the bovine papillomavirus distal "enhancer" is not cis essential for
TRANSCRIPT

Vol. 5, No. 11MOLECULAR AND CELLULAR BIOLOGY, Nov. 1985, p. 3310-33150270-7306/85/113310-06$02.00/0Copyright ©D 1985, American Society for Microbiology
The Bovine Papillomavirus Distal "Enhancer" Is Not cis Essentialfor Transformation or for Plasmid Maintenance
PETER M. HOWLEY,'* ELAINE T. SCHENBORN,2 ELSEBET LUND,2 JANET C. BYRNE,' ANDJAMES E. DAHLBERG2
Laboratory of Tumor Viruis Biology, National Cancer Institute, Bethesda, Maryland 20205,' and Department ofPhysiological Chemistry, University of Wisconsin-Madison, Madison, Wisconsin 537062
Received 17 June 1985/Accepted 19 August 1985
We constructed a mutant of bovine papillomavirus type 1 (BPV-1) DNA that lacked a transcriptionalenhancer located 3' to the polyadenylation site of the early viral RNAs expressed in transformed cells. Thismutant DNA, when separated from the procaryotic sequences, transforms mouse cells with an efficiencycomparable to that of the full BPV-1 genome, and it exists as a stable multicopy plasmid in transformed cells.The BPV-1 distal enhancer suppresses the effects of a cis-inhibitory element in pML2 sequences but is notessential for the expression of the viral genes involved in cellular transformation or plasmid maintenance.
Transcriptional enhancers are regulatory elements whichare able to increase the transcriptional activity of linkedgenes in eucaryotic cells (2, 21). This activation can occurover large distances and is largely position and orientationindependent (2, 9, 21, 27). Enhancers were first discoveredin the genomes of the DNA viruses simian virus 40 (SV40)(2, 3, 9, 12) and polyoma virus (6) and have subsequentlybeen shown to be important transcriptional regulatory com-ponents of other viral genomes as well as of cellular genes(see reference 14 for a review). Most enhancer elementshave been defined by their ability to activate a heterologouspromoter in a position- and orientation-independent manner,and these elements have been shown to be cis-essentialcomponents of the viral genomes or cellular genes in whichthey have been identified.A transcriptional enhancer element was first described in
the bovine papillomavirus type 1 (BPV-1) genome by Luskyet al. (16). When linked to the herpesvirus thymidine kinasegene, this enhancer increases the efficiency of stable trans-formation of a Rat 2 cell line from a Tk- to a Tk+ phenotype.In addition, this element can substitute for the SV40 72-base-pair repeat enhancer in a sensitive indirect assay for SV40large-T-antigen expression (16). This element, which werefer to as the distal enhancer, is located between bases 4390and 4451, approximately 200 base pairs 3' to thepolyadenylation site of a set of genes expressed in BPV-1-transformed mouse cells and in the nonproductively infectedcells of a bovine fibropapilloma (1, 8, 13). A 600-base-pairfragment containing this element functions as a transcrip-tional enhancer in stable transformation assays using theherpesvirus thymidine kinase gene (4) or the rat pre-proinsulin gene (N. Sarver, R. Muschel, J. C. Byrne, G.Khoury, and P. M. Howley, submitted for publication), andin transient expression studies with the herpesvirus thymi-dine kinase gene (26).
In the initial study of the BPV-1 distal enhancer, it wasargued that the expression of early genes of BPV-1 wasdependent upon this element, since deletion of this sequencefrom a chimeric BPV-1 pML2 plasmid resulted in the loss oftransform'ing activity (16). Reinsertion of the BPV-1 distalenhancer, the SV40 enhancer, or a polyomavirus enhancerfragment at a remote site restored transforming activity of
* Corresponding author.
