the and acanthamoeba account for to c · 2014-08-07 · l.m. macheskyetal. offilaments (pollard...
TRANSCRIPT

CELL REGULATION, VOl. 1, 937-950, November 1990
The affinities of human platelet and Acanthamoebaprofilin isoforms for polyphosphoinositides account fortheir relative abilities to inhibit phospholipase C
Laura M. Machesky,* Pascal J. Goldschmidt-Clermont,t and Thomas D. Pollard*Johns Hopkins University School of Medicine*Department of Cell Biology and Anatomyand tDepartment of Medicine(Division of Cardiology)Baltimore, Maryland 21205
In light of recent work implicating profilin from hu-man platelets as a possible regulator of both cyto-skeletal dynamics and inositol phospholipid-me-diated signaling, we have further characterized theinteraction of platelet profilin and the two isoformsof Acanthamoeba profilin with inositol phospholip-ids. Profilin from human platelets binds to phos-phatidylinositol-4-monophosphate (PIP) and phos-phatidylinositol-4,5-bisphosphate (PIP2) withrelatively high affinity (Kd- 1 gM for PIP2 by equi-librium gel filtration), but interacts only weakly (ifat all) with phosphatidylinositol (PI) or inositol tris-phosphate (IP3) in small-zone gel-filtration assays.The two isoforms of Acanthamoeba profilin bothhave a lower affinity for PIP2 than does humanplatelet profilin, but the more basic profilin isoformfrom Acanthamoeba (profilin-l1) has a much higher(- 10-MM KdJ affinity than the acidic isoform (profilin-1, 100 to 500-MM KQ). None of the profilins bind tophosphatidylserine (PS) or phosphatidylcholine (PC)in small-zone gel-filtration experiments. The differ-ences in affinity for PIP2 parallel the ability of thesethree profilins to inhibit PIP2 hydrolysis by solublephospholipase C (PLC). The results show that theinteraction of profilins with PIP2 is specific with re-spect to both the lipid and the proteins. In Acan-thamoeba, the two isoforms of profilin may havespecialized functions on the basis of their identical(- 10 M) affinities for actin monomers and differentaffinities for PIP2.
Introduction
Profilin was originally identified as a small actinmonomer-binding protein (Carlsson etal., 1977),but has been recently been shown to associate
with plasma membranes (Hartwig, et al., 1989)and to inhibit soluble phospholipase C (PLC) bybinding to phosphatidylinositol 4,5 bisphos-phate (PIP2) (Goldschmidt-Clermont et al.,1990). Profilin may provide a link between theprocesses that regulate transmembrane sig-naling and the cytoskeleton, but sorting out thecomplex relationships at these regulatorycrossroads will require careful characterizationof a variety of molecular interactions, a task stillin its infancy.Members of the profilin family vary consid-
erably in their primary structures. Only 18 of125 residues are conserved among the profilinsfrom vertebrates (Nystrom et al., 1979; Kwiat-kowski and Bruns, 1988), Acanthamoeba (Ampeet al., 1985, 1988), and yeast (Oechsner et al.,1987). But comparison of any two profilins (suchas human platelet and yeast) yields considerablymore sequence identity. There are single iso-forms in yeast and man, but at least three iso-forms in Acanthamoeba (2 neutral and 1 basic).
In spite of these differences in primary struc-ture, all well-characterized profilins bind actinmonomers with micromolar affinity (Pollard andCooper, 1986), increase the rate of exchangeof ATP bound to actin (Mockrin and Korn, 1980;Nishida, 1985), inhibit the actin monomerATPase (Tobacman and Korn, 1982), and inhibitactin polymerization (Reichstein and Korn, 1979;Dinubile and Southwick, 1985; Kaiser et al.,1986). Formation of a nonpolymerizable 1:1complex with actin monomers was originallythought to explain all of these effects on actin,but more recent data (Pollard and Cooper, 1984;Kaiser et al., 1986; Goldschmidt-Clermont, un-published observations) show that the mecha-nism of action is more complex. For example,in Acanthamoeba, binding to actin monomerscan account for the inhibition of nucleation andelongation at the pointed end of actin filaments,but not for the weak inhibition of elongation atthe barbed end (Tseng and Pollard, 1982; Tsenget al., 1984; Tilney et al., 1983; Kaiser et al.,1986). These observations can be explained bylow-affinity binding of profilin to the barbed end
© 1990 by The American Society for Cell Biology 937

L.M. Machesky et al.
of filaments (Pollard and Cooper, 1984). In thecell, these properties of profilin should suppressspontaneous nucleation but allow actin fila-ments to grow at one end. However, theseeffects of profilin require stoichiometric con-centrations of profilin, whereas the actinconcentration in both human platelets andAcanthamoeba exceeds the profilin concentra-tion by at least two- to sixfold (Tseng etal., 1984;Lind et al., 1987; Goldschmidt-Clermont, un-published observations). On the other hand, thecatalytic effect of profilin on exchange of bothnucleotide and divalent cation on actin mono-mers could contribute to the regulation of actinpolymerization, even at the substoichiometricconcentrations found in cells.The potential regulatory roles of profilin in
cells were expanded by the discoveries thatPIP2 dissociates the actin-profilin complex(Lassing and Lindberg, 1985, 1988) and thatprofilin binds to PIP2 in a manner that protectsthe lipid from hydrolysis by a soluble, phos-phoinositide-specific PLC-Il from humanplatelets (Goldschmidt-Clermont et al., 1990).Because platelet profilin has a higher affinityfor PIP2 than actin, much of the profilin mightbe bound to lipids and regulate both phospha-tidylinositol (PI) turnover and actin dynamics.These findings have raised many questionsabout the specificity of the interaction betweenprofilin and lipids, the mechanism of dissocia-tion of the profilin-PIP2 complex, and the sig-nificance of both the profilin-actin and profilin-PIP2 associations.
In this paper, we provide evidence that theinteraction of profilin with polyphosphoinosi-tides is specific for both the lipid and the protein.On the lipid side, profilins bind to phosphoino-sitides with a phosphate in the 4 position, butnot to Pi or to other acidic phospholipids, suchas phosphatidylserine (PS). On the basis of thelow affinity of profilin for inositol trisphosphate(OP3), we think that the affinity for individualphosphoinositide head groups is very low.Binding is achieved by the association of profilinwith small clusters of PIP2. On the protein side,two basic profilins (human platelet profilin andAcanthamoeba profilin-l1) bind to PIP2 muchmore strongly than profilin-l, the acidic isoformfrom Acanthamoeba. This is the first evidencefor a functional difference between profilin-I andprofilin-Il. Given that these profilins bind actinmonomers with equal affinity, we hypothesizethat profilin-ll is mainly membrane associatedand involved with phosphoinositide metabolism,while profilin-l acts primarily as a regulator ofthe actin cytoskeleton. The single isoform of
profilin in platelets has a high affinity for bothactin and phosphoinositides and may have dualregulatory functions.ResultsThree profilins interact differently withpolyphosphoinositides during small-zone gelfiltrationIn small-zone gel-filtration experiments, humanplatelet profilin (Goldschmidt-Clermont et al.,1990), Acanthamoeba profilin-l, and Acantha-moeba profilin-ll all bound to PIP2 micelles, buteach bound in a different manner (Figure 1). Agiven concentration of PIP2 micelles bound hu-man platelet profilin > profilin-ll > profilin-l. Theconcentration of each profilin carried from thestarting sample by the micelles was directlyproportional to the concentration of PIP2, butthe slopes of the plots differed for each profilin(Figure 1A). One human platelet profilin boundfor an average of 4.3 PIP2 molecules. This is aslightly larger ratio than the 1:7 ratio in our pre-vious report (Goldschmidt-Clermont et al.,1990). The lower ratios for profilin-ll (' 1:15) andprofilin-l (- 1:50) are unlikely to result from dif-ferences in the stoichiometries, as the data inthe following sections show that this can be ex-plained by lower affinities. Because of the loweraffinities of the Acanthamoeba profilins for PIP2,dissociation of the PIP2-profilin complex occursduring chromatography, and higher concentra-tions of PIP2 are required to bind these profilins.
