium dithionite. Extracts of nif F- mutants can
be rendered active by providing Azotobacter flavodoxin. It is thus assumed that the product
of nif F is a flavodoxin. The nif J product has been shown to be the enzyme pyruvate :
flavodoxin oxidoreductase, which catalyses the oxidation of pyruvate to produce reduced
flavodoxin:
Pyruvate + CoA + flavodoxin ----- acetyl CoA + flavodoxin + CO2
ox red
With flavodoxin and the pure enzyme in the reaction mixture, the reduction of nitrogenase
and then acetylene can be achieved, when the flavodoxins from Azotobacter is used, the
activity is one third of that with the flavodoxin from Klebsiella, which demonstrates thatthere is some specificity for the reductant and that flavodoxins from different species may
differ.
The genes which control the synthesis of the nitrogenase proteins will be present in all the
species that fix nitrogen. However, the genes concerned with electron transport will differ, as
the provision of electrons depends upon the metabolism of pyruvate. It is interesting to note
that the genes nif F and J for electron transport to nitrogenase are transcribed in the opposite
direction to the other nif genes.
The genes nif X and nif Y have been identified by means of their polypeptide products from
cloned fragments of the nif region. The function of these genes has however not beenestablished. The remaining two genes are nif A and nif L. The purpose of these genes is to
control the expression of the other genes in the nif region (Fig. 5a & 5b).
7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 18/28
18
Fig. 5: The nitrogenase system (a) Steps in nitrogen fixation: reduction of N2 to 2 NH3.
(b) The genetic structure of the nif regulation in Klebsiella pneumoniae, the best
studied nitrogen fixing organism. (Biology of Microorganisms)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 19/28
19
Symbiotic N2 fixation
Two members are required for the association, a plant and a microorganism. The classical
example of such a symbiosis is that between leguminous plants and bacteria of the genus
Rhizobium. The seat of the symbiosis is within the nodules that appear on the plant roots.
Legumes, the most important plant group concerned in symbiotic N2 fixation, are
dicotyledonous plants of the family Leguminosae. For successful symbiotic N2 fixation, a
healthy plant growing in sufficient light and an effective nodule forming bacterium are
required. The nodule fixes N2 only for a short duration when it is in the highest symbiotic
relationship with the plant. The energy requirement for biological N2 fixation appears to be
high and this becomes the limiting factor in the quantity of N2 fixed in different legume-
Rhizobium combinations. Fifty percent of natural nitrogen fixation is accomplished by the
Rhizobium legume association. Different symbiotic associations are Rhizobium-legumes;
Rhizobium non legumes, frankia and Angiosperm and cyanobacterial associations.
RhizobiumThe rhizobia are soil organisms that inhabit the rhizosphere of legumes and other plants.
There are two main types of rhizobia, “fast growers” and “slow growers”. The division of
Rhizobium into species is based on the interaction with plants; those bacteria which nodulate
clovers, are put in R. trifolii and those that nodulate peas and vetches are put in R.
