the catholic university of america ectopic bone matrix
TRANSCRIPT

THE CATHOLIC UNIVERSITY OF AMERICA
Ectopic Bone Matrix Mineralization: Unveiling the
Osteoinductive Nature of Crab Cuticle
A DISSERTATION
Submitted to the Faculty of the
Department of Biomedical Engineering
School of Engineering
Of The Catholic University of America
In Partial Fulfillment of the Requirements
For the Degree
Doctor of Philosophy
By
Tiffany Suella Omokanwaye
Washington, D.C.
2014

Ectopic Bone Matrix Mineralization: Unveiling the
Osteoinductive Nature of Crab Cuticle
Tiffany Suella Omokanwaye, Ph.D.
Director: Otto C. Wilson, Jr., Ph.D.
Large bone defects do not heal spontaneously and often require substitute materials. Ideally, a
bone replacement material should mimic bone tissue from a mechanical, chemical, biological and
functional point of view, and facilitate new bone formation. No single existing synthetic material
possesses all the necessary properties required in an ideal bone implant. Using biomimetic
principles and the kinship among biologically derived hard tissues, crustacean exoskeleton
emerged as a natural material for bone implant because of its similarities to bone in composition,
structure, and function. The purpose of this work is to serve as a preliminary investigation of the
role in which crab shell, from Callinectes sapidus or Chesapeake blue claw crab, can play in
bone healing. Soft tissue implantation studies, in rats, were used to investigate the osteoinductive
potential of the crab cuticle.
Crushed crab cuticle was subcutaneously implanted in the abdominal region of 28-day-old
Sprague-Dawley rats and aged for time periods ranging from 1-30 days by our collaborators at
Howard University. Tissue samples which grew in in the region of the crushed crab shell implant
were harvested and processed for microscopy and transmission electron microscopy (TEM)
analysis. This work focuses on characterizing the crystalline nature and physical characteristics
of the mineral phase which formed in the implant samples. Fascinating structures and
architectures were observed in TEM mode --- collagen fibers with the characteristic 67 nm
banding pattern, collagen bundles, fibroblasts, dark regions of crystal-like particles, and 20 x 40

nm nanocrystals. X-ray microanalysis of 20 x 40 nm nanocrystals showed an average
calcium:phosphorus ratio of 1.81 ± 0.37. Selected area diffraction (SAD) was initially used to
determine the degree of crystallinity of mineral phases. Dark electron-dense regions found
around collagen produced diffraction patterns indicative of amorphous solids. Upon further
inspection using high resolution transmission electron microscopy (HRTEM), approximately 2-4
nm crystalline-nano-building-blocks with lattice spacings of 0.95 nm were revealed.
Nanodiffraction was employed to investigate these 2-4 nm nano-structures with lattice spacings
of 0.95 nm in more detail. Nanodiffraction clearly indicated the particle was a single crystal.
Both the END pattern of the crystalline-nano-building-blocks and the SAD pattern of the 20 x 40
nm nanocrystal were both indexed and found to be of the apatite family. Compellingly, the SAD
pattern of the 20 x 40 nm nanocrystals displayed speckled rings made up of discrete spots. This
suggest that there are many oriented single crystals and that the larger crystals are made up of an
assembly of smaller single crystals. This gives evidence for the mesocrystal model of
crystallization for biologically derived hydroxyapatite (HAP). Arguably, our study is the first of
its kind to find biologically produced HAP crystals approximately 2-4 nm in size with evidence
they assemble to make larger HAP crystals based on the mesocrystal model.

ii
This dissertation by Tiffany Suella Omokanwaye fulfills the dissertation requirement for the
doctoral degree in Biomedical Engineering approved by Otto C. Wilson, Jr., Ph.D. as Director,
and by Patrick Mehl, Ph.D., Isabel k. Lloyd, Ph.D., Pamela L. Tuma, Ph.D., and Victor Frenkel,
Ph.D. as Readers.
–––––––––––––––––––––––––––––––––
Otto C. Wilson, Jr., Ph.D., Director
–––––––––––––––––––––––––––––––––
Patrick Mehl, Ph.D., Reader
–––––––––––––––––––––––––––––––––
Isabel K. Lloyd, Ph.D., Reader
–––––––––––––––––––––––––––––––––
Pamela L. Tuma, Ph.D., Reader
–––––––––––––––––––––––––––––––––
Victor Frenkel, Ph.D., Reader

iii
Dedication
This dissertation is dedicated to the following people:
the loving memory of my mother and best friend, Sandra Gertrude Hamilton, who
provided me unmeasurable love and support and who emphasized the importance of
education;
my patient husband and other best friend, Olushina Omokanwaye, who was extremely
supportive of my work and who endured many challenges and sacrifices while
completing this dissertation;
my awesome children, Mayowa, Lolade, Ayomide, Ekundayo, and Oladipo, for their
wholehearted attempt to be understanding while their mother spent so much time working
on this dissertation.

iv
Table of Contents Preface.......................................................................................................................................... xiii
Acknowledgements ..................................................................................................................... xvii
Chapter 1 Research Paradigm for Crab Cuticle Osteoinductive Potential ..................................... 1
Hypothesis................................................................................................................................... 1
Methodology ............................................................................................................................... 1
Rationale for using crab cuticle for bone implant study ......................................................... 1
Hypothesis Testing.................................................................................................................. 5
My role .................................................................................................................................... 6
Limitations ............................................................................................................................ 10
Hypothesis Implications........................................................................................................ 13
Chapter 2 Literature Review: Bone and Crab Cuticle and TRIZ .................................................. 15
Introduction ............................................................................................................................... 15
Ideal Bone Implant .................................................................................................................... 18
Biomimetics .............................................................................................................................. 18
Bone and Crab Cuticle Functional Similarities using TRIZ ..................................................... 24
Transferring Biology to Engineering ........................................................................................ 29
General Comparison of Bone and Crab Cuticle ....................................................................... 34
Concluding Remarks ................................................................................................................. 35
Chapter 3 Ties that Bind: Evaluation of Collagen I and α-chitin ................................................ 37
Introduction ............................................................................................................................... 37
Framework of chitin and collagen ............................................................................................ 39
Chitin Forms ............................................................................................................................. 40
Collagen Forms ......................................................................................................................... 42
Chemistry .................................................................................................................................. 43
Polysaccharides and Proteins .................................................................................................... 43
Protein and Polysaccharide Interactions ................................................................................... 48
Proteoglycans ............................................................................................................................ 51
Hierarchy................................................................................................................................... 57
Liquid Crystal Characteristics................................................................................................... 59
Synthesis ................................................................................................................................... 74

v
Properties .................................................................................................................................. 80
Applications .............................................................................................................................. 85
Characterization of Collagen and Chitin................................................................................... 88
Materials and Methods .......................................................................................................... 88
Elemental Analysis ............................................................................................................... 88
Zeta Potential Analysis ......................................................................................................... 89
Thermal Analysis (TGA) ...................................................................................................... 89
Surface Morphology (SEM) ................................................................................................. 89
Results ....................................................................................................................................... 90
Chemical analysis ................................................................................................................. 90
Thermal Analysis (TGA) ...................................................................................................... 93
Zeta potential ........................................................................................................................ 97
Scanning Electron Microscopy (SEM) ................................................................................. 99
Conclusion .............................................................................................................................. 101
Chapter 4 Tools of Biomineralization: Calcium Carbonate and Calcium Phosphate ................. 103
Introduction ............................................................................................................................. 103
Definition ................................................................................................................................ 105
Mineral Function/Strength/ Properties .................................................................................... 107
Biomimetics and biomineralization ........................................................................................ 111
Biomineralization Models ....................................................................................................... 113
Biologically Induced ........................................................................................................... 114
Mesocrystals model ............................................................................................................ 119
Brick and mortar model ...................................................................................................... 122
Cellular Role ....................................................................................................................... 122
Matrix Vesicles (MV) ......................................................................................................... 124
Amorphous Mineral Component ........................................................................................ 128
Mg stabilized amorphous precursors .................................................................................. 130
Effect of lipids/proteoglygans/alkaline phosphates ............................................................ 131
Biopolymers ............................................................................................................................ 131
Stages of Crystallization ......................................................................................................... 133
Crystallization Control ............................................................................................................ 137
Concluding Remarks ............................................................................................................... 139

vi
Chapter 5 Osteoinductive Nature of Blue Claw Crab ................................................................. 141
Introduction ............................................................................................................................. 141
Bone Grafts and Bone ............................................................................................................. 144
Osteoinduction, Osteoconduction, and Osteogenic ................................................................ 149
Ossification ............................................................................................................................. 152
Calcification/Mineralization ................................................................................................... 153
Inflammation ........................................................................................................................... 155
Experimental Section .............................................................................................................. 158
Materials ............................................................................................................................. 158
Crab integument preparation............................................................................................... 158
Characterization .................................................................................................................. 159
In vivo crab shell implantation ........................................................................................... 161
Some Preliminary Results ................................................................................................... 164
Analysis............................................................................................................................... 168
Particle Size ........................................................................................................................ 182
Nanodiffraction ................................................................................................................... 183
Indexing the nanodiffraction pattern ................................................................................... 189
Mathematical Method for Hexagonal ................................................................................. 190
Possible mechanism ................................................................................................................ 212
Concluding Remarks ............................................................................................................... 216
Chapter 6 Final Remarks ............................................................................................................ 219
References ................................................................................................................................... 224

vii
List of Figures
Figure 1-1 Composition percentages of bone (Chai, et al., 2012). ................................................ 4
Figure 2-1 Making an Ideal Bone Graft (Stocker & Wolinetz, 2009). ........................................ 17
Figure 2-2 Expanded description of the field of biomimetics (Sarikaya, 1994). ......................... 20
Figure 2-3 Biological (right column) and Engineering (left column) materials are very different
in the way they are developed (Allen, 2010), (Fratzl, 2007). ....................................................... 22
Figure 2-4 This table depicts bone and crab cuticle system, contribution, level or organization,
medium, parameters & interactions, and resource. (Bottom) This diagram ewxhibits a closer
inspection of the functions of bone and crab cuticle, conflicts to accomplishing those functions,
and the tricks or circumventions around those conflicts. .............................................................. 26
Figure 2-5 A contradiction describes the predicament caused by an attempt to improve one
property of a system that induces the degradation of another property (Stanley, Zlotin,
Bolckmans, & Zusman, 2005). ..................................................................................................... 28
Figure 2-6 How TRIZ works (Stanley, Zlotin, Bolckmans, & Zusman, 2005). .......................... 29
Figure 2-7 Schematic of steps to identify a function/concept/idea and to transfer the function
into engineering solution (Vincent, 2008), (Shimomura, 2010). .................................................. 31
Figure 2-8 Bone and Crab Shell Similarities. .............................................................................. 33
Figure 3-1 Diagram showing the direction of the polymer chains in the three crystallographic
forms of chitin. (a) α-chitin. (b) β-chitin. (c) γ-chitin (Wainwright, 1982). ................................. 41
Figure 3-2 Chemical Structure of Collagen showing the glycine, proline, and hydroxyproline
residues (Brodsky, Werkmeister, & Ramshaw, 2005). ................................................................. 44
Figure 3-3 Fragment of chitin chain (Ehrlich, 2010). .................................................................. 45
Figure 3-4 Arrangement of the protein subunits around the chitin core in the microfibril
perpendicular to the fiber axis and along the fiber axis. The 61 helix of protein subunits repeats in
3.06nm (Blackwell & Weih, 1980). .............................................................................................. 50
Figure 3-5 Proteoglycan aggregate (Kierszenbaum, 2007). ........................................................ 52
Figure 3-6 Hierarchical levels of collagen I in bone and α-chitin-protein matrix in crab
exoskeleton. Depicting points of differences in the types of bonds and triple helix structure of
collagen and linear structure of chitin. .......................................................................................... 58

viii
Figure 3-7 Representation of a typical liquid crystal model. ....................................................... 62
Figure 3-8 Depiction of rod-like liquid crystal phases: smectic, nematic, blue phase, and
cholesteric or chiral nematic. ........................................................................................................ 64
Figure 3-9 Illustration of aspects of lyotropic liquid crsytals. ..................................................... 65
Figure 3-10 Liquid crystalline organization of collagen at different concentrations. .................. 70
Figure 3-11 Collagen and Chitin Metabolism (Cohen, 1993). .................................................... 76
Figure 3-12 Chitin and Collagen Synthesis (Laurent, 1987). ...................................................... 77
Figure 3-13 Collagen and chitin unique features and ones they share. ....................................... 84
Figure 3-14 Collagen and Chitin/Chitosan Tissue Engineering Applications (Monzack,
Rodriguez, McCoy, Gu, & Masters, 2011). .................................................................................. 86
Figure 3-15 Other Biomedical Applications. In the diagram some of the collagen applications
are highlighted in yellow (Prashanth & Tharanathan, 2007). ....................................................... 87
Figure 3-16 Thermogram of Collagen and Chitin. ...................................................................... 95
Figure 3-17 Zeta potential (ξ) of collagen. Collagen trend line has been taken from (Andrade,
Ferreira, & Domingues, 2004). ..................................................................................................... 98
Figure 3-18 Chitin from Crab Exoskeleton SEM Micrographs. ............................................. 100
Figure 3-19 Collagen from Bovine Achilles Tendon SEM Micrographs ............................... 101
Figure 4-1 The materials property chart (Dunlop & Fratzl, 2010). ........................................... 108
Figure 4-2 Biomineral crystallographic properties are highly regulated during biomineralization.
These properties provide some indication of potential processing strategies (Gower, 2008). ... 110
Figure 4-3 There exist three primary divisions of biomimetic materials chemistry inspired by
biomineralization studies (Mann, 1995). .................................................................................... 113
Figure 4-4 Schematic of the mineralized collagen fibrils, the basic constituents of bone.
Nanocrystals of HAP are incorporated between collagen molecules (Dorozhkin & Epple, 2002).
..................................................................................................................................................... 118
Figure 4-5 Illustrative representation of classical and non-classical crystallization (Xu, Ma, &
Colfen, 2007). ............................................................................................................................. 121

ix
Figure 4-6 An illustration of the role of inorganic and organic constituents in controlling
biomineral formation (Gower, 2008). ......................................................................................... 123
Figure 4-7 Diagram outlining a proposed mechanism for bone mineral formation
(Boonrungsiman, et al., 2012)..................................................................................................... 125
Figure 4-8 Hierarchical structural organization of bone: (a) cortical and cancellous bone; (b)
osteons with Haversian systems; (c) lamellae; (d) collagen fiber assemblies of collagen fibrils;
(e) bone mineral crystals, collagen molecules, and non-collagenous proteins (Rho, Kuhn-
Spearing, & Zioupos, 1998). ....................................................................................................... 136
Figure 5-1 Schematic of the design strategy of tissue-engineered biomimetic nanocomposite
bone graft (Murugan & Ramakrishna, 2007). ............................................................................. 146
Figure 5-2 Depiction of the past, present and future of tissue repair (Hench, 1998). ................ 147
Figure 5-3 Diagram illustrating hypothesized mechanisms behind osteoinduction by
biomaterials. Physico-chemical and/or structural properties of osteoinductive biomaterials may
prompt the mechanism responsible for heterotopic bone formation (Barradas, Yuan, van
Blitterswijk, & Habibovic, 2011)................................................................................................ 154
Figure 5-4 Schematic depicting the material, interface, and biological enviroment components.
It emphasizes the importance of the interface. ............................................................................ 156
Figure 5-5 The implantation study has an aim to assess the histological response to the crab
cuticle and determine its potential as as a bone biomaterial. ...................................................... 162
Figure 5-6 Transmission electron microscopy image of in vivo study of demineralized crushed
Calinectes Sapides (Blue Claw) crab shell implanted subcutaneously in abdominal region of 28
day old Sprague–Dawley rats. Aging time was one day and image displays a crab shell particle
in the bottom of the image with a macrophage in close proximity (Wilson, Gugssa, Mehl, &
Anderson, 2012). ......................................................................................................................... 164
Figure 5-7 TEM pictures depicts macrophages surrounding the Bouligand structure of
demineralized crushed Calinectes Sapides (Blue Claw) crab shell (Wilson, Gugssa, Mehl, &
Anderson, 2012). ......................................................................................................................... 165
Figure 5-8 Transmission electron microscopy image of collagen bundles that formed after
subcutaneous implantation of demineralized crab shell for 6 days. The image shows 67 nm
banding pattern and regions of electron dense mineralized particles on the collagen fiber surface
and interspersed in between collagen fibers (Wilson, Gugssa, Mehl, & Anderson, 2012). ....... 166
Figure 5-9 (a) SEM micrograph and a schematic drawing of a cross-sectional surface showing
three different layers in the crab exoskeleton: epicuticle, exocuticle, and endocuticle (Chen, Lin,
McKittrick, & Meyers, 2008); (b) TEM micrograph of demineralized crushed Calinectes Sapides

x
(Blue Claw) crab shell displaying prominent Bouligand nested arc patterns and a schematic
drawing of the Bouligand structure showing the arc pattern on oblique surface. ....................... 169
Figure 5-10 Transmission electron microscopy image of collagen bundles that formed after
subcutaneous implantation of mineralized crab shell for 28 days. Similar to the image of
(Wilson, Gugssa, Mehl, & Anderson, 2012), it depicts collagen fibers with the characteristic 67
nm banding pattern. .................................................................................................................... 170
Figure 5-11 Transmission electron microscopy image Collagenous fibers, collagenous bundles,
and fibroblasts. Fibers oriented in longitudinal directions exhibit the characteristic 67 nm
banding pattern of collagen. Lying between regions of many collagen bundles and fibers,
fibroblasts often present an elongated form. Fibrocytes are essentially mature fibroblasts.
Fibrocytes can easily be recognized as such by their dense nucleus, long and thin cytoplasmic
processes and scarcity of cell organelles (Ebe & Kobayashi, 1972). ......................................... 171
Figure 5-12 Transmission electron microscopy image depicting collagenous fibers, collagenous
bundles, and fibroblasts. Fibers oriented in longitudinal directions exhibit the characteristic 67
nm banding pattern of collagen. Lying between regions of many collagen bundles and fibers,
fibroblasts often present an elongated form. Fibrocytes are essentially mature fibroblasts.
Fibrocytes can easily be recognized as such by their dense nucleus, long and thin cytoplasmic
processes and scarcity of cell organelles (Ebe & Kobayashi, 1972). ......................................... 172
Figure 5-13 Micrograph depicting regions of selected area diffraction. This image contains
collagenous fibers, fibroblasts. ................................................................................................... 176
Figure 5-14 (a) Selected area diffraction (SAD) coincides with 0021 SAD region from figure
13. (b) ) Selected area diffraction (SAD) coincides with 0023 SAD region from figure 13. (c)
Selected area diffraction (SAD) coincides with 0025 SAD region from figure 13. (d) Selected
area diffraction (SAD) coincides with 0028 SAD region from figure 13. All of these SAD images
display broad or diffuse rings which indicate amorphous-like phase. ........................................ 177
Figure 5-15 (a) Transmission electron microscopy image of dark region adjacent to collagenase
matrix and (b) its SAD pattern which shows broad diffuse rings that are indicative of a
amorphous solid. ......................................................................................................................... 179
Figure 5-16 SAD pattern of dark region adjacent to collagenase matrix depicted in figure 5-15
which shows broad diffuse rings that are indicative of a amorphous solid. ............................... 180
Figure 5-17 HRTEM image of potential Carbonated HAP depicting the short range or local
order. ........................................................................................................................................... 184
Figure 5-18 HRTEM image of small-size particle. Determined that nanosized particles of
different size preserve stoichiometric HA-like crystal structure (Biggemann, Prado da Silva,
Rossi, & Ramirez, 2008). ............................................................................................................ 185

xi
Figure 5-19 HRTEM nanodiffraction image of potential Carbonated HAP depicting a single
crystal. ......................................................................................................................................... 188
Figure 5-20 TEM image of possible polycrystalline HAP with interplanar or lattice spacings 197
Figure 5-21 SAD pattern of a polycrystalline HAP ................................................................ 198
Figure 5-22 Elemental Analysis with Weight and Atomic Percentages of oxygen, phosphorus
and calcium. ................................................................................................................................ 203
Figure 5-23 Elemental Analysis with Weight and Atomic Percentages of oxygen, phosphorus
and calcium. ................................................................................................................................ 204
Figure 5-24 Schematic diagrams representing the HAP unit cell (Menéndez-Proupin, et al.,
2011) ........................................................................................................................................... 206
Figure 5-25 Schematic depiction of one hexagonal building unit of HAP (Li, et al., 2007). ..... 210
Figure 5-26 Black lines connect Ca(I) columns in hexagonal networks. Cyan and magenta
triangles connect staggered Ca(II) atoms (Xia, Lindahl, Lausmaa, & Engqvist, 2011), (Boanini,
Gazzano, & A, 2010). ................................................................................................................. 211
Figure 5-27 The HAP unit cell has symmetry and order. Table of average distances of atom shell
from calcium atom via x-ray and EXAFS (Harries, Huskins, & Hasnain, 1986). ...................... 211
Figure 5-28 Diagram for wound healing response to crushed crab carapace (Falanga, 2005),
(Salgado, et al., 2011). ................................................................................................................ 213

xii
List of Tables
Table 1-1 Some osteoinduction papers. Only first author listed. ............................................... 3
Table 1-2 Thickness of skin strata in rat, mice and humans (Godin & Touitou, 2007) .......... 12
Table 3-1 Principal Components of Crustaceans (Crab) Exoskeleton and Bones ................... 38
Table 3-2 Key Articles that Highlight Collagen and Chitin .................................................... 40
Table 3-3 Some Macromolecules (polymers) and their primary biological functions (Smith &
Wood, 1991). ................................................................................................................................ 45
Table 3-4 Molecular Weight of Chitin and Collagen .............................................................. 47
Table 3-5 Description of Cholesteric Geometries ................................................................... 71
Table 3-6 Collagen and chitin empirical formula, tissue distribution, microscopic appearance,
ultrastructure, synthesis site, interaction, and function. ................................................................ 82
Table 3-7 Elemental Composition of Collagen. ...................................................................... 93
Table 4-1 Examples of the Diversity of Biominerals (Gower, 2008). ................................... 106
Table 4-2 Characterization of Biologically Controlled Extra-, Inter-, and Intra-cellular
Mineralization ............................................................................................................................. 116
Table 4-3 Designation of layers in the crab cuticle .............................................................. 137
Table 5-1 The composition of the inorganic and organic phases of bone ............................ 145
Table 5-2 Term [A] calculation for various values of hk ..................................................... 193
Table 5-3 Calculation of l2 and l2/(c/a)2 ................................................................................ 194
Table 5-4 Evaluation of Peaks. ............................................................................................. 194
Table 5-5 Allowed hkl and calculated d spacings ................................................................ 195
Table 5-6 d spacing values calculated for crystalline harvested sample. a (Fleet, Liu, & King,
2004). b (Wilson, Elliot, & Dowker, 1999) ................................................................................. 201

xiii
Preface
Bone tissue engineering seeks to regenerate the lost or damaged tissue by making use of the
interactions between cells and biomaterials. Bone formation is a complex process on which
inorganic biomineral (calcium phosphate) precipitation seems to be associated, initially, with
matrix vesicles and subsequently with organic collagen I fibers (Anderson, 1989). For large bone
defect repair, bone formation far from the host bone bed should occur by osteoinduction, a kind
of bone formation that does not start directly from osteogenic cells. Osteoinductive biomaterials
provide a good environment for cells to form bone (Reis & Weiner, 2004). Therefore, one of the
main goals for bone tissue constructs, when implanted in vivo, is to promote osteoinduction.
Bone formation, in soft tissues where no osteogenic cells exists, gives the true indication of
osteoinduction. This type of bone formation is called ectopic bone formation, heterotopic bone
formation, or heterotopic ossification (Reis & Weiner, 2004).
Skeletal conditions are becoming an increasing health concern in the aging population
(Giordano, Sanginario, Ambrosio, Silvio, & Santin, 2006), and reconstruction of bone defects is
one of the major therapeutic goals in various clinical fields. Clinicians and physical, biological,
and material scientists and engineers strive to develop bone biomaterials with novel properties
that perform like natural bone. The composition and nanostructure are believed to influence
biomaterial and biological environment interactions. Some novel materials are first designed
through exploratory research; next their properties are determined; and then potential uses are
identified. This is typically known as discovery based product development. However, the
scientific world is progressing toward a more application based model where the existing needs
are assessed and a design is established to address specific needs. Despite the method,

xiv
biomaterial design is based on an understanding of the biological material being replaced, the
material used as the replacement, and the material/ biological interface. Biological systems such
as bone and crustacean integument have many length scales of fundamental and structural
significance. Bone and crustacean integument are both naturally derived nanocomposites that
share unique attributes which can be used as sources for many lessons. As lessons are revealed,
crab cuticle emerges as an inspiration for developing functionally advanced biomaterial implants
(FAB). In Chapter 1, biomimetics and design concepts of TRIZ (Theory of Inventive Problem
Solving) are discussed highlighting the functional and general similarities of bone and crab
integument.
Bone and crustacean cuticle are biological materials. The term biological material is very
common, because it includes organic phases, inorganic phases, and composites of both organic
(e.g., protein–protein, protein–polysaccharide) and inorganic (e.g., mineral–protein, mineral–
polysaccharide) phases with amazing diversity of forms, shapes, and dimensions. Bone is a
composite that consist of organic and inorganic phases, collagen and hydroxyapatite,
respectively. Crustacean cuticle is also a composite that consist of organic and inorganic phases,
chitin and calcium carbonate, respectively. Bone and arthropod cuticle possess astonishingly
similar principles in organization: the ability to self-assemble; production of fibrillar and fiber-
like structures with hierarchical organization from nano- up to macro- levels; the capability to
perform the role of scaffolds; and the capacity to serve as templates for biomineralization and
formation of the rigid skeletal structures (Ehrlich H. , 2010). Chapter 2 compares the chitin and
collagen, the organic phases of crab cuticle and bone tissue. Chapter 3 surveys calcium
carbonates and calcium phosphate, the inorganic phases of crab cuticle and bone tissue, as tools

xv
of biomineralization. Thus, chapters 1-3 provides an understanding of biomimetics, TRIZ, the
inorganic-organic phases of bone tissue and how this awareness can be utilized to develop FAB.
Chapter 4 focuses on a general biocompatibility study to determine crab cuticle’s suitability
as a bone substitute material. Natural bone uses osteogenic (progenitor osteoblasts or osteocytes)
cells; osteoconductivity (structural support system); and osteoinductivity (such as growth factors)
to produce a mineralized collagenase matrix that hardens (Stocker & Wolinetz, 2009). Ideally, a
bone replacement material should mimic bone tissue from a mechanical, chemical, biological
and functional point of view, and facilitate new bone formation. No single existing synthetic
material possesses all the necessary properties required in an ideal bone implant. Emerging as
valid therapy approach, bone tissue engineering is based on understanding hard tissue formation
and targets induction of new functional tissues. Using inspiration from Nature and exploiting the
kinship that exists among biologically derived hard tissues, crustacean exoskeleton emerged as a
natural material, similar to bone in composition, structure, and function. The crab carapace is a
candidate material for enhancing the healing, remodeling, and engineering of bone.
The basic premise is that the properties and components of crab cuticle, specifically, the blue
claw crustacean, Callinectes sapidus, found in the Chesapeake Bay, satisfy a number of criteria
to make it a successful bone substitute material. Chapter 4 describes the result of an implantation
study in which crab cuticle has been harvested from Sprague-Dawley rats and analyzed to
provide evidence of mineralized collagen matrix formation in an attempt to assess crab shell’s
osteoinductive nature, bioactivity and viability as a bone material substitute. Special emphasis is
given to the characterization techniques transmission electron microscopy (TEM), high-

xvi
resolution transmission electron microscopy (HRTEM), X-ray diffraction (XRD), electron
nanodiffraction (END), and electron microanalysis - energy-dispersive spectrometry (EDS).
The final chapter, the epilogue, offers concluding remarks and discusses future directions.
Drawing inspiration from natural materials and examining the process involved in bone
morphogenesis, this research examines the possibility that crab exoskeleton is an osteo-friendly
material with promising bone bioactivity. This dissertation aims to illuminate the osteoinductive
attributes of blue claw crab, Callinectes sapidus, cuticle and to provide the first general
biological performance profile about the suitability of crab cuticle as a bone biomaterial. This is
a novel contribution to the fields of biomimetics, biological material science, orthopedic
medicine, and bone tissue engineering.

xvii
Acknowledgements
Foremost, I would like to thank my God and Savior Jesus Christ for giving me the strength,
wisdom, and perseverance to complete this dissertation.
The completion of this dissertation would not have been possible without the love and support
of my entire family --- parents, aunties, uncles, cousins and friends. Thanks for all the prayers,
for all the words of encouragement, and for all the hours of babysitting. I am especially grateful
for my dad, Sam Hamilton, Jr., for assisting me in his own unique ways. I am particularly
thankful for my Aunties Sinobia Gilliard and Wilhelmena Middleton and Uncle Sam Middleton
for constantly checking on me, listening to me, and helping me in whatever ways I asked.
I thank my dissertation committee led by Dr. Otto C. Wilson, Jr. No words can express how
much I truly appreciate your guidance and direction throughout this process. I am very thankful
for your patience and for your belief in me. My readers, Dr. Mehl, Dr. Lloyd, Dr. Tuma and Dr.
Frenkel, I thank you for your invaluable feedback. I am also very appreciative for Dr. Kilic and
Dr. Namazi serving as secretary and chair.
A special thanks to Dr. Anderson and Dr. Gugssa of Howard University for their invaluable
collaboration. Thank you for the use of your laboratory, animals, supplies, and instrumentation. I
am truly indebted for all of your input, feedback, direction and most importantly time.
The authors wished to thank the Nanoscale Imaging, Spectroscopy, and Properties (NISP)
Laboratory at the Kim Engineering Building at the University of Maryland at College Park, Dr.
Lai and Dr. Shang with my data analysis. We wish to thank Dr. Lloyd for the use of Schimadzu
TGA-50The financial support was granted by NSF DMR Biomaterials Grant # DMR-0645675.