the deleted genome. The procaryotic pML2 element in theseDNAs was pBR322 deleted of sequences that are cis inhib-itory to SV40 DNA replication in monkey cells (17) and totransformation of mouse cells by the complete BPV-1genome (18, 22). Despite the deletion of these sequences,pML2 DNA is not inert when linked to BPV-1 DNA se-quences. This point is best exemplified by the fact that thepML2 sequences are cis inhibitory for transformation ofmouse cells by the 69% subgenomic BamHI-HindIII frag-ment of BPV-1 DNA, which is by itself sufficient for trans-formation (7, 22).We were prompted to examine whether the BPV-1 distal
enhancer is cis essential for BPV-1 gene expression in theabsense of pML2 DNA sequences because of recent resultsfrom each of our laboratories. First, Schenborn et al. (24)described two chimeric DNAs which contain segments of ahuman Ul RNA gene linked to the BPV-1 transformingregion but which lack the distal enhancer. These DNAs caneach replicate as plasmids and induce morphologic transfor-mation in. mouse C127 cells, suggesting that either the distalenhancer is not cis essential for early viral gene expressionor that its function is provided by Ul RNA gene sequences.Second, Spalholz et al. (25) recently showed that the BPV-1noncoding region located 5' to the early open reading framescontains a transcriptional enhancer which is trans activatedby the putative viral E2 gene product. Although studies havenot yet established whether this noncoding region enhanceris itself cis'essential for viral gene expression, the presenceof this second enhancer in the BPV-1 genome suggests thatthe role of the BPV-1 distal enhancer should be furtherevaluated.To assess whether the distal enhancer is indeed a cis-
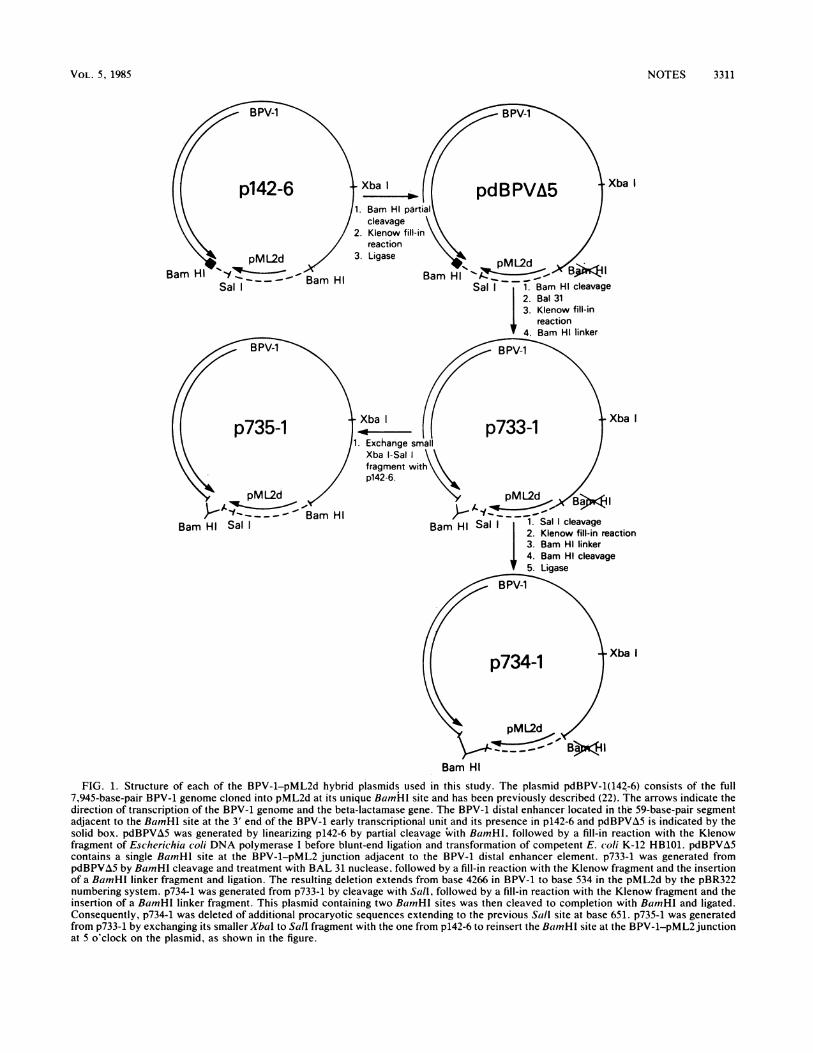
essential element for BPV-1 early gene expression, wedeleted the region containing it (see Fig. 1). The startingplasmid is pdBPV-1(142-6) which consists of the full BamHIlinear genome of BPV-1 DNA cloned into pML2d at itsunique BamHI site. This hybrid plasmid can efficientlytransform mouse C127 cells and can stably replicate in thesecells as well as in bacteria (22). The distal enhancer wasdeleted by BAL 31 nuclease treatment of pdBPVA5 afterBamHI cleavage (Fig. 1), and a BamHI linker fragment wasadded before ligation to generate p733-1. Sequence analysisof this mutant revealed that the deletion in BPV-1 DNAextended to base 4266 (Fig. 2), eliminating the distal en-
3310
NOTES 3311
Bam Hi