Platelet profilin and Acanthamoeba profilin-llbound to micelles of PIP2 under a variety of con-ditions. By small-zone gel filtration, binding wassimilar in 0 and 75 mM KCI at pH 7.5 and overa range of pH 5.5-8.5 in 75 mM KCI (data notshown).When incorporated into large unilamellar
vesicles made by an extrusion technique(LUVETS) of mixed composition, PIP2 boundhuman platelet profilin > profilin-l1 > profilin-I(Goldschmidt-Clermont et al., 1990; and Figure1 B). The PIP2 concentration dependence ofbinding was similar for LUVETS and micelles.The slopes of the plots gave ratios of 1 humanplatelet profilin per 5 PIP2 (Goldschmidt-Cler-mont et al., 1990) and 1 Acanthamoeba profilin-11 per 12 PIP2 (Figure 1B). Binding of profilin-lrequired much more PIP2 than we could use inthis assay. Because steric hindrance should notaffect the binding of profilin to large unilam-mellar vesicles composed mostly of PC, we at-tribute these differences in binding ratios to thelower affinities of the Acanthamoeba profilinsfor PIP2 and the resulting dissociation of thelipid-profilin complex during chromatography.
CELL REGULATION938

Specific interaction of profilin PIP2
20
16
12
8
[Micellar PIP2] ,IM
20
16
12
8
4
[Acid-P-Lipid] jM
20
16
12
0 100 200 300[P-inositide] jM
Figure 1. Binding of various profilins to lipids measLby small-zone gel filtration. Small-zone gel filtrationperformed with human platelet profilin, Acanthamoebafilin-I, and Acanthamoeba profilin-ll with micelles of I
PIP2 or PIP, SUVs of Pi, or LUVETS of mixed lipid consition. Conditions: samples of 100 ul containing 23profilin and various concentrations of lipids in VB vpreincubated for 15-30 min and chromatographed on ax 7-cm column of Sephadex G-1 00 equilibrated with V22°C. The concentration of free profilin was measurethe Bradford assay (Bradford, 1976) and the fraction bcto lipid calculated by difference. (A) Profilin from huiplatelets (-), amoeba profilin-l (0), or amoeba profilin-Iwith micelles of pure PIP2. (B) Amoeba profilin-l with LUVof PC:PIP2 5:1 (O) and PC:PS 5:1 (0) and amoeba profil
Binding of profilins is specific for phosphati-dylinositol-4-monophosphate (PIP) and PIP2. Insmall-zone gel-filtration experiments, humanprofilin bound equally well to micellar PIP andPIP2, but only weakly to small unilamellar vesi-cles (SUVs) of Pi (Figure 1 C). The stoichiometryone profilin per three or four molecules of PIP,and the fact that binding was directly propor-tional to the concentration of lipid added, areonly compatible with a high-affinity interaction.There was no detectable interaction between
400 human platelet profilin and IP3 by gel filtration(Figure 2), so the affinity of profilin for individualPIP2 headgroups is low relative to the affinityfor PIP2 pentamers in a membrane.No detectable Acanthamoeba profilin-l or
B profilin-l1 bound to LUVETS of phosphatidyl-choline (PC):PS (5:1) during small-zone gel fil-tration. This agrees with our studies using hu-man platelet profilin, where we also showed nodetectable binding to LUVETS of PC:PS (5:1) orPC: Pi (5:1) (Goldschmidt-Clermont et al., 1990).These results support the specificity of the in-teraction between profilin and polyphosphoino-sitides.
400 Affinities of profilins for PIP2 determined byequilibrium gel filtrationWe used equilibrium gel filtration to measurethe affinities of the three profilins for PIP2 and
- other lipids (Figure 3 and Table 1). These mea-surements confirmed that human platelet pro-filin has the highest affinity for PIP2, profilin-lIhas an intermediate affinity, and profilin-l fromAcanthamoeba has the lowest affinity for PIP2,in agreement with the small-zone gel-filtrationassays. As in the small-zone gel-filtration as-says, no detectable interaction occurred be-tween LUVETS of PC:PS and either Acantha-moeba profilin-l1 (Figure 3, C and D) orAcanthamoeba profilin-I (not shown).
400 Because this is a novel application of themethod of Hummel and Dreyer (1962), a fewcomments are in order regarding the equilibrium
ured gel-filtration method. This method is tradition-was ally used to measure the binding of small mol-pro- ecules to macromolecules. We adapted it topurenpo- measure the binding of small proteins to largeAM lipid assemblies (LUVETS). First, a sample con-vere0.7
'B atd bylundman11(-)FETSlin-l1
with LUVETS of PC:PIP2 5:1 (m) and PC:PS 5:1 (-). [Acidic-P-Lipid] corresponds to the concentration of the acidicphospholipid species in the outer leaflet of the vesicles (as-sumed to be 50% of the total concentration), e.g., PIP2, Pi,or PS. (C) Human platelet profilin with micelles of PIP (e),PIP2 (0), and SUVs of Pi (O).
Vol. 1, November 1990
0
C
0
c
a-0~4-0
939
L.M. Machesky et al.
1.2
1.0
0 0.8
S 0.6IL_ 0.4
0.2
0.0
.I-I
P-
cL_l
0 10 20 30 40
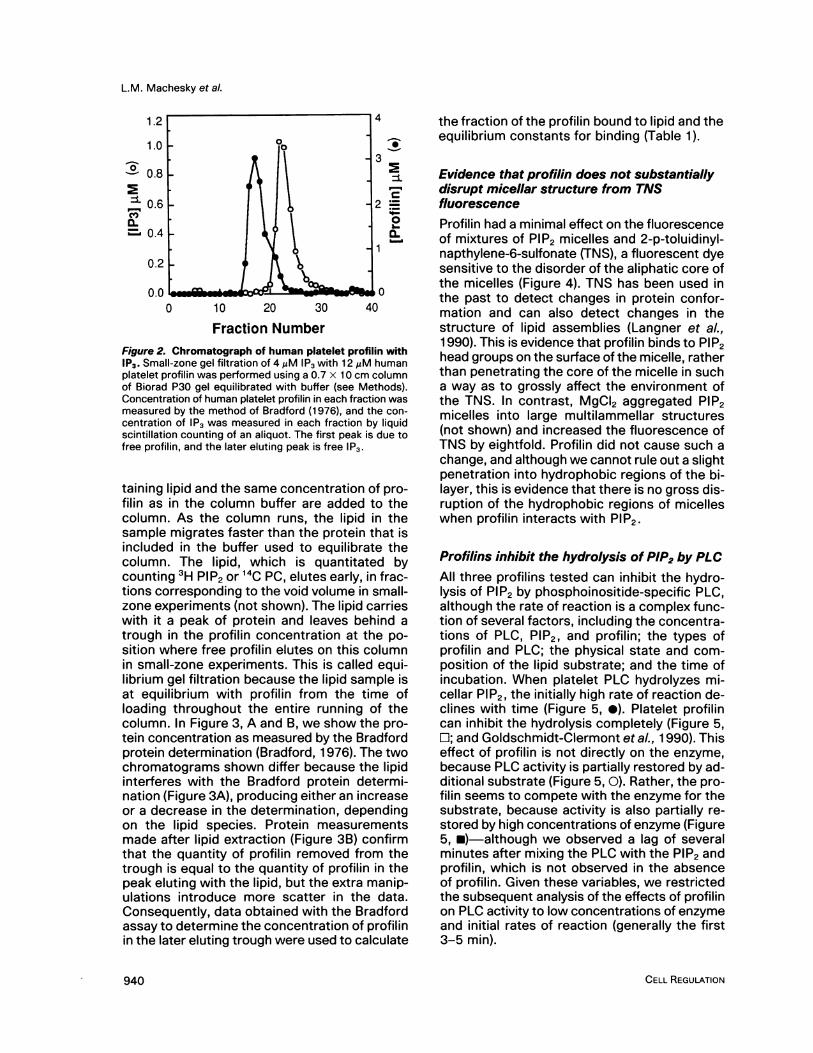
Fraction NumberFigure 2. Chromatograph of human platelet profilin withIP3. Small-zone gel filtration of 4 ,uM IP3 with 12 gM humanplatelet profilin was performed using a 0.7 x 10 cm columnof Biorad P30 gel equilibrated with buffer (see Methods).Concentration of human platelet profilin in each fraction wasmeasured by the method of Bradford (1976), and the con-centration of IP3 was measured in each fraction by liquidscintillation counting of an aliquot. The first peak is due tofree profilin, and the later eluting peak is free IP3.
taining lipid and the same concentration of pro-filin as in the column buffer are added to thecolumn. As the column runs, the lipid in thesample migrates faster than the protein that isincluded in the buffer used to equilibrate thecolumn. The lipid, which is quantitated bycounting 3H PIP2 or 14C PC, elutes early, in frac-tions corresponding to the void volume in small-zone experiments (not shown). The lipid carrieswith it a peak of protein and leaves behind atrough in the profilin concentration at the po-sition where free profilin elutes on this columnin small-zone experiments. This is called equi-librium gel filtration because the lipid sample isat equilibrium with profilin from the time ofloading throughout the entire running of thecolumn. In Figure 3, A and B, we show the pro-tein concentration as measured by the Bradfordprotein determination (Bradford, 1976). The twochromatograms shown differ because the lipidinterferes with the Bradford protein determi-nation (Figure 3A), producing either an increaseor a decrease in the determination, dependingon the lipid species. Protein measurementsmade after lipid extraction (Figure 3B) confirmthat the quantity of profilin removed from thetrough is equal to the quantity of profilin in thepeak eluting with the lipid, but the extra manip-ulations introduce more scatter in the data.Consequently, data obtained with the Bradfordassay to determine the concentration of profilinin the later eluting trough were used to calculate
the fraction of the profilin bound to lipid and theequilibrium constants for binding (Table 1).