leguminosarum. The bacteria are Gram negative, non-spore forming, aerobic rods, 0.5 to 0.9
µ wide and 1.2 µ to 3.0 µ long. They are typically motile and utilize several carbohydrates,
sometimes with the accumulation of acid but never of gas. Rhizobium bacteria stimulate
leguminous plants to develop root nodules, which the bacteria infect and inhabit. The nodules
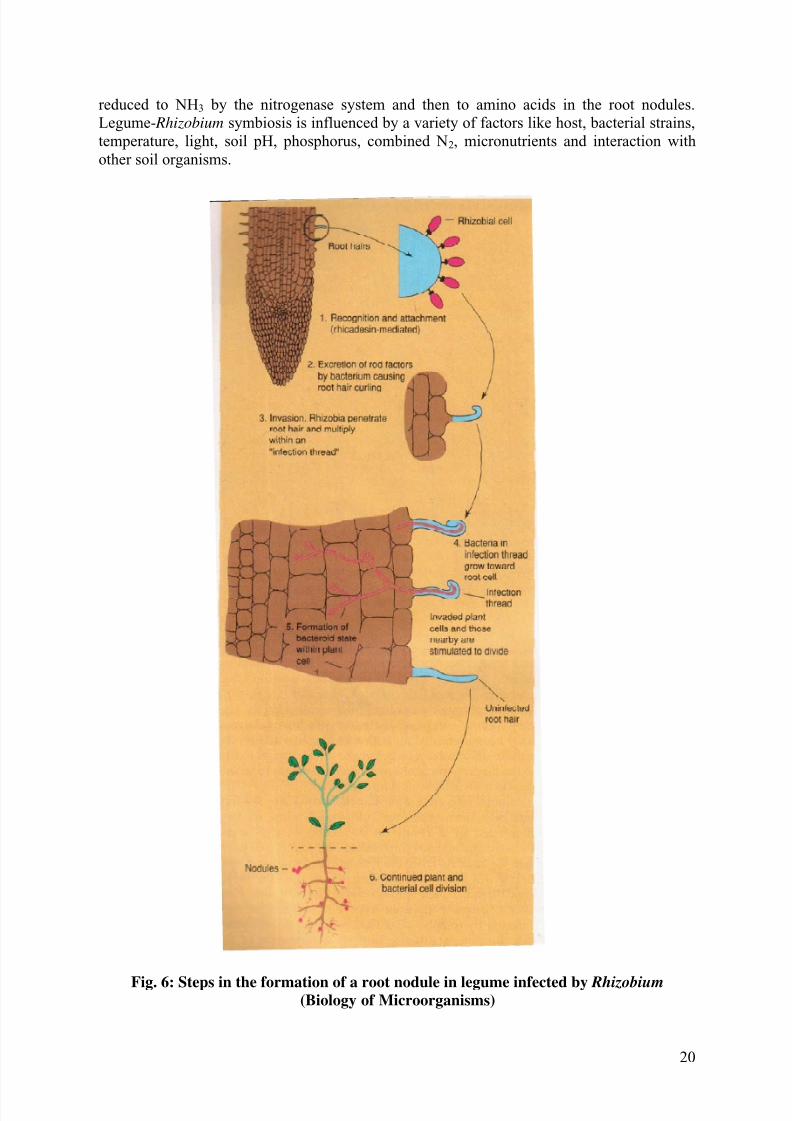
develop in a complex series of steps (Fig. 6).
(i) Recognition of the correct partner on the part of both plant and bacterium and
attachment of the bacterium to root hairs.
(ii) Invasion of the root hair by the bacterial formation of an infection thread.
(iii) Travel to the main root via the infection thread.
(iv) Formation of deformed bacterial cells, bacteroids, within the plant cells and
development of the nitrogen-fixing state.
(v) Continued plant and bacterial division and formation of the mature root nodule.
The roots of leguminous plants secrete a variety of organic materials that stimulate the
growth of a rhizosphere microflora. If there are rhizobia in the soil, they grow in the
rhizosphere and build up to high population densities. Attachment of bacterium to plant in thelegume – Rhizobium symbiosis is the first step in the formation of nodules. A specific
adhesion protein called rhicadhesin which is present on the surface of Rhizobium is a calcium
binding protein and binds calcium complexes on the root hair surface. Rhizobium cells
penetrate into the root hair via the root hair tip. Following binding, the root hair curls as a
result of the action of substances excreted by the bacterium called Nod factors and the
bacteria enter the root hair and induce formation by the plant of a cellulosic tube, called the
infection thread, which spreads down the root hair. Root cells adjacent to the root hairs
subsequently become infected by rhizobia and Nod factors stimulate plant cell division,
eventually leading to formation of the nodule. The bacteria multiply rapidly within the plant
cells and are transformed into swollen, misshapen and branch forms called bacteroids.
Bacteroids become surrounded simply or in small groups by portions of the plant cellmembrane. Only after the formation of bacteroids does nitrogen fixation begins. N2 fixation
takes place in these nodules and the effective nodules are pink in color. Atmospheric N2 gets
7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 20/28
20
reduced to NH3 by the nitrogenase system and then to amino acids in the root nodules.