FIG. 1. Structure of each of the BPV-1-pML2d hybrid plasmids used in this study. The plasmid pdBPV-1(142-6) consists of the full7,945-base-pair BPV-1 genome cloned into pML2d at its unique BamHI site and has been previously described (22). The arrows indicate thedirection of transcription of the BPV-1 genome and the beta-lactamase gene. The BPV-1 distal enhancer located in the 59-base-pair segmentadjacent to the BamHI site at the 3' end of the BPV-1 early transcriptional unit and its presence in p142-6 and pdBPVA5 is indicated by thesolid box. pdBPVA5 was generated by linearizing p142-6 by partial cleavage with BamnHI, followed by a fill-in reaction with the Klenowfragment of Escherichia coli DNA polymerase I before blunt-end ligation and transformation of competent E. coli K-12 HB101. pdBPVA5contains a single BamHl site at the BPV-1-pML2 junction adjacent to the BPV-1 distal enhancer element. p733-1 was generated frompdBPVA5 by BamnHl cleavage and treatment with BAL 31 nuclease, followed by a fill-in reaction with the Klenow fragment and the insertionof a BamHI linker fragment and ligation. The resulting deletion extends from base 4266 in BPV-1 to base 534 in the pML2d by the pBR322numbering system. p734-1 was generated from p733-1 by cleavage with Sall, followed by a fill-in reaction with the Klenow fragment and theinsertion of a BamHI linker fragment. This plasmid containing two BamnHI sites was then cleaved to completion with BamHI and ligated.Consequently, p734-1 was deleted of additional procaryotic sequences extending to the previous Sall site at base 651. p735-1 was generatedfrom p733-1 by exchanging its smaller Xbal to Sall fragment with the one from p142-6 to reinsert the BamnHI site at the BPV-1-pML2 junctionat 5 o'clock on the plasmid, as shown in the figure.
VOL. 5, 1985

MOL. CELL. BIOL.
hancer but retaining the BPV-1 early polyadenylation signalAATAAA at base 4181 (5) and the polyadenylation site atbase 4203 (29). A decamer BamHI linker fragment separatesthe BPV-1 DNA (base 4266) from the pML2 DNA segment(base 534, by the pBR322 numbering system) (Fig. 2). Topermit separation of BPV-1 DNA from the pML2 sequences,the BamHI site at 5 o'clock was reconstituted in the distalenhancer deletion mutant to generate p735-1 (Fig. 1). A moreextensive procaryotic plasmid deletion (in p734-1) was alsointroduced which resulted in the joining of the BPV-1genome at base 4266 to the pML2 sequence at the formerSalI site at base 651 (Fig. 1).The B3PV-1 DNA segment from which the distal enhancer