Evidence that profilin does not substantiallydisrupt micellar structure from TNSfluorescenceProfilin had a minimal effect on the fluorescenceof mixtures of PIP2 micelles and 2-p-toluidinyl-napthylene-6-sulfonate (TNS), a fluorescent dyesensitive to the disorder of the aliphatic core ofthe micelles (Figure 4). TNS has been used inthe past to detect changes in protein confor-mation and can also detect changes in thestructure of lipid assemblies (Langner et al.,1990). This is evidence that profilin binds to PIP2head groups on the surface of the micelle, ratherthan penetrating the core of the micelle in sucha way as to grossly affect the environment ofthe TNS. In contrast, MgCI2 aggregated PIP2micelles into large multilammellar structures(not shown) and increased the fluorescence ofTNS by eightfold. Profilin did not cause such achange, and although we cannot rule out a slightpenetration into hydrophobic regions of the bi-layer, this is evidence that there is no gross dis-ruption of the hydrophobic regions of micelleswhen profilin interacts with PIP2.
Profilins inhibit the hydrolysis of PIP2 by PLCAll three profilins tested can inhibit the hydro-lysis of PIP2 by phosphoinositide-specific PLC,although the rate of reaction is a complex func-tion of several factors, including the concentra-tions of PLC, PIP2, and profilin; the types ofprofilin and PLC; the physical state and com-position of the lipid substrate; and the time ofincubation. When platelet PLC hydrolyzes mi-cellar PIP2, the initially high rate of reaction de-clines with time (Figure 5, 0). Platelet profilincan inhibit the hydrolysis completely (Figure 5,OL; and Goldschmidt-Clermont etal., 1990). Thiseffect of profilin is not directly on the enzyme,because PLC activity is partially restored by ad-ditional substrate (Figure 5, 0). Rather, the pro-filin seems to compete with the enzyme for thesubstrate, because activity is also partially re-stored by high concentrations of enzyme (Figure5, *)-although we observed a lag of severalminutes after mixing the PLC with the PIP2 andprofilin, which is not observed in the absenceof profilin. Given these variables, we restrictedthe subsequent analysis of the effects of profilinon PLC activity to low concentrations of enzymeand initial rates of reaction (generally the first3-5 min).
CELL REGULATION940

Specific interaction of profilin PIP2
8
61
41
2
0
8
6
4
2
o0 10 20 30 40 0 10 20 30 40
Fraction NumberFigure 3. Binding of Acanthamoeba profilin-ll to lipids measured by equilibrium gel filtration. In these experiments, the0.7 x 50-cm column of Sephadex G-1 00 was equilibrated with VB containing 5 AM profilin-l1; and 600-il samples, containingboth Acanthamoeba profilin-ll (5 ,uM) and lipids, were chromatographed through the column. (A) Chromatogram of Acan-thamoeba profilin-ll with micellar PIP2. The profilin concentration in 0.45-ml fractions was assayed by the method of Bradford(1976). The first trough is due to negative interference of the lipid with the assay, whereas the second trough represents thedepletion of profilin because of interaction with lipid. (B) The same chromatogram as in A, assayed for protein by the BCAmethod after extraction of the lipids. This demonstrates that the profilin concentration is higher in the fractions where thelipids elute (here fractions 15-20, corresponding to the void volume in small-zone experiments). (C) The sample contained300 AM PS in PC:PS (2:1) LUVETS. No interference was observed of the PC:PS LUVETS with the Bradford assay. (D) Thesame chromatogram as in C, assayed by the BCA method after extraction of the lipids.
The ability of a given profilin to inhibit PIP2hydrolysis by PLC (Figure 6 and Table 2) cor-relates well with its affinity for PIP2 as measuredby equilibrium gel filtration (Table 1), with humanplatelet profilin being the strongest inhibitor andAcanthamoeba profilin-l being the weakest. Thetype and concentration of profilin are major de-terminants of the PLC activity (Figure 6, A-C).We obtained our cleanest data for all three pro-filins with PLC-y from bovine brain (Figure 6A),where the inhibition of PIP2 hydrolysis parallelsthe affinities of the three profilins for PIP2. Theapparent dissociation constants for a complexof profilin with a pentamer of PIP2 can be esti-mated from the theoretical curves in Figure 6Ato be -3,M for platelet profilin, 70 ,M for
Acanthamoeba profilin-ll, and 500 ,uM for Acan-thamoeba profilin-l.The composition and concentration of lipids
in the vesicles are major determinants of thePLC activity in the presence of profilins, so theapparent affinity of profilin for PIP2 varies de-pending on the lipid substrate. Although PLChydrolyzed PIP2 at the same rate with a high(Figure 6C) or low (Figure 6B) mole fraction ofPIP2 in the outer leaflet of the LUVETS (data notshown), the Acanthamoeba profilins inhibitedthe enzyme more strongly when assayed witha high mole fraction of PIP2 in the outer leaflet.These Acanthamoeba profilins with low affinityfor PIP2 may not compete well with PLC for PIP2when the PIP2 is widely dispersed in the bilayer.
Vol. 1, November 1990
A
*as________?g is
c
g~~~~~w-S-s- -l-ff Ofl
l . I lC
6-0L..IL B1
0
II I
D
o- m-- -
0
941

L.M. Machesky et al.
Table 1. Determination of binding constants by equilibrium gel filtration
Kd zM
PIP2 micelles PC:PIP2 LUVETS
Acanthamoeba profilin-I 143 (D = 17, n = 2) 101 (D = 15, n = 2)Acanthamoeba profilin-lI 8 (SD = 3, n = 5) 3 (SD = 2, n = 3)Human platelet profilin 0.6 (D = 0.3, n = 2) 3 (n = 1)
Dissociation constants (Kd) for profilin and PIP2 (7:1 PIP2:profilin for micelles,and 5:1 PIP2:profilin for LUVETS) as determined by equilibrium gel filtration. SDcorresponds to the standard deviation where more than two replicate experi-ments were performed (n > 2); D, the deviation from the mean where twoexperiments were performed (n = 2).
Profilin inhibited equally well the two solublePLCs that we tested, phosphoinositide-specificPLC-,y from bovine brain (Figure 6A) and solublephosphoinositide-specific PLC-Il from humanplatelets (Figure 6, B and C). In contrast, profilindid not inhibit PLC beta from bovine brain, anenzyme thought to be associated with mem-branes (Goldschmidt-Clermont, unpublishedobservations).