Legume- Rhizobium symbiosis is influenced by a variety of factors like host, bacterial strains,
temperature, light, soil pH, phosphorus, combined N2, micronutrients and interaction with
other soil organisms.
Fig. 6: Steps in the formation of a root nodule in legume infected by Rhizobium (Biology of Microorganisms)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 21/28
21
Symbiotic process is controlled by a number of nod genes of which some are host specific
and the other are common nod genes. Some of the nod genes induce the host plant to react by
producing nodulins which are flavanoids. Some nod genes are required for root-hair curling
and for cell division. The genes for N2 fixation, called nif genes, are found in both symbiotic
and free living systems, but in the Rhizobium-legume system, the nif genes are distinct. The
plant supplies carbon compounds, derived from photosynthesis in the shoot, which helps the bacteria to produce ATP, which is required in large quantities for N2 fixation. The
leghaemoglobin (Lb) present in the nodule binds O2 so as to facilitate N2 fixation by the
bacterium, since presence of O2 inhibits nitrogenase enzyme of the bacterium. The bacteria in
the nodule undergo limited DNA replication and division and then transform into bacteroids.
The plant-derived peribacteroid membrane (PBM) forms the envelop for the bacteroids.
There is specific gene-controlled interaction taking place between the bacteroids and the
PBM.
The symbiotic activation of nif -genes in the Rhizobium is dependent on low O2 concentration,
which in turn is regulated by another set of genes called fix genes which are common for both
symbiotic and free-living N2 fixation systems. The Rhizobium bacterial cells and the hostcells cooperate intimately in respect of cellular metabolism with the required energy and
growth regulation, accompanied by the genetic transcription and translation through DNA.
The exchange of carbon sources with that of the nitrogenous substances is balanced as to
make the bacterium a symbiont instead of a pathogen i.e. the bacterium through infective,
should ultimately come under the control of the host (Fig. 6).
Frankia
The Alder tree (genus alnus) has N2-fixing root nodules that harbor a filamentous,
streptomycete like, N2 fixing organism called Frankia. The members of this genus areactinomycetes : most of these bacteria at sometime in their life cycle have a filamentous habit
which often superficially bears some morphological resemblance to the fungi. They are
however, prokaryotes with hyphae of smaller dimensions – in Frankia typically less than 2
µm diameter-than fungi.
An important feature of Frankia is that many strains can fix N2 at normal O2 concentration at
rates sufficient to support growth in culture. although when assayed in cell extracts the
nitrogenase of Frankia is sensitive to molecular O2, like intact cells of Azotobacter. Intact
cells of Frankia fix N2 at full O2 tensions. This is because Frankia protects its nitrogenase by
localizing it in terminal swellings on the cells called Vesicles. The vesicles contain thick
walls of laminated structure that act as a barrier to O2 diffusion, thus maintaining the O2 tension within vesicles at levels compatible with nitrogenase activity. Frankia vesicles
resemble the heterocysts produced by some filamentous cyanobacteria as localized sites of N2
fixation. N2 fixation in such cultures is inhibited by the addition of combined N2.
Like Alder, Frankia nodulates a number of other small woody plants. This root nodule
symbiosis has been reported in at least 8 families of plants, many of which show no
evolutionary relationships to one another. This suggests that the nodulation process in the
Frankia symbiosis is more of a generalized phenomenon than the highly specific process
observed in the Rhizobium-legume symbiosis and this holds promise for experimental
attempts to expand the Frankia symbiosis to agriculturally important plants (Fig. 7).

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 22/28
22
a b
Fig. 7(a&b): Frankia nodules and Frankia cells. (a) Root nodules of the common alder
Alnus glutiosa (b) Frankia culture purified from nodules of Componia pergrina. (Biology
of Microorganism)
Non-symbiotic N2 fixation
Biological N2 fixation is brought about by free-living bacteria or blue-green algae, which
make use of N2 by non-symbiotic means. A number of environmental factors govern the rate
and magnitude of non-symbiotic N2 fixation and the transformation is markedly affected by
the physical and chemical characteristics of their habitat:
• Microorganisms that assimilate N2 have the ability to utilize ammonium and sometimes
nitrate and other combined forms of nitrogen. In fact, ammonium salts are used
preferentially and often at a greater rate than molecular N2 so that the presence of
ammonium, ineffect, inhibits the fixation, i.e. the bacteria use the N2 salt rather than N2
from the atmosphere.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 23/28
23
• Many inorganic nutrients are necessary for the development of the microorganisms.