1
Chapter 1 Research Paradigm for Crab Cuticle Osteoinductive
Potential
Hypothesis
My initial research question focused on the issue of whether crab shell possess the ability to
induce bone formation in an osteoinductive manner. As my investigation progressed, my
hypothesis developed along the lines of identifying one of the key features in hard tissue
formation: Mineralization. My hypothesis can be stated as follows. Can the electron dense
regions containing nanophase material which form around the collagen-like fibers produced
after implantation of crushed crab cuticle be identified as a biomineral phase related to bone
formation? The rest of my work focused on this aspect.
Methodology
Rationale for using crab cuticle for bone implant study
Large bone defects do not heal spontaneously and often require substitute materials. Bone is
osteoconductive and osteoinductive with osteogenic cells and this combination produces a
collagen matrix that mineralizes and hardens. Ideally, a bone replacement material should mimic
bone tissue from a mechanical, chemical, biological and functional point of view, and facilitate new
bone formation. No single existing synthetic material possesses all the necessary properties required
in an ideal bone implant.
Using biomimetic concepts, TRIZ and insight from previous studies, crab cuticle surfaced as
contender for another xenograft solution for the healing of large bone defects. Biomimetic and TRIZ

2
help to illuminate crab cuticle similarities to bone in regard to composition, structure, and
function. A literature search unearthed a wealth of information that provided even more
inspiration and credence to the possibility of crab cuticle prospects in bone healing.
The ancient Mayan practice of using nacre as bone implant material (Westbroek & Marin,
1998) serve as one source of inspiration. The literature revealed the use of other marine
organisms for dental and bone implants (Demers, et al., 2002). Additionally, derivatives of crab
shell such as chitin/chitosan have been used in bone tissue engineering investigations. Urist’s
(1965) discovery that demineralized bone matrix induced new bone formation when implanted
intramuscularly certainly peaked our interest. Urist and Strates (1971) presented ‘osteoinduction’
to the scientific and medical communities. Reddi and Anderson (1976) further expounded on
Urist's work and provided a compelling explanation of the functional role of purified organic
bone matrices (Gruskin, Doll, Futrell, Schmitz, & Hollinger Jeffrey, 2012).
Natural bone uses osteogenic (progenitor osteoblasts or osteocytes) cells; osteoconductivity
(structural support system); and osteoinductivity (such as growth factors) to produce a mineralized
collagen matrix that hardens (Stocker & Wolinetz, 2009). Crab exoskeleton has many similarities
to bone, but it does not have osteogenic cells. Osteoconduction relates to the ability to support
bone growth on a structure and our crushed crab shell in our experiments was not going to serve
a scaffold. Osteoinduction relates to the ability to create an in vivo miceroenvironment that
promotes new bone formation. Therefore, osteoinduction is essential to new bone formation and
made it the logical test choice.
Bone formation, through osteoinduction, does not start directly from osteogenic cells and
bone formation at ectopic sites has been found after implantation (Huggins, 1931), (Bertelsen,

3
1944), (Urist & McLean, 1952). Urist continued his groundbreaking research on osteoinduction
and later with the assistance of Strates came to the conclusion that protein, in particular bone
morphogenetic protein (BMP), was involved in the cascade of chemotaxis, mitosis,
differentiation, and finally bone formation (Urist & Strates, 1971).
There have been various publication that have illustrated osteoinduction in many animals by
diverse calcium phosphate biomaterials. Some of these papers are listed below in Table 1-1:
Table 1-1 Some osteoinduction papers. Only first author listed.
Author Title Year
Yamasaki
Ripamonnti
Yang
Ripamonnti
Yuan
Habibovic
Ripamonnti
Heterotopic bone formation around porous
hydroxyapatite ceramics in the subcutis of dogs
The Induction of Bone in Osteogenic Composites of
Bone Matrix and Porous Hydroxyapatite Replicas:
An Experimental Study on the Baboon
Osteogenesis in extraskeletally implanted porous
calcium phosphate ceramics: variability among
different kinds of animals
Osteoinduction in Dorous hydroxyapatite implanted
in heterotopic sites of different animal models
Bone formation induced by calcium phosphate
ceramics in soft tissue of dogs: a comparative
study between porous a-TCP and b-TCP
Osteoinductive biomaterials – properties and
relevance in bone repair
The induction of bone formation by coral-derived
calcium carbonate/hydroxyapatite constructs
1990
1991
1996
1996
2001
2007
2009

4
It could be concluded that osteoinduction is a general phenomenon (Yuan & Groot, 2004).
In 1969, Winter and Simpson (1969) observed bone induction by a sponge consisting of
polyhydroxyethyl methacrylate (poly-HEMA) in the soft tissue of pigs. These researchers noted
that the implanted sponge became calcified prior to bone formation in both pigs and rats. Bone
induction by the polymeric sponge cannot be explained by Urist’s BMPs theory. Crab shell like
this synthetic sponge do not contain nor can they produce BMPs. Thus, this suggest calcification
might play a role in the process of osteoinduction (Habibovic & de Groot, 2007).
The mineral phase makes up approximately 60% bone, figure 1-1. Bone matrix calcification
or mineralization is a critical stage of bone formation. Coupled with probable role of
calcification in osteoinduction and mineral percentage of bone, the presence of mineral became
an integral part of my hypothesis.
Figure 1-1 Composition percentages of bone (Chai, et al., 2012).

5
The biomimetic principles, TRIZ analysis, and literature review, help to solidify the notion
that crab shell might have osteoinductive potentials worth exploring.
Hypothesis Testing
There was a need for a relatively simple model in which osteoinductive potential could be
tested in animals. Across species, numerous general cellular properties are common in all
organisms. With each animal model having unique advantages and disadvantages, no single
animal model would be appropriate for all purposes, nor can a model be dismissed as
inappropriate for all purposes (Pearce, Richards, Milz, Schneider, & Pearce, 2007). The rat
model was attractive and selected based on factors such as the size of the material to be
implanted; cost to acquire and care for animals; tolerance to captivity and surgery; ease of
handling and housing; low maintenance care; resistance to infection and disease; adequate
facilities and support staff; and an existing database of biological information for the species.
The rat model would serve as an approximation of the physiological human clinical condition.
The rat subcutaneous tissue implant site has proven to be a high-through-put, relatively low-cost
screening technique for testing the initial tissue response to new materials.
Bone formation by osteoinduction is initiated by soluble and insoluble signals that trigger a
complex cascade of molecular and cellular morphogenetic processes that ultimately leads to the
sculpture of precisely organized mineralized structures. Bone formation, in soft tissues where no
osteogenic cells exists, gives the true indication of osteoinduction (Yuan & Groot, 2004).
Therefore, a pilot study, using the rat subcutaneous tissue model, offered a rapid, cost-effective

6
means of indicating how crab shell interacts with a living system and how this interaction
produced the desired expectation of bone formation.
The implantation studies were conducted at Howard University with the assistance of Dr.
Gugssa under the guidance of Dr. Anderson. Approximately 48 rats were used to address the
question of crab shell influencing bone formation using an osteoinductive model involving
subcutaneous implantation of crushed crab shell. The focus of my study involved analyzing
tissue samples from the crushed crab cuticle implant region that had been aged for 28 days, n =
2. A detailed account of implantation study is provided in the Experimental Section of Chapter 4.
My role
To understand the ramifications of this harvested tissue, the knowledge was acquired about
the similarities between crab cuticle and bone and their constituent parts. My literature search
uncovered many shared characteristics of bone and crab cuticle. Biomimetics and TRIZ helped
illuminated and communicate these similarities.
To get a better understanding of bone and crab shell, their compositions were explored and
some properties were characterized. I performed scanning electron microscopy (SEM), zeta
potential, and thermogravimetric analysis of the organic constituents of bone and crab cuticle.
Carbon, hydrogen, and nitrogen (CHN) analysis were performed by an outside lab, Prevalere
Life Sciences. SEM furnished information about the sample’s surface topography and
composition. Zeta potential analysis was used to ascertain the stability of colloidal dispersions of

7
chitin. TGA were done to determine the thermal decomposition and degradation behavior of our
samples. The CHN analysis provided information such as the purity of our sample.
A technician, Dr. Lai, assisted me with the preparation of samples and operated the SEM
during my sessions at the NISP Laboratory at the Kim Engineering Building at the University of
Maryland at College Park analysis. I communicated the magnification and the areas on the
sample that I wanted to observe. Dr Lai took and saved micrographs I wanted to analyze later.
Some of these images and analysis are reported in Chapter 2.
The zeta potential measurements were performed at Catholic University in Dr. Wilson’s
B.O.N.E. C.R.A.B lab. pH was varied systematically from about 2 to 12 in order to determine
how it influences the ζ-potential of chitin. The data was plotted and the analysis is reported in
Chapter 2.
After training, I performed TGA analysis at Dr. Lloyd’s lab at University of Maryland at
College Park. The data was plotted and the analysis is reported in Chapter 2.
Transmission electron microscopy (TEM) offered broad range of characterization techniques
with high spatial and analytical resolution that could be done on the same small sample. I was
charged/commissioned to create and analysis TEM grids samples from the harvested rat tissue in
the region of crushed crab shell implantation.
I participated in cutting some of the 90 nm samples at Howard University after training was
provided by Dr. Gugssa. The main task of my work was to evaluate the samples for
mineralization. Using a Leica Microsystems Instrument outfitted with a diamond blade, at a
speed of 3-4 mm/s and an approach of 0.5 µm, 490 nm samples were cut. 6-8 490 nm samples
were placed on a slide that contained a drop of water and placed on a hot plate at about 30°C to

8
let dry. The dried samples were stained with 25 % Basic Fuchsin Toludine Blue and allowed to
dry on the hot. The samples were rinsed and allowed to dry again on the hot plate. The stained
dried samples were checked under an optical microscope to verify that a viable sample was
obtained. After a section of viable samples was reached, 90 nm slices were generated using the
same above procedure. 90 nm samples were placed on TEM grids.
Technicians, Dr. Lai and later Dr. Shang, assisted me with the preparation of samples and
operated the TEM during my sessions at the NISP Laboratory at the Kim Engineering Building
at the University of Maryland at College Park analysis. I communicated the technique I wanted
to employ, the magnification and the areas on the sample that I wanted to observe. Dr Lai and
later Dr. Shang took and saved micrographs I wanted to analyze later. Some of these images and
analysis are reported in Chapter 4.
I examined many of the TEM grids produced from our pilot study for many hours. I began to
focus on the TEM grids retrieved from the subcutaneous abdominal region of the rat implanted
with the mineralized crushed crab shell because of the interesting structures and particles
observed. Specifically after observing regions of collagen-like structures with the characteristic
67 nm banding pattern, the main question was are these structures mineralized?
I utilized high resolution transmission electron microscopy (HRTEM), selected area
diffraction (SAD), electron nanodiffraction (END), electron microanalysis - energy-dispersive
spectrometry (EDS) at the NISP characterization facility at the University of Maryland. These
techniques assisted me in answering the mineral presence question.
SAD was used in many regions. SAD can be useful for the determination of lattice parameter
information and unit cell details. Lattice structure gives information about the long range

9
composition of a material. Restrictive factors of SAD are that a good crystal must be found for
high accuracy of crystallographic structures and that only limited information about the
structure’s dynamic behavior can be obtained from one single diffraction experiment. Resolution
is an important limitation of SAD. Small particle size can lead to peak broadening and erroneous
interpretation of a SAD pattern (Ferrell Jr. & Paulson, 1977).
In an effort to locate regions of potential mineral formation, dark sections adjacent to
observed collagen bundles were evaluated. The images depicted a region that appears to harbor
material with crystalline characteristics. To assess its crystalline nature, SAD was performed.
The SAD pattern, displayed diffuse rings. This is typically indicative of amorphous solids.
However, using HRTEM clearly showed lattice spacing with long range order despite the fact
the reflections observed in selected area electron diffraction pattern exhibited broad peaks or
diffuse rings. Once I became aware of the nanodiffraction, Dr. Shang was asked to perform this
technique on the samples that had visible long-range order in HRTEM images even though their
SAD patterns suggested they were amorphous. Nanodiffraction offered an avenue to characterize
the ultrastructure of these nanocrystals. The nanodiffraction image depicted a single crystal. This
established that the nanosized matetial had crystalline characteristics. The nanodiffraction pattern
was indexed with the help of computer software and it was determined that the regular pattern of
widely spaced strong spots came from the apatite family.
I also communicated other areas to perform x-ray microanalysis and SAD. Plate shaped nano
-sized particles with lengths and widths of approximately 40 x 20 nm were observed with lattice
spacing measuring 0.95 nm. X-ray microanalysis was performed on the 40 x 20 nm nano
particles. This revealed an average calcium:phosphorus ratio of 1.81 ± 0.37. SAD was taken of

10
the observed nanocrystals, indexed, and determined to be of the apatite family. Moreover, this is
a very interesting diffraction pattern because it displays speckled rings made up of discrete spots.
This suggest that there are many oriented single crystals in this SAD pattern. One can surmise
that this diffraction pattern displaying many oriented single crystals is evidence for the
mesocrystal model of crystallization. Highly oriented subunits distinguish a mesocrystal from a
randomly oriented polycrystal, and the identifiable nano-sized building units distinguish it from a
single crystal. Often, it is difficult to distinguish a mesocrystal from a single crystal because the
mesocrystal shows an identical scattering pattern and behavior to a single crystal. Mesocrytsals
can be indexed like a single crystal. Therefore, it is difficult to define the border between a single
and mesocrystalline substance. However, mesocrystals were shown to be intermediates between
a classical single crystal and a polycrystal.
Limitations
Obvious strengths of a small pilot study are that the research question can be addressed in a
relatively short space of time, obtaining ethical and institutional approval is easier in small
studies, and it avoids spending too many resources. The main problem with small studies is
interpretation of results, in particular confidence intervals and p-values. Ideally all intervals
should be as narrow as 95%, but usually only large studies can produce such precise results.
Therefore, the data must be interpreted and reported carefully. The lack of statistical significance
does not mean there is no effect. There needs to be a prudent balance between not dismissing
outright what could be a real effect and also not making undue claims about the effect. While

11
small studies can provide results quickly, they do not normally yield reliable or precise estimates.
It is important not to make strong conclusions. Data from small studies should be used to design
larger confirmatory studies (Hackshaw, 2008) making methodical, logistical and financial
estimates. Pilots are rapidly becoming an essential pre-cursor. While there are weaknesses, this
pilot will be used to streamline our study to reduce the waste of resources and time.
No one animal model fully authenticates the processes occurring in human tissue repair. Only
humans can be used to reflect accurately the events of human healing, but even human studies
are subject to the pitfalls of laboratory methods and genetic or environmental variables that may
affect the results (Cohen & Mast, 1990). There is a great deal of concern regarding the ability to
transfer animal research data to the human clinical situation. Rats and humans share the
following skin characteristics: the presence of an epidermis, basement membrane, hair follicles,
and dermis. Among rodents, rat skin has more structural similarities to human tissue. Regarding
the rat skin, permeation kinetic parameters are frequently comparable with human skin, table 1-2
(Godin & Touitou, 2007). Obviously, there are numerous anatomical and physiological
differences between human and rat. Among the differences is the fact that rats do not form
keloids or hypertrophic scars but people of certain ethnic backgrounds, such as African-
Americans and Asians, are predisposed to excessive scarring (Dorsett-Martin & Wysocki, 2008).
Although most mammalian species simulate human healing with collagen deposition as a
predominant feature, the healing process in these animals is certainly not identical to humans.
Another difference in healing between animals and humans is their nutritional requirements.
Unlike humans, who require a dietary supply of ascorbic acid, rats are able to synthesize this

12
Table 1-2 Thickness of skin strata in rat, mice and humans (Godin & Touitou, 2007)
important cofactor needed for the production of collagen. (Cohen & Mast, 1990). It must be
recognized that humans to whom the results are being extrapolated are genetically highly
variable due to cultural, dietary, and environmental differences (Rand, 2008).
Characterizing the structure that were assessed to be collagen based on their characteristic
banding pattern was beyond the scope of my work and would not have provided insight on
mineral presence. The presence of mineral would distinguish the collagen production in this
study from typical wound healing collagen.
The amount of tissue available from harvested tissue is often a limiting factor. The limited
amount of harvested tissue available makes performing various test on each sample an issue. It
would have difficult to perform biological assays such as staining for calcium using alizarin red s
and the von Kossa technique and create TEM grids with such a small amount of harvested tissue
available. Additionally, it is nearly impossible to excise these wounds for any analysis without
including some surrounding normal skin. Therefore, contamination of harvested tissue by
uninjured skin can add a degree of inaccuracy in the evaluation.
Standardization in reporting, could facilitate comparisons and may initiate additional research
that favors the inevitable comparisons between studies. This increased knowledge base would be

13
vital in transferring animal-derived data to human clinical situations (Dorsett-Martin & Wysocki,
2008).
Hypothesis Implications
This was part of a preliminary study. Despite its exploratory nature, this study offers some
insight into osteoinductive potential of the crab cuticle. Conclusive evidence for osteoinduction
is characterized by heterotopic bone formation, that is, bone formation in tissues or organs where
bone does not naturally grow. Conclusive evidence for osteoinduction was not provided in this
study. No firm conclusions were made about crab shell’s osteoinductive potential. Although the
current study is based on a small sample of participants, the findings suggest the following:
The degree of crystallinity changes as bone matrix matures, can be masked by the size of
the crystal, and varies with location;
The results from TEM, HRTEM, XRD, SAD, END, and EDS, in this study indicate that
crushed shell from C. sapidus exhibits some unique interactions in vivo using Sprague–
Dawley rats;
The study indicates that crab cuticle in vivo interactions can result in the formation a
matrix containing collagen and mineral components;
These results further suggest that crab shell may be suitable for use in developing
functionally advanced bone implants;
Since bone formation can take months, this material warrants further study to ascertain its
potential osteoinductive properties fully.

14
Therefore, through the presented hypothesis, we suggest that that crab cuticle may be an
effective material for stimulating bone growth. Further extensive studies are warranted on large
randomized controlled trials to further assess the possible application and provide more
definitive evidence.

15
Chapter 2 Literature Review: Bone and Crab Cuticle and TRIZ
Introduction
Bone and crab cuticle are two dynamic and vital biomineralized composite structures. At first
glance, there appears to be more dissimilarities among these two tissues but when one takes a
closer look, there is an extensive array of similarities and compelling relationships. An
understanding of these similarities and relationships can point to a rich and abundant source of
inspiration for development of functionally advanced biomaterial (FAB) implants.
Large bone defects resulting from a multitude of causes, including aging, trauma, deformity,
and disease, are commonplace. These defects usually do not heal spontaneously and
interventions with bone grafts are often required. Bone grafts are substitute material that is
transplanted to the defective bone area to aid in healing, strengthening or improving function.
The clinical gold standard and preferred method for bone repair is grafts from the individuals
themselves, an autologous graft. Although effective and safe due to the low risk of disease
transmission or immunological rejection, autograft bone is limited by the availability of
sufficient donor tissue and problems with donor site morbidity (Braddock, Houston, Campbell,
& Ashcroft, 2001). There are several categories of bone grafts and substitutes that encompass a
variety materials, material sources and origins (Nandi, et al., 2010). All available grafts have
advantages and disadvantages and no single existing material possesses all the necessary
properties required in an ideal bone implant. Biocompatible implanted materials have provided
options in many cases, but the reaction of the body to these devices is far from perfect.

16
Complications, such as thrombosis, infection inflammatory reaction, impaired function,
loosening, and pain, are unfortunate for the patient and costly to the health care system. But, for
biomaterials researchers, these shortcomings with existing materials also represent opportunities
and challenges to engineer improved therapies. (Ratner, 2001). A suitable bone graft material of
proper quality, that is readily available in unlimited quantities, is still needed (Wise, Trantolo,
Lewandrowski, Gresser, & Cattaneo, 2002). Bone defects can be addressed by tissue engineering
techniques (Braddock, Houston, Campbell, & Ashcroft, 2001) that draw from the best material
scientist known --- Nature. This review will cover biomimetics and the uncanny similarities
between bone and crab cuticle.
The overall goal of using bone as a biomimetic subject is to learn from its unique structure
and function and use this insight to develop bone inspired implants that enhance healing and
remodeling of bone (Wilson Jr., 2008). Adapted as a bone implant material and early
biomimetics, nacre’s ability to integrate with bone was noted as early as 600 A.D. in the ancient
Mayan civilization (Westbroek & Marin, 1998). Drawing from this inspiration and accentuating
the interesting similarities to bone tissue, the crab shell cuticle itself has been underestimated as a
candidate material for engineering of bone. Biomimetics and design concepts of TRIZ (Theory
of Inventive Problem Solving) reveals crab cuticle as candidate material for development of FAB
bone implant. In this paper, bone and crab shell integument functional and general similarities
will be highlighted.

17
Figure 2-1 Making an Ideal Bone Graft (Stocker & Wolinetz, 2009).

18
Ideal Bone Implant
Materials designed by humans pale in comparison to those created by nature (Smith & Wood,
1991). Bone, one of nature’s masterpieces, is a remarkable, living, mineralized, connective
tissue, which is characterized by its strength, its resilience, and its ability to remodel and repair
itself (Hing, 2004). Bone is composed of an organic matrix of fibrous protein and collagen (30–
35% of the weight); inorganic calcium phosphate (65–70% of the weight); and water (Kalfas,
2001). Ideally, a bone replacement material should mimic bone tissue from a mechanical,
chemical, biological and functional point of view, be disease free, and facilitate new bone
formation.
There are three characteristics inherent to natural bone: osteoconductivity; osteogenicity; and
osteoinductivity as depicted in figure 2-1 (Stocker & Wolinetz, 2009). Osteogenesis is the ability
of the graft to produce new bone and is dependent on the presence of live bone cells.
Osteoconduction is the physical property of the graft to serve as a scaffold for viable bone
healing and allows for vasculature and the infiltration of osteogenic precursor cells.
Osteoinduction is the ability of graft material to induce stem cells to differentiate into mature
bone cells (Kalfas, 2001). Thus, to facilitate new bone formation an ideal bone substitute
material should have the characteristics inherent to natural bone.
Biomimetics
Material design and development for any replacement or regeneration application should be

19
based on the thorough understanding of the structure to be substituted. Material researchers can
gain valuable knowledge from mineralized tissues. Typically, the main characteristics of the
route by which the mineralized hard tissues are formed are that the organic matrix is laid down
first and the inorganic reinforcing phase grows within this organic matrix/template. Among the
wide variety of biomineralized materials engineered by living organisms, bone and crab wide
variety of biomineralized materials engineered by living organisms, bone and crab exoskeleton
are organic/inorganic composite structures with analogous mechanical properties. Non-biological
methods have not been able to duplicate the elegance of the biomineral assembly mechanisms or
the rather complex composite microarchitectures of hard mineralized tissues like bone and crab
(Reis & Weiner, 2004).
Uniformity of defining and expressing terms or concepts varies depending on discipline. For
example, cuticle is also known as exoskeleton, integument, shell, or carapace, depending on
discipline. Additionally, crab is also referred to as crustacean or arthropod. Many times these
terms are expressed interchangeably. Biomimetics is no stranger to this phenomenon of non-
uniformity. One definition of biomimetics describes it as the study of how Nature designs,
processes and assembles/disassembles molecular building blocks to fabricate high performance
materials such as mineral-polymer composites (e.g., mollusk shells, bone, etc.) and then applies
these designs and processes to engineer new molecules and materials with unique properties
(Reis & Weiner, 2004). Some relate two ways in which biological and engineering systems can
be related --- bioinspiration and biomimetics: (1) bioinspiration is describes as how inspiration is
gained from nature as a basis for developing engineering solutions to problems which requires an

20
Figure 2-2 Expanded description of the field of biomimetics (Sarikaya, 1994).
Biomimetics
(Nature) Understanding, Transfer, Implementation (Engineering)
Repurposing
existing
biological
materials
Using engineered
materials to mimic
(Structure, Energy, Etc.)
Design
Biorecycling Biomimicking
Fields of Operations
(Structure, Energy, Etc.) Bioduplication
Bioinspiration
Biologistics
Novel Synthesis
& Processing
Mastering
molecular
synthesis and
processing
mechanism
New Materials with Tailored Structures
Existing Materials with
Tailored Purpose

21
understanding of the natural system and the operational envelope within which it operates; and
(2) biomimetics is defined as mimicking nature in some way. Inspiration is considered the
dominant of the two (Allen, 2010). Others divide biomimetics into two categories based on
design approaches --- biomimicking and bioduplication: (1) biomimicking deals with the
investigation of the structure of a biomaterial at all possible length scales of spatial resolution
and then deducing and ultimately mimicking the fundamentals of the biomaterial’s unique design
structure; and (2) bioduplication deals with the mastery of the molecular synthesis and
processing mechanisms of biomaterials and applying these newfound methodologies to produce
new technological materials (Sarikaya, Liu, & Aksay, 1995).
However, the argument can be made that fabrication via biomimetics must first start with
inspiration. Secondly, the biomimetic design process is followed by studying or deducing how
Nature designs, processes, and assembles/disassembles molecular building blocks to
manufacture materials. Then the process involves applying the fundamental knowledge gained
through planning and management. Finally, the process creates a novel purpose, strategy, or
material by techniques such as biorecycling, biomimicking (structure, energy, etc.), or
bioduplication. An expanded definition of biomimetics is provided in figure 2-2.
By studying biological principles of nature, new ways of dealing with engineering problems
can be unlocked. Nature or biology finds the most economical and efficient ways to achieve its
objective in terms of energy, cost, and materials. Biology frequently uses a few simple materials

22
Figure 2-3 Biological (right column) and Engineering (left column) materials are very different
in the way they are developed (Allen, 2010), (Fratzl, 2007).
Biological Material
Light common Elements
H, C, N, O, Si
Ca, Na, P, S Cl, K, ...
Growth
by Biologically Controlled Self-Assembly
(Approximate/Variable Design)
Environmentally Influenced Self-Assembly
Hierachical Structuring at all size levels
Interfaces allow separate control of stiffness & fracture
Adaption of form & structure to the function
Responsive Design
Environmentally Responsive; adaptive in function & morphology; Self-repair
Engineering Material
Many Heavy Elements, Some rare
H, C, N, O, Si
Fe, Ni, Al, Zn, Cr, ...
Fabrication
from Melts, Powders, Solutions, Etc.
(Exact Design)
Externally Imposed Form
Mostly Monolithic; Little or No Hierarchy; Forming Parts & Microstructuring
Materials
Few Interfaces; therefore poor fracture control
Selection of material according to function
Secure Design
Considering possible max load; Very little environmental response; Self-repair (little
to nonexistent)

23
arranged in a particular manner. On the other hand, engineering uses much more energy and
often achieve less impressive results (Allen, 2010). Biological and engineering materials are very
different in how they are developed, as shown figure 2-3.
Natural materials use free energy while constructing and operating under conditions of low
temperature (0-40°C), atmospheric pressure, and neutral pH (Smith & Wood, 1991). Biological
materials use few polymers (protein, polysaccharide, etc.), with ceramics (calcium salts, silica,
etc.), and a few iron-containing ceramics (FeS, Fe3O4, etc.), whereas human-made materials
require high temperatures and hundreds of polymers (Allen, 2010). Consequently, the design
strategies of biological materials are not immediately applicable to the design of novel
engineering materials. One major difference is in the range of choice of elements. Biological
materials consist of relatively few constituent elements, which are used to synthesize a variety of
polymers and minerals. Although the choices are far greater for the engineer, man-made
materials are characterized by the use of many more elements and usually cannot match natural
performance of nature’s materials. Another major difference is the way in which materials are
made. While the engineer selects a material to fabricate a component according to an exact
design, nature grows both the material and the whole organism using principles of biologically
controlled self-assembly. This is certainly a key to the successful use of polymers and
composites as structural materials and provides control over the structure of the material at all
levels of hierarchy (Fratzl, 2007). Biology, like engineering, is reliant on materials for the
structures it makes. These structures have to be cheap and reliable (Allen, 2010).
Fundamental science, such as biology, has the goal to observe, describe, classify and analyze.