Discussion
Mechanism of interaction of profilin withpolyphosphoinositidesThe available evidence suggests that membersof the profilin family interact specifically withPIP and PIP2 but that the mechanism is com-plicated because the protein binds to a smallcluster of lipid molecules. In this section we willdefine the overall mechanism as we now un-derstand it.The lipid requirements for binding profilin in-
clude an inositol head group with at least onephosphate in addition to the phosphodiesterlinkage to the glycerol. A phosphate in the 4position is apparently sufficient. Single phos-phates at other positions have not been tested.Neither PI nor PS is a good ligand, so the in-teraction is not simply an electrostatic associ-ation with a negatively charged head group. Thelack of detectable binding of IP3, at concentra-tions where profilin binds to PIP2, provides animportant clue discussed further below. Thephysical state of the phosphoinositide does notappear to be important, because PIP2 bindsprofilins equally well, whether it is concentratedin small micelles or dispersed among neutrallipids in a bilayer. The concentration of lipid isimportant, as in any association reaction. Themole fraction of polyphosphoinositide in a bi-layer is probably an important variable with re-
spect to the ability of profilin to interact withlipid clusters, as indicated by the PLC experi-ments with amoeba profilins (Figure 6C), but wehave not yet studied this systematically.The stoichiometry of the molecules in the
complex is one of the most important parame-ters. From the small-zone gel-filtration bindingassays and the PLC inhibition experiments withhuman platelet profilin, we estimate the stoi-chiometry to be one profilin bound to four orfive PIP2s. This number needs further verifica-tion, because neither assay is accurate enoughto determine the stoichiometry to within morethan ±±2 lipid molecules. The experimentswith the amoeba profilins provide no evidenceregarding the stoichiometry, because their af-finity is too low to obtain an accurate measure-ment. Therefore, we assumed that the stoichi-ometry of the complex of amoeba profilins withPIP2 is also 1:5.Our current interpretation of this data is that
profilins have multiple weak binding sites for in-dividual phosphoinositide head groups and thata stable complex is formed by possibly sequen-tial association with up to five lipid head groups.We have quantitated this interaction by assum-ing that each profilin molecule binds simulta-neously to a pentamer unit of lipid molecules.Such clusters with the appropriate geometry areprobably readily available on the surface of purePIP2 micelles, but when the PIP2 is dispersed ina bilayer with other lipids, the cluster of PIP2probably has to form by multiple associationsof individual lipid molecules with the protein. Wespeculate that this would start with a weaklybound 1:1 complex and grow as PIP2 moleculesdiffuse into association with the protein. Con-sequently, the dissociation constants that wemeasure are not equilibrium dissociation con-stants in the sense of a typical bimolecular re-action. Rather, these apparent equilibrium con-
CELL REGULATION942

Specific interaction of profilin PIP2
0
enc0)L..0
370 430 490 550 610
Wavelength (nm)
Figure 4. The effect of human platelet profilin on thestructure of PIP2 micelles measured by TNS fluorescence.The fluorescence emission spectrum of 30 MM TNS excitedat 360 nm, in the presence of PIP2 micelles and variousmolecules, was measured to analyze their effect on thestructure of the micelles. Conditions: (a) 100MgM TNS alone;(b) TNS with 10 uM PIP2; (c) TNS and 7MAM profilin; (d) TNSwith 7 MM profilin and 10 MM PIP2; (e) 10 mM MgCI2 withTNS and 10 AM PIP2. Profilin does not significantly changethe fluorescence of TNS in the presence of PIP2 micelles,whereas Mgcl2 has a marked effect on TNS fluorescencein the presence of PIP2.
stants are a composite value reflecting multiple,possibly cooperative, interactions. Given the lowaffinities, we suspect that these complexes ofprofilin with PIP2 have a relatively short lifetime,but the weak binding may also be the conse-quence of slow association rates rather thanrapid dissociation rates. We hope that the for-mation and dynamics of these complexes canbe studied further by both kinetic and structuralmethods.The evidence suggests that electrostatic in-
teractions between the negatively chargedphosphosphate on the inositol and the proteinare important but not sufficient to explain theobserved binding. Because other acidic headgroups do not bind profilin (Figures 1, B and C;
and 3, C and D), the geometry of the phospho-inositide must play an essential role. The stablebinding of platelet profilin and profilin-ll in 0-75mM KCI over the pH range 5.5-8.5 does notrule out electrostatic interactions, becausethere is evidence that PIP2 is expected to havea net charge of - -3 (Toner et al., 1988) overthis pH range in 75 mM KCI. However, there isalso evidence suggesting that in 100 mM NaCI,the pK of PIP2 is variable over this pH range(van Paridon et al., 1986).
Clues about polyphosphoinositide bindingfrom comparison of profilin sequencesThe sequences of all profilins are weakly ho-mologous from end to end, but the vertebrateprofilins differ so much from the amoeba pro-filins that a detailed sequence comparison toexplain human platelet profilin's higher affinityfor PIP2 was not useful in the absence of knowl-edge about their three-dimensional structures.Nonetheless, the two profilins that bind well toPIP2 have one common feature that may be rel-evant. Both human platelet profilin and Acan-thamoeba profilin-ll have basic isoelectric points(pl - 9), whereas Acanthamoeba profilin-l isacidic (pI - 5.5).Because Acanthamoeba profilin-l and profilin-
11 differ at only 22 out of 125 residues (Table 3),
1.5 -
00
Ec.-I
0U)z
C-
1.0
0.51p
0.0'0 2 4 6 8 10
Minutes
Figure 5. Influence of platelet profilin, substrate, and en-zyme concentrations on the time course of PIP2 hydrolysisby platelet PLC. Release of IP3 from micellar PIP2 was as-sayed according to Methods. Conditions: (0) Control with47 Mg/mI PLC and 7.9 MM micellar PIP2; (O) 47 Mg/ml PLC,7.9MM PIP2, and 1.25MuM profilin, which completely inhibitsactivity; (0) 47 Mg/ml PLC, 1.25MgM profilin, and excess (19.1MM) PIP2, which restores activity in the presence of theprofilin; (m) 7.9 MM PIP2, 1.25 MM profilin in the presence ofa high (1 10 Mg/ml) concentration of PLC, which gives a lowlevel of activity.
Vol. 1, November 1990
011-71",
0
943

L.M. Machesky et al.
13 these differences provide some clues about the° o Kd 10-fold difference in their affinities for PIP2.[50SM Most of these substitutions involve uncharged
residues or the exchange of two residues with80jM the same charge. Substitutions at residues 24,l6tt-g1M 50, and 66 result in differences in charge, ac-
count for the higher isoelectric point of profilin-11, and give it a net charge of +2 higher thanprofilin-l at pH 7. Because profilin-llbinds well
3P>zM to PIP2 at pH 5.5 and 8.5, lysine-50 is more likely., . , . , . to account for its higher affinity than histidine-
0 10 20 30 40 24 (leucine in profilin-l) or arginine-66 (histidinein profilin-l).Although the substitution of a lysine for an
l 3nKd asparagine may seem like a minor difference13
Kd
° 500pM between profilin-l and profilin-l1, single chargedresidues can have a profound effect on the af-finity of peptides for acidic phospholipids. In**20gM model studies of the binding of small positivelycharged peptides derived from protein kinaseC (PKC) to vesicles composed of PS and PS:PC, each lysine or arginine residue contributed
lO\ about one order of magnitude (a free energy ofO.1gM 1.4 kcal/mol) to the affinity (Kim et al., unpub-.o lished observations). PKC peptides are an im-
0 5 10 15 20 perfect model for profilin-lipid interactions, be-cause they bind to vesicles containing PS orphosphatidylglycerol (PG), but the conclusionregarding the energic contribution of a singleelectrostatic bond may be sufficient to explain
13 Kd the higher affinity of profilin-1l for PIP2.* E # A Mechanism of PLC inhibition by profilin
o3 The best explanation for the inhibition of PLCby profilin is sequestration of substrate PIP220 W (Figure 7). Under a wide variety of conditions,the ability of the three profilins to inhibit theenzyme parallels their affinity for PIP2 measured
.______._____._____._____.____ in binding reactions (Tables 1 and 2). The0 5 10 15 20 25 agreement is not precisely quantitative, likely
[profilin] gM because of the complexity of the interactionsamong PIP2, profilin, and PLC in the enzyme
Inhibition by profilins of the hydrolysis of PIP2 assay. For example, even at high concentra-a PLC. The samples contained profilin, a PLC iso- tions, Acanthamoeba profilin-l1 never inhibitedJ various lipids (micelles or LUVETS). Theoretical ' . .PLC activity were calculated by a simple model PLC as well as platelet profilin (Figure 6). Per-
;sociation constants (Kd) between 0.1 and 1000 haps the subtle difference in the affinity of hu-dicated. (A) Activity of bovine brain PLC-y using man platelet profilin and Acanthamoeba profilin-)f PC:PIP2 (5:1) in the presence of (O) Acantha- 11 for PIP2 is magnified when competing withofilin-l, (0) Acanthamoeba profilin-ll, or (0) human PLC. This seems particularly likely, given therofilin. Theoretical curves for a complex of profilin . . . . opentmerardrwn or KS o 50 zM 80,uM possibly cooperative Interaction of one profilinpentamers are drawn for Kds of 500,MM, 80 MAM,
wt iePP oeue.Frhrsseaiid 3 MM. (B) Activity of human platelet PLC using with five PIP2 molecules. Further systematicif PC:PE:PIP2 (5:5:1) in the presence of (L) Acan- studies of both the binding reactions and thewprofilin-l, (0) Acanthamoeba profilin-ll, or (0) hu- enzyme activity will be required to substantiateblet profilin. Theoretical curves are drawn for Kds the mechanism of inhibition.In ..KA OX ..MA nnl n I ..M lr\ Antivitti nf hi imanUl oJW AIVI, I UV AIvi, LU AIVI, GIlU U. I fIVI. 1%..) m;IVIly Ul numat
platelet PLC using LUVETS of PC:PE:PIP2 (1:1:1) in thepresence of (0) Acanthamoeba profilin-l, or (0) Acantha-moeba profilin-l1. Theoretical curves are drawn for 50 ,Mand 20 MM.