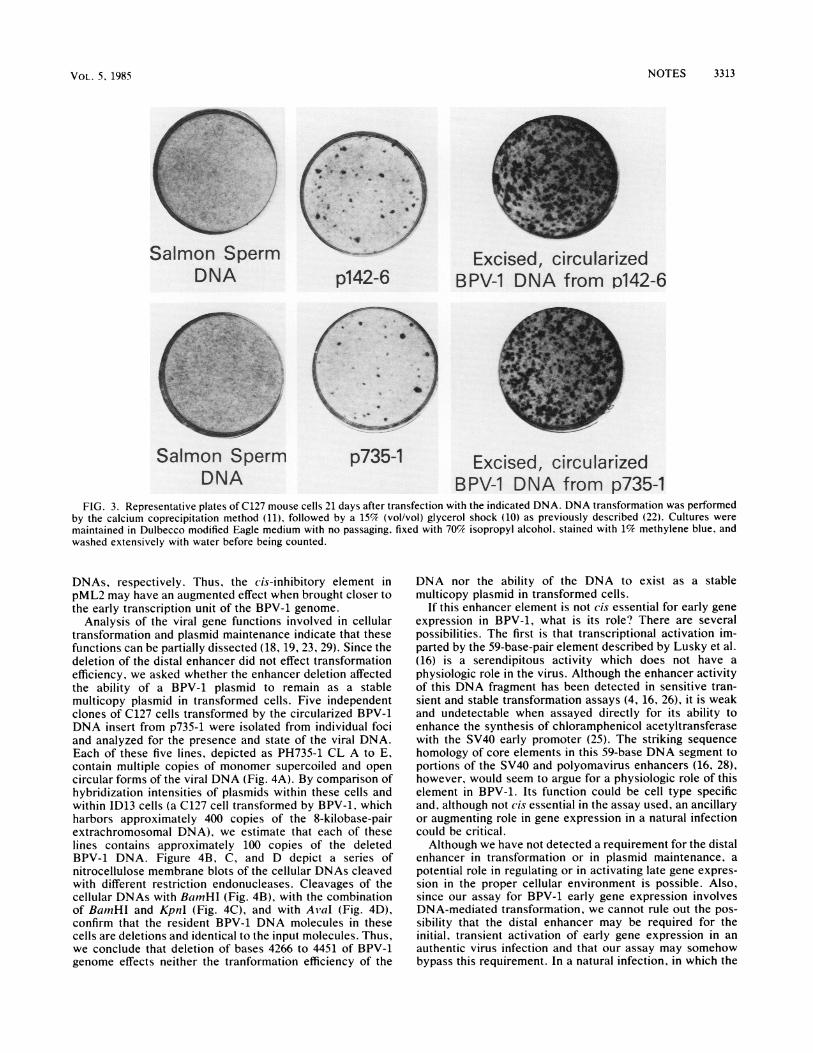
is deleted can be released from the pML2 procaryoticsequences by digestion of p735-1 with BamHI. The BPV-1DNA insert was gel purified and circularized with T4 DNAligase at a dilute DNA concentration and assayed for trans-formation of C127 cells. The BPV-1 insert from p735-1transformed these cells at approximately the same efficiencyas the full BPV-1 DNA insert from p142-6, with or withoutcircularization (Table 1; Fig. 3). Furthermore, the time ofappearance arid the morphology of the transformants werethe same for these two DNAs. We therefore conclude thatthe distal enhancer is not cis essential for expression of theBPV-1 transforming functions.
This conclusion is contrary to that of Lusky et al. (16),who assayed a deletion mutant (Bal 2) in which the BPV-1
G G+A T+C C
BPV-1
OG
TACGC
TA \ .TA \CG \ - -ATCG \ '
..CG(4266)CG _CGGCGCAT
TACGCGGCGCCG(534)CGGO
GCGCGCGCAT I #
PML2
FIG. 2. Sequence analysis of BPV-1-pML2 junction in p733-1after treatment with BAL 31 nuclease. The DNA was 5' end labeledat the Sall site, and the 900-base-pair Sall to Kpnl fragment wasisolated and sequenced by the technique of Maxam and Gilbert (20).The BPV-1 DNA junction is at base 4266, followed by a singleBainHI decamer linker, and the pBR322 sequence junction is at batse534.
TABLE 1. Transformation of mouse C127 cells by cloned BPV-1DNAs
No. ofDNA foci per
plate"
p142-6" .......................... 72, 58BPV-1 insert from p142-6, linear ..........................221, 248BPV-1 insert from p142-6, circular'.........................291, 217p735-1............................40, 36BPV-1 insert from p735-1, linear ..........................182, 186BPV-1 insert from p735-1, circular .........................232, 300Salmon sperm only .......................... 0, 0p733 lb .......................... 33, 31p734-lb .......................... 6, 5Bal 2d.......................... 2, 0
" Each plate received 0.5 ml of a DNA precipitate containing 1 ,ug of theplasmid DNA or modified insert DNA in the presence of high-molecular-weight salmon sperm DNA carrier (20 FJg/ml). Results are for duplicate plates.
I Circular DNA containing pML2 sequences as shown in Fig. 1.'The BarnHI-cleaved BPV-1 DNA linear fragments were gel purified and
ligated into covalently closed monomer circular forms with T4 ligase underdilute ligation conditions.dThis Bal 2 mutant has been described by Lusky et al. (16) and was
generously provided by M. Lusky and M. Botchan, University of California,Berkeley.