24
On the contrary, engineering has the intention to act, prescribe, utilize and manage (Vincent J. F.,
Bogatyreva, Pahl, Bogatyrev, & Bowyer, 2005). Thus, the transfer of a concept, process, or
mechanism from living to nonliving systems is not a straightforward matter. The advantage of a
biomimetic approach is that biology has already done a lot of the design prototyping; the
disadvantage is that every case has to be approached newly (Allen, 2010).
Bone and Crab Cuticle Functional Similarities using TRIZ
Different approaches have been developed to solve the same problems. There is no clear
roadmap or single path to the correct way to solve a given problem. Traditional problem solving
techniques focus on what is needed to solve the problem. Thus, the classical problem solving
loop involves: (1) identifying the problem; (2) exploring information generated from a specific
area of expertise; (3) creating ideas based on a specific background; selecting the best idea(s); (4)
building and testing idea(s); and (5) analysis and evaluation of results.
An innovative approach to creative problem solving involves carefully identifying the ideal
result in terms of a delivered function then working out what is needed to get to that ideal.
Altshuller developed a rigorous set of methods that concentrate on defining the function to be
delivered, the working environment or context in which it is to be delivered, and the resources
available within that environment (Allen, 2010). After studying thousands of patents, he
developed a set of methods based on the idea that the solution is best delivered by altering the
environment, rather than by altering the immediate mechanism of delivery and called this set of

25
methods “The Theory of Inventive Problem Solving” (in Russian, Theoriya Resheniya
Izobreatatelskikh Zadatch, or TRIZ). His approach showed that any branch of technology could
provide a solution to a specific problem, even if that branch does not appear to be related to the
problem or its apparent solution. Defining the innovation in terms of the function it is desired to
deliver and expressing it in simple relatively abstract terms, destroys the barriers between various
areas of knowledge (Allen, 2010).
TRIZ is an available design tool that establishes a system into which all known solutions can
be classified in terms of function. A database or information fund exists along with software
packages that facilitate the use of this tool. This database reflects a concentrated collection of
experience and knowledge that can be used to leverage natural and/or technical insight. To date,
the database contains limited biological information. As biological knowledge increase, the area
where nature/biology meets engineering, shrinks (Vincent & Mann, 2002), improving the
creation and application of biomimetic designs, concepts, or solutions.
The typical TRIZ iterative problem solving process involves: (1) identify or define
problem/function; (2) problem/function analysis and definition, using the database and language
of TRIZ, that provides insightful information; (3) contradiction/conflict analysis using
standardized factors (weight, speed, ease of use, etc.) — TRIZ theory contains 39 design
parameters and 40 generic principles for solving the contradictions derived by Altshuller; (4)
analysis of how contradictions/conflicts were circumvented or eliminated; (5) feasibility analysis
of problem resolutions of conflicts; and (6) iteration, if valid solution not found after conflict
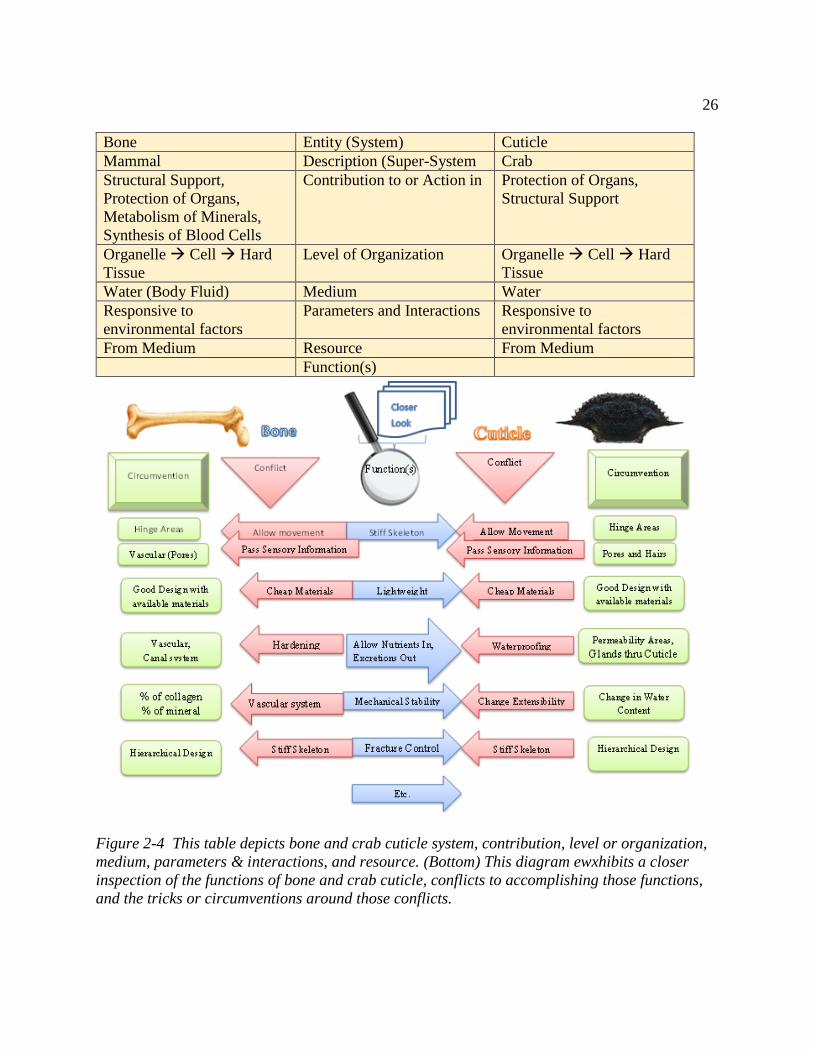
26
Bone Entity (System) Cuticle
Mammal Description (Super-System Crab
Structural Support,
Protection of Organs,
Metabolism of Minerals,
Synthesis of Blood Cells
Contribution to or Action in Protection of Organs,
Structural Support
Organelle Cell Hard
Tissue
Level of Organization Organelle Cell Hard
Tissue
Water (Body Fluid) Medium Water
Responsive to
environmental factors
Parameters and Interactions Responsive to
environmental factors
From Medium Resource From Medium
Function(s)
Figure 2-4 This table depicts bone and crab cuticle system, contribution, level or organization,
medium, parameters & interactions, and resource. (Bottom) This diagram ewxhibits a closer
inspection of the functions of bone and crab cuticle, conflicts to accomplishing those functions,
and the tricks or circumventions around those conflicts.

27
elimination (Yeh-Liang, Hung, & Yin, 2006). These factors, and the resolutions of the conflicts
which was defined, represent a collection of engineering best practice (Vincent, 2005). In short,
TRIZ is a systematic and repeatable method for broadening the field of thinking across
disciplines and for enhancing creative options.
Using aspect of a biological case study conducted by Vincent based on TRIZ analysis of a
relatively simple natural composite material—the outer covering of arthropods or cuticle,
similarities between the functions of bone and crab cuticle can be highlighted. Produced by a
single layer of epithelial cells, this layer of material is charged to provide a large number of
functions such as protection, shape, structure, hinges, barrier, filter, etc. (Vincent, 2005). Bone is
charged with many of the same functions. In his study, a list, not necessarily exhaustive or
unbiased, of functions and associated characteristics of cuticle was generated partly by reference
to literature and partly from experience with insects and insect cuticle over the years. In this case,
a short list of functions, conflicts, and circumventions shared by both bone and crab are
illustrated in figure 2-4. It can be easily demonstrated that bone and crab cuticle use very similar
approaches to the solution of design and integration, functionally speaking. Figure 2-5 illustrates
the idea of a contradiction or mixed messaging. A contradiction characterizes the conflicting
goals materials strive to satisfy.
From the point of view of biomimetics, if, by defining functions and comparing the ways in
which biology and engineering deliver a range of functions, an independent assessment can be
made whether engineering stands any chance of being environmentally benign by comparison to
Nature (Allen, 2010).

28
Figure 2-5 A contradiction describes the predicament caused by an attempt to improve one
property of a system that induces the degradation of another property (Stanley, Zlotin,
Bolckmans, & Zusman, 2005).
TRIZ is versatile and can vary drastically in its level of complexity. TRIZ analysis can not
only illuminate that bone and crab exoskeleton performs many of the same functions but
circumvents many the same contradictions in a similar manner. For example, both bone and crab
cuticle have the property of being stiff. However, bone and crab cuticle have to allow movement
and pass sensory information. These contradictions are circumvented with hinge areas and a
system of pores and vascularity. As one delves deeper into the functions, contradictions, and
circumventions, the level of complexity can vary from a general surface analysis to a more
thorough approach. As an illustration, if the discussion were to continue regarding stiffness,
movement, sensory information flow, pores, etc., the discussion could include the entire sensory
network system --- cells, channels, piezoelectric quality, homeostasis, etc. At the same token, the
list of functions can expand. In essence, TRIZ analysis can start and stop with small building
blocks or it can burgeon into a complex framework, figure 2-6.

29
Figure 2-6 How TRIZ works (Stanley, Zlotin, Bolckmans, & Zusman, 2005).
Transferring Biology to Engineering
Properties of materials allow communication of requirements when selecting materials for
implants. An organized system of property information then becomes a valuable commodity. The
functional knowledge coupled with other property information can form a powerful database that

30
(1) equips with new wisdoms and connections to ask and to possibly answer novel questions; and
(2) has the potential of advancing science, pioneering new methods, and ensuring quality of
biomaterials and other products.
TRIZ has been traditionally used to show how technical solutions compare to each other.
Combining biomimetic concepts with TRIZ allows inspection of the way nature solved problems
and see how that compares with technical solutions to the same or similar problems. TRIZ
establishes a systematic scheme in which problems and their solutions can be defined and
classified. Transferring a concept or mechanism from living to nonliving systems can be
daunting because nature exists within a different putative context than nonliving systems. Bone
represents an example of a biological putative prototype that is continually recycled or repaired
for the duration of its life inside the body and profoundly difficult to replicate. Therefore, some
approach or form of interpretation or translation is needed to make the leap from biology to
technology (Vincent, 2008).
Biomimetics is not a brand-new was of adapting ideas from biology, but its approach is
currently guided by experience. TRIZ introduces a set of common principles that builds on
current successes and has the ability to establish a transparent method for technologists to access
biology. TRIZ is a systems approach for engineering and biology is regarded as a system. TRIZ
involves the transfer of various inventions and solutions from one field to another, while
biomimetics involves the transfer of functions, mechanisms and principles from one field to
another. Therefore, TRIZ is a superior candidate to be an ideal bridge to assist in identifying
biomimetic principles, concepts, and functions and transferring them from nature to

31
Figure 2-7 Schematic of steps to identify a function/concept/idea and to transfer the function
into engineering solution (Vincent, 2008), (Shimomura, 2010).
ENGINEERING *BIOLOGY
Total Mimicry
Partial Mimicry
Abstraction
Mechanics
Physics
Inspiration
_______optimizations________
Identify the challenge/function
Interpret challenge/function in terms of biology
Discover biological model that performs same function or deals with same challenge
Analyze the patterns and processes of the biological model
Transfer biological ideas, functions, processes to engineering *
To
Develop ideas and solutions based on biological model
Biological Model: Function/Property Mimicked Product/Solution Lotus Effect: Superhydrophobicity Paint, Self-Cleaning
Sharkskin: Fluid Resistance Swim Suit
Termite Den: Temperature Maintenance Building Architecture
Plant Burr: Cling Velcro (two-sided fastener)
Whale Fin: Reduce Drag Turbine blades
Kingfishers Beak: Travels Quickly & Smoothly Between Contrary Mediums Bullet Train

32
technology/engineering, as shown in figure 2-7 (Vincent J. F., Bogatyreva, Bogatyrev, Bowyer,
& Pahl, 2006).
Although TRIZ is a promising system for use in biomimetics, there are considerable issues.
Biology and technology solve problems in rather different ways. TRIZ is derived from the
environment of things artificial, non-living, technical and engineering while biology is in the
realm of the living while biomimetics operates across the border between living and non-living
systems. Living organism are highly complex. They have great adaptability and versatility, but at
the expense of the predictability of the system’s behavior. Generally, unpredictability in
technical systems are typically unacceptable and avoided.
Two of the more pronounced challenges between biology and technology deals with
hierarchy and energy. Typically living systems are based on a hierarchical structure. Complexity
and added functionality are achieved by adding levels of hierarchy in Nature, due to processing
limitations current technologies are either not significantly hierarchical, or ignore any
hierarchical structure. The hierarchical approach is also difficult to model in engineering and is
not as well developed as it is in biology. Energy is also a testament to the challenges of
transferring biology to technology. Biological functions and processes are less reliant on energy
(Vincent J. F., Bogatyreva, Bogatyrev, Bowyer, & Pahl, 2006), than their technological
counterparts. As biomimetics and TRIZ tools are developed and used more frequently, functions,
ideas, or concepts can hopefully be readily translated from biology to engineering.

33
Figure 2-8 Bone and Crab Shell Similarities.
Hierarchical structuring at all size levels
Biomineralized composites
Ca2+ based inorganic phase
Each inorganic pahse (hydroxyapatite, calcium carbonate contains 40% Ca2+ by mass
Organic ploymer matrix pahse (collagen, chitin)
Highly loaded inorganic phase (50-70 wt%)
Textured, crystallographic orientation of inorganic phase
Protein and muccopolsaccharide constituents
Relatively hard and damage tolerant
liquid crystalline behavior of orgainc matrix
Structural support role
Cellular control
Self-healing behavior
Piezoelectric propertiies (ability to convert stress to electrical signals)
Similarities between the biomineralized hard tissues bone and crab shell

34
General Comparison of Bone and Crab Cuticle
Crab shell integument shares chemical and structural features that are closely related to bone.
The similarities of bone and crab cuticle are outlined in figure 2-8. Both hard tissues are formed
via biomineralization processes and contain calcium as a major component, with calcium
phosphate present in bone and calcium carbonate in crab exoskeleton. The mineral phase of bone
is often referred to as carbonated apatite because carbonate substitutes readily for both phosphate
and hydroxyl anions in biological hydroxyapatite. On a macroscopic level, bone and crab cuticle
provide structural support and protect internal organs. Bone tissue customarily resides inside the
body while crab integument functions as an external protective armor. These hard tissues both
serve as storage sites for calcium, phosphate, carbonate, and other biologically relevant ionic
species. The nanoscale level of bone and crab shell have features that are truly remarkable from a
materials design viewpoint. Bone and crab cuticle are composed of an organic matrix embedded
with mineral network where proteins contribute to the controlled nucleation and growth of
inorganic phase. In exoskeleton of crustaceans, the polysaccharide chitin associates with calcite
while in bone, the protein collagen associates with calcium phosphate (Giraud-Guille, Belamie,
& Mosser, 2004). Both of these associations produce biomineral crystals that appear as thin
platelets (Weiner & Wagner, 1998). Both bone and crab cuticle serve as a framework for the
activity of specialized cells. Bone cells --- osteoblasts, osteoclasts, and osteocytes --- activities
include orchestrating signalling and producing extracellular matrix proteins that coordinate
cellular and tissue interactions while crab cuticle epidermal cells produce and secrete organic
matrix for maintenance of integument integrity and function through various molt cycles and

35
survival events. Bone has a role in providing a reservoir for the production and storage of
hematopoietic and mesenchymal stem cells. This reservoir of cells in the body suggest that bone
is capable of osteogenesis throughout life (Aubin, 1998).
Hierarchical structure and biomineralization phenomena play an integral role in crab shell and
bone tissue physiology and function. In both hard tissues, the manner in which organic
framework molecules, collagen and chitin, influence biomineral formation (nucleation, growth,
morphology, etc.) is a truly intriguing study. This will be discussed in the next chapter.
A fundamental understanding of bone and crab shell structure and biology can be gained by
studying the chemical, physical, and mechanical structure of biomineralized tissues bone and
crab integument with a focus on the organic phase, the inorganic phase, and the inter-relations
between and among these phases. This expanded understanding is a necessary course to develop
successful bioimplants.
Concluding Remarks
Biology and technology/engineering use very different approaches to the solution of similar
problems or functions of design and integration. However, many of nature’s systems use the
same exquisite and superior design principles. As hard tissue systems are studied, the similarities
between systems such as bone and crab cuticle become obviously clear. Because technological or
engineering designs cannot, to date, match the performance of nature’s materials, biomimetic
design strategies, such as biorecycling allows exploitation of natural environmentally friendly
materials, such as marine wastes (Hirano, Nakahira, Nakagawa, & Kim, 1999). This exploitation

36
takes advantage of the economical and efficient ways nature achieves its objective in terms of
energy, cost, and materials. In addition, most of the design prototyping and fabrication has been
done.
If the desired functionally advanced biomaterials based on crab cuticle are to be produced
through biomimetics, further knowledge is needed about: (1) the micro- and nano- structure; (2)
variations of composition and structural organization of the matrix; (3) responses of phases
(organic matrix, and interfaces) under various mechanical stresses; (4) toughening,
strengthening, and hardening mechanisms; (5) biomineralization; and (6) growth mechanism
based on geometry, crystallography, hierarchy, and size. (Sarikaya, Liu, & Aksay, 1995).
Consequently, as the understanding of how nature designs many of its complex systems is
illuminated, it becomes obviously more transparent that systems and their functions in biology
can be adapted to technological ends. Therefore, biomimetics can offer novel and valid
alternatives to technological problems (Vincent, 2005). As biomimetics, TRIZ and other
techniques evolve, they can further illuminate that the solution to many of our technological
problems has been hiding in plain sight and that nature has a multitude of recommendations.

37
Chapter 3 Ties that Bind: Evaluation of Collagen I and α-chitin
Introduction
Bone and crab cuticle are two dynamic and vital biomineralized composite structures.
Studying the similarities and differences between bone and crab shell at various structural levels
allows a greater understanding of the inter-relationships between biomineralized tissues and how
they function to repair, restore, and regenerate. Bone and crab shell integument is comprised of
an organic component and an inorganic component. Calcium phosphate and calcium carbonate
are the inorganic or mineral constituents of bone and crab cuticle respectively. Likewise,
collagen and chitin are the organic phases of bones and crab cuticle, respectively. Table 3-1
exhibits the principle components of two similar biological systems: crab exoskeleton and bone.
Focusing on the organic constituents, some interesting similarities and some unique qualities can
be elucidated. Thus, chitin and collagen are two fascinating matrix framework molecules that
play a key role in the hierarchical development of hard tissue. For a thorough and systematic
design of a bone inspired bio-implant, it is fundamental to study the influence of individual
components (Daamen, et al., 2003) of biological systems.
The key chemical and structural features of chitin and collagen were studied in order to gain
insight into the functional role of important organic matrix material of hard tissue. Both collagen
and chitin are constructed with polymeric fibers. Chitin is the second most abundant natural

38
Table 3-1 Principal Components of Crustaceans (Crab) Exoskeleton and Bones Mineral Organic
Biological Composite Hydroxyapatite
(calcium phosphate)
Calcium Carbonate Collagen Chitin
Bones
X X
Crustacean (Crab) Exoskeleton
X
X
fibrous polymer while collagen is among the first. One of the more conspicuous differences is
the biochemistry of collagen I, a protein, and α-chitin, a polysaccharide. There is a considerable
amount of information about the structure of chitin and collagen. However, all the mechanism
involved in the self-assembly of chitin and collagen molecules into solid fibrous structures stills
lacks complete clarity. Both collagen and chitin fall under the class of molecules described by
Bouligand as ‘biological analogues of liquid crystals’ (Bouligand Y. , 1972). Experiments have
accentuated the importance of liquid crystal characteristics with respect to self-assembly. Thus,
the more intriguing similarities involve hierarchical structuring, liquid crystal characteristics and
self-assembly. Advances made in regard to the identification of molecules involved in cellular
and morphogenetic regulation have provided a window into how collagen, chitin, and similar
biological components interact to form fibrous composites. The chemistry, hierarchy, synthesis,
liquid crystal characteristics, biomedical applications and other properties were evaluated to aid
in a deeper understanding and development of concepts for biological, hierarchical, composite
design. In this work, our focus is on inter-relationships between the biomolecular building blocks
chitin and collagen and their respective roles in the architectural development of molecular
organic matrix (MOM).

39
A growing body of work highlights bone and crab exoskeleton including aspects of collagen
and chitin, see Table 3-2 (Bouligand Y. , 1972), (Giraud-Guille M.-M. , 1994), (Giraud-Guille,
Belamie, & Mosser, 2004), (Belamie, Mosser, Gobeaux, & Giraud-Guille, 2006), (Meyers, Lin,
Seki, Chen, & Bodde, 2006), (Meyers, Chen, Lin, & Seki, 2008). One group highlights the
liquid crystal properties of α-chitin and collagen. However, studies that feature a more
comprehensive view of solely collagen I and α-chitin is limited. This article will examine the
organic constituent of bone and crab exoskeleton, collagen I and α-chitin, respectively. Collagen
I and α-chitin have many similarities as well as differences, and. hence forth these fibrous
composites will be referred to primarily as collagen and chitin. Collagen and chitin work
together on various levels sometimes leading, other times playing a more supportive role, but
always integrally linked in establishing form and functionality in biological structures and
systems. Specifically, collagen interacts with polysaccharides via extracellular matrix (ECM)
(collagen and ECM in bone structure (various roles in cell signalling, biomineralization, etc.);
Chitin interactions with proteins (chitin/protein complex (beta pleated sheets in nacre, etc. This
paper is organized to address the similarities and differences at various level of organization of
collagen and chitin.
Framework of chitin and collagen
Chitin was first isolated from mushrooms in 1811 by the French scientist Henri Braconnot.
In 1823, Odier found the same compound in the cuticles of insects and named it chitin
(Stephen, 1995). At least 10 gigatons (1 × 1013 kg) of chitin are synthesized each year making

40
Table 3-2 Key Articles that Highlight Collagen and Chitin
Author Year Title
Bouligand 1972 Twisted Fibrous Arrangements in Biological Materials and
Cholesteric Mesophases
Giraud-Guille 1994 Liquid Crystalline Order of Biopolymers in Cuticles and Bones
Giraud-Guille 2004 Organic and mineral networks and biomimetic materials
Belamie 2006 Possible transient liquid crystal phase during the laying out of
connective tissues: α-chitin and collagen as models
Meyers 2006 Structural Biological Composites: An Overview
Meyers 2008 Biological materials: Structure and mechanical properties
chitin the second most abundant natural polymer on earth. It is found in the exoskeleton of
crustaceans, cuticle of insects, and cell wall of fungi (Jolles & Muzzarelli, 1999). On the other
hand, collagen appears to have been first reported by Zachariades in 1900 (Slack, 1955).
Collagens are found in all multicellular organisms (Gobeaux, et al., 2008). Chitin and collagen
play an important role in the hierarchical control of the biomineralization processes and in the
structural component that provides support and protection.
Chitin Forms
Three polymorphic forms of chitin (α-, β-, and γ-chitins) have been distinguished due to their
crystal structure that differ in packing and polarities of adjacent chains in successive sheets,
figure 3-1. α-Chitin, the most abundant form found in nature, is arranged in an anti-parallel

41
Figure 3-1 Diagram showing the direction of the polymer chains in the three crystallographic
forms of chitin. (a) α-chitin. (b) β-chitin. (c) γ-chitin (Wainwright, 1982).
configuration; β-chitin is organized in a parallel configuration; and γ-Chitin is a mixture of α-
and β-chitin. The anti-parallel configuration gives a highly ordered crystalline structure with
strong hydrogen bonding between chitin chains, leading to the rigid and insoluble properties of
α-chitin (Meyers, Chen, Lin, & Seki, 2008).
α-Chitin types vary as to the length of their N-acetylglucosamine chains, as to the degree of
crystallinity, and as to the dependence on the presence of accessory inorganic compounds or
proteins. As a consequence, the consistency of the chitinous layers in organisms like arthropods
differs considerably (Jolles & Muzzarelli, 1999). A chitinous exoskeleton is diagnostic of the
crustaceans. It is secreted by a single layer of cells (the epidermis) and gains its stiffness and
structural complexity mainly by being folded and curved in many complex ways (Vincent J. F.,
Arthropod cuticle: a natural composite shell system, 2002).
α-chitin β-chitin γ-chitin
(a) (b) (c)

42
Collagen Forms
More than 28 different types of collagen have been identified (Gobeaux, et al., 2008) and
classified primarily according to their physiological structure. Additionally, considering the
relationship between the inner structure of collagen fibrils, their diameter, their spatial layout and
the functional requirements they have to withstand, Ottani et al. proposed that collagen fibrils of
fibrous connective tissue may also belong to two different, mutually exclusive forms indicated as
“T-type” and “C-type”. The first class, consisting of large, heterogeneous fibrils, parallel tightly
packed, subjected to tensile stress along their axis is found in highly tensile structures such as
tendons, ligaments and bone. The other class, consisting of small, homogeneous fibrils, helically
arranged, resisting multidirectional stresses, is mostly present within highly compliant tissues
such as blood vessel walls, skin and nerve sheaths (Ottani, Raspantib, & Ruggeri, 2001).
Vendors, such as Sigma Aldrich, continue to refer to collagens both by the widely used
‘‘modern’’ type classification system defined by the chain types and by types as defined in the
early 1970s by the researchers who first separated collagen chains (Miller type II cartilage). The
earlier collagen classifications tended to be defined more by the tissue type from which the
collagen had been extracted than the collagen chain content (Abraham, Zuena, Perez-Ramirez, &
Kaplan, 2008). Collagen type I usually consists of three coiled subunits: two α1(I) and one α2(I)
chains forming fibrils of 50 nm in diameter (Kolacna, Bakesova, Varga, Kostakova, & Planka,
2007). The family of fibrillar collagens is of the biochemical types I, II, III, V, and XI (Ottani,
Matin, Franchi, Ruggeri, & Raspanti, 2002). All the approximately 44 genes for 28 collagen
types were described up to now; they are divided according to their supramolecular structure.

43
Alpha-chains descend from a common ancestor and are characteristic for each collagen type: the
chains are numbered according to the respective collagen type. The structure of the heterotrimer
in collagen I is [α1(I)2 α2(I)], and the structure of the homotrimer in collagen I is [α1(I)]3]
(Abraham, Zuena, Perez-Ramirez, & Kaplan, 2008). Collagen type I, the main constituent of
bone, tendon, skin, and vessel walls, is synthesized by fibroblast, smooth-muscle cells,
chondroblasts, odontoblasts, and osteoblasts. Osteoblast synthesize only type I collagen (Kuhn
& Glanville, 1980).
Both α-chitin and collagen I are crystalline. While there are three polymorphic forms of
chitin, there are 28 identified types of collagen defined by chain types. Additionally collagen
can be further classified according to its physiological structure.
Chemistry
Collagen I and α-chtin both consist of relatively few constituent elements: H, C, O, and N
(Meyers, Chen, Lin, & Seki, 2008). The chitin monomer general empirical formula is
[C8H13O5N]n. The empirical chemical formula for collagen is [C12H24O4N3].
Polysaccharides and Proteins
The basic chemistry of collagen and chitin has its origins in the basic building blocks of cells.
Major cellular compounds are lipids, carbohydrates and amino acids; these compounds mostly

44
combine to form nucleotides, polysaccharides, and proteins, Table 3-3. Chitin is a polysaccharide
while collagen is a polypeptide or protein.
Polypeptides or proteins are organic compounds made of amino acids arranged in a linear
chain polymer and joined together by peptide bonds between the carboxyl and amino groups of
adjacent amino acid residues. A particular protein always has a specific number and sequence of
amino acid residues. The basic unit of collagens is a polypeptide consisting of the repeating
sequence (G- X - Y)n glycine (G) is small, and is the only amino acid that fits in the crowded
interior of the triple helix. X is usually proline and Y is usually hydroxyproline. See figure 3-2.
Figure 3-2 Chemical Structure of Collagen showing the glycine, proline, and hydroxyproline
residues (Brodsky, Werkmeister, & Ramshaw, 2005).