Molybdenum, calcium and iron are critical for the fixation reaction.
• Molybdenum is required for the metabolism of N2, but microorganisms do not use nitrate
unless molybdate is present although the molybdenum requirement for nitrate utilization
is less than for N2 fixation.
• In like manner, Fe salts are implicated in the N2 metabolism of Azotobacter, Clostridium, Algae, Aerobacter and Achromobacter , but the specific requirement for N2 metabolism is
often difficult to establish because Fe is required, to a lesser extent, for growth upon fixed
compounds of N2.
• A requirement for Ca has been demonstrated during N2 assimilation by blue-green algae
and some Azotobacter spp., but the need for calcium can sometimes be replaced by
strontium.
• Azotobacter is characteristically sensitive to high hydrogen ion concentrations. Their
absence is associated directly with pH. As a rule, environments more acid than pH 6.0 are
free of the organism or contain very few Azotobacter cells. Similarly, the bacteria
generally, will neither grow nor fix N2
in culture media having a pH below 6.0.
• Beijerinckia spp. do not possess the acid sensitivity like Azotobacters and they develop
and fix N2 from pH 3 to 9.
• Blue-green algae bacteria, however, develop poorly in media and are sparse in soils more
acid than approximately pH 6.0 whereas the acid tolerance of Clostridium falls between
Azotobacter and Bcijerinckia.
• There is some evidence that the occurrence of Azotobacter is also related to the available
PO4 content of soils. About 1 mg of phosphorus must be assimilated by Azotobacter for
each 5 to 10 mg of N2 fixed.
• The distribution of blue-green algae in wet paddy fields is likewise associated with the
PO4 content of the soil.
Azotobacter and Azospirillum
Azotobacter is a free living heterotrophic nitrogen fixing bacterium encountered in neutral to
alkaline soil conditions. The bacterium not only provides the nitrogen but produces a variety
of growth promoting substances. These include indole acetic acid (IAA), gibberellic acid
(GA), vitamin-B and anti fungal substances. Another important characteristic of Azotobacter
associated crop improvement is excretion of ammonia in the rhizosphere in the presence of
root exudates and help in modification of nutrient uptake by the plants. These strains are
better competitors than the non excreting strains. The genus Azotobacter is highly versatile in
utilizing carbon sources therefore, application of organic carbon containing sources to the soilimproves the asymbiotic N2 fixation capacity by the diazotroph. The benefits of Azotobacter
inoculation are: enhanced branching of roots, production of plant growth hormones,
enhancement of uptake of NO3, NH4+, H2PO4
-, K +, Rb++ and Fe++, improved water status of
plants, increased nitrate reductase activity and antifungal compounds.
Members of this genus are strict aerobes: O2 is required for metabolism and also to fix N2. N2
fixation therefore occurs in an aerobic environment and there must be a mechanism to
prevent the access of O2 to the O2-sensitive proteins. Azotobacter has a very high rate of
respiration and when the organism is deprived of respirable substrate, as when it is grown on
a medium low in carbon, the nitrogenase is more susceptible to O2. N2 fixation is inhibited
when the organism is suddenly exposed to an O2 concentration higher than that in which ithas been grown. Azotobacter therefore, grows better at O2 concentration lower than
atmospheric when fixing N2. Where there is sufficient substrate it is likely that the O2

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 24/28
24
concentration will be lower than that of atmospheric because of the respiration of both
Azotobacter and of other microorganisms in the vicinity.