Functions of profilins in cellsThe actin and lipid binding capabilities of pro-filins suggest that they are unique regulatory
CELL REGULATION
120
0a-'.'
C.)
120
100
80
60
40
20
0
Figure 6.by solublezyme, anccurves forwhere disMM are inLUVETS cmoeba pr(platelet prwith PIP260 WM, anLUVETS athamoebaman platen%f Rlnn I'M
120
o n -
944

Specific interaction of profilin PIP2
Table 2. Affinity of profilins for PIP2 estimated from their effects on PLC activity
Substrates
Kd AM (profilin-LUVETS) Kd AM (profilin-micelles)
1:1:1, PC:PE:PIP2 5:5:1, PC:PE:PIP2 5:1, PC:PIP2 PIP2
Platelet PLC-IlAmoeba profilin-I 40-50 100-500Amoeba profilin-11 15-20 20-100Platelet profilin 0.1-10 0.1-1.0 1-5 0.01-0.1
Brain PLC--yAmoeba profilin-l 1000Amoeba profilin-Il 60-80Platelet profilin 1-10
We measured the activity of PLC-Il from human platelets and PLC-'y from bovine brain over a range of profilin concentrationsand vesicle compositions. The Kds of the profilin:PIP2 complex (7:1 PIP2:profilin for micelles and 5:1 PIP2:profilin for LUVETS)were estimated from theoretical curves, calculated assuming that all bound PIP2 is inaccessible to the PLC (Figure 6 andMaterials and methods).
proteins functioning at the interface of a majorsignaling pathway and the actin cytoskeleton,and our results show that these functions canbe partitioned in at least three different ways.Platelet profilin appears to be bifunctional withrelatively high affinities for both polyphospho-inositides and actin monomers; Acanthamoebaprofilin-Il has moderate affinities for both Ii-gands; and Acanthamoeba profilin-I seems tobe specialized for binding to actin. This is thefirst known functional difference between thetwo isoforms of profilin from Acanthamoeba.Not enough is known about profilin isoformexpression in other cells to predict whether thevertebrate or amoeba solution is more wide-spread.To estimate the potential physiological sig-
nificance of these interactions, we calculatedthe concentrations of the various species pres-ent at equilibrium in a mixture with cellular con-centrations of profilin, actin, and PIP2 (Table 4).Accepting for now that cells are much morecomplicated than three-component systems,these calculations show that platelet profilinwould be partitioned between unpolymerizedactin monomers and membrane PIP2, whereasamoeba profilin-Il would mostly be associatedwith PIP2, and amoeba profilin-I would be largelybound to actin monomers. In all cases, most ofthe profilin is bound to one of its ligands, but inno case is all of the unpolymerized actin ormembrane PIP2 complexed with profilin.These predictions, based on binding con-
stants measured with purified proteins, agreein a general way with observations on cells and
crude cellular fractions. For example, about one-third of total platelet profilin is associated withactin when isolated on p0ly-L-proline Sepharose(Lind et al., 1987; Goldschmidt-Clermont, un-published observations). Second, by electronmicroscopy, part of the profilin in blood cells isassociated with the plasma membrane (Hartwiget al., 1989). Third, part of cellular PIP2 is notaccessible for binding neomycin, presumablybecause it is already bound to a protein (Kinget al., 1987; Toner et al., 1988), perhaps profilin.It is an open question how many other proteinsmay also sequester PIP and PIP2. Three can-didates are the plasma membrane protein gly-cophorin (Anderson and Marchesi, 1985) andthe cytoplasmic proteins gelsolin (Janmey andStossel, 1987) and cofilin (Yonezawa et al.,1990).These new insights regarding interactions of
profilin, actin, and phosphoinositides provide anopportunity to reevaluate the potential physio-logical functions of profilin. With the partial in-formation currently available, it is not yet clearwhich molecules are the regulators and whichare regulated. For example, does profilin regu-late phosphoinositide metabolism, or do thephosphoinositides regulate profilin, or is theregulation reciprocal?
Originally profilin was viewed as an actinmonomer sequestering protein responsible infull or in part for the pool of unpolymerized actinin nonmuscle cells. However, given the affinityof the profilins foi lipids and the expected largeexcess of unpolymerized actin over the con-centration of profilin-actin complexes (Table 4),
Vol. 1, November199094945

L.M. Machesky et a!.
Table 3. Comparison of the variable residues of the three isoforms of Acanthamoeba profilin
22 24 38 41 43 44 47 50 51 53 54 57 58 61 65 66 70 77 82 84 94 97
Profilin-IA L L T Q Q T S N N D P A S D V H L I G A S VProfilin-IB L L T Q T T S N N D P A G D V H L G S A VProfilin-ll I H S N A A N K D T A S N E T R I V S A A I
The primary structures of profilin-IA and profilin-IB were determined by amino acid sequencing of a mixture of the two proteins(Ampe et al., 1985) and from the nucleotide sequence of profilin-IA (Pollard and Rimm, unpublished observations). Thesequence of profilin-l1 was determined by amino acid sequencing (Ampe et al., 1988) and confirmed and corrected by cDNAsequencing (T.D. Pollard and D.L. Rimm, unpublished observations). Because there are only three amino acid differencesbetween profilin-IA and profilin-IB, and because these isoforms cannot be separated by our current purification procedure,we have referred to them collectively as profilin-l throughout this paper. Differences at residues 24, 50, and 66, involvingcharged residues, may be particularly important in determining the differences between the affinities of profilin-l and profilin-11 for the polyphosphoinositides.
it is unlikely that the profilins alone account forthe pool of unpolymerized actin monomer incells. The predicted concentration of unpoly-merized actin not bound to profilin is well abovethe critical concentration required for polymer-ization, so it seems likely that part of theunpolymerized actin is bound to other seques-tering proteins, such as members of the acto-phorin/depactin/actin depolymerizing factor(ADF) group (Giuliano et al., 1988; Mabuchi,1983; Cooper et al., 1986), or to a recently dis-covered 5-kDa protein (Safer et al., 1990). Ifprofilin is not adequate to sequester the un-polymerized actin in the cell, we should considerother roles that are compatible with the lowconcentration of free profilin in the cytoplasm.One possibility is fine-tuning actin turnover inthe cell by enhancing nucleotide exchange.Profilin accelerates the exchange of nucleotideon actin monomers (Mockrin and Korn, 1980;Nishida, 1985). The profilin can accomplish thisat low concentrations because it acts catalyti-cally (Goldschmidt-Clermont, unpublished ob-servations). At the time of cell activation, profilinappears to associate with actin (Lind et al.,1987), presumably as a result of the activationof PLC, which allows it to overcome the profilinblock (Goldschmidt-Clermont, unpublished ob-servations) and thus begin rapid Pi-turnover andsubsequent release of profilin from the mem-brane. There is no information about the poolsof ADP-actin in cells, but if present, this releasedprofilin could promote the exchange of its ADPfor ATP and stimulate polymerization of actin.Profilin could also inhibit spontaneous nucle-ation of the ATP-actin (Pollard and Cooper,1984), so that polymerization takes place in anorganized fashion from preformed nucleatingsites in the cell. In these mechanisms we en-vision the phosphoinositides as the regulatorsof profilin, which in turn regulates the assemblyof actin.
946
The other side of this scheme is that profilinparticipates in the regulation of phosphoinosi-tide metabolism. In both platelets and Acan-thamoeba, profilin binds to PIP and PIP2 with ahigh enough affinity that a substantial portionof these lipids could be complexed with profilinin a cell (Table 4). Together with other phos-phoinositide-binding proteins, profilin may pro-tect PIP2 from hydrolysis by PLC until a cellbecomes activated (Forscher, 1989; Gold-schmidt-Clermont et al., 1990). We have inves-tigated only the effects of profilin on the hydro-lysis of PIP2 by PLC, but because profilin alsobinds to PIP, it may regulate phosphoinositidemetabolism in other ways, including the syn-
ProfilinA' PIP2
0 J
.
Phospholipid Bilayer
Figure 7. Model for the mechanism of PLC inhibition byprofilin. When profilin is added in a high concentration, itbinds to clusters of PIP2 on the micelles or outer leaflet ofvesicles, forming complexes that are poor substrates forhydrolysis by soluble PLC.