DNA was linked to pML2 sequences. The Bal 2 mutant hadbeen constructed by linearizing a plasmid identical to p142-6at its unique Sall site (see Fig. 1) followed by treatment withBAL 31 exonuclease and blunt end ligation (16). The dele-tion resulted in the removal of approximately 100 bases ofthe BPV-1 genome from the BamHI site, including the distalenh4ncer, and would be expected to join the BPV-1 DNA topML2 sequences at approximnately base 1025 by the pBR322numbering system.The discrepancy between the results of our study and that
of Lusky et al. (16) could be explained if the linked pML2sequences were cis inhibitory to the BPV-1 genome deletedof the distal enhancer. Each of the pML2d-linked but en-hancer-deleted DNAs (i.e., p733-1, p734-1, and p735-1) was,therefore, tested for its ability to transform C127 cells. TheBal 2 mutant described previo6sly (16) was also tested and,in agreement with the published results, was significantlyimnpaired in its ability to transformnmouse C127 cells (Table1). Likewise, transformation by each of the distal enhancer-deleted BPV-1 DNAs cloned in pML2 was impaired. Nev-ertheless, with various efficiencies, they all were able toinduce some transformed foci, indicating that there is noabsolute requirement for the distal (nhancer for the expres-sion of the BPV-1 transforming gen ' even in the presence ofthe procaryotic sequences. The IIB 2 plasmid was about60-fold less efficient than p142-6 in the focus assay (Table 1);however, the p733-1 and p735-1 intact plastnids were onlytwofold less efficient in transformation than the full BPV-1genome clone, despite the fact that the deletion in BPV-1sequences in p733-1 and p735-1 is more extensive.The degree to which the cis-inhibitory sequences in pML2
decrease the efficiency of transformation by these distalenhancer deletion mutants appears to be a function of theproximity of these sequences to the BPV-1 genome. Bal 2DNA, which is the least efficient in transformation (about1%, compared with p142-6 or p735-1 DNAs), has the largestdeletion of procaryotic sequences, with the BPV-1-pML2junction at approximately base 1025 (in pBR322). Plasmidp734-1 and p733-1, with smaller deletions of procaryoticDNA (junctions at bases 651 and 534 of pBR322, respec-tively) are 10 and 50% as efficient as p142-6 and p735-1
3312 NOTES
NOTES 3313
Salmon SpermDNA
*; ' V
p142-6
Excised, circularizedBPV-1 DNA from p142-6
Salmon SpermDNA
p735-1 Excised, circularizedBPV-1 DNA from p735-1
FIG. 3. Representative plates of C127 mouse cells 21 days after transfection with the indicated DNA. DNA transformation was performedby the calcium coprecipitation method (11). followed by a 15% (vol/vol) glycerol shock (10) as previously described (22). Cultures weremaintained in Dulbecco modified Eagle medium with no passaging. fixed with 70% isopropyl alcohol, stained with 1% methylene blue, andwashed extensively with water before being counted.
DNAs, respectively. Thus, the c is-inhibitory element inpML2 may have an augmented effect when brought closer tothe early transcription unit of the BPV-1 genome.
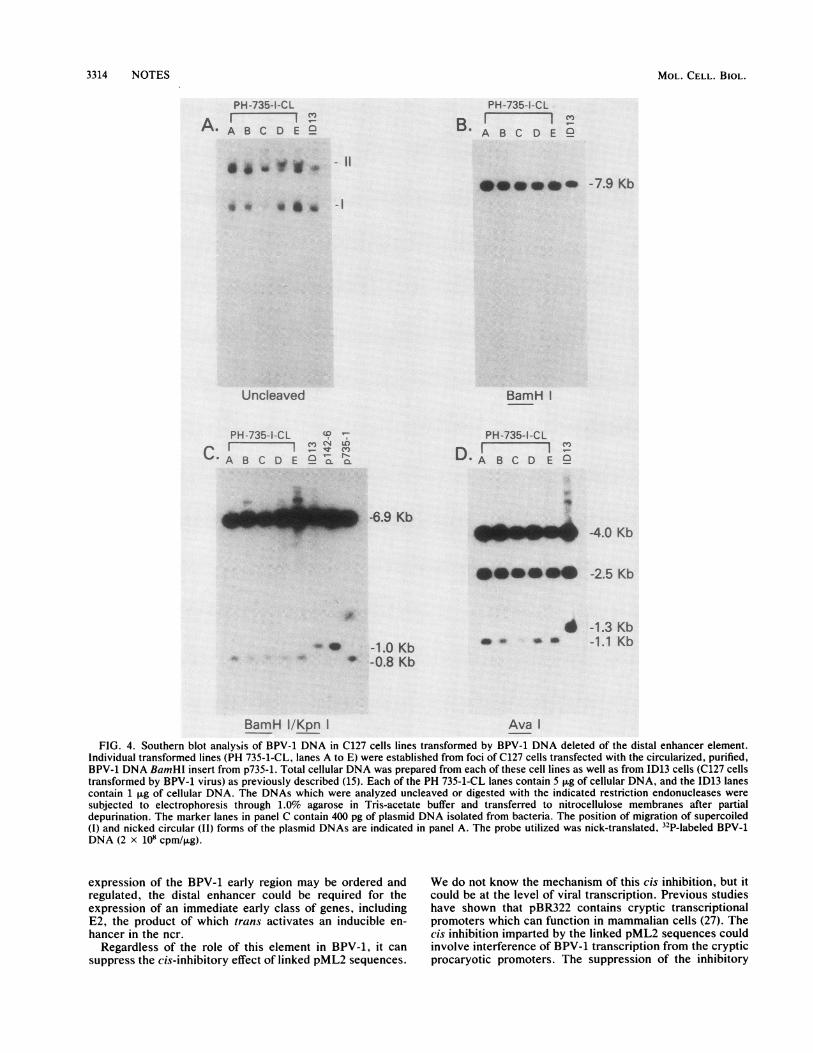