The genus Azotobacter comprises large, Gram-negative, obligately aerobic rods capable of
fixing N2 non-symbiotically. The tropical soils have much higher Azotobacter populations
than those found under temperate climates. In soil, their numbers vary from a few to a fewhundred per gm of soil. Application of nitrogenous fertilizers drastically reduces the
Azotobacter population in soil. Inoculation of soil or seed with Azotobacter is effective in
increasing yields of crops in well manured soil with high organic matter content.
Azospirillum: an associative micro aerophilic N2 fixer, commonly found in association with
roots of cereals and grasses has received great interest as a biofertilizer. Its useful characters
include high N2 fixation capacity, low energy requirement and tolerance to high soil
temperature for its suitability under tropical conditions. Azospirillum is a mesophyllic
bacterium and is reported to occur in association with crops grown in acidic to alkaline pH
range. Azospirilla are metabolically versatile and can grow vigorously in the presence of
nitrogenous compounds present in the soil but as soon as the external N2 supply is exhausted the bacteria shift to diazotrophy. Use of Azospirillum inoculum under saline alkaline
conditions is possible because strains adapted to these stress conditions maintained high N2
activity. Crops grown to pre-treated seed give increased yields up to about 10 to 30%.
Azospirillum participates in all steps of the N2 cycle except nitrification. It can fix
atmospheric N2 in pure culture and under microaerophilic conditions. Azospirillum spp. have
been isolated from the rhizosphere of a large number of monocotyledons and a few
dicotyledon plants.
Azospirillum lipoferum has been observed to fix atmospheric N2 in the cortical cells of the
roots of maize. Substantial increases in yield were reported following the inoculation of
sorghum and pearl millet with Azospirillum brasilense under several agro-climatic conditions
in India. In addition to N2 fixation, hormonal effects have also been shown to be responsible
for at least part of yield increase following inoculation with Azospirillum. It has also been
shown that Azospirillum and Azotobacter , besides enhancing N2 uptake by plants, increase
the number of root hairs and root hormone exudation. This genus of spirally curved Gram
negative bacteria is interesting as its members not only live in the rhizosphere of grasses but
can also enter the root cortex. These organisms use root exudates for their carbon and energy
source while fixing N2. Azospirillum is a O2-sensitive and can fix N2 only at low O2
concentrations. It is a tropical bacterium and has a high optimum temperature so that it does
not occur to any great extents in temperature latitude. It has a wide host range.
Mycorrhizae
Mycorrhizae are fungus root associations, first discovered by Albert Bernhard Frank in 1885.
The word mycorrhizae comes from the Greek words meaning fungus and roots. These
microorganism contribute to plant functioning in natural environments, agriculture and
reclamation. The roots of 95% of all kinds of vascular plants are normally involved in
symbiotic associations with mycorrhizae (Fig 8).
Five types of mycorrhizae can be recognized:
(i) Ectomycorrhizae which form a sheath around roots but lack intracellular penetration of
the cortical cells; three types of Endomycorrhizae: (ii) ericoid, (iii) orchid and (iv)

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 25/28
25
vesicular-arbuscular mycorrhiza which colonize the root cortical cells intracellularly and
(v) Ectendomycorrhizae which form sheath and produce intracellular penetrations (Fig 9).
Fig. 8: Components of the mycorrhizal symbiosis. Phosphate enters the plant, alongwith other mineral nutrients, both directly from the soil and through the fungus.
(Advances in Agricultural Microbiology)
Ectomycorrhizae: are found mainly in forest-trees, especially conifers, beeches and oaks and
are most highly developed in temperate forests. In a forest, almost every root of every tree is
mycorrhizal. The root system of a mycorrhizal tree is composed of both long and short roots.