CELL REGULATION

Specific interaction of profilin PIP2
Table 4. Estimates of the concentrations of the species present in mixtures of physiological concentrations of profilin,actin, and PIP2
Bound to
Total uM Profilin-l uM Profilin-lI MM Actin MM PIP2 UM Free M
AcanthamoebaProfilin-l 67 50.9 3.2 12.9Profilin-ll 33 9.9 20.5 2.6PIP2 240 15.8 102.7 121.5Actin 200 50.9 9.9 100* 39.2
Bound to
Total MM Profilin MM 5 kDa MuM Actin MuM PIP2 MM Free zM
PlateletsProfilin 40 6.0 32.0 2.0PIP2 240 160 80Actin 200 6.0 86.4 100* 7.65 kDa 200 86.4 113.6
Concentrations and equilibrium dissociation constants were obtained from the following sources:Concentrations: Acanthamoeba profilins-Tseng and Pollard, 1982; Kaiser et al., 1986; Acanthamoeba actin-Gordon et al.,1976; Tseng and Pollard, 1982; Acanthamoeba PIP2-P.A. Allen, personal communication; platelet profilin and actin-Lindet al., 1987; Goldschmidt-Clermont, unpublished observations; platelet PIP2-Cohen et al., 1971; and platelet 5 kDa-Saferet al., 1990.Dissociation constants: Profilins and PIP2-this paper and Goldschmidt-Clermont et al., 1990; Acanthamoeba profilin andactin-Pollard and Cooper, 1984; platelet profilin and actin-Lind et al., 1987; Goldschmidt-Clermont, unpublished obser-vations; and platelet profilin and 5 kDa-Safer et al., 1990; Goldschmidt-Clermont, unpublished observation.We used the KINSIM program of Barshop et al. (1983) to calculate equilibrium distributions of each species. We understandthat cells are much more complex than simple four-component systems, and, actually, the free species are probably interactingwith other components of the cell. Until numbers are available for affinities and concentrations of other actin-binding proteinsas well as profilin's affinity for PIP and PIP's concentration in cells, however, a better model is not possible. * Refers to actinfilament concentration found in a resting cell.
thesis of PIP2 from PIP. Profilin or other PIP2-binding proteins may also be responsible formaintenance of a hormone-insensitive pool ofPIP2 (Koreh and Monaco, 1986). Furthermore,it will be interesting to examine the interactionof the profilins with phosphatidylinositol-3,4-bisphosphate and other minor phosphoinosi-tides, because so little is known about thesenovel phospholipids, and profilin may actuallyhave a higher affinity for one or more of theseminor species than for the common isomersthat we have studied.
Insight about the physiological relevance ofthe profilin-phosphoinositide interactions inAcanthamoeba will have to await further studies.Accurate quantitation of PIP and PIP2 concen-trations in the membrane has not been done,although estimates have been made that putthe concentration of PIP2 close to that found inother cells (>0.5% of total lipid, P.G. Allen, per-sonal communication), which would be -140-240 qM (Cohen et al., 1971). Acanthamoebachemotax toward bacteria on an agar substrate
(Mcintyre and Jenkins, 1969), and PI turnoverincreases with phagocytosis of either yeast orhigh-mannose glycoproteins (P.G. Allen, per-sonal communication), but neither the PLC norother parts of the signaling machinery havebeen identified. This information will be nec-essary to appreciate fully the meaning of thedifferences between the two profilin isoformsfrom Acanthamoeba. The available evidencepoints to two different roles, with profilin-ll beingmore like human platelet profilin and profilin-lbeing the major actin monomer-sequesteringprofilin.Perhaps the most interesting model is that
the interactions of phosphoinositides and pro-filins are part of a reciprocal regulatory network.The interaction of profilin with PIP2 might helpto explain the heterogeneity in cellular phos-phoinositide pools available for hydrolysis byPLC, whereas the interaction of profilin with ac-tin (regulated by the phosphoinositide concen-trations) might be key in reorganization of thecytoskeleton.
Vol. 1, November 1990 947

L.M. Machesky et al.
Materials and methods
Preparation of proteinsProfilins were purified from Acanthamoeba and outdatedhuman platelets by affinity chromatography on poly-L-prolineagarose (Kaiser et al., 1989). The profilin-l and profilin-ll iso-forms from Acanthamoeba were separated by carboxy-methylSepharose affinity chromatography (Kaiser et al.,1986). Profilins were dialyzed into vesicle buffer (VB; 10mMtris(hydroxymethyl)aminomethane [Tris], pH 7.5,75 mM KCI,0.5 mM dithiothreitol, and 1.8 mM NaN3). Protein concen-trations were measured by ultraviolet absorbance with anextinction coefficient of 0.015 OD * gM-' -cm-' at 280 nm.Phosphoinositide-specific PLC-Il was purified from humanplatelet cytosol (Baldassare et al., 1989) (a generous gift ofDr. J.J. Baldassare) and PLC-y from bovine brain (Sung etal., 1987) (a generous gift of Dr. S.G. Rhee).
Sources of lipidsLipids were obtained from the following sources: PIP2 fromCalbiochem (La Jolla, CA); PIP from Sigma Chemical (St.Louis, MO); PC, PS, phosphatidylethanolamine (PE), and PIin chloroform from Avanti Polar Lipids (Pelham, AL); phos-phatidyl [2-3H] inositol-4,5-bisphosphate ([3H] PIP2) in di-chloromethane:ethanol:water 20:10:1 (v/vlv), L-3 phospha-tidylcholine 1,2-di(1 14C) oleoyl ([14C] PC) in toluene:ethanol1:1 (v/v) IP3, and [3H]IP3 from Amersham (Arlington Heights,IL). The purity of the phosphoinositides PI, PIP, and PIP2was confirmed by thin-layer chromatography using 10 ,ugof each lipid loaded onto silica gel 60 plates in a 90:90:7:22chloroform:methanol:ammonium hydroxide:water solventsystem.
Preparation of lipid substratesHomogenous PIP2 and PIP micelles were prepared by sus-pending 1 mg of PIP2 or PIP in 0.5 or 1 ml of deionized waterand sonicating in a Bransonic 32 (bath type) sonicator for5 min at room temperature. SUVs of PI were prepared bydrying the lipid under a stream of nitrogen, resuspending inthe appropriate amount of deionized water by vortexing,and sonicating in a Bransonic 32 (bath type) sonicator asabove. Large unilamellar vesicles of various compositionswere made by an extrusion technique (LUVETS, Hope etal.,1985; Mayer et al., 1986). Lipids obtained in organic solvents,including the radiolabeled lipids (see below) were dried in aglass tube under a stream of nitrogen. PIP2, which had beenresuspended in deionized water, was added to the driedlipids, along with enough additional deionized water to pro-duce the desired final concentration; all lipids were thenresuspended by vortexing. After five cycles of freezing inliquid nitrogen and thawing in a 370C water bath, sampleswere passed 10 times through a filter (polycarbonate, 0.1-gM pore size, Nucleopore, Pleasanton, CA) in an Extruder(Lipex Biomembranes, Vancouver, Canada) under a pressureof 400 psi. The concentration of lipid in each mixture wasmeasured by liquid scintillation counting of an aliquot of thesample after extrusion.
Gel-filtration assay for profilin binding tolipidsFor small-zone experiments, samples were prepared in VBat room temperature and incubated for 15-30 min beforechromatography. Micelles and LUVETS with profilin wererun on a 0.7 x 7-cm column of Sephadex G-100, preequili-brated with VB, at room temperature at 20 ml/h, collectingfractions of 0.45 ml. Protein concentrations of each fraction
were measured with the Bradford Coomassie Blue dye-binding assay (Bradford, 1976) with purified profilin as thestandard. Because some lipids interfere with the Bradfordassay, we used only the free profilin peak in our calculations.Lipid concentrations of each fraction were measured by liq-uid scintillation counting of an aliquot. The fraction of profilinbound to lipids was calculated as the difference betweenthe total applied to the column and the amount recoveredin the entire peak of free protein.