45
Figure 3-3 Fragment of chitin chain (Ehrlich, 2010).
Table 3-3 Some Macromolecules (polymers) and their primary biological functions (Smith &
Wood, 1991).
Monomer Polymer Examples Functions
Monosaccharide Polysaccharide Chitin Structural
Amino Acids Proteins Collagen
Enzymes
Hormones
Structural
Catalysis
Messengers
Nucleotides Nucleic acids DNA
RNA
Information
Storage
Information
Transfer,
Structural

46
The basic unit of chitin is long linear polysaccharide chain of N-acetylglucosamine joined
together by glycosidic bonds, figure 3-3. The glycosidic link joins the number 1 carbon of the
N-acetylglucosamine to the number 4 carbon of the second N-acetylglucosamine. Joining the N-
acetylglucosamine monomers in a different way would result in a different polysaccharide. β1-4
linked residues tend to form an extended ribbon. Individual polysaccharides rarely have a
specific Mi but rather show a range or distribution of M.
One of the structural features distinguishing polysaccharides from proteins is the great
variation possible in the configuration of the glycosidic bond between adjacent sugar residues.
The many possibilities for linking allow a variety of shapes and surfaces to be formed from just a
few sugar residues. Using a limited range of N-acetylglucosamine, a range of unique molecular
shapes may be generated. With some exceptions, the function of polysaccharide structure lie in
the uniform and repeating shapes and surfaces rather than size itself. In contrast, the result of
protein folding is to produce a unique but irregular surface, a feature that requires a precise
sequence and fixed number of amino acids. Proteins can produce quaternary structures. Both
proteins like collagen and polysaccharides like chitin can produce a range of unique shapes and
surfaces.
Fibrous protein such as collagen have distinctive amino acid compositions, in which a very
few amino acid residues predominate, and usually there is a short sequence of residues that is
repeated many times. They often contain unusual, modified amino acid residues and tend to be
highly cross-linked, giving them strength. Compared with other fibrous proteins, the

47
composition of collagen is unusual. There is a very high content of proline. Additionally, and
uniquely, a large proportion of the proline in collagen is hydroxylated (Smith & Wood, 1991).
Another difference between proteins and polysaccharides is in the variety of components.
Each polysaccharide typically comprises only a few different types of sugars. Often, there is but
one. Sequences of repeating blocks of mono-, di-, tri-saccharides, etc. produces polysaccharides
of simple structures such as ribbons. There is a limited array of groups on the polysaccharide
surface capable of interaction with other molecules. This can be contrasted with much greater
range of chemical or physical interactions displayed by amino acid side chains on the surface of
a protein. Their very simplicity allows them to interact other polymers to form large molecular
networks with important and desirable properties, especially in the extended linear
conformations.
Although there is a limited range of polysaccharide types, the glycosidic bond between
adjacent residues is not confined to just one group in the sugar ring. In contrast to proteins, a
number of glycosidic bond arrangements are found between sugar residues where identical
Table 3-4 Molecular Weight of Chitin and Collagen
Molecule Molecular Weight (Daltons) Reference
Chitin 1.0 x 106
2.0 x 106
Muzzarelli 1976
Morganti 2008
Collagen 1.4 x 105
3.0 x 105
Ying 2001
Worthington Biochemicals 2009

48
peptide bonds join amino acid residues. The availability of different linkages means that the
preferred conformation or shape of the polysaccharide is dependent on the glycosidic bond, in
addition to the nature of the residues themselves. For example, glucose residues, linked α1-4,
produce a polymer with a shape and solubility quite distinctive from one with identical residues,
linked 1-4, but with a β-configuration (Smith & Wood, 1991). Table 3-4 lists molecular weights
of chitin and collagen.
Protein and Polysaccharide Interactions
Chitin and collagen perform similar structural roles in animals. Collagen often exists in
vertebrates as a complex of ordered collagen (protein) molecules in a matrix of water and
polysaccharide. On the other hand, chitin consists of poly-N-acetyl-glucosamine
(polysaccharide) chains which aggregate into crystallites, these crystallites being embedded in a
matrix of water and protein (Wainwright, 1982).
Chitin always occurs associated with proteins (Trim, 1941)and no sample of purified chitin
has been found to be free of amino acids (Wright & Arthur, 1987). The precise nature of the
interaction of cuticular proteins with chitin fibers and a detailed structure has not yet been
resolved. Certain sequence motifs occur in cuticular proteins from even distantly related species
and such conserved motifs have common and important roles for the proper function of cuticle.
Further work is needed to learn if the assignment of protein class to cuticle type is universal
(Trim, 1941). The relative proportions of protein and chitin in cuticles change during processes
such as cuticulogenesis (Wright & Arthur, 1987).

49
A few models attempt to describe the chitin-protein complex. While studying a rubber like
cuticle (locust prealar arm), Neville compared resilin and chitin content. From the measurements
of the unit cell and the diameter of the crystallite, calculation the percentage of chitin present in
the sample were based on 3 assumptions: (1) the shape of the crystallite in cross-section - round,
square or rectangular; (2) the degree of shrinkage of the specimen during preparation; and (3) the
amount of chitin which is 'disorganized' and outside the periphery of the observed crystallite. He
hypothesized that little chitin is found outside the crystallite, though the possibility that some
may be present cannot be completely eliminated. For cuticle to behave as a composite material it
would be expected that the chitin crystallites are strongly bonded to the protein matrix, and there
is some chemical evidence for this (Neville A. C., 1976). Removal of protein from the cuticle
reduced the heavily stained matrix. Neville, when examining the staining properties of structures
composed of chitin and protein resilin, concluded that cuticle is composed of microfibrils of
chitin in a matrix of proteins. High resolution micrographs of cuticular microfibrils show them
to be poorly stained, not unstained as is usually stated, which indicates that either chitin takes up
some stain or the presence of other material, e.g. protein. Because of the strong bonding between
chitin chains it is assumed that the microfibrillar core consists of crystalline chitin and the only
the peripheral chains are associated with protein. Proteins may also be associated with chitin
chains within the core. Although the microfibrils in many cuticle contain chitin, there is no
convincing evidence that any microfibrilar core is pure chitin (Wright & Arthur, 1987). The
presence of proteoglycan coats around fibrils is very likely in most arthropod carapaces, insects
and crustaceans, and this could be the origin of the examples of epidermal control on fibril

50
Figure 3-4 Arrangement of the protein subunits around the chitin core in the microfibril
perpendicular to the fiber axis and along the fiber axis. The 61 helix of protein subunits repeats
in 3.06nm (Blackwell & Weih, 1980).
direction. The complex transformations of filaments after their secretion by the epidermis in crab
cuticle suggest the presence of associated macromolecules which could be involved in the
control of fibril diameters and of their diverse associations (Bouligand Y. , 2008).
Blackwell and Weih have developed a model for the chitin-protein complex as it occurs in
ovipositors of the ichneumon flies Megarhyssa lunator and M. atrata. The model shows a sheath
of proteins subunits arranged in a helix (6 subunits per turn) around a chitin core, figure 3-4. Not
all cuticular proteins are bound to chitin, some form the protein matrix in which is embedded the
chitin-protein complex. The chitin-protein complexes are ordered structures (Blackwell & Weih,
1980), (Wright & Arthur, 1987). More elaborate modelling and experimental future work is
needed both to correlate the models of chitin–protein interaction proposed with the model
proposed by Blackwell and Weih (Blackwell & Weih, 1980). It is also evident from the data

51
overall that each species has its own characteristic protein binding matrix. Apparently there is no
simple relation between chitin content and the amount of covalently bound protein in the chitin-
protein complex (Austin, Brine, Castle, & Zikakis, 1981).
Proteoglycans
Collagen fibrils of extracellular matrices are known to be surrounded by a continuous coat
made of proteins and polysaccharides, called proteoglycans. This coat is quasi-fractal
construction, associating long chains of hyaluronic acid, with attached globular and linear
proteins, and also second-order lateral chains of chondroitin and keratan sulfates. These branched
structures can be arranged to form a cylindrical protection, a sort of sheath coat around each
collagen fibril. Bouligand theorized that: (1) this was possibly selected as being well devised to
stop the assembly of collagen molecules at a given diameter of fibrils; (2) this might be also a
way to have equal interdistances between fibrils and therefore an even distribution of minerals
between fibrils, as required in fiber-reinforced composites; and (3) these proteoglycans could be
involved in the cell control of fibril direction. This direction is initiated by the liquid crystalline
self-assembly, but might be modulated by links associating the cytoskeleton with integrins,
fibronectins and finally this proteoglycan coat and the collagen itself (Bouligand Y. , 2008). The
specific relationship of proteoglycans with the collagen fibrils in sclera closely resembled that
previously described in tendon and in articular cartilage, lending support to the view that the
association of proteoglycans with collagen may be consistent in a majority of connective tissues,
irrespective of their diverse functional specializations (Young, 1985).

52
Figure 3-5 Proteoglycan aggregate (Kierszenbaum, 2007).
Figure 3-5 shows that the extracellular matrix is comprised of proteoglycans attached to a
backbone of hyaluronic acid that is intertwined among collagen fibrils. Proteoglycans have both
chondroitin-sulfate- and keratin-sulfate-rich regions and link proteins facilitate binding of
hyaluronic acid.
Divergent views also exist as to how the carbohydrate portion is linked to collagen.
Different opinions have been expressed as to the functional role of carbohydrates in the collagen
structure. According to some researchers, the bulky carbohydrate group might direct the regular
stagger of tropocollagen molecules by requiring a particular fit. The polysaccharides of
connective tissue constitute the aqueous phase or ground substance of the extracellular space in
which collagen fibrils are embedded. The ground substance is believed to perform many
functions in the tissue. It provides the actual homeostatic environment of the cells and takes up
their metabolites and acts as a barrier against bacterial invasion. The proportion of the various
polysaccharides in ground substance varies in different tissues. The polysaccharides complexes

53
of connective tissue can be divided into two groups: (1) the glycoproteins consisting of protein
molecules to which monosaccharides or oligosaccharides are covalently bound; and (2),
proteoglycans consisting of polysaccharide-protein complexes, in which the polysaccharide
makes up a major part of the whole molecule (Chichester, 1982). List of polysaccharides or
glycosaminoglycan include hyaluronic acid, chondroitin, chondroitin 4-sulfate, chondroitin 6-
sulfate, dermatan 4-sulfate, keratin sulfate, heparin, and heparin sulfate. These glycosamino-
glycan imparts some degree of flexibility to calcified tissue like bone. The functioning of the
various tissues depends not only on the amount and types of collagen but also on the relative
proportion of other associated constituents. The mucopolysaccharides, associated with collagen,
are largely responsible for the water sorptive and retentive capacity of the tissues and for the
creation of osmotic pressures, at physiological concentration of a magnitude that is important to
the living organism. Polysaccharides also have the ability to affect the diffusion of ions through
the connective tissues (Chichester, 1982).
In the animal kingdom the fibers are usually proteins --- collagen. The fibrous soluble
polymer appears always to be predominantly carbohydrate, although of very varying
composition, especially in the invertebrates. This pattern, of a fiber-reinforced composite
material, has great versatility. By changing the ratio of fiber to soluble polymer, or by altering
the spatial relations between the two, quite different situations can be coped with, without
altering the chemical composition of the participants. Of the two elements, the fibers have
tensile strength, i.e. resistance to forces which would extend them. The interfibrillar polymer
expands the meshwork, and thus provides resistance to compressive forces. Movement of small

54
molecules can take place more or less freely in the polymer compartment, to sustain life in the
cells of the matrix. Apart from their physical roles proteoglycans and glycosamino-glycans are
able to interact physicochemically with, for example, collagen in ways which show considerable
specificity, and which presumably are important in the laying down of the fibrous network as
well as in maintaining its mechanical integrity. Systems of insoluble fibrils and soluble polymers
take stresses of movement and the maintenance of shape (Scott J. E., 1975). Chondroitin
sulphates are produced by sponges, crabs and mammals. The extent to which the sizes and
shapes of the polymer molecules correspond to those of the holes in the fiber meshwork is
clearly important in determining the properties of the tissue. Small-scale or very flexible
polymers would be able to move bodily through the meshwork, resulting in an easily deformable,
inelastic matrix.
Sulphated polysaccharides/glycosaminoglycans can change their structure much more
flexibly than a protein β-sheet. Determining the crystal structure in vitro is still quite problematic
because the crystal structure is influenced by temperature, calcium ion concentration, the
concentration of negative charged acidic macromolecules, carbonate ions, water, and other
cations. Sulphate groups for calcification mostly involve sulphated GAG (and some sulphated
glycoproteins), and these are certainly water soluble (Furuhashi, Schwarzinger, Miksik, Smrx, &
Beran, 2009).
Among the sulfated glycosaminoglycans, heparan sulfates, dermatan sulfates, and chondroitin
sulfates seem to be ubiquitous components of mammalian and other vertebrate tissues as well as
of a variety of cells in culture (Nader, et al., 1984). It has been suggested (Scott & Tigwell, 1978)

55
that hyaluronate and chondroitin sulphate differ from dermatan sulphate in being able to form
interresidue hydrogen-bonded sequences along the carbohydrate chain, which stiffen them.
Chondroitin sulphate and hyaluronate are thus particularly suited to occupying and maintaining
large domains in solution against deforming pressures. The creation of a 'buffer volume' between
the thin fibres that serves to keep them well separated and relatively free of anastomosis may be
a manifestation of this property. As a corollary, dermatan sulphate has more capacity to form
intermolecular liaisons, since it is not involved in intramolecular bonding to the same extent. Its
role in binding specifically to collagen may be aspect of this capacity (Scott, Orford, & W,
1981).
Chondroitin sulphate proteoglycans (CSPGs) in mineralized bone tissue of human, bovine
and other species have been isolated and characterized. It has also been suggested by an electron
microscopy that core proteins of proteoglycans interact with collagen fibrils. Since the occupied
region of collagen fibrils is suggested to be an initiation site of mineralization, proteoglycans
possibly inhibit the mineralization process (Takeuchi, Matsumoto, Ogata, & Shishiba, 1990).
Processed extracellularly by proteolytic cleavages at the N- and C-terminals, collagens are
biosynthesized as procollagens before aggregating efficiently to fibrils. There exist a number of
stages at which collagens might interact with proteoglycans, from procollagen, through various
aggregates, to the complete fibril. The first stages are in solution while the later interactions are
surface phenomena (Scott J. E., 1988).
The proteoglycans are soluble polymers in the simple scheme of connective tissues
consisting of a polypeptide 'core' attached to one or more glycan chains (Poole, 1986).

56
Consisting of repeating disaccharide units , in which one residue is always hexosamine, the
glycan chains are of three types, (1) heparan, (2) keratan, and (3) chondroitin dermatan.
Chondroitin-dermatan glycans contain N-acetylgalactosamine, with D-glucuronic and/or L-
iduronic acid, but contains no L-iduronic acid. Found in all connective tissues, a fourth glycan,
hyaluronan, similar in structure to the chondroitins. Hyaluronan is not attached to a peptide
chain, nor is it sulphated; but it contains glucuronic acid and N-acetylglucosamine and is usually
of molecular mass 105-l07 Da (Scott J. E., 1988), (Stern, Asari, & Sugahara, 2006).
Scott reviewed several model experiments dealing with glycosaminoglycans and summarized
a consensus of the findings: (1) Glycosaminoglycans interact electrostatically with type I
collagen under physiological conditions. Additional short range bonding cannot be excluded.
Linear charge density is important, as is glycosaminoglycan shape (determined by iduronate:
glucuronate ratios). (2) Strongly interacting glycosaminoglycans accelerate fibrillogenesis. (3)
Glycosaminoglycans are not incorporated into types I and II collagen fibrils. (4) Chondroitin
sulphate and dermatan sulphate proteoglycans interact strongly with type I collagen,
electrostatically via glycosaminoglycan chains, and by protein-protein interactions. (5)
Chondroitin sulphate and dermatan sulphate proteoglycans are incorporated into type I collagen
precipitates, if present during fibril growth. (6) Chondroitin sulphate and dermatan sulphate
proteoglycans inhibit fibrillogenesis (Scott J. E., 1988).
Tissues composed of collagen type I possess small amounts of proteoglycans which contain
almost exclusively dermatan sulfate; while tissues containing only collagen type II have high
amounts of chondroitin sulfates. The interaction among macromolecules is also important in the

57
extracellular matrix, and point at collagen-proteoglycan interaction as being essential to tissue
resiliency (Junqueira & Montes, 1983).
Hierarchy
Structural order in collagen occurs at several discrete levels. The primary structure denotes
the complete sequence of amino acids along each of three polypeptide chains. The secondary
structure is the local configuration of a polypeptide chain (Ratner, Hoffman, Schoen, & Lemons,
1996). An essential feature for the formation of the triple helical structure is that in the central
part every third position along the chains is occupied by glycine, which leads to the tripeptide
unit Gly-X-Y (Kuhn & Glanville, 1980). Tertiary structure refers to the global configuration of
the polypeptide chain into the triple helical collagen molecule. The fourth-order or quaternary
structure denotes the repeating supermolecular unit structure, comprising several molecules
packed in a specific lattice, which constitutes the basic element of the five molecule microfibril.
Several microfibrils aggregate end to end and also laterally to form a collagen fiber. Collagen
fibers exhibit a characteristic banding pattern with a period of about 65 nm (Ratner, Hoffman,
Schoen, & Lemons, 1996).
The exoskeleton materials of arthropods are complex composites that are hierarchically
structured and multifunctional. The linear chitin chains align anti-parallel and form α-chitin
crystals at the molecular level. Several of α-chitin crystals which are wrapped by proteins form
nanofibrils of about 2–5 nm in diameter and 300 nm in length. A bundle of chitin–protein

58
Figure 3-6 Hierarchical levels of collagen I in bone and α-chitin-protein matrix in crab
exoskeleton. Depicting points of differences in the types of bonds and triple helix structure of
collagen and linear structure of chitin.
Molecular
unit
Peptide
bonds
Nanofibrils
3-chain
coiled
helix
Aggregation of
fibers
Fibrous Bundles
Structure
Linear
Crystals
Glycosidic
bonds
Collagen Type I α-Chitin

59
nanofibrils then form chitin–protein fibers of about 50–100 nm in diameter. These chitin–protein
fibers align together forming planar layers which stack up helicoidally (Meyers, Chen, Lin, &
Seki, 2008). This structure is called a twisted plywood or Bouligand structure (Bouligand Y. ,
1972).
The basic hierarchical sequence for both collagen and chitin is Molecule: Chain of molecules:
Microfibril: Fibril Aggregation: Fibrous Bundle: ‘Twisted Plywood’. The major difference in
the hierarchical scheme lies within the chain of molecules. Collagen has peptide bond that create
a triple helix while chitin has glycosidic bonds that creates a linear crystal structure. See figure 3-
6.
Liquid Crystal Characteristics
All biological structures form in an environment rich in water. How rigid crystalline
structures are formed in biologically aqueous environments is not fully understood. Chemical
reactions in water can produce a precipitate that is insoluble in water. Crystalline structures can
form in water if the concentration of the substance exceeds its saturation point. The crystalline
precipitate can form a rigid structure under certain conditions. In fact, such processes are
biologically important, especially in the formation of crystalline structures, such as bone, cuticle,
and their building blocks.
Biological solid structures do not allow molecules to diffuse around so that all the chemical
and physical processes necessary for life can take place. Fluids allow molecules to move around

60
in a way that allows interactions with other molecules. Succinctly, biological structures must be
rigid enough to add stability and function properly as well as fluid enough to allow the
movement of molecules and all the necessary processes to take place quickly. Many biological
structures achieve a delicate balance between rigidity, order, and fluidity by liquid crystalline
(LC) structures. Membranes surrounding the outer cell, the nucleus, mitochondria, and the
endoplasmic reticulum of animal cells demonstrate this balance and serve many functions: (1)
confines the contents (rigidity); (2) discriminate between various substances to be transported
(order); and (3) control the quick movements of molecules and rapid reactions (fluidity).
Therefore, it is a safe assertion that liquid crystalline structures are major building blocks within
animal cells.
In the liquid crystal phase, molecules move freely about in much the same manner as in a
liquid, but tend to remain oriented in a certain direction. This partial alignment represents a
degree of order not present in liquids. States of matter are different from each other because the
molecules in each state possess different amounts of positional order and orientational order. In a
solid, the molecules are constrained to occupy only certain positions, creating a phase possessing
positional order. Additionally, the specific positioned molecules in a solid are constrained in
how they orient themselves with respect to other molecules, creating a phase possessing
orientational order. Both positional and orientational orders are lost completely, when a solid
melts to a liquid and the molecules chaotically move and tumble. However, when a solid melts
to a liquid crystal, the positional order may be lost but some of the orientational order remains.

61
Molecules likely to be liquid crystalline at a certain temperature: (1) typically are elongated in
shape (significantly longer than it is wide); (2) have some rigidity in its central region; (3) tend to
have somewhat flexible ends. A representative model of a typical liquid crystal molecule is
depicted in figure 3-7. Elongated molecules customarily have stronger attractive forces when
aligned parallel to one another and they bump into each other less when they all tend to point in
the same direction. The flexibility appears to allow one molecule to position itself more easily
between other molecules as they all randomly move about (Collings, 2002).
Liquid crystals can be classified by their dependence on temperature and/or concentration.
Thermotropic LCs present phase transitions, as temperature is changed. When the liquid crystal
phase is dependent on the concentration and temperature of the LC molecules in a solvent, it is
called a lyotropic LC. A classic example of a lyotropic liquid crystal mixture is one with a
molecule that has end groups with different properties; one end of the molecule shows an affinity
for water, while the other end tends to exclude water. Both thermotropic and lyotropic LCs
consist of organic molecules. Although many compounds exhibit only one LC phase, it is not
unusual for a single substance to possess more than one (Collings, 2002).
Membranes surrounding the outer cell, the nucleus, mitochondria, and the endoplasmic
reticulum of animal cells owe its structure to the LC nature of the bilayer phospholipid/water
mixture. Amphiphilic molecules are composed of hydrophobic (water-fearing) and hydrophilic
(water-loving) parts; and are also known as surfactants. As the amount of amphiphilic (loving

62
Figure 3-7 Representation of a typical liquid crystal model.
both kinds) material is mixed with water, it is possible for: (1) a small amount to go into
solution; and (2) an increasing amount to form micelles or vesicles. The fluid mosaic model is
the standard model for the cell membrane. The concentration of one constituent with respect to
the other is far more important in these lyotropic LCs.
Three liquid crystals classifications are relevant to biological composites: nematic,
cholesteric, and blue-phase, figure 3-8. Long rod-shaped molecules line up in parallel in nematic
liquid crystals. In cholesteric liquid crystals (twisted or chiral nematic), long rod-shaped
molecules have helicoidal distribution that is only apparently layered; in a minute volume the
molecules run almost in parallel. Nonetheless, there is a slight angular change between
neighboring molecules, always in one sense of direction for a specified set of conditions. Blue
phase liquid crystals are similar to cholesteric; however the twist is cylindrical, whereas the twist
is planar in cholesteric liquid crystals. Changes in temperature or in concentration may bring
about a change in plane adopted (Neville A. , 1993). Although many compounds exhibit only
one liquid crystal phase, it is not unusual for a single substance to possess more than one. These
liquid crystal phases are formed by rodlike molecules, the most common and well known LC
phases (Collings, 2002). In 1977, researchers in Indian discovered disk-like molecules also form

63
liquid crystal phases and are called them discotic LCs (Chandrasekhar, Sadashiva, & Suresh,
1977).
There are indications that the crystalline ordering of polymer material and proteins is the first
step in the creation of very hard biological structures such as cuticle and bone. The organic
portions of cuticle and bone are chitin and collagen, respectively. These organic polymers form
fibrous material that share many characteristics of LCs. For example, liquid crystals and fibrous
composites are both mobile, have similar architectures or geometries, can self-assemble, and
have similar optical properties. Fibrous materials can be composed of a single polymer or a
mixture of polymers (Neville A. , 1993).
A polymer is any large molecule composed of a long sequence of repeating units. The
monomer is the basic building block and is the unit repeated in the molecule. Proceeding in a
variety of ways, the polymerization process is the chemical reaction in which these bonds
between monomers form. The polymerization process stops when the number of reactive end
groups decreases to the point where there is little chance for further reactions between them.
Polymer LCs can achieve extremely high orientational order; respond to electric and magnetic
fields; are relatively inexpensive and stable; and can be easily fabricated into thin films.
Chitin and collagen are polymer LCs. Chitin is a polymer consisting of a linear chain of
repeating units. The unit is derived from N-acetyl-glucosamine monomer. Notice that there are
chemical entities (-H, -OH, and -CH2OH) attached to the central part of the monomer. As
reactions occur with chitin’s chemical entities, the linear chain adopts a helical structure, gaining
rigidity and the capacity to produce LC phases. Amino acids form the basic building block for

64
the collagen polymer LC. A typical amino acid possesses two quite different reactive groups.
The carboxyl group (-COOH) on the end of one amino acid can react with the amino group (-
NH2) on the end of another amino acid to form a chain of amino acids. The resulting
macromolecules are called polypeptides. The collagen structure is basically a triple helical
Figure 3-8 Depiction of rod-like liquid crystal phases: smectic, nematic, blue phase, and
cholesteric or chiral nematic.
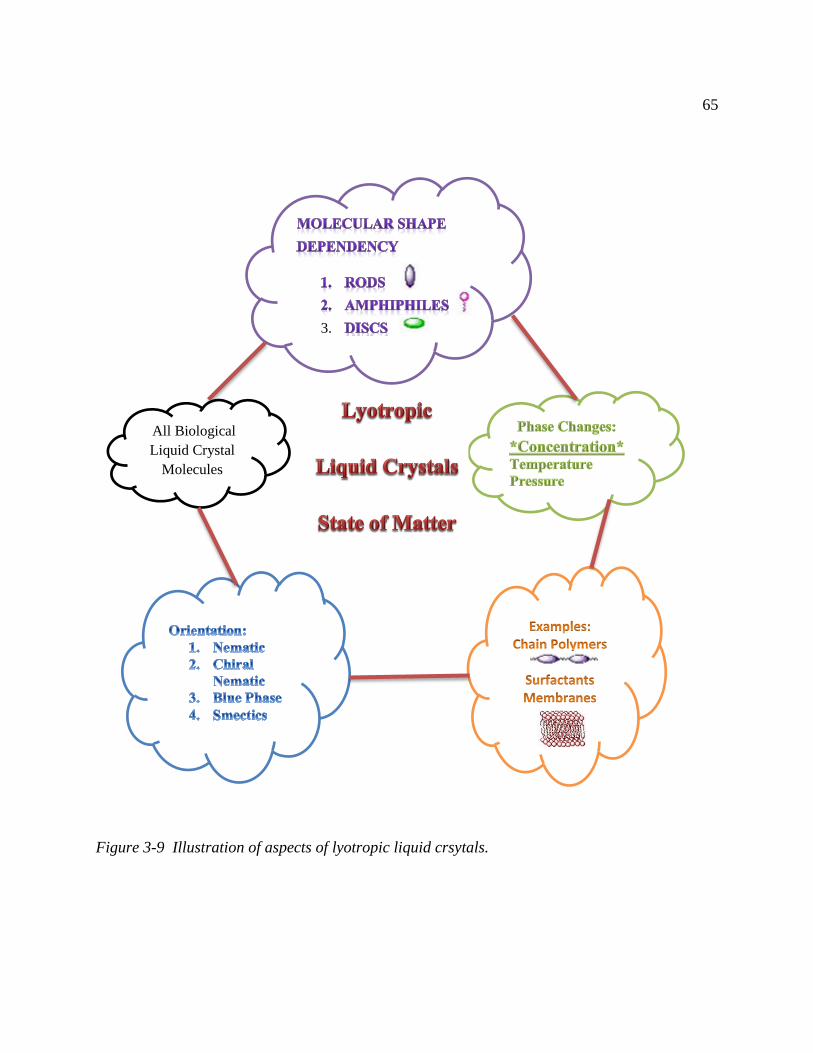
65
Figure 3-9 Illustration of aspects of lyotropic liquid crsytals.
3.
All Biological
Liquid Crystal
Molecules

66
chain, which causes the macromolecule to be fairly rigid and capable of forming LCs. Because
these macromolecules have helical structures, they interact in a fashion which prefers that the
molecules not be exactly parallel to one another. Consequently, some molecules form chiral
nematic phases. The forces between these large molecules are due to the helical molecules
themselves and to the interactions between these large molecules and the solvent molecules
(Collings, 2002). Chitin and collagen macromolecules must dissolve in the solvent. LC phases
only form when the concentration is high enough so that these macromolecules constantly
interact with one another. This cannot be achieved if the macromolecules do not dissolve in the
solvent to high enough concentrations (Collings, 2002). Chitin and collagen fulfill
therequirements of rigidity and of being solvent-dissolved; thus, they, like all biological
structures, are lyotropics LCs. Characteristics of lyotropics LCs is depicted in figure 3-9.
Works published by Bouligand, Giraud-Guille, and Neville reviewed liquid crystalline order
in biological materials such as chitin, collagen, and other fibrous composites (Bouligand Y. ,
1972), (Giraud-Guille M.-M. , 1994), (Giraud-Guille M.-M. , 1992), (Giraud-Guille M. M.,
1996), (Giraud Guille, Mosser, Helary, & Eglin, 2005), (Neville & Luke, 1971). After studying
fibrous structures such as the cuticles of crabs, Bouligand (1972) found that several fibrous
biological materials are: (1) made of fibrils arranged in highly organized geometric patterns ---
‘twisted plywood model’; (2) solids with elongated molecules that show a strong anisotropy; (3)
still more or less fluid phases between two well defined temperatures and may show
birefringence; and (4) capable of forming LCs in solutions. These vary among the factors that led
Bouligand to postulate that the secretions of fibrous structures pass through a brief liquid phase