Analysis of the viral gene functions involved in cellulartransformation and plasmid maintenance indicate that thesefunctions can be partially dissected (18, 19, 23, 29). Since thedeletion of the distal enhancer did not effect transformationefficiency, we asked whether the enhancer deletion affectedthe ability of a BPV-1 plasmid to remain as a stablemulticopy plasmid in transformed cells. Five independentclones of C127 cells transformed by the circularized BPV-1DNA insert from p735-1 were isolated from individual fociand analyzed for the presence and state of the viral DNA.Each of these five lines, depicted as PH735-1 CL A to E,contain multiple copies of monomer supercoiled and opencircular forms of the viral DNA (Fig. 4A). By comparison ofhybridization intensities of plasmids within these cells andwithin ID13 cells (a C127 cell transformed by BPV-1. whichharbors approximately 400 copies of the 8-kilobase-pairextrachromosomal DNA), we estimate that each of theselines contains approximately 100 copies of the deletedBPV-1 DNA. Figure 4B, C, and D depict a series ofnitrocellulose membrane blots of the cellular DNAs cleavedwith different restriction endonucleases. Cleavages of thecellular DNAs with BamHI (Fig. 4B), with the combinationof BainHI and KpnI (Fig. 4C), and with AvsaI (Fig. 4D),confirm that the resident BPV-1 DNA molecules in thesecells are deletions and identical to the input molecules. Thus,we conclude that deletion of bases 4266 to 4451 of BPV-1genome effects neither the tranformation efficiency of the
DNA nor the ability of the DNA to exist as a stablemulticopy plasmid in transformed cells.
If this enhancer element is not cis essential for early geneexpression in BPV-1, what is its role? There are severalpossibilities. The first is that transcriptional activation im-parted by the 59-base-pair element described by Lusky et al.(16) is a serendipitous activity which does not have aphysiologic role in the virus. Although the enhancer activityof this DNA fragment has been detected in sensitive tran-sient and stable transformation assays (4, 16, 26), it is weakand undetectable when assayed directly for its ability toenhance the synthesis of chloramphenicol acetyltransferasewith the SV40 early promoter (25). The striking sequencehomology of core elements in this 59-base DNA segment toportions of the SV40 and polyomavirus enhancers (16, 28),however, would seem to argue for a physiologic role of thiselement in BPV-1. Its function could be cell type specificand, although not c is essential in the assay used, an ancillaryor augmenting role in gene expression in a natural infectioncould be critical.
Although we have not detected a requirement for the distalenhancer in transformation or in plasmid maintenance, apotential role in regulating or in activating late gene expres-sion in the proper cellular environment is possible. Also,since our assay for BPV-1 early gene expression involvesDNA-mediated transformation, we cannot rule out the pos-sibility that the distal enhancer may be required for theinitial, transient activation of early gene expression in anauthentic virus infection and that our assay may somehowbypass this requirement. In a natural infection, in which the
VOL. 5, 1985
MOL. CELL. BIOL.