The short roots, which are characteristically dichotomously branched, show the typical fungal
sheath, whereas long roots are usually uninfected. Most mycorrhizal fungi do not attack
cellulose and leaf litter but instead use simple carbohydrates for growth and usually have one
or more vitamin requirements; they obtain their nutrients from root secretions. The
mycorrhizal fungi are never found in nature except in association with roots and hence can beconsidered obligate symbionts. These fungi produce plant growth substances that induce
morphological alterations in the roots, causing characteristically short dichotomously
branched mycorrhizal roots to be formed. Despite the close relationship between fungus and
root, there is little species specificity involved, a single species of pine can form mycorrhizae
with over 40 species of fungi. Ectomycorrhizal fungi penetrate intra cellularly and partially
replaces the middle lamellae cortical cells of feeder roots. These fungi form a dense mycelial
net around and between the plant cells termed Hartig net. Ectomycorrhizal associations are
also characterized by a dense, generally continuous hyphal network over the feeder root
surface called a fungal mantle. This fungal mantle varies from one to two hyphal diameters to
as many as 30 or 40 depending upon the fungal associate, the host and the environmental
conditions. Example of plant spp. forming ectomycorrhizal associations are spp. in thefamilies Pinaceae, Salicaceae, Betulaceae and Fugaceae. Most ectomycorrhizal fungi are

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 26/28
26
basidomycetes (primarily of the families Amanitaceae, Boletaceae, Cortinariaceae,
Russulaceae, Tricholomataceae, Rhizopogonaceae and Sclerodermataceae). Mycorrhizal
hyphae have been found to be very efficient in the uptake of phosphorus from the soil, which
would otherwise be unavailable to the plant. The ectomycorrhizal fungi help in the
phosphorus nutrition of plants through increased surface area of absorption, offer protection
against some of the soil-borne plant pathogens and enhance rooting and survival of cuttingsthrough production of growth hormones.
Endomycorrhizae: are distinguished by the fact that the fungus penetrates the cortical cells of
feeder roots and may form large vesicles and arbuscles (hence the term vesicular-arbuscular
mycorrhizae (VAM). These fungi do not form dense fungal mantles, but they do develop a
loose, intermittent arrangement of mycelium on the root surface. Endomycorrhizae are
formed by most agronomic, horticultural and ornamental crops, as well as some forest tree
spp. that do not form ectomycorrhizae. The fungal spp. are phycomyces many of which are in
the genus Endogone. VAM colonization originates from hyphae arising from soil borne
propagules. On reaching the cortex, hyphae grow into cells by tree like dichotomous
branching to give arbuscules. When colonization is well established, oval structures called vesicles may form which have storage functions. Vesicles appear to be organelles for the
storage of lipid and energy reserves and arbuscules, which resemble the haustoria of rust
fungi and mildews are complex ramifications of small branches of the fungus that provide
sites for nutrient exchange. VA mycorrhizal fungus grown on lettuce can infect maize,
grasses, beans, citrus and almost any other plant spp. that can form mycorrhizal associations
of the VA type. Ectomycorrhizal fungi have more limited host ranges and there are plant spp.
which are not infected by endomycorrhizal fungi.
Endomycorrhizae are of particular interest, as it has not been possible to grow these fungi,
usually members of the zygomycetes, without the plant. In this association the fungal hyphae
penetrate the outer cortical cells of the plant root, where they grow intracellularly and form
coils, swellings, or minute branches. Endotrophic mycorrhizae are found in wheat, corn,
beans, tomatoes, apples, oranges and many other commercial crops, as well as most pasture
and rangeland grasses. Recent studies show that plant flavonoids may stimulate spore
germination and this could lead to the development of plant-free cultures of these
mycorrhizae.
Mycorrhizal fungi have been observed to improve plant growth through better uptake of P
and Zn from soil. The VAM fungi penetrates the outermost cortex region, when the plant is
well supplied with phosphorus, but in phosphorus deficient plants they penetrate deep into the
cortex and help the plant to obtain the nutrient from the soil. Recent studies have shown thatthey stimulate beneficial organisms like Rhizobium, Azotobacter and phosphate solubilizers
in the rhizosphere and suppress the growth of root pathogenic fungi and nematodes. In
addition, the mycorrhizal fungi are reported to increase the availability of water to plants,
resulting in more vigorous growth under drought conditions.
The ericoid mycorrhiza is seen in members of heath family like blueberry and Erica.
Pezizella ericae, an ascomycete, is the most common fungal symbiont, which can be
cultured. Currently, several laboratories are studying this association, as blueberry is an
important cash crop.