67
providing an intriguing mechanism in morphogenesis for certain fibrous materials. Neville also
suggested biological fibrous composites might have developed by self-assembly via a liquid
crystalline phase (Neville A. , 1993). This idea was further strengthen by the work of Giruad-
Guille (1989, 1992). Despite their similarities, notable differences exist between biological
fibrous composites and liquid crystals. The fibrous component consists of small parallel bundles
of molecules (crystallites), while liquid crystals consist of individual molecular chains. Another
difference is that biological composites that have reached fruition exist in a solid state, while
liquid crystals exist in a state with properties intermediate between those of a solid and liquid
(Wright & Arthur, 1987).
The structure of many other biological materials is undeniably related to that of liquid
crystals, despite they are not at all fluid. In most skeletal tissues, the extracellular matrices are
made up of fibrils which also are stabilized analogues of either nematic or cholesteric liquid
crystals (Bouligand Y. , 2008). Cuticles also fall into this class, non-lamellae cuticle (preferred
orientation of microfibril) corresponds to the nematic phase and helicoidal model to the
cholesteric phase (Wright & Arthur, 1987). They are considered as stabilized analogues of liquid
crystals.
Liquid crystalline self-assembly was observed in concentrated solutions of the main polymers
extracted and purified from these materials: chitin and collagen. Liquid crystals appear to be
involved in most physical aspects of biological morphogenesis. Patterns draw their origin from
properties of liquid crystals, mainly geometrical. Certain patterns are very common in thin
sections of biological tissues. The ‘‘arced fibrils’’ were first described in carapaces of

68
arthropods, mainly insects and crustaceans. The crab carapace also called cuticle is strongly
mineralized, and samples must be decalcified in general before dehydration, embedding and
microtomy. The thin section crustaceans showed a fibrous material made of superposed lamellae,
each one being made of a long series of nested arcs, with concavities oriented along a common
direction. The fibrils are made of chitin, a nitrogenous polysaccharide, associated with diverse
proteins. This was rather surprising to observe a similar organization in so distant biological
materials, differing by their chemical composition (chitin, collagen, DNA), their function and
their phylogenetic position (Bouligand Y. , 2008).
In arthropod cuticle, the helicoidal building block is the chitin polymer. Within each
horizontal layer these rod shaped polymers are packed parallel to each other. These layers of
parallel rods are stacked on top of one another, each layer rotated slightly to adjacent layers. The
angle of rotation between the layers is more or less constant, giving a regular helicoidal structure.
The 3D structure is reminiscent of plywood. This multiply construction gives the helicoid the
ability to resist strain in most directions (Murray & Neville, 1997). Similar patterns were
observed in the sections of compact bone in TEM. The collagen network appears very dense,
with fibrils either arranged in two alternating directions forming an orthogonal plywood, or
regularly changing orientation within the section plane, forming arced patterns. Thus, two
osteon models coexist in compact bone; the first obeys the classical description of osteons
forming orthogonal plywood, the second follows the twisted plywood model, where collagen
fibrils show small and regular changes in their orientation. Arced figures are a direct
consequence of what is described as the twisted plywood model. Parallel and equidistant straight

69
lines represent the molecular directions and from one level to the next, the lines turn by a small
and constant angle; series of nested arcs appear on oblique sides of the model (Giraud Guille,
Mosser, Helary, & Eglin, 2005).
In compact bone osteons, the collagen fibrils form arced patterns as a consequence of their
three-dimensional organization; this structure is analogous to a geometry found in certain liquid
crystals (Giraud-Guille M.-M. , 1992). These self-assemblies were analyzed between crossed
polars in optical microscopy since they form birefringent fluids over long distances. Self-
assembled liquid crystalline phases of collagen generated crimp morphologies (Giraud-Guille &
Besseau, 1998). This characteristic and striking architecture must have a common physical origin
in these phyletically and functionally very different biological materials (Bouligand Y. , 1972).
Bouligand discussed a possible mechanism for a liquid crystalline secretion in
crustaceans and in other arthropod cuticles. Bouligand postulated that the fibrous matrix is
produced by a secretion which is a highly concentrated solution of chitin and proteins. Thin
filaments which associate these two components differentiate at the epidermal cells’ membrane
surface, and align in this plane. This corresponds to an ordered secretion which is liquid
crystalline, since alignment and fluidity are realized at the close contact of epidermis. The
parallelism of filaments with the epidermis involves also that of cholesteric layers themselves
with this epidermis. Bouligand believed that cell membranes create strong boundary conditions,
with a considerable influence on textures. The cuticle layers lie parallel or nearly parallel to the
epidermis and rapidly the shape of the epidermis is maintained by the cuticle itself. Cells
maintain a control at the time of the liquid crystalline secretion, mainly by the faculty of

70
changing molecular orientations, through the interactions between the cytoskeleton and the
extracellular matrices, across some specialized areas of cell membranes. Questions still remain
concerning the differentiation of biological analogues of cholesteric liquid crystals, what comes
from liquid crystallinity of the initial secretion, and what is controlled by cells. As research
continues on liquid crystalline phases of fibrous materials, progression to the answers is expected
(Bouligand Y. , 2008).
Neville (1993) reported that ultrastructural observations of arthropod cuticles above the
epidermal cells reveal uniform series of arced patterns, attesting to fluidity at the time of
Figure 3-10 Liquid crystalline organization of collagen at different concentrations.
Isotropic
20-50 mg/ml
Loose
Cholesteric
50-400
mg/ml
Dense
Cholesteric
400-800
mg/ml
Banded
Structures
800-1000
mg/ml

71
secretion. Indeed, products of neighboring cells join up neatly if the system is liquid; otherwise,
many irregularities and faults would appear along the cell boundaries. Directly visible on the
oblique sides of the pyramid are what appear to be superposed series of parallel nested arcs. The
concavities of the arcs are reversed on opposite sides of the model. In biological systems this
particular geometry has often been described as a twisted plywood, but depending on its
appearance in optical or electron microscopy, or on the research field of the authors, a variety of
Table 3-5 Description of Cholesteric Geometries
Descriptions of Cholesteric Geometries
Terms used by physicists or chemists (liquid crystal and polymer field)
Cholesteric mesophases
Cholesteric texture
Chiral nematic structure
Terms used by microscopists (aspect in optical or electron microscopy)
Fingerprint-like patterns
Arced patterns
Terminology introduced by Bouligund and colleagues in France
Twisted plywood
Multidirectional plywood
Twisted fibrillar phase
Terminology introduced by Neville and colleagues in the United Kingdom
Helicoidal
Helicoidal architecture
Helicoidal arrays
(Giraud-Guille M. M., 1996)

72
descriptions of the cholesteric structure are found in the literature, table 3-5. The possibility of
slowly increasing the collagen concentration of a solution has allowed the following of the
consequent changes in the organization of collagen, figure 3-10. This work shows that a
relationship exists between collagen concentration and the diameter of fibrils and the discrete
rotational angle of subsequent layers in collagen rich matrices (Mosser, Anglo, Helary,
Bouligand, & Giraud-Guille, 2006).
Collagen and chitin can readily form analogues of amphiphilic molecules due to the presence
of reactive groups on its backbone. A variety of amphiphilic copolymers containing hydrophobic
and hydrophilic segments, are very active due to their spontaneous self-assembly behavior in
aqueous media. This is explained by the presence of a great number of different hydrophilic and
hydrophobic reactive groups in these structures. Hydrophilic groups include hydroxyl, carboxy
groups and hydrophobic groups include hydrocarbons chains. The items created by amphiphiles
can be rod-like. These anisotropic self-assembled structures can order themselves in a similar
fashion that liquid crystals do, forming large-scale versions of LC phases.
Amphiphilic properties of polysaccharides such as chitin can be varied by manipulating their
hydrophilic and hydrophobic groups. Since most polysaccharides are polyelectrolytes. Their
hydrophilicity can be controlled by varying charge density and their hydrophobicity can be
controlled by the introduction of branches (alkylation, acylation, grafting, crosslinking)
(Desbrieres & Babak, 2008). Amphiphilic chitin derivatives are soluble in both organic and
aqueous solvents and under appropriate conditions they are able to self-assemble (Aranaz,
Harris, & Heras, 2010).

73
The term ‘‘peptide-amphiphile’’ can be used to describe amphiphilic peptides consisting only
of amino acids such as collagen. It has been shown that linking peptides to synthetic hydrophobic
tails enables them to self-assemble into biomimetic films with highly organized interfaces, and
enhances their ability to attain well-defined and stabilized three-dimensional structures that
effectively promote cell adhesion, spreading, migration, growth and differentiation. By changing
any of the structural segments of an amphiphilic molecule, one can control the morphology,
characteristics, surface chemistry, and function of the molecule. Amphiphiles can self-assemble
into a variety of different structures such as micelles, vesicles, bilayers, ribbons, twisted ribbons,
and patterned membranes to minimize unfavorable interactions with their surroundings. Triple-
helical peptide-amphiphiles are very stable and such model collagen peptide-amphiphiles follow
some of the trends observed for other more common surfactants. The long alkyl chains
characteristic of peptide-amphiphiles provide a convenient mechanism for self-assembly and
association with other hydrophobic substrates. Enabling optimal presentation of active binding
sites, the composition and orientation of molecules on the surface is well controlled. The self-
assembly of the hydrophobic tails of the peptide-amphiphiles is a driving force (hydrophobic
effect) that stabilizes the three-dimensional structure of the peptide headgroup into triple helices,
and a-helices, giving rise to protein-like molecular architectures (Kokkoli, et al., 2006).
Bouligand, Neville and Giraud-Guille outlined a progression of phases that collagen and
chitin undergo to create ordered, fibrous structures. The ordering seems to be in layers, with the
preferred direction in each layer rotating slightly in going from layer to layer. At first the order is
liquid crystalline, but as the concentration of the polymer and protein increases, the material

74
becomes more of a gel. The more rigid a molecule is, the less soluble it is. Crystalline bundles of
parallel polymers can also form, causing the material to harden. Such a substrate seems to be
necessary for minerals to begin to accumulate, thus allowing shells, bone, etc., to form (Collings,
2002).
Synthesis
The biosynthesis and degradation of both collagen and chitin are stepwise procedures
governed by enzymatic activity; these pathways consist of catalytic units, precursors,
biotransformations, polymerization, hydrolysis, and fibrillogenesis. Both collagen and chitin
share many common properties at the molecular level of organization. For example, they both
have components formed in the intracellular and in the extracellular space during the synthesis
process. Both synthesis pathways involve changes in the cytoplasmic matrix, the rough
endoplasmic reticulum, processing in the Golgi apparatus and transport through vesicles
culminating in exocytosis into an assembly zone. Both use enzymes to transform. In the
assembly zone, both collagen and chitin molecules form fibrous, hierarchical structures. Both of
these extracellular fibrous molecules are synthesized in a precursor form and their active
structure is polymeric. Furthermore, the resulting tissue formation, i.e. bone for collagen and
cuticle for chitin, follow a common developmental course, namely that the major organic
constituent collagen and chitin are synthesized, extruded from the cell, and then self-assembled
in the extracellular space before mineralization begins. For this reason, bone and cuticles are
good examples of an ‘‘organic matrix-mediated’’ mineralization process (Cui, Li, & Ge, 2007).

75
Collagen is synthesized by osteoblasts and fibroblast as well as smooth-muscle cells,
chondroblasts and odontoblasts. Osteoblasts synthesize only type I collagen (Kuhn & Glanville,
1980). The biological synthesis, secretion, and assembly into definitive extracellular structures of
collagen can be described briefly as follows. Pre-procollagen chain is the starting point for a
chain of bioconversions which include major steps such as non-lysosomal degradation,
hydroxylation, glycosylation, and disulphide bond formation. This metabolic pathway culminates
in the formation of procollagen. Collagen polypeptides are synthesized on the rough endoplasmic
reticulum as propolypeptides, over 100 amino acid long. The individual polypeptide chains,
called pre-procollagen chains, are synthesized on membrane-bound ribosomes in the cisternae of
the rough endoplasmic reticulum. These chains have three major domains: the α-chain, the
amino-terminal peptide, and the carboxy-terminal peptide. These pro-α-chains are modified after
synthesis by the removal of prepeptide sequences, by hydroxylation of proline and lysine, by
glycosylation of propeptides and by disulfide bond formation. Each α-chain has an amino
terminal and a carboxy terminal that direct the spontaneous assembly of the α-chains into the
triple helix, procollagen. After all these post-translational modifications take place, individual
procollagen molecules are transported to the Golgi apparatus, packaged into granules, and
secreted into the extracellular space by exocytosis. The collagen producing cells like fibroblasts,
for example, secrete enzymes which catalyze the hydrolysis of the extension peptides from
procollagen to produce the triple helix called tropocollagen – a rigid rod 300nm long and 1.5nm
in diameter. The extension peptides at both ends of the molecule must be cleaved before
polymerization can occur and the fibers can be assembled. Tropocollagen molecules

76
Figure 3-11 Collagen and Chitin Metabolism (Cohen, 1993).
ENZYMES
turnover
POLYMERIZATION
PRECURSORS Biotransformation:
Collagen
Hydroxylation
Glycosylation
Disulphide bond formation
Collagen
prolyl-3-hydroxylase
prolyl-4-hydroxylase
lysl hydroxylase
Chitin
chitin synthase
Synthesis Translocation Insertion Activation
Chitin
Phosphorylation
Amination
acetylation
Procollagen formation
UDP-GlcNAc formation
Collagen
Limited Proteolysis
(crystallization?, fibrillogenesis)
ORIENTATION OF MICROFIBRILS
Chitin Hydrogen Bond Formation (crystallization, fibrillogenesis)
SUPRAMOLECULAR ORGANIZATION
(covalent and noncovalent interactions)
(with proteins, with polysaccharides)
DEGRADATION
(Collagen: collagenases)
(Chitin: chitinase, chitobiase, chitodextrinase)
RECYCLING
MINERALIZATION

77
Figure 3-12 Chitin and Collagen Synthesis (Laurent, 1987).

78
spontaneously associate in a quarter-staggered manner, all aligning in the same direction, to form
microfibrils. Microfibrils pack closely together to form mature collagen fibers 50 nm thick with
the characteristic banding pattern (Smith & Wood, 1991).
The biosynthesis of the crustacean cuticle or exoskeleton involves the synthesis and assembly
of component lipids, carbohydrates, and proteins by the epithelial cell layer which lies just
underneath the cuticle. Chitin synthesis is directly governed by an enzymatic activity called
chitin synthase. To provide a flow of substrate molecules for the functional catalytic units, a
cascade of cytoplasmic biotransformations of sugar precursors takes place. Chitin synthesis in
the crustacean epithelial cell begins with the glycosylation of a protein acceptor to form a primer
molecule for chitin synthetase. Catalytic units synthesized at the endoplasmic reticulum
compartment are packaged in clusters which involve the Golgi apparatus. The assembled
multiunits are translocated and integrated into the plasma membrane at the apical compartments
of epidermal cells. Trehalose, glucose, or the polysaccharide glycogen, circulating in the
hemolymph, could be the starting points for a chain of bioconversions which include major steps
such as phosphorylation, amination, and acetylation. This metabolic pathway culminates in the
formation of UDP-GlcNAc, the ultimate substrate of chitin synthase. Transformation of fructose-
6-phosphate to glucosamine-6-phosphate by glutamine-fructose-6-phosphate aminotransferase
can be considered as the first turning point on the pathway to chitin synthesis. Next, there is
molecular organization of the chitin synthase units in the cell membrane, and an intricate process
which involve the translocation of chitin polymers across this barrier into the extracellular space
where chemical modification of the noncrystallized chitin occurs (Cohen, 1993). Synthesis of

79
chitin occurs either intracellularly or at the interphase with the extracellular medium. The
modified chitin molecule associates with other molecules like protein. It has become clear that
protein is covalently attached to the structural polysaccharide, chitin, in the crustacean. The last
step in chitin biotransformation involves the formation of microfibrils at the cell surface,
following the coagulation of adjacent developing chitin chains via extensive hydrogen bonding.
Chitin polymerization and crystallization are coupled; they are consecutive processes. The
resulting chitin-protein structure matures through formation of secondary covalent bonds among
its components, creating chitin fibers (Jolles & Muzzarelli, 1999). Studies done by Horst
supports the conclusion that crustacean chitin is secreted as a glycoprotein complex and that
concurrent protein synthesis is required for chitin deposition to continue (Horst, 1989). Liquid
crystal mobility may account for changes in orientation of the chitin microfibrils and, thus, for
the helicoidal appearance. The cyclic phenomenon of molting in crustaceans involves a
concerted and coordinated flow of hormonally-controlled biochemical events governed by
enzymatic axtivity. One such major event is the degradation of the endocuticular chitin, followed
by the subsequent recycling of the aminosugar monomers into newly-formed polymer units
(Cohen, 1993).
There are differences in the biotransformation processes involved in the synthesis of the
molecules secreted into the extracellular matrix of the cells that make collagen and chitin. The
biotransformation of collagen involves non-lysosomal degradation, hydroxylation, glycosylation,
and disulphide bond formation while chitin’s biotransformation involves phosphorylation

80
amination acetylation. While collagen undergoes limited proteolysis after polymerization, chitin
undergoes hydrogen bonding, figure 3-11.
Figure 3-12 is a schematic representation of cells showing the generalized pathway of
collagen and chitin biosynthesis. The sequential changes in the cytoplasmic matrix, RER, and
processing in the Golgi apparatus and transport through vesicles culminating in exocytosis into
the assembly zone are depicted (Laurent, 1987), (Cohen, 1993), (Jolles & Muzzarelli, 1999).
Properties
Collagen fibers transmit forces, dissipate energy, prevent premature mechanical failure and
provide biological signals to adjacent cells that regulate function. Moreover, collagen is
resorbable, it has high water affinity, low antigenicity, very good cell compatibility and ability to
promote tissue regeneration (Kolacna, Bakesova, Varga, Kostakova, & Planka, 2007). Isolated
collagen fibers have high tensile strength that is comparable to nylon (50-100MPa) and an elastic
modulus of approximately 1 GPa (Enderle, Blanchard, & Bronzino, 2005). Cross-linking
increases with age, and collagen becomes less flexible. Collagen often exists in combination with
other proteins.
The mechanical properties of the cuticle are conferred by the proportion of chitin, by the degree
of sclerotization and by the sequences of its proteins (Iconomidou, Willis, & Hamodrakas, 2005).
Chitin microfibrils, being composed of long molecular chains bound firmly to another,
contribute significantly to the mechanical strength of cuticles and their final orientation appears
to be related to the stresses which will be placed on the cuticle. The helicoidal model gives a

81
plywood structure to the cuticle, each ply (or sheet) being made up of high modulus chitinous
fibrils in a low modulus protein matrix. A plywood is a mechanically resistant material, which
associates several wood sheets, stuck together, with the grain of adjacent layers lying at right
angle to each other, so that the fibers follow two alternating directions (Bouligand Y. , 2008).
This would give a very strong structure, one which would resist bending forces in all
directions. On the other hand microfibrils with preferred orientation provide a structure strong in
tension (when the strain is on chitin) but weak in the direction normal to the chitin chains (when
the strain is on the protein) (Wright & Arthur, 1987). Examples of these arced patterns were
found in diverse tissues of animal and plant groups, mainly extracellular matrices, connective
and skeletal tissues, but also in several intracellular differentiations. Variations are considerable
within these materials, simply in the same crustacean carapace for instance, according to the
position in the body and also according to the distance from the outer surface. This is an aspect
of biodiversity, with considerable changes also between species. Research will be necessary in
this field, and the list of these twisted fibrous materials needs to be completed: the presence of
fibrils is often masked but some appropriate methods, as chemical treatments, staining and use of
contrast agents can make the arcs visible. Polarizing microscopy also is essential to detect these
structures (Bouligand Y. , 2008). Table 3-6 provides microscopic information regarding collagen
and chitin.
Collagen and chitin provide the backbone for mineralized tissue such as bone and crab
exoskeleton (Chen, et al., 2008). Similarites between liquid crystal and collagen and chitin were

82
Table 3-6 Collagen and chitin empirical formula, tissue distribution, microscopic appearance,
ultrastructure, synthesis site, interaction, and function.
(Junqueira & Montes, 1983)
realized by Bouligand, Giraud-Guille, and Neville (Bouligand Y. , 1972), (Giraud-Guille M.-M. ,
1992). Collagen and chitin readily form spherulites in vitro (Krebs, et al., 2004).Other common
features of collagen and chitin are that they are both biodegradable, biocompatible,
multifunctional, non-toxic, play a role in mechanical support, have limited reactivity; highly
insoluble and birefringement (Jolles & Muzzarelli, 1999), (Köster, Evans, Wong, & Pfohl,
2008).

83
Features unique to chitin are that it is a polysaccharide polymer found in crab exoskeleton,
has glycosidic bonds, has a linear chain, and has three forms exist (Jolles & Muzzarelli, 1999).
Another feature unique to chitin is that it is antibacterial. One study proclaimed that chitin
sutures resisted attack in bile, urine, and pancreatic juice (Dutta, Ravikumar, & Joydeep, 2002).
The fungicidal and bactericidal action of chitin appears to be mediated by the electrostatic forces
between the protonated NH2 group and the negative residues at cell surfaces. The number of
protonated NH2 groups present increases with increased degree of deacetylation (DD).
Therefore, the DD influences antimicrobial activity (Tsai, Su, Chen, & Pan, 2002). Features
unique to collagen are protein molecule, peptide bond, triple helix coiled chain with cross-
striation pattern, over 27 other types, found in bone (Köster, Evans, Wong, & Pfohl, 2008). It is
interesting that collagen is not antimicrobial like chitin. The simplest explanation can be
attributed to the structure of collagen; in that it does not afford enough electrostatic forces.
There also exist differences in the swelling behavior of chitin and collagen. Chitin does not
swell in water. Collagen may or may not swell in water depending on which tissue is used to
extract the collagen. For example, bone collagen (which normally calcifies) does not swell in
water whereas tendon collagen (which normally does not calcify) does (Bonucci, 1992). Several
crystalline forms of chitin have been ascribed and they differ in packing and polarity of the
adjacent chains. The failure of chitin to swell in water is explained by the very extensive inter-
and intramolecular bonding. This strong bonding leads to the formation of long microfibrils with
high tensile strengths and accounts for chitin’s fibrous character. The extents of the molecular

84
Figure 3-13 Collagen and chitin unique features and ones they share.
bonding differ in different cuticles which can lead to some variation in the observed chemical
and physical properties of the chitins present in different species (Wright & Arthur, 1987).
Figure 3-13 highlights common and unique features of collagen and chitin.

85
Applications
Collagen and chitin have numerous tissue engineering applications, ranging from structural to
metabolic organs (Monzack, Rodriguez, McCoy, Gu, & Masters, 2011), see figure 3-14. In
addition, a wide variety of other applications, figure 3-15, for chitin and collagen have been
reported over the last four decades (Enderle, Blanchard, & Bronzino, 2005), (Singh & Ray,
2000), (Ma & Elisseeff, 2006), (van Blitterswijk, 2007), (Hin, 2004).
The poor solubility of chitin is the major limiting factor in its utilization, and the investigation
of its properties and structure. Despite these limitations, various applications of chitin have been
reported in the literature. For example, fibers made of chitin have been useful as absorbable
sutures and wound-dressing materials. These chitin sutures resist attack in bile, urine, and
pancreatic juice, which are difficult with other absorbable sutures. It has also been claimed that
wound dressings made of chitin fibers accelerate the healing of wounds by up to 75% (Dutta,
Ravikumar, & Joydeep, Chitin and Chitosan for versatile Applications, 2002). Collagen has been
3 N HCl at the boil for 1.5 h. A typical ratio of 3 N HCl to chitin was 10 ml /g. After acid
viewed as an ideal biomaterial, as it is the major component of the extracellular matrix, and
because it can be processed into a wide variety of structures and shapes (sponges, fibers, films,
3D gels). A vast number of publications can be found in the literature, covering a diversity of
clinical applications, such as general surgery, orthopedics, cardiovascular, dermatology, urology,
dentistry, ophthalmology, plastic and reconstructive surgery. Although these examples
encourage applications of collagen in the biomedical arena, the risk of infection, its low
antigenicity and fast degradation when implanted in the human body are, to some extent, limiting

86
Figure 3-14 Collagen and Chitin/Chitosan Tissue Engineering Applications (Monzack,
Rodriguez, McCoy, Gu, & Masters, 2011).
Eye
Collagen
Chitin (Chitosan)
Muscle
Collagen
Chitin (Chitosan)
Tendon
Collagen
Chitin (Chitosan)
Bone and Cartilage
Collagen
Chitin (Chitosan)
Skin
Collagen
Chitin (Chitosan)
Vessels
Collagen
Chitin (Chitosan)
Heart
Collagen
Chitin (Chitosan)
Adipose
Collagen
Chitin (Chitosan)
Nerve
Collagen
Chitin (Chitosan)
Metabolic Organs
Collagen
Chitin (Chitosan)

87
Figure 3-15 Other Biomedical Applications. In the diagram some of the collagen applications
are highlighted in yellow (Prashanth & Tharanathan, 2007).
Chitin/Chitosan
Collagen
Biorenewable
Biocompatible
Biodegradable
Biofunctional

88
the clinical applications of this natural biomaterial. (van Blitterswijk, 2007), (Ma & Elisseeff,
2006), (Atala, Lanza, Thomson, & Nerem, 2008).
Characterization of Collagen and Chitin
Materials and Methods
Collagen samples were obtained from Worthington and Sigma. Chitin, from crab
exoskeleton, was obtained from Sigma. Chitin samples were purified into chitin crystallite
suspensions or chitin whiskers. The samples were boiled in 5% KOH solution for 2 hours to
remove residual protein. The samples were washed in deionized water until the pH of the
sample was 7. Chitin suspensions were prepared by hydrolyzing the purified chitin sample with
hydrolysis, the suspensions were washed with deionized water by centrifugation and decanting
of the supernatant. This process was repeated several times until the suspension spontaneously
transformed into a colloidal state (Li, Revol, & Marchessault, 1996) and the pH of the
suspension was approximately 6.
Elemental Analysis
Elemental analysis to determine the Carbon, Hydrogen, and Nitrogen composition was
performed by Prevalere Life Sciences, LLC, Whitesboro, NY.

89
Zeta Potential Analysis
A Malvern Zetasizer 3000 was used to measure the zeta potential and conductivity of the
collagen and chitin samples. Samples were created varying the pH. Samples tested consisted
98ml of 0.005 M solution of KNO3; 1 ml of either [0.1, 0.25, 0.5, 0.75, 1] M of HNO3 or [0.1,
0.25, 0.5, 0.75, 1 M] of KOH; and 1 ml of 0.001 wt% of chitin whiskers suspension. One sample
consisted of 99 ml of0.005 M solution of KNO3 and 1 ml of 0.001 wt% of chitin whiskers
suspension. Data points were plotted.
Thermal Analysis (TGA)
Thermogravimetric analysis was carried out on dried samples using a Shimadzu TGA-50H
Thermogravimetric Analyzer (Kyoto, Japan). Heating was performed in an alumina pan in an air
flow (20 ml /min) at a rate of 10 °C/min up to 800°C. Percentage of weight loss was calculated
using the formula 𝑊𝑡
𝑊𝑜× 100. The percentage weight loss was plotted as a function of temperature
for both collagen and chitin in Figure 2.
Surface Morphology (SEM)
Collagen and Chitin samples were sputtered with an ultrathin layer of gold and studied with a
Hitachi SU-70 Schottky field emission gun scanning electron microscope (The Nanoscale
Imaging, Spectroscopy, and Properties (NISP) Laboratory the Kim Engineering Building at the
University of Maryland at College Park).