PH -735-1-C Lr-m~A B C D E 9
isis ##ifI - 11
0 0 41 16 a -I
Uncleaved
PH-735-l-CL I9C I
r-BC Dn*A BOCD E 9'S. a
PH -735-1-CL
B. IA B C D E 9
*[email protected] -7.9 Kb
BamH I
PH-735-1 -CL
DI.T*A B C D ECa)0
-6.9 Kb
S
- e -1.0 Kbo -0.8 Kb
*
4.0 Kb
*- - -@ -2.5 Kb
* -1.3 Kbe. * * -1.1 Kb
BamH I/Kpn Ava I
FIG. 4. Southern blot analysis of BPV-1 DNA in C127 cells lines transformed by BPV-1 DNA deleted of the distal enhancer element.Individual transformed lines (PH 735-1-CL, lanes A to E) were established from foci of C127 cells transfected with the circularized, purified,BPV-1 DNA BamHl insert from p735-1. Total cellular DNA was prepared from each of these cell lines as well as from 1D13 cells (C127 cellstransformed by BPV-1 virus) as previously described (15). Each of the PH 735-1-CL lanes contain 5 pug of cellular DNA, and the ID13 lanescontain 1 p.g of cellular DNA. The DNAs which were analyzed uncleaved or digested with the indicated restriction endonucleases weresubjected to electrophoresis through 1.0% agarose in Tris-acetate buffer and transferred to nitrocellulose membranes after partialdepurination. The marker lanes in panel C contain 400 pg of plasmid DNA isolated from bacteria. The position of migration of supercoiled(I) and nicked circular (11) forms of the plasmid DNAs are indicated in panel A. The probe utilized was nick-translated, 32P-labeled BPV-1DNA (2 x 1io cpm/,ug).
expression of the BPV-1 early region may be ordered andregulated, the distal enhancer could be required for theexpression of an immediate early class of genes, includingE2, the product of which trans activates an inducible en-hancer in the ncr.
Regardless of the role of this element in BPV-1, it cansuppress the cis-inhibitory effect of linked pML2 sequences.
We do not know the mechanism of this cis inhibition, but itcould be at the level of viral transcription. Previous studieshave shown that pBR322 contains cryptic transcriptionalpromoters which can function in mammalian cells (27). Thecis inhibition imparted by the linked pML2 sequences couldinvolve interference of BPV-1 transcription from the crypticprocaryotic promoters. The suppression of the inhibitory
3314 NOTES

NOTES 3315
effect by the BPV-1 distal enhancer might, therefore, be dueto an augmentation of transcription of the BPV-1 transform-ing region.
We are grateful to George Khoury for critical comments concern-
ing this manuscript and to Nan Freas for its preparation.This work was supported in part by National Science Foundation
grant PCM 83-09618 and Public Health Service grant GM 30220 fromthe National Institutes of Health to J.E.D. and E.L. and by NationalResearch Service Award 5-T32-CA-09075 and Damon Runyon-Walter Winchell Cancer Fund Fellowship DRG-715 to E.T.S.
LITERATURE CITED1. Amtmann, E., and G. Sauer. 1982. Bovine papillomavirus
transcription: Polyadenylated RNA species and assessment ofthe direction of transcription. J. Virol. 43:59-66.
2. Banerji, J., S. Rusconi, and W. Schaffner. 1981. Expression of a
beta-globin gene is enhanced by remote SV40 DNA sequences.Cell 27:299-308.
3. Benoist, C., and P. Cambon. 1981. In i'i'o sequence require-ments of the SV40 early promoter region. Nature (London)290:304-310.
4. Campo, M. S., D. A. Spandidos, J. Lang, and N. M. Wilkie.1983. Transcriptional control signals in the genome of bovinepapillomavirus type 1. Nature (London) 303:77-80.
5. Chen, E. Y., P. M. Howley, A. D. Levinson, and P. H. Seeburg.1982. The primary structure and genetic organization of thebovine papillomavirus (BPV) type 1. Nature (London)299:529-534.
6. de Villiers, J., L. Olson, C. Tyndall, and W. Schaffner. 1982.Transcriptional "enhanceres" from SV40 and polyoma virusshow a cell type preference. Nucleic Acids Res. 10:7965-7976.
7. DiMaio, D., R. Treisman, and T. Maniatis. 1982. A bovinepapillomavirus vector which propagates as an episome in bothmouse and bacterial cells. Proc. NatI. Acad. Sci. USA79:4030-4034.
8. Engel, L. W., C. A. Heilman, and P. M. Howley. 1983. Tran-scriptional organization of the bovine papillomavirus type 1. J.Virol. 47:516-528.