Dissociation constants for the profilin-PIP2 complex weremeasured by equilibrium gel filtration (Hummel and Dreyer,1962) with a 0.7 x 50-cm column of Sephadex G-1 00 equil-ibrated with 3-20 MM profilin in VB. Samples of 600 AI con-tained various concentrations of lipid preequilibrated for 15-30 min with the same concentration of profilin as that usedin the column buffer. The following lipid concentrations andcompositions were used: micelles of pure PIP2 at 150-420MM, LUVETS of PC:PIP2 2:1 with 100-300 MM PIP2 in theouter leaflet, and LUVETS of PC:PS 2:1 with 300,MM PS inthe outer leaflet. The column was run at 20 ml/h, and 0.45-ml fractions were collected. Profilin bound to lipid was mea-sured as the amount of profilin depleted from the fractionswhere free profilin elutes (approximately fractions 21-29).Profilin bound to lipid was measured directly for two rep-resentative experiments, one with micelles and one withLUVETS. In these control experiments, 100-Ml aliquots ofeach fraction were acidified to 12 mM HCI and the lipidswere extracted with 10 vol of acetone for 1 h at 20°C. Afterspinning for 10 min in an Eppendorf centrifuge at 4°C, weremoved acetone with a pasteur pipette, and the proteinpellet was dried in a Speed Vac centrifuge (Savant, Farm-ingdale, NY). Pelleted protein was assayed by the BCAmethod (Pierce Chemical Co., Rockford, IL) using 96-wellplates and a microplate reader at 540 nm. Dissociation con-stants for the profilins and PIP2 were calculated assuminga stoichiometry of one profilin molecule per seven PIP2 mol-ecules in micelles and one profilin molecule per five PIP2molecules with LUVETS.
Gel filtration of IP3 and profilinThe interaction of profilin with IP3was tested by small-zonegel filtration using a 0.7 x 10-cm column of Biorad P30 gelpreequilibrated with 2 mM Tris pH 7.3, 50 mM KCI, 100 ,uMCaCI2, 100 MM NaN3, and 0.5 mM dithiothreitol. A 400-ulsample of 4 MM IP3 and 12 MM human platelet profilin, incolumn buffer, was run at 4°C; and 0.45-mI fractions werecollected. Trace amounts of [3Hl IP3 (specific activity 0.1 Ci/mole) were added to detect the position of elution of IP3,whereas protein concentration in each fraction was mea-sured by the Bradford assay.
TNS fluorescenceFluorescence of TNS (30 MAM in buffer: 2 mM Tris pH 7.5,0.2 mM ATP, 0.2 mM CaCI2, 0.5 mM dithiothreitol) wasmeasured at 220C with a Perkin-Elmer 650-10S Fluores-cence Spectrophotometer. Fluorescence excitation was at360 nm and emission was scanned from 360-560 nm(McClure and Edelman, 1966; Goldschmidt-Clermont et al.,1987). The signal obtained with TNS alone was comparedwith that obtained with TNS in the presence of PIP2 (10MM)and/or human platelet profilin (7 AM) and in separate assays,MgCI2 (10 mM) and/or PIP2 (10 M). MgCI2 was chosen be-cause it aggregates PIP2 into large multilammellar struc-tures, thus disrupting the micellar structure.
PLC assaysThese assays were run at 370C in VB with 80 AM CaCI2added for human platelet PLC and 20 MM CaCI2 for bovine
CELL REGULATION948

Specific interaction of profilin PIP2
brain PLC--y. PLC from human platelets was used at variousconcentrations (47-172 jg/ml) in experiments with micellesand at -78 ;g/ml in experiments with LUVETS. PLC--y frombovine brain was used at -4 ug/ml. Micelles of pure PIP2or LUVETS of mixed lipid composition were made using [3H]-PIP2 as the labeled substrate for PLC. The following com-positions and concentrations of LUVETS were used: PC:PE:PIP2 1:1:1 at 0.3 mg/ml total lipid, with 45 MM PIP2 in outerleaflet; PC:PE:PIP2 5:5:1 at 0.2 mg/ml total lipid, with 2.3UM PIP2 in outer leaflet; and PC:PIP2 (5:1) at 0.24 mg/mltotal lipid, with 18 MM PIP2 in outer leaflet. We assumedthat the PIP2 would distribute equally between inner andouter leaflets, except with the PC:PE:PIP2 (5:5:1) LUVETS,where we added PIP2 to the outer leaflet by incubating theLUVETS of PC:PE with micelles of PIP2 (Goldschmidt-Cler-mont et al., 1990). Micelles of pure PIP2 were used at variousconcentrations, as indicated in Figure 5. Hydrolysis wasstopped by addition of ice cold methanol/chloroform/HCIto separate the IP3 in the aqueous phase from lipids in theorganic phase (Dawson, 1965; Baldassare and Fisher, 1986;Raben, 1987). The [3H]-1P3 in the aqueous phase was mea-sured by liquid scintillation counting. Because the concen-tration of PIP2 was much lower than the Km of the enzyme(Goldschmidt-Clermont et al., 1990), the rate of hydrolysiswas directly proportional to the PiP2 concentration accordingto the Michaelis-Menton equation V = (Vmax[S])/(IS] + Kin).The effect of PIP2 sequestration by profilin on the hydrolysisrate can be calculated directly from the Michaelis-Mentonequation where [S] is replaced by 1/2{([Kd + PT - ST]2 + 4KdX ST)1/2 - [Kd + PT - ST] }, where PT is the total concentrationof profilin, ST is the total concentration of PIP2, and Kd isthe dissociation constant for the profilin:PIP2 complex. Eachpoint corresponds to one individual time course.
AcknowledgmentsThis work was supported by research grant GM-26338, anN.S.F. graduate fellowship to L.M.M., and an A.H.A. fellow-ship to P.J.G.C. We are grateful to Dr.'s R.E. Pagano andD.M. Raben for their helpful advice regarding lipids and toDr.'s S.G. Rhee and J.J. Baldassare for their gifts of phos-pholipases.
Received: July 18, 1990.Revised and accepted: September 10, 1990.
ReferencesAmpe, C., Vandekerchove, J., Brenner, S.L., Tobacman, L.,and Korn, E.D. (1985). The amino acid sequence of Acan-thamoeba profilin. J. Biol. Chem. 260, 834-840.
Ampe, C., Sato, M., Pollard, T., Vandekerckhove, J. (1988).The primary structure of the basic isoform of Acanthamoebaprofilin. Eur. J. Biochem. 170, 597-601.
Anderson, R.A., and Marchesi, V.T. (1985). Regulation ofthe association of membrane skeletal protein 4.1 with gly-cophorin by a polyphosphoinositide. Nature 318, 295-298.
Baldassare, J.J., and Fisher, G.J. (1986). Regulation of mem-brane-associated and cytosolic phospholipase C activitiesin human platelets by guanosine triphosphate. J. Biol. Chem.261, 11942-11944.
Baldassare, J.J., Henderson, P.A., and Fisher, G.J. (1989).Isolation and characterization of one soluble and two mem-brane-associated forms of phosphoinositide-specific phos-pholipase C from human platelets. Biochemistry 28, 6010-6016.
Barshop, B.A., Wrenn, R.F., and Frieden, C. (1983). Analysisof numerical methods for computer simulation of kineticprocesses: development of KINSIM-a flexible portablesystem. Analytical Biochem. 130, 134-145.
Bradford, M.M. (1976). A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248-254.
Carlsson, L., Nystrom, L.E., Sundkvist, I., Markey, F., andLindberg, U. (1977). Actin polymerizability is influenced byprofilin, a low molecular weight protein in non-muscle cells.J. Mol. Biol. 1 15, 465-483.
Cohen, P., Broekman, M.J., Verkley, A., Lisman, J.W.W.,Derksen, A. (1971). Quantification of human platelet inosi-tides and the influence of ionic environment on their incor-poration of orthophosphate 32P. J. Clin. Invest. 50, 762-772.
Cooper, J.A., Blum, J.D., Williams Jr., R.C., and Pollard, T.D.(1986). Purification and characterization of actophorin, anew 1 5,000-dalton actin-binding protein from Acanthamoebacastellanii. J. Cell Biol. 261, 477-485.
Dawson, R.M.C. (1965). 'Phosphatido-peptide'-like com-plexes formed by the interaction of calcium triphosphoino-sitide with protein. Biochem. J. 97, 134-138.
Dinubile, M.J., and Southwick, F.S. (1985). Effects of mac-rophage profilin on actin in the presence and absence ofacumentin and gelsolin. J. Biol. Chem. 260, 7402-7409.
Forscher, P. (1989). Calcium and polyphosphoinositidecontrol of cytoskeletal dynamics. Trends Neurosci. 12, 468-474.
Goldschmidt-Clermont, P.J., Williams, M.H., and Galbraith,R.M. (1987). Altered conformation of Gc (vitamin D binding-protein) upon complexing with cellular actin. Biochem. Bio-phys. Res. Commun. 146, 611-617.