90
Results
Chemical analysis
Collagen I and α-chtin both consist of relatively few constituent elements: H, C, O, and N
(Meyers, Chen, Lin, & Seki, 2008) . Results of the elemental composition measurements are
shown in Table 3-7. The elemental analysis revealed the nitrogen content of collagen derived
from bovine achilles tendon was 7.03% while the %N of chitin derived from a crab carapace was
14.61. The theoretical values for % N for collagen and chitin were calculated to be 15.33 % and
6.90 %, respectively. According to Muzzarelli, elemental analysis of chitin isolates typically
have nitrogen content close to 7% and N/C ratio 0.146 for fully acetylated chitin (Muzzarelli,
1999). Degree of acetylation (DA) corresponds to the mole fraction of acetyl units within the
polymer chains. The carbon/nitrogen mole ratio varies from 5.145 in completely N-deacetylated
chitosan to 6.861 in the fully N-acetylated chitin. The degree of deacetylation, DDA, was
therefore calculated according to %𝐷𝐷𝐴 =6.861−𝐶/𝑁
6.861−5.145 (Wanchoo, Thakur, & Sweta, 2008)
(Abdou, Nagy, & Ellsabee, 2008)
However, an explanation for increased nitrogen content lies in the source of the chitin [crab].
Studies indicate that nitrogen content can vary based on the diet, season, or moulting stage; in
females, the production of egg cells also creates a nitrogen difference. Nitrogen content appears
to be the highest for intermoult stages and healthy diets (Linton & Greenaway, 2000), (Linton &
Greenaway, 1997). The experimental values of N% are near the theoretical (6.89) calculated for

91
a chitin that is completely deacetylated. The nitrogen content is also a measure of the protein
amount still present in the chitin. A low experimental value of N is indicative of the minimum
amount of protein left. The sample composition and purity of polysaccharides are in the range of
commercial chitins (Cardenas, Cabrera, Taboada, & Miranda, 2004). The quantity and
characteristics of these constituents such as nitrogen can vary between species and between
individuals of the same species as functions of growth stage, gender, feeding, and other
environmental conditions (Diaz-Rojas, et al., 2006). After purification, the nitrogen, carbon and
hydrogen contents as well as N/C ratio in purified chitin significantly increased. After conversion
of the nitrogen content to the chitin content by a theoretical factor of 203/14, which is the
molecular weight ratio of a monomer (C8H13O5N = 203) to nitrogen (N = 14) in the chitin
(general formula [C8H13O5N]n), the chitin contents in crude and purified crab chitin were 77.1
and 90.0%, respectively. It seems that acid and alkaline treatments followed by decolorization
with potassium permanganate were an effective method in improving the purity of purified
chitin. However, purified chitin contained some components other than chitin (Yen, Yang, &
Mau, 2009).
Collagen degrades. One common archeological indictor for extent of collagen degradation is
the atomic carbon-to-nitrogen ratio, C/N. Modern vertebrate collagen has C/N of 2.8. When
C/N of archeological collagen is greater than 3.1, the isotopic values generally fall outside the
observed ranges for modern animals, meaning that the collagen has degraded sufficiently, that it
is no longer an accurate reflection of diet. These changes in C/N must be associated with
changes in molecular structure. The nature of collagen degradation products and pathways is

92
still largely unknown. Infrared spectroscopy and amino acid analysis have shown loss of amide
bonds and of net amino acid content in degraded collagen, but they do not reveal new organic
constituents generated by the degradation process. Collagen undergoes denaturation,
deamination of R group nitrogen, peptide bond hydrolysis, and condensation reactions of the
hydrolyzed amino acid. The hydrolysis, deamination, and condensation reactions appear to be
components of a concerted process driven by bacteria which leave biomarkers in degraded
collagen. Samples with identical C/N may not have undergone exactly the same degradation
process (Hinman, Markarewicz, asara, Cody, & Tuross, 2008).
Theoretically, nitrogen content of collagen with glycine, proline, and hydroxyproline can be
derived from the empirical chemical formula (C12H24N3O4=274). Therefore, the molecular
weight ratio of the collagen molecule, (C12H24N3O4=274), to nitrogen (N=42) is 274/42 or 6.5.
Nitrogen values were reported on a water- and ash-free basis for Achilles Tendon-Bovine
origin; trimmed from extraneous fat and muscle tissue, hashed, and dried in vacuum; total
nitrogen, 16.9 percent (Graham, Waitkoff, & Hier, 1948).
For Collagen, theoretically, Carbon (144/274), Hydrogen (24/274) and Nitrogen (42/274)
should represent approximately 85% of the sample. However the collagen sample is total
54.27% CHN. Degradation, Contamination, Other than oxygen what other elements are present.
For Chitin, theoretically, Carbon (96/203), Hydrogen (13/203) and Nitrogen (14/203) should
represent approximately 60.59% of the sample. However the collagen sample is total 62.35%
CHN. Degradation, Contamination, Other than oxygen what other elements are present.

93
Table 3-7 Elemental Composition of Collagen. Sample
Collagena
% C
52.55
%H
8.76
%N
15.33
%CHN
76.64
C/N
3.43
Collagenb 41.48 5.76 7.03 54.27 5.90
Chitinc 47.29 6.40 6.90 60.59 6.86
Chitind 41.22 6.13 14.61 61.96 2.82
aCollagen, Theoretical bCollagen derived from Bovine Achilles Tendon, Worthington c Chtin, Theoretical dChitin derived from Crab Carapace, Sigma
Thermal Analysis (TGA)
In the thermogram of chitin (figure 3-16), two thermal events could be observed. The first
occurs in the range of 50–100 °C, and is attributed to water loss. The second occurs in the range
of 300–400 °C and could be attributed to the degradation of the polysaccharide structure of the
molecule, including the dehydration of polysaccharide rings and the polymerization and
decomposition of the acetylated and deacetylated units of chitin. Based on the work of
Jayakumar and Tamura, this is the thermal behavior expected of chitin (Jayakumar & Tamura,
Synthesis, characterization and thermal properties of chitin-g-poly(e-caprolactone) copolymers
by using chitin gel, 2008) (Tanodekaewa, et al., 2004).
In the thermogram of collagen (figure 3-16), three thermal events could be observed. The
first occurs in the range 50-100 °C, and is attributed to water loss. The second event occurs in
the range of 250-350 °C and could be attributed to the breakdown of the triple helix molecule in
chitin. The third event occurs in the range of 450- 600 °C and could be attributed to the further

94
denaturation of collagen to gelatin. Similar behavior was observed in frog skin, amniotic
membrane, and calfskin collagen (Shanmugasundaram, Ravikumar, & Babu, 2004).
Both the collagen and chitin molecules are crystalline which is indicated by thermal shrinkage
behavior due to reorganization, chain folding, recrystallization and other structural changes.
However, chitin has two thermal transitions while collagen has three. Both collagen and chitin
have a thermal event associated with the evolution of freely bound water. Chitin is linear
molecule; hence it does not require the extra thermal step that collagen molecules require to
break down its triple helix. In collagen, the transition from an extended, triple-helical structure to
a random coil is thermodynamically first-order transition, characteristic of melting in crystalline
materials. The temperature at which this transition takes place varies in collagens from different
animals. The denaturation temperature has been correlated with the amount of proline and
hydroxyproline and found to higher in collagens with a higher proportion of these amino acids
(Wainwright, 1982).
Collagen denaturation may also be extremely slow because complete unfolding of the helix
may occur in many steps and via different pathways. Type I collagen is thermodynamically
rather than kinetically stable. Apparently, helices confined in fibers cannot melt completely
because their confinement would not allow chains to gain as much entropy as in solution
(Leikina, Mertts, Kuznetsova, & Leikin, 2002).
Products from the proteins consisted of aromatic hydrocarbons, their nitrogen-containing
analogs, nitriles, aniline, and phenols. Like most organic materials subjected to high
temperatures, the two proteins examined produced aromatic hydrocarbons. The study described

95
Figure 3-16 Thermogram of Collagen and Chitin.
confirms the expectation that most, if not all, organic material will produce polynuclear aromatic
hydrocarbons on exposure to high temperatures. The formation of simple pyridine bases, nitriles,
aniline, and aromatic hydrocarbons as a result of protein pyrolysis is again difficult to rationalize
mechanistically, and studies of pyrolysis of individual amino acids may be helpful. Many of the
products in the protein pyrolyzates bear structural relationships to amino acid units comprising
the original proteins (Higman, Schmeltz, & Schlotzhauer, 1970).
In previous studies, difference in collagen denaturation temperatures have been correlated
with proline (Pro) and hydroxyproline (Hyp) content which are believed to play a substantial role
in the stabilization of the triple helix due to the non-covalent bonding of their pyrrolidine ring.
Therefore, the higher value of Pro + Hyp contributed to a higher thermal stability. Purified
0
10
20
30
40
50
60
70
80
90
100
0 100 200 300 400 500 600 700 800
Weig
ht
Lo
ss (
%)
Temperature (°C)
Collagen
Chitin

96
collagen usually contains traces of non-collagenous proteins and glycosaminoglycan.
Glycosaminoglycans are negatively charged heteropolysaccharides that serve a function in the
formation of the matrix to hold together the collagen fiber of connective tissue. Therefore, the PS
collagen is not only associated with a high degree of glycosaminoglycan but this also affects
other physico-chemical properties. The amino acid composition directly influenced the thermal
stability of different collagen species (Lin & Liu, 2006).
There were over 4500 data points created for each TG curve. Errors can be introduced in
TGA measurements if there are factors such as external vibrations, excessive external heat, slow
recorder/readout equipment, or the sample is packed tightly.
Results from the programmed operation of a thermo balance are represented by a plot of
weight changes versus temperature of time. The plot is referred to as a TG curve. The horizontal
portion indicates the regions where there is no weight change. From this, thermal stability can be
gauged. Instrumental factors that affect a TG curve include furnace heating rate, recording or
chart speed, furnace atmosphere, geometry of sample holder/location of sensors, sensitivity of
recording mechanism, and composition of sample container. Sample factors that affect a TG
curve include amount of sample, solubility of evolved gases in sample, particle size, heat of
reaction, sample packing, thermal conductivity, and sample properties. The strengths of the TGA
include the fact that any type of solid can be analyzed with minimal sample preparation, a
minimum amount of sample is required, and analysis can be qualitative or quantitative. Some
limitations include that the sample must be initially be a solid and data interpretation may not be
straightforward requiring information from other techniques.

97
Zeta potential
Particles interact according to the magnitude of the zeta potential not their surface charge.
Zeta potential values can be used to predict dispersion stability; gives insight into the range of
pH that the substance can operate over without worrying about issues of aggregation and
fluctuations; and gives insight in long term stability. The higher the value of zeta potential, the
more stable the particle dispersion is likely to be. Zeta potential values of 30mV should
remain stable (Kaszuba, 2008).
Figure 3-17 provides the zeta potential of chitin whiskers as a function of pH. The chitin
whiskers are likely to be stable below ~pH 5. From the study of Li, Revol and Marchessault , the
zeta potential of chitin crystallite suspensions at pH 4 in 10-3 M NaCl is 47 mV with a specific
conductance of 0.180. This is of the same order as at pH 2.8 with a zeta potential value of 44
with a specific conductance of 0.699 (Li, Revol, & Marchessault, 1996). These values show that
chitin whiskers are stable below ~pH 5. The isoelectric point of chitin is ~pH 8. According to
the work of Huang and Chen, the isoelectric point of chitosan is pH 8.7. Chitosan and chitin
whiskers are similar derivatives of chitin.
Figure 3-17 also provides the zeta potential of collagen as a function of pH. The isoelectric
point of collagen is ~pH 9-10 (Gobeaux, et al., 2008), (Andrade, Ferreira, & Domingues, 2004),
which is very similar to the isoelectric point of chitin whiskers.

98
Figure 3-17 Zeta potential (ξ) of collagen. Collagen trend line has been taken from (Andrade,
Ferreira, & Domingues, 2004).
As more alkali is added to the suspension the particles tend to become more negatively
charged. As acid is added to the suspension, a point is reached where the charge will be
neutralized. As more acid is added, a buildup of positive charge occurs. There is a point where
the curve passes through zero. The point where the curve passes through zero is called the
isoelectric point and it is where the system is least stable. It can be seen in the chitin curve (blue)
when the dispersion pH below 4 or above 9, there is sufficient charge to confer stability. If the
system is between 4 and 9, the dispersion may be unstable. This instability is most likely around
pHIEP 8 (the isoelectric point). If the system is between 7 and 10, the dispersion may be unstable.
-20
-10
0
10
20
30
40
0 1 2 3 4 5 6 7 8 9 10 11 12 13
Zeta
Po
ten
tial
(m
V)
pH
Chitin
Collagen

99
The trendline for both collagen and chitin indicate that basicity does not improve stability much.
It should be noted that collagen has a greater affinity to be less stable in the basic pH region. This
is probable due to the fact the collagen is more basic molecule with hydroproline, more hydrogen
and amine groups.
The error bar indicates the standard deviation for 3 runs of Zeta Potential measurements
It does not change my evaluation of the information the standard deviation adds to the
information about how the degree in which zeta values within the sample differ from the sample
mean. The collagen trendline was taken from Andrade et al. no standard deviation or error bars
were reported. Many in the literature give a couple values of zeta potentials in regard to pH
instead of a complete trend line. One of those points is typically the isoelectric point. Common
limitations to generating high quality zeta potential measurements are very small particles, very
dilute samples, concentrations that are too high, dissolved solids, or residual reactants.
Scanning Electron Microscopy (SEM)
Chitin SEM micrograph shows chitin fibers [see figure 3-18]. Chitin and collagen [figure 3-
19] SEM micrographs both display lattice-like structures with nucleation sites. The fibers in both
collagen and chitin micrographs were oriented parallel, perpendicular, or a variation in between,
to the surface, on successive planes. Multidirectional or ‘twisting plywood’ system (Neville A.
C., 1993) was observed. Bouligand and Neville observed similar helicoid systems while
examining crustaceans (Bouligand Y. , 1972). One major difference, between the chitin and

100
collagen fibers, is that the collagen fibers of SEM micrographs demonstrate the characteristic 67
nm banding pattern of a collagen fiber (Franchi, et al., 2008).
Advantages of the SEM include its wide-array of applications, detailed topographical
imaging, and the fast nature of the instrument. One disadvantages is the fact the preparation of
samples can result in artifacts. SEMs are limited to solid samples small enough to fit inside the
vacuum chamber and able to handle vacuum pressure. Special training is required to operate an
SEM as well as prepare samples.
Figure 3-18 Chitin from Crab Exoskeleton SEM Micrographs.

101
Figure 3-19 Collagen from Bovine Achilles Tendon SEM Micrographs
Conclusion
Collagen and chitin follow the same extraordinary storyline. They do not act alone; collagen
is a triple helix embedded in an inorganic matrix and chitin is embedded in a protein helix.
Compared to typical structures of their kind, both collagen and chitin are unusual. Some other
significant points about collagen and chitin include:
Chitin is a polysaccharide; Collagen is a protein;
Collagen and Chitin associate with proteoglycans;
Collagen and chitin complex composites are hierarchically structured and multifunctional
Microfibril: Fribil Aggregation (Fiber): Fibrous Bundle: ‘Twisted Plywood”;

102
Chitin and collagen are lyotropic liquid crystals that form LC phases in solution. Liquid
crystals and fibrous composites such as collagen and chitin are both mobile, have similar
ordered geometries, can self-assemble, and have similar optical properties;
The biosynthesis and degradation of both collagen and chitin are stepwise procedures
governed by enzymatic activity; these pathways consist of catalytic units, precursors,
biotransformations, polymerization, hydrolysis, and fibrillogenesis;
Both collagen and chitin are synthesized by sequential changes in the cytoplasmic matrix,
RER, processing in the Golgi apparatus and transport through vesicles culminating in
exocytosis across the membrane where chemical reactions occur and association into
fibril, fibers, and fibrous bundle;
Collagen and chitin share properties; they are both biocompatible, biodegradable, non-
toxic, highly insoluble, and birefringement;
Chitin and collagen have a wide variety of medical applications.
An argument can be made that collagen is what holds bone together. The same argument can
be made for chitin and crab exoskeleton. Since collagen and chitin share so many similarities
could they be used interchangeably as implants? Previous reports indicate that chitin induced
fine collagen fibers histologically. Chitin was found to enhance synthesis of types I, III, and IV
collagens (Kojima, et al., 2004), (Minami, et al., 1996). If chitin can, does this ability extend to
its source which in this case includes crab shell integument?

103
Chapter 4 Tools of Biomineralization: Calcium Carbonate and Calcium
Phosphate
Introduction
A vast array of organisms are known to precipitate minerals (Lowenstam H. A., 1981). The
formation of inorganic minerals by biological systems, e.g. in bone, teeth, and mollusk shells, is
called biomineralization (Neues, Ziegler, & Epple, 2007). Until the early 1980s the field was
known as “calcification,” reflecting the predominance of biologically formed calcium-containing
minerals. As more and more biogenic minerals were discovered that contained other cations, the
field became known as “biomineralization” (Weiner & Dove, 2003), and is characterized by a
remarkable level of molecular control of the particle size, structure, morphology, aggregation,
and crystallographic orientation of the mineral phases. Biomineralization results in complex
materials (Mann, et al., 1993). One of the major challenges in the field of biomineralization is to
understand the mechanism(s) by which biological systems determine which polymorph will
precipitate (Weiner & Dove, 2003). When investigating any mineralized tissue, one must be
cognizant of the complex nature of the material: 1) the chemical nature and crystalline form of
the mineral; 2) the nature and form of the organic matrix; 3) the relationship between the organic
and inorganic components of the tissue and the influence of the matrix on crystal morphology; 4)
the sources of mineral for deposition; 5) the pathways for mineral movement into or out from the
mineralized structures; 6) rates of mineral deposition; and 7) the nature and location of

104
nucleation sites within the matrix and mechanisms for control or cessation of crystal growth
(Roer & Dillaman, 1984).
Biomineralization provides an intelligent strategy to generate functional materials such as
mineral skeletons and cuticles (Tao, Zhou, Zhang, Xu, & Tang, 2009) that exhibit tight structural
control at the nanoscale (Barthelat, 2007). Skeletons and cuticles are formed in a wide variety of
shapes, sizes, and compositions of organic and mineral components (Omelon, et al., 2009).
Mineralization takes time depending on the animal species (Skinner & Jahren, 2003). In nature,
crustacean animals periodically undergo cyclic molt of the exoskeleton, which is essential for
their growth, metamorphosis, and reproduction. The layers in cuticle are deposited and
mineralized at distinctly different stages: premolt, intra-molt, postmolt, and inter-molt (Tao,
Zhou, Zhang, Xu, & Tang, 2009). Calcium carbonate is used by many invertebrate organisms to
mineralize a large variety of structures. The delicately structured exoskeleton or cuticle protects
arthropods from environmental hazards like desiccation, osmotic and mechanical stress, and
predation. In addition, it serves as support for organs, provides sites for muscle attachment and
takes up mechanical tension during muscle contraction. These functions require an adequate
mechanical strength of the cuticle (Neues, Ziegler, & Epple, 2007).
Vertebrate skeletons are instead composed of a calcium phosphate mineral known as apatite.
The vertebrate skeleton must satisfy a wide range of demands, including structural integrity,
metabolic activity, growth, and continual repair of wear and damage caused by locomotion
and/or trauma. These demands require that vertebrates continually resorb and rebuild their
mineralized skeleton. The high functionality and metabolic activity of the vertebrate skeleton

105
suggest that apatite offers an advantage to the vertebrate organism that other minerals do not
(Omelon, et al., 2009).
To date, approximately 60 biogenic minerals (i.e. generated through biological processes)
have been identified (Dunlop & Fratzl, 2010), see table 4-1 for examples of biomineral diversity.
The calcium carbonate minerals are the most abundant biogenic minerals, both in terms of the
quantities produced and their widespread distribution among many different taxa. Phosphates
comprise about 25% of the biogenic mineral types. The most abundantly produced phosphate
mineral is carbonated hydroxyapatite (Weiner & Dove, 2003).
The vertebrate skeleton or bone is extremely enterprising and continually being rebuilt,
repaired, and resorbed during growth and normal remodeling (Omelon, et al., 2009). In
Crustacea, molting includes a cycle of demineralization of the old and mineralization of the new
cuticle. Crustaceans and bones are excellent models for the study of dynamic biomineralization
processes (Neues, Ziegler, & Epple, 2007). The detailed processes of biomineralization are as
varied as the organisms themselves (Weiner & Dove, 2003).
Definition
Landis and Glimcher defined biomineralization as the process by which living forms
influence the precipitation of mineral materials. The process creates heterogeneous
accumulations, composites composed of biologic (or organic) and inorganic compounds, with

106
Table 4-1 Examples of the Diversity of Biominerals (Gower, 2008).
Biogenic
minerals
Formula Organism Biological
Location
Biological
Function
Calcium
carbonates
(calcite, vaterite,
aragonite,
amorphous)
CaCO3,
(Mg,Ca)CO3,
CaCOnH2O
Many marine
organisms,
aves, plants,
mammals
Shell, test,
eye lens,
crab cuticle,
eggshells,
leaves, inner
ear
Exoskeleton,
optical,
mechanical
strength,
protection,
gravity receptor,
buoyancy
device, Ca store
Calcium
phosphates
(hydroxylapatite,
dahllite)
Ca10(PO4)6(OH)2,
Ca5(PO4CO3)3(OH),
Ca8H2(PO4)6
Vertebrates,
mammals,
fish, bivalves
Bone, teeth Endoskeleton,
ion store,
cutting/grinding,
protection,
precursor
inhomogeneous distributions that reflect the environment in which they form (Landis &
Glimcher, 1978).
According to a modern definition, minerals are elements or compounds, amorphous or
crystalline, formed through biogeochemical processes. This definition acknowledges that some
biominerals are barely crystalline but avoids a discussion of amorphous. Biominerals have
distinct morphologies and make unique contributions to well-known life forms. Many
biomineralized tissues are composite materials, containing a biologically produced organic
matrix and nano- or microscale amorphous or crystalline minerals. During the processes of
biomineralization the organic material acts in various roles as nucleator, cooperative modifier,

107
and matrix or mold for the mineral ions. The resulting tissue has properties very different from
those of the pure minerals themselves. The stiff mineral prevents the organic matrix (proteins,
peptides, polysaccharides, lipids) from yielding, while the organic matrix hinders crack
propagation (Ehrlich, Koutsoukos, Demadis, & Pokrovsky, 2008).
Mineral Function/Strength/ Properties
With unique combinations of stiffness and strength, hard tissues can serve a variety of
functions: mechanical support (bones) or armored protection (cuticles). In nature, the most
common route taken to make stiff materials out of soft protein networks and tissues is to
incorporate minerals, which are much stiffer (Barthelat, 2007). Therefore, most natural materials
are composites based on biopolymers and some minerals (Dunlop & Fratzl, 2010). The effect of
combining soft proteins with stiff minerals can be seen in figure 4-1, which is a material
properties map for a selection of natural ceramics, biopolymers and their composites. Their
mineral content makes them 100–5000 times stiffer than soft proteins (Barthelat, 2007). Despite
the relative scarcity of these constituents, their combination yields materials with a great
variation in functionality. A particular characteristic of biological composites is their
multifunctionality (Dunlop & Fratzl, 2010).
Many mineralized tissues fulfill structural functions. The vast majority of these, such as
bones, teeth, and shells, utilize crystalline minerals to stiffen and strengthen the tissue (Addadi,
Sefi, & Weiner, 2003). In one example, organic fiber networks are reinforced by mineral, as in
the case of the crab cuticle, which consists of a plywood-like arrangement of chitin fibers and

108
Figure 4-1 The materials property chart (Dunlop & Fratzl, 2010).

109
inclusions of calcite and amorphous calcium carbonate. Another example is bone, in which
collagen fibrils are reinforced by hydroxyapatite platelets of only a few nanometer thickness
(Bouligand, 1972), (Dunlop & Fratzl, 2010). The calcium phosphate mineral phase of bone has
two functions: (1) providing structural stability to the skeleton by infusing the organic matrix of
bone to form a rigid structural material; and (2) operating as a storage site for calcium, inorganic
orthophosphate, sodium, magnesium, carbonate, and other ions. Being involved in both the
biomechanical and metabolic functions of osseous tissue, bone mineral acts as a reservoir
providing the body fluids with these ions to maintain the biologically required levels and as a
detoxifying depository to store ions unwanted in the body fluids (Posner, 1985). The relationship
between toughness and modulus for natural materials is shown in figure 4-1. Materials vary
widely for both properties. The materials property chart illustrates many mechanically important
materials where both stiffness and toughness are large (Dunlop & Fratzl, 2010).
The process of mineral phase formation is regulated by solubility, surface chemistry, and
bioavailability of elemental and molecular species such as calcium, carbonate, and phosphate
(Wilson Jr., 2008). The mineral phase is thought to dominate the stiffness of bone, which
increases more than linearly with mineral content. Mineral platelets deform elastically to almost
twice the bulk failure strain. Load-bearing organs or protective elements require a high modulus
to transmit loads (bone) or to minimize deformation (cuticle). High stiffness is readily achieved
by the incorporation of mineral phases (e.g., calcite and hydroxyapatite) and of stiff, fibrous
elements (collagen or chitin) and through extensive cross-linking. (Dunlop & Fratzl, 2010). The

110
main mechanical feature of hard biological tissues is their ‘hardness’, although ‘stiffness’
(resistance to deformation) may generally be a more accurate term (Barthelat, 2007).
Having formed under controlled conditions, biomineral phases often have properties such as
shape, size, crystallinity, isotopic and trace element compositions quite unlike its inorganically
formed counterpart. The term “biomineral” reflects all this complexity (Weiner & Dove, 2003).
Biology is limited to using a small number of biopolymers, some minerals and ions, and various
cross-linking agents to make its materials. Despite these limitations, natural materials present an
incredible variability in their properties. Mechanical properties of biological composites based
on polymers and mineral, are usually very superior to those of the components (Dunlop & Fratzl,
2010). Bone mineral is an integral part of bone tissue having physical and chemical properties
Figure 4-2 Biomineral crystallographic properties are highly regulated during
biomineralization. These properties provide some indication of potential processing strategies
(Gower, 2008).

111
that directly affect those of the tissue as a whole (Termine J. D., 1972). Figure 4-2 depicts some
properties that are regulated during biomineralization.
Biomimetics and biomineralization
The variation in biological mineralization is enormous. These variations represent alternative
solutions to similar problems. The adaptation of biomineralization processes in the laboratory
highlights the potential of a biomimetic approach to crystal science. In particular, the concept of
molecular recognition at inorganic-organic interfaces involved in nucleation and crystal growth
is providing a perspective in which the classical approaches to inorganic crystallization can be
developed within a structural and sterochemical context. Moreover, the integration of organic
supramolecular chemistry, self-assembly, and inorganic materials chemistry provides the
opportunity to develop rational synthetic routes to products with uniform particle sizes,
nanoscale dimensions, tailored morphologies, crystallographic orientation, and organized
microarchitectures (Mann, et al., 1993).
Mann coined the term “molecular tectonics” and defined it as a process that strives towards
the chemical construction of higher-order architectures, albeit on different length scales.
Furthermore, he proposed that the study of the molecular processes that result in the construction
of higher-order architectures in biomineralization is relevant to the development of new
strategies for the controlled synthesis of organized inorganic and composite materials. He
presented a commentary, using representative examples, on the essential aspects of molecular

112
tectonics that connect materials chemistry and biomineralization, and ultimately their
assimilation within a biomimetic approach (Mann, 1993).
The three constructional processes of biomineralization- supramolecular pre-organization,
interfacial molecular recognition (templating) and cellular processing – represent a sequence of
increasing tectonic complexity. This leads to three main divisions of biomimetic materials
chemistry inspired by studies of biomineralization, which are highlighted in figure 4-3. First, the
specific chemical and structural properties of biomolecules involved in biomineralization
processes can be utilized in synthetic reactions. A second area of biomimetic research is in the
use of living organisms, as intact biosystems, for the fabrication of highly ordered composites. A
third biomimetic approach involves the use of biological concepts as inspiration for new ideas in
the synthesis of inorganic materials with controlled properties, such as uniform particle size,
polymorph selectivity, tailored morphology, oriented nucleation and organized assembly (Mann,
1995).
These materials processing rates are achieved through contrasting assembly strategies that are
instructive to the materials scientist. Understanding the inherent complexity of the molecular
systems controlling the biological synthesis is a major challenge to materials scientists, who want
to copy the structure, property, and performance (function) relations of these elegant structures
(Heuer, et al., 1992).