9. Fromm, M., and P. Berg. 1982. Deletion mapping of DNAregions required for SV40 early region promoter function invivo. J. Mol. Genet. 1:457-481.
10. Frost, E., and J. Williams. 1978. Mapping-temperature sensitiveand host-range mutations of adenovirus type 5 by markerrescue. Virology 91:39-50.
11. Graham, F. L., and A. J. van der Eb. 1973. A new technique forthe assay of infectivity of human adenovirus 5 DNA. Virology52:456-461.
12. Gruss, P., R. Dhar, and G. Khoury. 1981. Simian virus 40tandem repeated sequences as an element of the early promoter.Proc. Natl. Acad. Sci. USA 78:943-947.
13. Heilman, C. A., L. Engel, D. R. Lowy, and P. M. Howley. 1982.Virus specific transcription in bovine papillomavirus trans-
formed mouse cells. Virology 119:22-34.14. Khoury, G., and P. Gruss. 1983. Enhancer elements. Cell
33:313-314.15. Law, M.-F., D. R. Lowy, I. Dvoretzky, and P. M. Howley. 1981.
Mouse cells transformed by bovine papillomavirus contain onlyextrachromosomal viral DNA sequences. Proc. Natl. Acad. Sci.USA 78:2727-2731.
16. Lusky, M., L. Berg, H. Weiher, and M. Botchan. 1983. Bovinepapillomavirus contains an activator of gene expression at thedistal end of the early transcription unit. Mol. Cell. Biol.3:1108-1122.
17. Lusky, M., and M. Botchan. 1981. Inhibitory effect of specificpBR322 DNA sequences upon SV40 replication in simian cells.Nature (London) 293:79-81.
18. Lusky, M., and M. Botchan. 1984. Characterization of thebovine papilloma virus plasmid maintenance sequences. Cell36:391-401.
19. Lusky, M., and M. R. Botchan. 1985. Genetic analysis of thebovine papillomavirus type 1 trans-acting replication factors. J.Virol. 53:955-965.
20. Maxam, A., and W. Gilbert. 1977. A new method for sequencingDNA. Proc. Natl. Acad. Sci. USA 74:560-564.
21. Moreau, P., R. Hen, B. Wasylyk, R. Everett, M. P. Gaub, and P.Chambon. 1981. The SV40 72 base pair repeat has a strikingeffect on gene expression both in SV40 and other chimericrecombinants. Nucleic Acids Res. 9:6047-607.
22. Sarver, N., J. C. Byrne, and P. M. Howley. 1982. Transforma-tion and replication in mouse cells of a bovine papil-lomavirus/pML2 plasmid vector that can be rescued in bacteria.Proc. Natl. Acad. Sci. USA 79:7147-7151.
23. Sarver, N., M. S. Rabson, Y.-C. Yang, J. C. Byrne, and P. M.Howley. 1984. Localization and analysis of bovine papil-lomavirus type 1 transforming functions. J. Virol. 52:377-388.
24. Schenborn, E. T., E. Lund, J. L. Mitchen, and J. E. Dahlberg.1985. Expression of a human Ul RNA gene introduced intomouse cells via bovine papillomavirus DNA vectors. Mol. Cell.Biol. 5:1318-1326.
25. Spalholz, B. A., Y.-C. Yang, and P. M. Howley. 1985.Transactivation of a bovine papillomavirus transcriptional reg-ulatory element by the E2 gene product. Cell 42:183-191.
26. Spandidos, D. A., and N. M. Wilkie. 1983. Host-specificities ofpapillomavirus, Moloney murine sarcoma virus and simian virus40 enhancer sequences. EMBO J. 2:1193-1199.
27. Wasylyk, B., C. Wasylyk, P. Augereau, and P. Chambon. 1983.The SV40 72 bp repeat preferentially potentiates transcriptionstarting from proximal natural or substitute promoter elements.Cell 32:503-514.
28. Weiher, H., and M. R. Botchan. 1984. An enhancer sequencefrom bovine papillomavirus DNA consists of two essentialregions. Nucleic Acids Res. 12:2901-2916.
29. Yang, Y.-C., H. Okayama, and P. M. Howley. 1985. Bovinepapillomavirus contains multiple transforming genes. Proc.NatI. Acad. Sci. USA 82:1030-1034.
VOL. 5, 1985