Goldschmidt-Clermont, P.J., Machesky, L.M., Baldassare,J.J., and Pollard, T.D. (1990). The actin-binding protein profilinbinds to PIP2 and inhibits its hydrolysis by phospholipaseC. Science 247, 1575-1578.
Gordon, D.J., Eisenberg, E., and Korn, E.D. (1976). Char-acterization of cytoplasmic actin isolated from Acantham-oeba castellanii by a new method. J. Biol. Chem. 251, 4778-4786.
Giuiliano, K.A., Khatib, F.A., Hayden, S.M., Daoud, E.W.R.,Adams, M.E., Amorese, D.A., Bernstein, B.W., and Bamburg,J.R. (1988). Properties of purified actin depolymerizing factorfrom chick brain. Biochemistry 27, 8931-8938.
Hartwig, J.H., Chambers, K.A., Hopica, K.L., Kwiatkowski,D.J. (1989). Association of profilin with filament-free regionsof leukocyte and platelet membranes and reversible mem-brane binding during platelet activation. J. Cell Biol. 109,1571-1 579.
Hope, M.J., Bally, M.B., Webb, G., and Cullis, P.R. (1985).Production of large unilammellar vesicles by a rapid extru-sion procedure. Characterization of size distribution, trappedvolume and ability to maintain a membrane potential.Biochim. Biophys. Acta 812, 55-65.
Hummel, J.P., and Dreyer, W.J. (1962). Measurement ofprotein-binding phenomena by gel filtration. Biochim. Bio-phys. Acta 63, 530-532.
Janmey, P.A., and Stossel, T.P. (1987). Modulation of gelsolinfunction by phosphatidylinositol 4,5-bisphosphate. Nature325, 362-364.
Vol. 1, November 1990 949

L.M. Machesky et al.
Kaiser, D.A., Sato, M., Ebert, R.F., and Pollard, T.D. (1986).Purification and characterization of two isoforms of Acan-thamoeba profilin. J. Cell Biol. 102, 221-226.
Kaiser, D.A., Goldschmidt-Clermont, P.J., Levine, B.A., andPollard, T.D. (1989). Characterization of renatured profilinpurified by urea elution from poly-L-proline agarose columns.Cell Motil. Cytoskeleton 14, 251-262.King, C.E., Stephens, L.R., Hawkins, P.T., Guy, G.R., andMichell, R.H. (1987). Multiple metabolic pools of phospho-inositides and phosphatidate in human erythrocytes incu-bated in a medium that permits rapid transmembrane ex-change of phosphate. Biochem. J. 244, 209-217.Koreh, K., and Monaco, M.E. (1986). The relationship ofhormone-sensitive and hormone-insensitive phosphatidyl-inositol to phosphatidylinositol 4,5-bisphosphate in the WRK-1 cell. J. Biol. Chem. 261, 88-91.Kwiatkowski, D.J., and Bruns, G.A. (1988). Human profilin.Molecular cloning, sequence comparison and chromosomeanalysis. J. Biol. Chem. 263, 5910-5915.Langner, M., Cafiso, D., Marcelja, S., and McLaughlin, S.(1990). Electrostatics of phosphoinositide bilayer mem-branes. Biophys. J. 57, 335-349.Lassing, I., and Lindberg, U. (1985). Specific interaction be-tween phosphatidylinositol 4,5 bisphosphate and profilactin.Nature 314, 472.
Lassing, I., and Lindberg, U. (1988). Specificity of the inter-action between PIP2 and the profilin:actin complex. J. CellBiochem. 37, 255.
Lind, S.E., Janmey, P.A., Chaponnier, C., Herbert, T.J., andStossel, T.P. (1987). Reversible binding of actin to gelsolinand profilin in human platelet extracts. J. Cell Biol. 105, 833-842.
Mabuchi, I. (1983). An actin-depolymerizing protein (depac-tin) from starfish oocytes: properties and interaction withactin. J. Cell Biol. 97, 1612-1621.
Mayer, L.D., Hope, M.J., and Cullis, P.R. (1986). Vesicles ofvariable sizes produced by a rapid extrusion procedure.Biochim. Biophys. Acta 858, 161-168.
McClure, W.O., and Edelman, G.M. (1966). Fluorescentprobes for conformational states of proteins. I. Mechanismof fluorescence of 2-p-Toluidinylnapthalene-6-sulfonate, ahydrophobic probe. Biochemistry 6, 1908-1918.
McIntyre, J., and Jenkins, C.R. (1969). Chemotaxis in thefree living amoebae (Hartmannela rhysodes). Aust. J. Exp.Med. Sci. 47, 625-632.
Mockrin, S.C., and Korn, E.D. (1980). Acanthamoeba profilininteracts with G-actin to increase the exchange of actinbound adenosine 5'-triphosphate. Biochemistry 19, 5359-5362.
Nishida, E. (1985). Opposite effects of cofilin and profilinfrom porcine brain on rate of exchange of actin-boundadenosine 5'-triphosphate. Biochemistry 24, 1160-1164.
Nystrom, L.E., Lindberg, U., Kendrick-Jones, J., and Jakes,R. (1979). The amino acid sequence of profilin from calfspleen. FEBS Lett. 101, 161-165.Oechsner, U., Magdolen, V., and Bandlow, W. (1987). ThecDNA and deduced amino acid sequence of profilin fromSaccharomyces cerevisiae. Nucleic Acids Res. 15, 9078.Corrigendum (1988). Nucleic Acids Res. 16, 1232.Pollard, T.D., and Cooper, J.A. (1984). Quantitative analysisof the effect of Acanthamoeba profilin on actin filament nu-cleation and elongation. Biochemistry 23, 6631-6641.Pollard, T.D., and Cooper, J.A. (1986). Actin and actin-bindingproteins. A critical evaluation of mechanisms and function.Annu. Rev. Biochem. 55, 987-1035.Raben, D.M., Yasuda, K., and Cunningham, D.D. (1987).Modulation of thrombin-stimulated lipid responses in cul-tured fibroblasts. Evidence for two coupling mechanisms.Biochemistry 26, 2759-2765.Reichstein, E., and Korn, E.D. (1979). Acanthamoeba profilin(a protein of low molecular weight from Acanthamoeba thatinhibits actin nucleation). J. Biol. Chem. 254, 6174-6179.Safer, D., Golla, R., and Nachmias, V. (1990). Isolation of a5-kilodalton actin-sequestering peptide from human bloodplatelets. Proc. Natl. Acad. Sci. USA 87, 2536-2540.Sung, H.R., Suh, P.G., Cho, K.S., Lee, K.Y., and Rhee, S.G.(1987). Bovine brain cytosol contains three immunogenicallydistinct forms of inositolphospholipid-specific phospholipaseC. Proc. Natl. Acad. Sci. USA 84, 6649-6653.Tilney, L.G., Bonder, E.M., Coluccio, L.M., Mooseker, M.S.(1983). Actin from Thyone briareus sperm assembles ononly 1 end of an actin filament: a behavior regulated by pro-filin. J. Cell Biol. 97, 112-142.Tobacman, L.S., and Korn, E.D. (1982). The regulation ofactin polymerization and the inhibition of monomeric actinATP-ase activity by Acanthamoeba profilin. J. Biol. Chem.257, 4166-4170.Toner, M., Vaio, G., McLaughlin, A., and McLaughlin, S.(1988). Adsorption of actions to phosphatidylinositol 4,5bisphosphate. Biochemistry 27, 7435-7443.Tseng, P.C.H., and Pollard, T.D. (1982). Mechanism of actionof Acanthamoeba profilin. Demonstration of actin speciesspecificity and regulation by micromolar concentrations ofMgCI2. J. Cell Biol. 94, 213-218.Tseng, P.C.H., Runge, M.S., Cooper, J.A., Williams, R.C., Jr.,and Pollard, T.D. (1984). Physical, immunochemical, andfunctional properties of Acanthamoeba profilin. J. Cell. Biol.98, 214-221.van Paridon, P.A., de Kruijff, B., Ouwerkerk, R., and Wirtz,K.W.A. (1986). Polyphosphoinositides undergo charge neu-tralization in the physiological pH range: a 3'P-NMR study.Biochim. Biophys. Acta 877, 216-219.Yonezawa, N., Nishida, E., lida, K., Yahara, I., and Sakai, H.(1990). Inhibition of the interactions of cofilin, destrin, anddeoxyribonuclease with actin by phosphoinositides. J. Biol.Chem. 265, 8382-8386.
CELL REGULATION950