113
Figure 4-3 There exist three primary divisions of biomimetic materials chemistry inspired by
biomineralization studies (Mann, 1995).
Biomineralization Models
Several theories have been proposed for the mechanism of mineralization of hard tissue such
as bone and cuticle (Lowenstam H. A., 1981), (Mann, 1983), (Anderson, 1984), (Nilsen, 1980).
BIOMIMETIC
MATERIALS CHEMISTRY
Biomolecules
Bioconcepts
Biosystems
proteins
polysaccharides
bone
cuticle
molecular recognition
vectorial regulation
cellular processing
preorganization

114
The classical model of biomineralization considers mineral formation as an amplification process
in which individual atoms or molecules add to existing nuclei or templates, while living
organisms may make use of proteins and peptides to deterministically modify nucleation, growth
and facet stability. This conventional concept for crystal growth has been challenged (Cai &
Tang, 2008). Thus, there is at present no unified theory of the mechanism of mineralization,
especially for bone (Posner, 1985). A discussion of various models is presented below.
Biologically Induced
The basic process of biologically induced mineral formation is characterized by bulk
extracellular and/or intercellular mineral formation, without the elaboration of organic matrices.
"Biologically induced" mineralization results in the minerals having crystal habits similar to
those produced by precipitation from inorganic solutions (Lowenstam H. A., 1981). In this
situation, cell surfaces often act as causative agents for nucleation and subsequent mineral
growth. The biological system has little control over the type and habit of minerals deposited,
although the metabolic processes employed by the organism within its particular redox
environment mediate pH, pCO2 and the compositions of secretion products. These chemical
conditions favor particular mineral types in an indirect way. In some cases, biological surfaces
are important in the induction stage because nucleation often occurs directly on the cell wall, and
the resulting biominerals can remain firmly attached. Heterogeneity is the hallmark of
biologically induced minerals. This heterogeneity includes variable external morphology

115
(typically poorly defined), water content, trace/minor element compositions, structure and
particle size (Lowenstam & Weiner, 1989), (Weiner & Dove, 2003).
Organic matrix-mediated
In “organic matrix-mediated” process, the organism constructs an organic framework or mold
into which the appropriate ions are actively introduced and then induced to crystallize and grow.
The mineral type, orientation of crystallographic axes, and microarchitectures are under genetic
control (Lowenstam H. A., 1981).
“Organic matrix-mediated” was generalized by Mann (Mann, 1983) to “biologically
controlled” mineralization. The organism uses cellular activities to direct the nucleation, growth,
morphology and final location of the mineral that is deposited in “biologically controlled”
mineralization. The degree of control varies across species, and almost all controlled
mineralization processes occur in an isolated environment. The results can be exceptionally
sophisticated, species-specific products that give the organism specialized biological functions.
Biologically controlled mineralization processes can be characterized as occurring extra-, inter-
or intracellularly (Table 4-2). In some cases, mineral formation begins within the cell and then
proceeds outside the cell (Weiner & Dove, 2003).

116
Table 4-2 Characterization of Biologically Controlled Extra-, Inter-, and Intra-cellular
Mineralization
Biologically controlled
extracellular mineralization
Biologically controlled
intercellular mineralization
Biologically controlled
intracellular mineralization The cell produces a
macromolecular matrix
outside the cell in an
area that will become
the site of
mineralization. (Matrix
refers to a group of
macromolecules
comprised of proteins,
polysaccharides or
glycoproteins that
assemble to form a
three-dimensional
framework).
The structures and
compositions of these
organic frameworks are
programmed to perform
essential regulating
and/or organizing
functions that will result
in the formation of
composite biominerals.
Almost all structures
that form by
extracellular processes
develop upon a pre-
formed matrix derived
from secretory products
of multicellular
epithelial tissues.
Examples of organisms
believed to mineralize
primarily by
extracellular
biologically controlled
processes include shells
of mollusks and bones
and teeth.
It typically occurs in
single-celled organisms
that exist as a
community.
The epidermis of the
individual organisms
serves as the primary
means of isolating the
site of mineralization.
The epithelial substrate
reproducibly directs the
nucleation and growth
of specific biomineral
phases over large areas
of cell surfaces.
Mineralization between
cells can become so
extensive as to
completely fill the
intercellular spaces, thus
forming a type of
exoskeleton.
The epidermis of
individual organisms
directs the polymorph
and shape of the
biomineral that forms.
An example is found in
calcareous algae that
nucleate and grow
calcite with a c-axis
orientation that is
perpendicular to the cell
surface.
Controlled
mineralization can also
occur also within
specialized vesicles or
vacuoles that direct the
nucleation of
biominerals within the
cell.
These
compartmentalized
crystallization
environments govern the
resulting biomineral
composition and
morphology.
The cell has a high
degree of control upon
the concentrations of
cation and anion
biomineral constituents
in an environment where
an organic matrix may
also be active as a
nucleating template.
The compartment
membrane also regulates
the pH, pCO2 and minor
element compositions.
Indicative of the general
scenario whereby
biominerals form
intracellularly before
eventually becoming
extracellular.
(Watabe, 1974), (Veis, 2003), (Weiner & Dove, 2003).

117
Biogenic minerals found in teeth and bones are synthesized by precise cell-mediated
mechanisms. These hard tissues have superior mechanical properties due to their complex
architecture. It is believed that the initial mineralization phase is amorphous and during the
nucleation and growth process, the initially formed metastable amorphous calcium phosphate
phase transforms into thermodynamically stable crystalline hydroxyapatite in a precisely
controlled manner. The organic matrix-mediated controlled transformation of amorphous
calcium phosphate into crystalline HAP has been confirmed by x-ray diffraction, selected area
electron diffraction (SAED or SAD), Raman spectroscopy, and elemental analysis (Gajjeraman,
Narayanan, Hao, Qin, & George, 2007).
Biomineralization of collagen involves functional motifs incorporated in extracellular matrix
protein\molecules. These molecules have the objective to stabilize the amorphous calcium
phosphate into nanoprecursors and to direct the nucleation and growth of apatite within collagen
fibrils, figure 4-4. For organic-inorganic nanocomposites such as bone, this self-assembly
process determines the framework and spatial constraints for nucleation and propagation of the
reinforcing mineral phase. By itself, type I collagen is insufficient to induce nucleation of
carbonated apatite from transient amorphous calcium phosphate (ACP) phases (Falini &
Fermani, 2004). The organic matrix exerts a great degree of crystallographic control over the
nucleation and growth of mineral particles. Type I collagen, the predominant matrix protein in
bone, possesses self-assembling properties and forms enclosed spaces within which the inorganic
reinforcing phase grows. When the degree of supersaturation of calcium and phosphate ions is
high, thermodynamically stable HAP is formed via intermediate precursor polymorphs such as

118
amorphous calcium phosphate (ACP) or octacalcium phosphate. It is well established that type I
collagen matrix does not have the capacity to induce matrix-specific mineral formation from
metastable calcium phosphate solutions that do not spontaneously precipitate but merely provide
the organizational framework and spatial constraint for crystal deposition. Noncollagenous
matrix macromolecules might be involved in the control of nucleation and growth of the mineral
phase. Type I collagen designates the space in which the crystal grows, and this constrained
space might be necessary for defining the crystal size and morphology (Gajjeraman, Narayanan,
Hao, Qin, & George, 2007).
Figure 4-4 Schematic of the mineralized collagen fibrils, the basic constituents of bone.
Nanocrystals of HAP are incorporated between collagen molecules (Dorozhkin & Epple, 2002).
Crystal of HA
( 50 x 25 x 4 nm3)
Mineralized collagen fibril:
few µm long and
100 nm in diameter
Collagen molecules (polypeptide chains)
300 nm long and 1.5 nm in diameter

119
In mollusks, more than 40 types of ultrastructural arrangement are known where many
crystals are embedded in an organic matrix and mechanical requirement adaptations are
reflected. Polymorphs of calcium carbonate (aragonite, calcite, and vaterite) have been observed
(Falini & Fermani, 2004). The location and polymorphism of the crystals are governed by the
super-saturation inside the compartmentalized space in the chitin. As the organic component, a
role of chitin is to reinforce the mineral phase. This is analogous to collagen, which is the main
organic component of bone. This property, in addition to the inherent biodegradability,
biocompatibility, and good mechanical properties of chitin, has paved the way for the use of
chitin–HA composites as a hard tissue substitute. These composites are useful as a bone material
substitute because they allow the introduction of osteoconductive HA into regions of bone loss,
with the chitin matrix acting as a binder to prevent postoperative migration of the HA particles
(Falini & Fermani, 2004). Other composites consisting of collagen, chitin, calcium phosphates,
and their derivatives have been used as hard tissue substitutes (Ge, Baguenard, Lim, Wee, &
Khor, 2004), (Wahl & Czernuszka, 2006), (Jayakumar, Prabaharan, Nair, & Tamura, 2010).
Because of their properties, these materials have the potential for use for a variety of medical
applications (Singh & Ray, 2000), (Dutta, Ravikumar, & Dutta, 2002).
Mesocrystals model
The concept of mesocrystals, which consist of crystallographic oriented crystallites connected
by polymers or surfactants, has been suggested in the studies of biomineralization. Under the

120
regulation of various organic molecules, the formation of mesocrystals has been previously
observed in the growth processes of CaCO3. In these mesocrystals, the building blocks are the
fully crystallized inorganic phase. The organic molecules connect the adjacent crystallites,
resulting in the superstructures. A crucial condition in the architecture of mesocrystals is the
interaction between crystallites and organic molecules (Cai & Tang, 2008).
Mesocrystals are colloidal crystals composed of crystalline nanoparticles in perfect three-
dimensional crystallographic register so that they scatter like single crystals but are clearly a
nanoparticle superstructure. Consistent with the model of a mesocrystal, nanoparticles were
suggested as building units of biominerals in combination with a brick-by brick formation
mechanism. Studies point to the fact that nano-clustered crystal growth, induced by organic
matrices, is a basic characteristic of biomineralization that enables the production of composite
materials with elaborate morphologies. These examples indicate that mesocrystal formation
plays a role in biomineralization, at least as an intermediate in the production of single crystals
with complex morphologies. This nanoparticle mediated crystallization pathway involving
mesoscopic transformation is called ‘‘non-classical crystallization’’. Contrasting a single
nucleation event to form a single crystal, this pathway involves multiple nucleation events of
nanoparticles which form a nanoparticle superstructure. Non-classical crystallization engages
self-organization of pre-formed nanoparticles to an ordered superstructure, that can later fuse to a
single crystal (Xu, Ma, & Colfen, 2007).

121
Figure 4-5 Illustrative representation of classical and non-classical crystallization (Xu, Ma, &
Colfen, 2007).
In figure 4-5, Pathway a) represents the classical crystallization pathway in which nucleation
clusters form and grow until they reach the size of the critical crystal nucleus growing to a
primary nanoparticle, which is amplified to a single crystal. Pathway b). The primary
nanoparticles can get covered by a polymer or other additive before they undergo a mesoscale
assembly, so they can form a mesocrystal. Pathway c). Mesocrystals can form from pure

122
nanoparticles. Pathway d). There also exist the possibility that amorphous particles are formed,
capable of transforming before or after their assembly to complicated morphologies (symbolized
by the question mark in path d) (Colfen & Mann, 2003), (Xu, Ma, & Colfen, 2007).
Brick and mortar model
A new model of ‘‘bricks and mortar’’ concerns the biological aggregation of apatite
nanoparticles. An inorganic phase, amorphous calcium phosphate (ACP), acts as ‘‘mortar’’ to
cement the crystallized ‘‘bricks’’ of nano-HAP. Meanwhile, biological molecules control the
nano-construction. By using HAP nanospheres as the building blocks, highly ordered enamellike
and bone-like apatite are hierarchically constructed in the presence of glycine (Gly) and
glutamate (Glu), respectively. During the progression of biological apatite, the amorphous
‘‘mortar’’ can be eventually turned into the ‘‘brick’’ phase by phase transformation to ensure the
integrity of biominerals (Cai & Tang, 2008)
Cellular Role
It is recognized that bone-related cells (especially osteoblasts and osteoclasts) play key roles
in the physiological formation of bone. Bone-related cells not only are theorized to take part in
the formation of biomineral and construction of bone, they also continuously regulate the
density, regeneration and degradation of bone. So understanding the relationship between bone-
related cells and calcium phosphates is a necessary step in order to elucidate the formation

123
Figure 4-6 An illustration of the role of inorganic and organic constituents in controlling
biomineral formation (Gower, 2008).
mechanism of bone (Cai & Tang, 2008). Cells are amply sensitive to their surroundings such that
signaling molecules or macro-, micro-, or nano-scale alterations in topography may elicit diverse
cell behaviors (Stevens & George, 2005).
Cellular processing administers spatial and temporal control over the delivery of the species
and their subsequent organic-inorganic interactions. In the physiological environment, there is a

124
requirement for mineral reactants and inorganic impurities can have a pronounced impact on the
process, especially in combination with the soluble organic additives, where promotion of the
amorphous precursor pathway can occur. The soluble additives are considered as having either
“process-directing” or “structure-based” interactions. Structure-based interactions may influence
the crystallographic texture, phase, and orientation (Gower, 2008). Thus, the role of inorganic
and organic constituents in controlling biomineral formation is illustrated in figure 4-6.
Matrix Vesicles (MV)
Bone apatite formation seemingly proceeds via a number of cooperative mechanisms. Current
hypotheses encompasses the utilization of: (i) Matrix vesicles which bud from the plasma
membrane and accumulate calcium (Ca2+) and phosphate (PO43−) ions extracellularly before
associating with the collagenous ECM (2); (ii) Noncollagenous proteins, associated with the gap
zones in collagen, mediate and foster mineral nucleation and its propagation within and along
collagen fibrils; and (iii) Amorphous calcium phosphate and ionic calcium stored in
mitochondria is transported via vesicles to the ECM before converting to more crystalline
apatite. In the depiction, “matrix vesicles” are purple, and collagen-mediated mineralization is
portrayed in the bottom left corner with calcium and phosphate ions highlighted in yellow and
red. Mitochondria are shown in green, and vesicles are orange and blue, with and without
mineral/ions, respectively. “N” identifies the cell nucleus (Boonrungsiman, et al., 2012). Figure
4-7 outlines this proposed mechanism for bone mineral formation.

125
Figure 4-7 Diagram outlining a proposed mechanism for bone mineral formation
(Boonrungsiman, et al., 2012).
Anderson (1967) and Bunucci (1967) demonstrated membrane-bounded vesicles in the matrix
of hard tissues having a diameter of about 100 nm. They proposed that the initial crystals

126
appeared inside these matrix vesicles (MV). Later two subtypes of these matrix vesicles have
been described, type I having a dark, lysosome-like content and type II having a content
resembling intracellular matrix. Several investigators have proposed that matrix vesicles
constitute the beginning stages of mineralization in bone (Nilsen, 1980).
Matrix vesicles are ubiquitous in the cytoplasm of eukaryotic cells and are diverse in the
human body. These vesicles are involved in a wide range of transport, storage and secretory
functions and are assembled with well-defined compositions. They may undergo a variety of
specific transactions involving fusion events, notably during protein trafficking between cisterna
and the Golgi stack. Matrix vesicles generally adopt a spherical morphology in the absence of
external scaffolds. They are extracellular, membrane-invested, and 100 to 200 nm in diameter.
Matrix vesicles are enriched in Ca and lipids and possess a unique assortment of
phospholipids characterized by an acidic phospholipid with a strong affinity for Ca2+. These
particles which have been described as the initial locus of calcium phosphate mineral deposition
in cartilage, growing bone and predentin of teeth. In addition, matrix vesicles are the initial locus
of calcification in all growing skeletal tissues in which studies of anhydrously prepared tissues
support this assertion.
Matrix vesicles have membranes that degrade during calcification by both enzymatic and
mechanical forces. Degradation explains the low matrix vesicle numbers occurring in calcified
zone of growth plate. These membranes that are replete with phosphatase activities including

127
alkaline phosphatase, ATPase, inorganic pyrophosphatase, and 5’-nucleotidase. The outer
membranes provides one of the few protected and controlled internal microenvironments that
exist outside cellular environments in which complex interactions between molecules can occur.
Matrix vesicles are programmed based on increasing evidence, whereby metabolic objectives
of the host cell may be pursued vigorously at a distance from the host cell and can be transferred
into other cells. Vesicle formation remains an area of some disagreement and varied opinion.
Their pattern of phospholipid incorporation strongly suggests that they take their origin from
adjacent cells. Thus, major theories of biogenesis include:
a. budding from plasma membrane ( much supported )
b. cell degeneration
c. extrusion of intracytoplasmic vesicles
d. extracellular subunit self- assembly
(Anderson, 1984), (Mann, 1993), (Anderson, Mulhall, & Garimella, 2010).
Calcium phosphate mineral formation in vertebrate bone formation is a multistep process.
Much evidence indicates that extracellular MV play a key role in the induction of the first solid
phase mineral. Previous biochemical and ultrastructural studies have shown that MV contain
large amounts of calcium and phosphate. However, the initial mineral phase has not yet been
characterized (Wu, et al., 1997). With regard to biomineralization, the enclosed intravesicular
space provides a means of controlling the size and location of discrete intracellular mineral
products, whereas ion pumps in the vesicle membrane are responsible for controlling the
physicochemical conditions (supersaturation, ionic strength, complexation, pH) pertaining to

128
crystal nucleation and growth (Mann, Molecular tectonics in biomineralization and biomimetic
materials chemistry, 1993).
Amorphous Mineral Component
Hydroxyapatite formation in aqueous environments has been intensively investigated for
studies both of biomineralization and synthesis of biomedical materials. A multistep strategy of
phase transformation has been suggested and highlights the enlistment of transient amorphous
precursors in biomineralization. Evidence of this, for example, are the first-formed amorphous
calcium carbonate (ACC) in sea urchin spicules, corals, and crustacean cuticles, etc. The
participation of amorphous calcium phosphate (ACP) in the early stages during bone generation
and the subsequent transformation into carbonated hydroxyapatite (HAP) have been clarified in
vivo. The precursor role of ACC and ACP in calcium mineralization has also been confirmed in
vitro by a number of laboratory studies. It is believed that these amorphous phases are the
reservoirs for crystallization and the mechanical strengtheners during biomineral formation (Xu,
Ma, & Colfen, 2007), (Tao, Zhou, Zhang, Xu, & Tang, 2009).
The macromolecules introduced inside the amorphous phase from the start, or at an
intermediate stage, may fulfill two distinct functions. Some macromolecules may contribute to
the transient stabilization of the amorphous phase that must be broken down or otherwise
neutralized in an accurately orchestrated series of events, to allow for crystallization. Other
macromolecules have the ability to modulate crystal growth and are presumably adsorbed at the

129
boundary planes as the crystal grows, influencing crystal growth in certain directions rather than
others. It is easy to overlook the presence of ACC or ACP especially when it co-exists with the
crystalline forms of calcium carbonate (Addadi, Sefi, & Weiner, 2003) or calcium phosphate.
Two modes have been postulated for the transition from an amorphous to a crystalline structure:
direct transformation and heterogeneous growth-transformation. Direct transformation proposes
the reorganization of the internal structure of amorphous particles into ordered crystalline states,
whereas heterogeneous growth transformation predicts that crystals grow incrementally on an
amorphous groundwork, eventually overgrowing the preexisting amorphous particles (Tsuji,
Onuma, Yamamoto, Iijima, & Shiba, 2008).
Control over biomineral properties can be accomplished by regulation of particle size, shape,
crystal orientation, and polymorphic structure. Biogenic minerals are assembled using a transient
amorphous mineral phase, and the stabilization of amorphous precursors during the growth phase
is important in biomineralization because transient amorphous minerals play an important role in
many organisms. Amorphous materials can be molded into various shapes. Moreover, a growing
body of evidence has shown that the shape of the biominerals is often controlled through
molding of solid or gelated amorphous precursors. Transition of amorphous phase to crystalline
phase is carried out in a controlled manner by the organism (Gajjeraman, Narayanan, Hao, Qin,
& George, 2007).

130
Mg stabilized amorphous precursors
Although the concentration varies considerably among species, magnesium and phosphate are
found in virtually all crustacean cuticles. Still unknown, the function of calcium phosphate
within the crustacean cuticle might enhance the ACC formation because in vitro experiments
showed that under physiological conditions, low concentrations of phosphate prevent calcite
crystal nucleation and formation. However, it is also possible that ACP is merely formed to alter
the mechanical properties of the cuticle. Differences within the composition of the cuticle
between species convey a chemical adaptation to their different biological requirements. Within
the cuticle, the mineral composition relies on the function and the habitat of the animal. In
combination with a high concentration of organic material and rather low amounts of calcite,
ACC occurs in species that require flexible cuticles (Neues, Ziegler, & Epple, 2007). The
formation of ACC occurs from a highly supersaturated solution when additives such as Mg, are
present that prevent deposition of the crystalline phases. By following the cuticle molt of
crustacean, it was established that magnesium switches off the phase transformation by
stabilizing amorphous precursors and the organic matrices. Both ACC and ACP can be well
stabilized by magnesium. It was found that the subsequent introduction of aspartate (Asp) can
effectively switch on the crystallization of these Mg-stabilized amorphous precursors in the
laboratory. The proposed magnesium-Asp based crystallization switch is another lesson inspired

131
from nature and this discovery provides an understanding of intelligent controls in
biomineralization (Tao, Zhou, Zhang, Xu, & Tang, 2009).
Effect of lipids/proteoglygans/alkaline phosphates
There is growing evidence supporting the hypothesis that phospholipids, are associated with
the initial mineralization of bone (Boskey & Posner, 1976). Other investigators have postulated
that the initiation of mineralization also involves the effect of proteoglycans, lipids, and alkaline
phosphatases (Nilsen, 1980).
Biopolymers
An underlying concept in biomineralization concerns the use of organized biopolymers to
exert detailed control over the nucleation and growth, the shape and size, and over the assembly
of crystals and other nanoscale building blocks into complex structures. Specific biopolymers
can dictate a strong influence on both crystal nucleation and growth rates as well as temperature,
pH, ionic strength and composition. One example of the influence of soluble biopolymers on a
crystallization event is the control of the CaCO3 crystal modifications using soluble
macromolecules extracted from the respective layers of a mollusk shell. Proteins can promote the
crystallization of a preferred mineral (Weiner & Dove, 2003). Another example is the
relationship between collagen and crystals which was first described by Robinson and Watson.
Crystal-like particles have been shown to be sequentially orientated in collagen fibrils (Nilsen,

132
1980). Glimcher and Krane suggested also that the collagen fibrils were the substrate for
nucleation and that the initial stages showed crystals in "holes" in the collagen fibrils (Nilsen,
1980).
The potential mechanisms for formation of organized biomineralized structures include
oriented crystal growth on templates, the aggregation of nanocrystals by oriented attachment, and
the assembly of inorganic nanoparticles mediated by organic molecules into aggregated
structures (Tao, Pan, Zeng, Xu, & Tang, 2007). The classical model of biomineralization
considers mineral formation as an amplification process in which individual atoms or molecules
are added to existing nuclei or templates, but a model involving aggregation-based growth has
gain traction recently and challenges the conventional concept of crystal growth. The brick and
mortar model attempts to explain the biological aggregation of nano-sized apatite by suggesting
that amorphous calcium phosphate acts as “mortar” to cement the crystallized “brick” of
nanosized HAP while biological molecules control the construction process. However, a new
concept of “mesocrystals” highlights the roles of nano-sized particles in biological crystal
engineering and is emerging as the current state of biomineralization theory (Dorozhkin S. ,
2012).
Nacre is one of the biological composites that contain both ceramic and biomacromolecular
phases. This unique structure is composed of alternating nanometer-scale laminated layers of
thin biomacromolecular matrix and CaCO3 (aragonite) platelets, all highly organized to produce
an excellent multifunctional material for the organism (Sarikaya, Liu, & Aksay, 1995). The
unique crystal structure of aragonite and the arrangement of the aragonite nanoparticles in

133
nacre’s platelets make the diffraction patterns of individual platelets show single-crystal
characteristics (Li & Huang, 2009). The tablet/platelet of nacre does diffract as a single crystal
but is made up of a continuous organic matrix which breaks the mineral up into coherent
nanograin which share the same crystallographic orientation (Rousseau, 2011).
Stages of Crystallization
Biogenic minerals may be amorphous, paracrystalline, or crystalline and may occur as a
single unit (such as a single crystal), numerous individual units, or aggregates. Mineralized
skeletal hard tissues are examples of such aggregates. The aggregated units are usually arranged
in an orderly fashion, and when crystalline, the crystallographic axes are partially or fully
aligned. These ordered structures are generally differentiated into a number of microarchitectural
units, each of which is enveloped by an organic matrix. Minerals formed at a particular site may
be (i) retained in place, (ii) transferred intact to other sites, (iii) excreted, (iv) dissolved and
replaced continuously, periodically, or only occasionally, or (v) continuously reconstituted.
Mineralized hard tissue can have a number of functions other than structural and mechanical
ones. It is well known that the endoskeletons of mammals act as a reservoir of calcium and
phosphorus and are intimately linked to the organism's metabolism (Lowenstam H. A., 1981).
The hierarchical structure of bone is illustrated in figure 4-8.
The formation of a mature skeleton composed of various shapes ultimately depends on
cellular control and the deposition of mineral within an organic matrix, and the creation of
specialized tissues. The mineral, a calcium phosphate, closely approximates hydroxylapatite,

134
Ca5(PO4)3(OH). The mineral in skeletal tissues is found as part of several different textures, e.g.,
woven, lamellar, haversian, trabecular and cortical bone that can be discriminated optically, or
histologically, and at higher resolutions using TEM or SEM. Bones and bone tissues fulfill their
structural function while protecting the marrow where essential cellular material is generated.
Sufficient bone stiffness is achieved by increasing the amount of calcium phosphate mineral
relative to the organic components in the tissue (Skinner & Jahren, 2003). The state in which
Ca2+ ions and PO43- ions are combined with the carboxylate groups on the organic matrices is the
earliest stage in crystal nucleation (Sato, 2007).
Crustaceans demonstrate drastic temporal differences within the same tissue with regard to
the extent of and capacity for mineralization. Such temporal differences raises questions about
the control of nucleation and mineralization. The cuticle is not homogeneous, but contains four
discrete layers. This observation was made more than a century ago (Williamson, 1860).
However, the terminology of Travis (1963) has come to be widely accepted. These layers from
the most external to the most internal are: the epicuticle, the exocuticle, the endocuticle and the
membranous layer (Roer & Dillaman, 1984).
Calcification of the crustacean cuticle is an extracellular phenomenon. Preparation for the
molt in the calcified crustaceans involves the resorption of the mineral and organic components
of the old cuticle and the simultaneous deposition of elements of the new cuticle. During
premolt, the entire organic matrix of the two outermost layers of the new cuticle (the epi- and
exocuticle) are elaborated beneath the old cuticle. The pre-exuvial layers do not calcify until
after the molt. The bidirectional nature of mineral transport and the sharp temporal transitions in

135
nucleating ability of the cuticular matrix provide ideal systems in which to study these aspects of
calcification (Roer & Dillaman, 1984).
During ecdysis, crabs periodically shed their rigid calcified exoskeleton. Immediately after
ecdysis, the cuticle is soft; thereafter the calcite mineral bestows progressive hardness to the
cuticle (Giraud-Guille, 1984). Calcification of the epi- and exo-cuticles begins in the most
external regions and proceeds proximally. Mineral apparently reaches the outer portions of the
cuticle via the pore canals. Calcium is concentrated in the distal portions of the epithelial cells
and appears to be extruded in vertical rows corresponding in position to the pore canals of the
new cuticle. Calcification is concurrent with matrix formation, each organic lamella being
mineralized as it is laid down. Mineral deposition continues through postmolt. Calcification
spreads throughout the exocuticle and eventually is found to pervade the walls of the pore canals.
The end of postmolt is marked by the deposition of the membranous layer and the cessation of
net calcium deposition (Roer & Dillaman, 1984).
The epicuticle is the outermost and thinnest layer of the cuticle. It consists of tanned
lipoprotein impregnated with calcium salts. The exocuticle immediately underlies the epicuticle.
The exocuticle is composed of chitin-protein fibers with biphase of calcite and amorphous
calcium carbonate, stacked in layers of continuously changing orientation. The exocuticle is
hardened by quinone tanning and calcification, with the mineral crystals situated between the
fibers (Roer & Dillaman, 1984). The crystallization in exocuticle is completed in postmolt stage
(Tao, Zhou, Zhang, Xu, & Tang, 2009). The endocuticle is the thickest and the most heavily
calcified layer of the cuticle. The endocuticle is composed of horizontal lamellae of chitin-

136
Figure 4-8 Hierarchical structural organization of bone: (a) cortical and cancellous bone; (b)
osteons with Haversian systems; (c) lamellae; (d) collagen fiber assemblies of collagen fibrils;
(e) bone mineral crystals, collagen molecules, and non-collagenous proteins (Rho, Kuhn-
Spearing, & Zioupos, 1998).
protein fibers with continuously changing orientation and is apparently not tanned, but hardened
solely by Ca salts (Roer & Dillaman, 1984). The endocuticle is calcified in late postmolt several
days after ecdysis. A membranous layer is also synthesized during postmolt and its completion
signals the onset of inter-molt. The membranous layer contains chitin and protein but is not
calcified. During the inter-molt stage, the isopod cuticle can be divided into epicuticle,
exocuticle, endocuticle, and membranous layer from outside to inside (Tao, Zhou, Zhang, Xu, &
Tang, 2009), Table 4-3.