All orchids are infected at some stage in their life cycle by the Orchidaceous mycorrhizalfungi. A recent study has shown that artificial inoculation of orchids with the mycorrhizal
fungus is not necessary as the fungus is abundantly present in nature.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 27/28
27
Ectodomycorrhizae: These mycorrhizae resemble ectomycorrhizae in forming a Hartig net
and a fungal mantle. A resemblance with the endomycorrhizae is associated with their
penetration of cortical cells. This mycorrhizal grouping is the least studied and the nature of
the fungal symbionts has not been totally elucidated.
A mycorrhizae is a mutualistic symbiosis between plant and fungus localized in a root or root-like structure in which energy moves primarily from plant to fungus and inorganic
resources move from fungus to plant. The formation of mycorrhizae is particularly
pronounced in land low in phosphorus and N2 and high nutrient levels are correlated with
poor mycorrhizal development. N2 fixing microorganisms can increase soil N2 while
mycorrhizal fungi effectively augment the absorbing surface of the roots. Many tropical soils
are so phosphorus deficient that they cannot respond to N2 until this deficiency is corrected. It
is here that endomycorrhiza takes on added significance, as it enhances the absorption of
phosphorus.
Mycorrhizae are symbiotic because the plant provides the fungus with organic nutrients and
the fungus facilitates the uptake of mineral nutrients and in particular PO4. Mycorrhizal fungican be so important that some species of host plant are almost dependent upon them to be
able to grow in soils low in PO4, citrus and cassava are particularly good example.
Mycorrhizae are thus something of an enigma. They are wide spread and show little sign of
specificity, yet they involve complex interactions and possibly most plants are partially
dependent upon them for their PO4 nutrition. These symbioses have evolved to enable the
fungus to invade roots and routinely colonize from 10% to 90% of the root length with no
obvious harmful effect on the plant.
Depending on the environment of the plant, mycorrhizae can increase a plant’s
competitiveness. In wet environments they increase the availability of nutrients, especially
phosphorus. In arid environments, where nutrients do not limit plant functioning to the same
degree, the mycorrhizae aid in water uptake, allowing increased transpiration rates in
comparison with nonmycorrhizal plants. These benefits have distinct energy costs for the
plant in the form of photosynthate required to support the plant’s “mycorrhizal habit”. Under
certain conditions the plant is apparently willing to trade photosynthate produced with the
increased water acquisition for H2O.
The beneficial effect on the plant of the mycorrhizal fungus is best observed in poor soils,
where trees that are mycorrhizal, thrive but non mycorrhizal one do not. When trees are
planted in prairie soils, which ordinarily lack a suitable fungal inoculum, trees that were
artificially inoculated at the time of planting grow much more rapidly than uninoculated trees.It is well established that the mycorrhizal plant is able to absorb nutrients from its
environment more efficiently than does a nonmycorrhizal one. This improved nutrient
absorption is probably due to the greater surface area provided by the fungal mycelium.
Suggested Reading
1. Avances in Agricultural Microbiology (1982) by Subba Rao.
2. Agricultural Microbiology (1996) by G. Rangaswamy and D.J. Bagayaraj.3. Brock Biology of Microorganisms (1997) by Michael T. Madigan, John M. Martinko, Jack Parker.4. Ecology of Microbial Communities (1987) by M. Fletcher, T.R.G. Gray & J.G. Jones.
5.
Introduction to Soil Microbiology (1961) by Martin Alexander.6. Microbiology (1999) by Lansing M. Prescott, John P. Harley & Donald A. Klein.7. Nitrogen Fixation in Plants by (1986) R.O.D. Dixon & C.T. Wheeler.

7/29/2019 Terrestrial Environment
http://slidepdf.com/reader/full/terrestrial-environment 28/28
8. Plant-Microbe Interactions. Molecular & Genetic Perspectives (1984) Vol. 1 by Tsune Kosnge &Eugene W. Nester.
9. Plant Microbe Interaction in Sustainable Agriculture (1995) by R.K. Behl, A.L. Khurana & R.C.Dogra.
10. Soil Microbiology by (1994) Robert L. Tate III.