137
Table 4-3 Designation of layers in the crab cuticle
Sequence of formation Chemical Composition
Preecdysial
layers
1 Epicuticle
2 Pigmented layer
Lipoprotein
Chitin-protein,
CaCO3
Ecdysis
Postecdysial
layers
3 Principle layer
4 Membranous layer
Chitin-protein,
CaCO3
Chitin-protein
(Giraud-Guille, 1984)
Crystallization Control
Kinetic control of crystallization can be achieved by modifying the interactions of nuclei and
developing crystals with solid surfaces and soluble molecules. Such processes influence the
structure and composition of the nuclei, particle size, texture, habit and aggregation, and stability
of intermediate phases. In biomineralization, for example, structured organic surfaces are
considered to play a key role in organic matrix-mediated deposition by lowering the activation
energy of nucleation of specific crystal faces and polymorphs through interfacial recognition.
Soluble macromolecules and organic anions, as well as inorganic ions can also have an important
kinetic effect on crystallization, particularly with regard to polymorph selectivity and habit
modification.
A central tenet of biomineralization is that the nucleation, growth, morphology, and
aggregation (assembly) of the inorganic crystals are regulated by organized assemblies of
organic macromolecules (the "organic matrix"). Control over the crystallochemical properties of

138
the bio-mineral is achieved by specific processes involving molecular recognition at inorganic-
organic interfaces.
Mann illuminated the role of molecular recognition in inorganic crystallization systems;
showed how organized organic surfaces can control the nucleation of inorganic materials by
geometric, electrostatic, and stereochemical complementarity between incipient nuclei and
functionalized substrates; and described how analogous interactions are responsible for the
morphological modification of inorganic crystals grown in the presence of anionic organic
additives. The surface lattice geometries, spatial charge distributions, polarity of hydration
layers, defect sites, and stereochemistries of the inorganic phase are all possible features that can
be recognized by the organic matrix (Mann, et al., 1993).
Biomineralization processes exhibit a high level of spatial control as the mineralization
usually takes place in a confined reaction environment. This confined reaction environment is
constructed by biomolecules—mainly polymers. They can build a scaffold for the mineralization
site as collagen in the example of bone. The mineralization event itself is also highly controlled
by biopolymers, which are usually soluble. The task of the soluble or functional matrix is the
control of nucleation, growth, polymorph and orientation of the inorganic compounds. It is
difficult to explore the function of the soluble matrix in biomineralization as it usually represents
a mixture of many biopolymers in tiny quantities so that a model system which mimics soluble
biomineralization polymers is an efficient way to understand biomineralization processes. The
functioning of the insoluble matrix and of the various components of the soluble matrix is a
highly synergetic process and only in some cases, the functions of the individual components of

139
the soluble matrix are already revealed. This makes the mimicking of biomineralization
processes difficult as often, only little is known about the formation of a biomineral archetype.
Furthermore, it is difficult to achieve the assembly of the structural matrix over several levels of
hierarchy because the synthetic chemist does not have cells at hand, which carry out this job in
nature. However, biomimetic mineralization can be successful if the formation of the natural
archetype is not yet completely understood. Frequently, a model system, which mimics soluble
and/or insoluble matrix is an efficient way to understand biomineralization processes and to
mimic them. Despite the complexity of the topic, characterization of biomineralization processes
and mechanisms has made significant progress in recent years often catalyzed by new findings or
concepts in this field (Xu, Ma, & Colfen, 2007).
Concluding Remarks
Biomineralization/Calcification/Mineralization/Crystallization is an important process in a
plethora of scientific disciplines. There is not a unified molecular and mechanistic descriptions of
crystal nucleation, growth, morphology. Biomineralization results in complex materials and
composites, such as bones and shells that are characterized by an extraordinary level of
molecular control of the particle size, structure, morphology, aggregation, and crystallographic
orientation of the mineral phases. Attempts to create synthetic materials that would imitate
mineral nucleation and growth have embraced a variety of approaches including biomimetic.

140
Understanding biomineralization of biological systems offers a genuine possibility for future
strategies that create higher-order architectures.

141
Chapter 5 Osteoinductive Nature of Blue Claw Crab
Introduction
There are many different concepts of engineering tissue. The desired properties of the
construct are defined by the intended clinical use (Kneser, et al., 2002). In 1993, Langer and
Vacanti summarized the early developments in this field and defined tissue engineering as “an
interdisciplinary field that applies the principles of engineering and life sciences toward the
development of biological substitutes that restore, maintain or improve tissue or organ function”
(Langer & Vacanti, 1993). Tissue engineering (TE) uses a set of tools at the interface of the
biomedical and engineering sciences to support the growth of living cells or attract endogenous
cells to aid in tissue formation or regeneration to produce therapeutic or diagnostic benefit. TE
also seeks to understand structure-function relationship in normal and pathological tissues; and to
control cell and tissue responses to injury, physical stimuli, and biomaterials surfaces through
chemical, pharmacological, mechanical, immunological, and genetic manipulation. The key
processes occurring during the in vitro and in vivo phases of tissue formation and maturation are:
(1) cell proliferation, sorting and differentiation; (2) extracellular matrix production and
organization; (3) degradation of the scaffold; and (4) remodeling and potentially growth of the
tissue (Schoen, 2013).
In bone tissue engineering, the emerging trend is to employ materials that play an active role
in regeneration, rather than passive or inert roles. Marine invertebrates offer inexhaustible
inspiration (Ehrlich H. , 2010). Marine sources such as coral, algae, and nacre have been

142
evaluated as potential orthopedic or dental implants. Evaluations of corals as potential bone graft
substitutes began in the early 1970s in animals and in 1979 in humans (Demers, et al., 2002).
Nacre investigations dates back as early as 600 A.D. with the ancient Mayan civilization
adapting nacre as a bone implant material (Westbroek & Marin, 1998).
Bone tissue engineering is based on the understanding of tissue formation and regeneration
and integrates knowledge from physics, chemistry, engineering, materials science, biology and
clinical medicine into a comprehensive interdisciplinary approach. The goal of bone tissue
engineering is to regenerate a functional structure containing osteoblast, osteocyte, and osteoclast
cells capable of building and of continuously remodeling the mineralized collagenous
extracellular matrix that functions structurally and biomechanically as bone tissue (Kneser, et al.,
2002).
Bone tissue engineering requires the interaction of cells, growth factors, and extracellular
matrix. Several studies both past and present demonstrate the effectiveness of these elements
(Hing, 2004). In 1889, Senn noticed that decalcified bone can induce healing of osseous defects
(Senn, 1889). Urist (1965) discovered that demineralized bone when implanted intramuscularly
induced new bone formation. In 1971, Urist and Strates presented ‘bone morphogenetic protein’
(BMP) and ‘osteoinduction’ to the scientific and medical communities (Urist & Strates, 1971)
and later Urist coined osteoconduction. By 1976, Reddi and Anderson further expounded on
Urist's work dealing with ‘induced osteogenesis’ (Reddi & Anderson, 1976) and provided a
compelling explanation of the functional role of purified organic bone matrices (Gruskin, Doll,
Futrell, Schmitz, & Hollinger Jeffrey, 2012).

143
Large bone defects, resulting from a multitude of causes, including aging, trauma, and
disease, are commonplace. These defects usually do not heal spontaneously and interventions
with bone substitutes are often required. Natural bone uses osteogenic (progenitor osteoblasts)
cells; osteoconduction (structural support system); and osteoinduction (growth factors) to
produce a mineralized collagen matrix that hardens (Stocker & Wolinetz, 2009). Ideally, a bone
replacement material should mimic bone tissue from a mechanical, chemical, biological and
functional point of view, and facilitate new bone formation. None of the existing implant
biomaterials are optimized in terms of performance in bone replacement (Hertz & Bruce, 2007).
Emerging as a valid therapeutic approach, bone tissue engineering is based on understanding
hard tissue formation and targets induction of new functional tissues. Among hard tissue
engineering strategies, the selection of a suitable material is paramount. Surveying the
characteristics of bone, crab carapace materialized as a material similar to bone based on factors
such as high strength, stiffness, and hierarchical structuring. Thus, crab cuticle is another
candidate material for enhancing the healing, remodeling, and engineering of bone. To date, our
lab conducted the first preliminary biocompatibility study to determine crab cuticle’s suitability
as a bone substitute material (Wilson, Gugssa, Mehl, & Anderson, 2012). The paper addresses
the osteoinductive nature of crab cuticle, specifically its ability to facilitate the formation of a
mineralized collagen matrix.

144
Bone Grafts and Bone
Bone grafts, synthetic, and natural materials are used to assist and to build bones after
damage from disease, trauma, or aging (Yuan & Groot, 2004). Bone is noted for its strength and
hardness and is characterized by its composite structure consisting of organic matrix (35%) and
inorganic elements (65%). The inorganic constituent of bone consists mainly of calcium
phosphate, considerable amounts of citrate and carbonate ions and traces of magnesium fluoride,
and sodium. The organic matter in bone is largely collagen. The calcium phosphate in bone
resembles mineral hydroxyapatite (HAP) (Yuan & Groot, 2004), and contains carbonates,
sodium, magnesium and fluoride salts (Hing, 2004). See Table 5-1.
A dynamic living tissue, bone consists of osteocytes and osteoblasts, living cells that build
bone. During bone remodeling, bone has osteoclasts, living cells that resorb bone (Yuan &
Groot, 2004). Bone is a remarkable composite, and its unique functional characteristics include a
high level of vascularity, the ability to remodel and repair itself, maintaining mineral
homeostasis and providing a source of stem cells, acting as a mineral and blood cell reservoir
(Hing, 2004).
Even when fully mature, bone is not static. There is a constant repairing, modeling, and
remodeling that continues throughout life (Summerlee, 2002). Bone can suffer from disease and
is subject to deterioration as a result of age or trauma (Hing, 2004). When complicated or large

145
Table 5-1 The composition of the inorganic and organic phases of bone
(Murugan & Ramakrishna, 2007)
fractures and defects occur in bone, the healing and repair often fails and requires surgical
intervention. Tissue engineering strategies are explored to repair or restore damaged bone tissue
with the assistance of biomimetics, biomaterials, cells, and/ or signal molecules (Janicki &
Schmidmaier, 2011), figure 5-1. There are three characteristics inherent to natural bone:
osteoconductivity; osteogenicity; and osteoinductivity (Stocker & Wolinetz, 2009).
Autologous bone grafts are the gold standard for current therapeutic strategies. Autologous
bone combines the three characteristics inherent to natural bone to promote growth and
regeneration: osteogenic cells, osteoinductive and osteoconductive properties. Autografts have
limitations such as donor site morbidity caused by surgical procedures from harvesting, risk of
infections, and chronic pain (Janicki & Schmidmaier, 2011). Allografts avoid some of the
disadvantages of autografts; however allografts have other limitations like transmission of
diseases from donor to recipient or immunogenic reactions (Janicki & Schmidmaier, 2011).

146
Xengografts are tissues harvested from one species and implanted into a different species (Bauer
& Muschler, 2000). Alloplasts include any synthetically derived graft material not derived from
animal or human origin (Suneelkumar, Datta, Srinivasan, & Kumar, 2008). Allografts,
xenografts, and alloplasts all have disadvantages and limitations. An alternative to auto-, allo-
and xenografts are tissue engineered (Janicki & Schmidmaier, 2011) or biomimetic bone
substitutes. Figure 5-2 depicts the past, present and future of tissue repair. Synthetic materials
have widely broadened the available tools and strategies for bone grafting and vary greatly in
osteoconductivity and osteoinductivity, and in mechanical strength, handling properties, and cost
(Bauer & Muschler, 2000).
Figure 5-1 Schematic of the design strategy of tissue-engineered biomimetic nanocomposite
bone graft (Murugan & Ramakrishna, 2007).

147
Bone substitutes can be generally divided into two main groups: biological and synthetic
materials. Biological bone substitutes consist of materials like natural polymers (like collagen
type I or demineralized bone matrix (DBM). Materials like porous metals or bioactive glasses,
synthetic polymers and calcium phosphates rank high amongst synthetic materials (Janicki &
Schmidmaier, 2011) used as bone biomaterials. Bone grafts types have limitations and
disadvantages that motivate the development of alternatives (Stocker & Wolinetz, 2009).
Figure 5-2 Depiction of the past, present and future of tissue repair (Hench, 1998).

148
The constraints of common bone defect treatment strategies have triggered exploration of new
biological (Holzapfel, et al., 2013) and biomimetic approaches. The three key ingredients for
both morphogenesis and tissue engineering are inductive signals, responding stem cells, and the
extracellular matrix (Reddi, 2000). The classic paradigm of tissue engineering comprises a cell
seeded scaffold, an in vitro stage of tissue formation, and an in vivo stage of tissue growth and
remodeling or an unseeded scaffold, with biological information such as growth factors, to
attract precursor cells and cause differentiation in vivo (Mendelson & Schoen, 2006). In a
biomimetic approach to tissue engineering, natural materials or lessons from nature are exploited
to solve tissue engineering problems. Natural materials can be used with the aspiration to control
the behavior cells and promote the attachment, migration, proliferation, and differentiation of
cell types of interest.
Materials harvested from natural sources are used extensively in bone tissue engineering.
These materials may be derived from a wide range of sources and possess an equally wide range
of physical and biological characteristics. For example, coral was used as a bone graft material in
the mid 1970’s. Coral’s porous CaCO3 exoskeleton displays a unique ability to bond with bone
(White, et al., 1975). Another natural material chitin is derived from the exoskeletons of
crustaceans such as crab and shrimp and is the second most abundant natural polymer after
cellulose. Chitosan, a derivative of chitin, elicits a minimal foreign body reaction, with little or
no fibrous encapsulation (Monzack, Rodriguez, McCoy, Gu, & Masters, 2011). Chitosan,
chitosan derivatives, and blends of chitosan have been used in bone tissue engineering

149
applications. Costa-Pinta et. al. provides an extensive review of chitosan’s role as a scaffold in
bone tissue engineering (Costa-Pinto, Reis, & Neves, 2011).
Osteoinduction, Osteoconduction, and Osteogenic
Giving the limitations associated with current bone graft technologies, implementation of
bone tissue repair strategies aims to incorporate the advantages of allografts and autografts
(Nicoll, 2011)as well as the characteristics inherent to bone. Osteoconduction highlights bone
formation towards the implant from the host bone bed; guided bone formation on material
surfaces resulting in bone bonding (sometimes called “bioactivity”, “osteocoalesce”, and
“osteoingreation”); and the absence of other tissues between the newly formed bone and
biomaterial surface (Yuan & Groot, 2004). A distinction can be made between, osteoconduction
and osteointegration. Both depend on biological factors, but osteoconduction may be rather short
lived and successful osteointegration maintains its bone anchorage over an extended period
(Albrektsson & Johansson, 2001). Ensuring the continuity of the repaired bone both structurally
and mechanically, osteoconductive biomaterials form a biological apatite layer in vivo on its
surface where osteogenic cells can easily attach and form bone on it (Yuan & Groot, 2004).
Osteoconductivity respresents structure and the ability of a biomaterial to serve as scaffolding
to support new bone formation and growth (Stocker & Wolinetz, 2009). Osteoconductive
biomaterials influences bone regeneration. On osteoconductive biomaterial surface, guided bone
formation is limited in its distance spotlighting that osteoconductive biomaterials alone may not
repair large bone defects (Yuan & Groot, 2004). Osteogenicity represents the presence of

150
osteoblasts or stem cells that can transform into cells responsible for bone formation (Stocker &
Wolinetz, 2009). Osteoinduction entails the recruitment and stimulation of primitive,
undifferentiated and pluripotent cells to develop into bone-forming cell lineage that ultimately
produce bone (Albrektsson & Johansson, 2001). Conclusive evidence for osteoinduction is
characterized by heterotopic bone formation, that is, bone formation in tissues or organs where
bone does not naturally grow (Barradas, Yuan, van Blitterswijk, & Habibovic, 2011) . Although
the exact processes involved in the mechanism of osteoinduction by biomaterials are still largely
unknown, work by many groups has shown that biomaterials need to meet very specific
requirements in terms of (a) macrostructure, (b) microstructure and (c) chemical composition in
order to be osteoinductive (Habibovic & de Groot, 2007).
This initial part of bone fracture healing response includes osteoinduction, a process that
starts immediately after the injury. Osteoinduction is responsible for the majority of newly
formed bone. Osteoconduction depends to a fairly large extent on previous osteoinduction
(Albrektsson & Johansson, 2001).
The phenomenon of bone formation upon implantation of various tissues heterotopically has
been documented: (Huggins, 1931), (Levander, 1934), (Bertelsen, 1944), (Urist & McLean,
1952), (Moss, 1958). Urist’s discovery that decalcified bone matrix induced bone formation in
muscles of mouse, rat, guinea pig and rabbit, subsequent identification of Bone Morphogenetic
Proteins (BMPs) as inducers of heterotopic bone formation, and definition of autoinduction, or
osteoinduction as “the mechanism of cellular differentiation towards bone of one tissue due to
the physicochemical effect or contact with another tissue” set a benchmark in his field of

151
research (Barradas, Yuan, van Blitterswijk, & Habibovic, 2011). Other osteoinduction
definitions have been proposed. Wilson-Hench described osteoinduction as the process by which
osteogenesis is induced (Barradas, Yuan, van Blitterswijk, & Habibovic, 2011).
Biomaterials with intrinsic osteoinductivity possess a great potential as alternatives to
biological approaches to bone regeneration. de novo bone origin is attributed to its osteoinductive
properties rather than to the osteoconductive. The capacity to repair bone defects can differ
greatly among materials from the same family. Apart from the chemical composition of the
material, the geometry and macrostructural properties have been shown to play an important
role. In the case of macrostructure, the most striking example is the importance of porosity.
(Barradas, Yuan, van Blitterswijk, & Habibovic, 2011)
Bone formation by osteoinduction is initiated by soluble and insoluble signals that trigger a
complex cascade of molecular and cellular morphogenetic processes that ultimately leads to the
sculpture of precisely organized multicellular mineralized structures (Ripamonti & Roden,
2010). Turing was the first to describe a system of chemical substances, called morphogens,
account for the main phenomena of morphogenesis (Turing, 1952). Secreted from a group of
cells or organizing center, morphogen molecules can provide spatial information and then moves
away. Cells can detect where they are in respect to the organizing center as the activity of the
morphogen decreases gradually as a function of its distance from the source. The cells are
induced by the morphogens to take on different fates according to their position (Vincent &
Perrimon, 2001). Moreover, it is believed that several mineralized and non-mineralized
extracellular matrices of mammalian tissues contain morphogens that direct specific cell

152
differentiation in responding stem cells generating tissue form and function (Ripamonti &
Roden, 2010).
Ossification
Bone forms in a process termed ossification. There are some distinctions that can be made
about bone development in two critical phases: either embryonically or postembryonically.
During embryonic or in utero development, bone tissue starts to form mesenchyme cells. During
the second phase, bone elongate and change shape. There are two types of embryonic
ossification: intramembranous ossification and endochondral ossification. Intramembranous
ossification involves mesenchymal cells migrating, becoming osteoblasts, forming aggregations
in organizing or ossification centers, producing bone matrix that undergoes calcification,
encasing osteoblasts to emerge as osteocytes, growing linear extensions called spicules from the
ossification centers, growing of blood vessels branching around the spicules, fusing spicules in a
network to eventually create flat bones. Endochondral ossification includes long, short, and
irregular bones and involves bone development from a pre-existing cartilage model where a rod
of cartilage develops in the expected final position and mimics the general shape of the adult
bone (Summerlee, 2002).

153
Calcification/Mineralization
Usually limited to bones and teeth in vertebrates, mineralization and calcification are terms
used interchangeably to describe the process in which tissues are converted to hard tissues.
Matrix mineralization and intact bone cells characterize physiological mineralization (Robey,
2011). Bone formation or osteogenesis can also occur outside the skeleton. Ectopic ossification
happens when precursor cells inappropriately receive signals to develop into bone cells, to
synthesize extracellular matrix and to create an environment facilitating mineralization. Matrix
mineralization is the final step of osteogenesis (Jahnen-Dechent, 2004). Figure 5-3 hypothesizes
a mechanism behind osteoinduction of biomaterials.
Pathologic calcification is a complex regulated process of mineralization that is similar to
bone formation and remodeling. Pathologic calcification refers to the deposition of calcium
phosphates or other calcific salts at sites, which would not normally have become mineralized.
The sizes of the mineral crystals in pathologic calcification are similar to those in bone; however,
there is much more mineral in the deposits than there is in bone (Ciftcioglu & McKay, 2010).
Pathologic calcification can thus occur before and after osteogenesis (Jahnen-Dechent, 2004).
There are two forms of pathologic calcification: dystrophic calcification and metastatic
calcification. Dystrophic calcification occurs in dying or dead tissue that is able to maintain
normal calcium homeostasis. Dystrophic calcification is distinguished by the lack of identifiable
osteoblasts and osteocytes, and lack of marrow (Robey, 2011). Metastatic calcification is the
deposition of calcium salts in otherwise normal tissues due to abnormal biochemistry with
disruptions in the calcium or phosphorus metabolism (Tam, Wang, & Fan, 2002).

154
Figure 5-3 Diagram illustrating hypothesized mechanisms behind osteoinduction by
biomaterials. Physico-chemical and/or structural properties of osteoinductive biomaterials may
prompt the mechanism responsible for heterotopic bone formation (Barradas, Yuan, van
Blitterswijk, & Habibovic, 2011).
Ectopic calcification is defined as biomineralization occurring in tissues where it is not
normally found. Ectopic calcification or ossification can be mistaken for pathological
calcification and can develop at the same site. There are two types of ectopic bone: heterotopic

155
and osseous metaplasia. Heterotopic bone is essentially normal osseous tissue found in an
abnormal location that is believed to originate from embryonic cells that persist in tissues during
development. Osseous metaplasia involves the transformation of a cell or tissue from one type to
another and can develop from connective tissues at sites of chronic inflammation (Shehab,
Elgazzar, & Collier, 2002).
Inflammation
All space within the body of an organism is filled; therefore the penetration of a biomedical
device is almost inevitably going to result in injury (Ramsden, 2008).
The interface between the implant and the biological environment is the location of proteins
and other biomolecules that interact with implant as well as the location of the body’s wound
healing process (Dee, Puleo, & Bizios, 2002). Materials implanted into tissues by surgical
techniques involve the creation of a wound. The inflammatory response is an early step in the
wound healing process. Just as the initiation and the magnitude of inflammation are regulated by
the characteristics of the biomaterial, the resolution of the inflammatory process is also
dependent on the physical and chemical properties of the biomaterial. These attributes will
determine the overall resolution of the wound healing process (Greco, 1994) and hence the
bioactivity of the implant.
Inflammation is qualitatively separated into acute and chronic inflammation. Acute
inflammation is a stereotypic response, generally of short duration and lasting less than a week.

156
Figure 5-4 Schematic depicting the material, interface, and biological enviroment components.
It emphasizes the importance of the interface.
It is characterized by an increased capillary permeability which results in the exudation of
plasma proteins and emigration of polymorphonuclear neutrophilic leukocytes (PMN) to the site
of inflammation. Chronic inflammation is of longer duration and is histologically associated
with macrophages, lymphocytes, proliferation of connective tissues, and deposition of
matrixproteins (Greco, 1994). The inflammatory response to biomaterials is determined by their
composition, purity, shape of the implant and its surface properties, implantation site, positional
stability at the implantation site, porosity, chemical stability, etc. (Mainil-Varlet, Gogolewski, &
Nieuwenhuis, 1996). The material-interface is an important factor in biological response, figure
ions
proteins
blood
enzymes
coagulation
inflammation
proliferation
remodeling
resolution
Biological Environment Interface Material

157
5-4, as it is the place of the first interactions with the biological environment. The time course
varies with severity and nature of the injury (Black, 1999).
Biologically, macrophage are perhaps the most important cell type in chronic inflammation.
Macrophages appear at the site of biomaterial implantation due to their role in wound healing in
response to surgery, as well as in response to the biomaterial itself. As a rule, macrophages can
be seen adhering to the biomaterial within 24 hours after implantation. The number of
macrophages attracted to the implant is in part dependent on the texture of its surface.
Macrophages are capable of producing a variety of growth factors, cytokines, chemotactic
agents, proteases, metabolites, oxygen free radicals, complement components, and coagulation
factors. These macrophage products may have a wide spectrum of effects, from cell death to cell
proliferation, and thus may impact on the course and outcome of the chronic inflammatory
process (Greco, 1994). Macrophages react to and generate, signals that influence growth,
differentiation, and death of other cells (Paul, 2008). The activated macrophage can undergo a
number of transformations. In addition to releasing lysozymes, both internally and externally,
related to attempts to digest foreign materials, activated macrophages synthesize and release a
wide range of biochemical factors that can mediate the activity of many other cells, including
lymphocytes, fibroblasts, osteoblasts, and osteoclasts (Black, 1999).
Osteoinductive materials provide a biological stimulus for recruitment, stimulation and
differentiation of primitive, undifferentiated and pluripotent cells into osteoblast or
preosteoblasts, which are the initial cellular phase of a bone-forming lineage. Bone formation is

158
essentially achieved by the interaction of cells, growth factors, and an implant material and the
facilitation of an appropriate cellular response (Di Silvio, 2009).
Experimental Section
Materials
Perfusion solutions for animal experiments were a normal saline solution and a fixative
solution (1:4 ratio). The saline solution consisted in a 0.9% NaCl solution with a checked pH of
7.4. The fixative solution is a 1%w/v Paraformaldehyde , 2%w/v Gluteraldehyde in a 0.1M
cacodylate buffer with pH adjusted to 7.4. The preparation of 200 ml/animal was sufficient for
perfusion. Various sets of C. sapidus (Blue Claw Crab) was purchased from commercial sources.
The Sprague-Dawley female rats were purchased from Harlan (Frederick, MD). The Sprague-
Dawley female rat experimental protocols are following the Animal Welfare Care and Use
Committee (IACUC) directives from Howard University. All animal experiments were
performed at Howard University in the Biology Department under the direction of Dr. Anderson.
Crab integument preparation
Carapace integument from fresh and fresh frozen C. sapidus (Blue Claw Crab) specimens
were removed and thoroughly cleaned with deionized water and allowed to dry. Carapace was
initially crushed using a commercial coffee grinder using a pulse cycle. Subsequent size
reduction was accomplished using a mortar and pestle. Crushed crab shell samples were washed

159
and aged in absolute ethanol or 70 vol.% ethanol as an initial step in sterilization. A portion of
the crushed crab shell was demineralized by aging 2 g of dry crab shell in 20 ml of 0.5 M HCl.
The crab shell granules were allowed to react in the HCl for 1–3 h. The demineralized crab shell
was washed with deionized water and finally rinsed with ethanol and allowed to dry in air
(Wilson, Gugssa, Mehl, & Anderson, 2012).
Characterization
Transmission electron microscopy (TEM), High resolution transmission electron microscopy
(HRTEM), selected area diffraction (SAD), electron nanodiffraction (END), electron
microanalysis - energy-dispersive spectrometry (EDS) was performed at the NISP
characterization facility at the University of Maryland; in vitro and in vivo studies were used to
investigate the osteoinductive nature of Blue claw crab.
TEM enabled observation of ultrastructure of our sample and allowed for histological
assessment. HRTEM allowed even finer detailed observation in the nanoscale. SAD was used to
identify crystal characteristics. END offered atomic resolution for nanosized crystal. X-ray
microanalysis was used to determine the elemental composition of the specimen.
TEM has different forms and offers a broad range of characterization techniques with high
spatial and analytical resolution. TEM is the central tool for characterization of nanoscale
materials. The advantages of using TEM is that a small amount of materials is needed, strong
diffraction with matter at an atomic resolution, can handle nano-size crystals, and precise method
of structure determination. Despite all the advantages, TEM bring accompanying drawbacks.

160
One drawback to TEM techniques is that many materials require extensive sample preparation to
produce a sample thin enough to be electron transparent, which makes TEM analysis a relatively
time consuming process with a low throughput of samples. Another disadvantage is that the
structure and chemistry of the sample may also be changed during the preparation process.
Additionally, the field of view is relatively small, raising the possibility that the region analyzed
may not be characteristic of the whole sample. The sampling abilities of the instrument is greatly
comprised with the higher resolution. Another problem is TEM displays 2D images of 3D
specimens. A single TEM image has no depth sensitivity. This problem can be circumvented
with a series of images taken a t different tilts to create 3D image. Other disadvantages include
image artifacts and defects. Electron beam damage, particularly in the case with biological
materials, is also a concern (Williams & Carter, 2009).
HRTEM is a form of TEM and has many of the same advantages and disadvantages. HRTEM
maximizes the useful detail in the image but at the cost of sampling abilities. HRTEM, SAD and
END were used in a complementary fashion. SAD and END were techniques employed inside
the TEM. SAD can be useful for the determination of lattice parameter information and unit cell
details. Lattice structure gives information about the long range composition of a material.
Atomic structure is a huge amount of data compared to what any other biochemical technique
could provide. Restrictive factors of SAD are that a good crystal must be found for high accuracy
of crystallographic structures and that only limited information about the structure’s dynamic
behavior can be obtained from one single diffraction experiment. Resolution is an important
limitation of SAD. Small particle size can lead to peak broadening and erroneous interpretation

161
of a SAD pattern. If a SAD pattern of a polycrystalline aggregate has many rings, the number of
rings may be too great and closely spaced for effective measurement. Thickness variation and
crystal defects can make complete and specific identification of each small crystal difficult even
impossible (Ferrell Jr. & Paulson, 1977). For nanodiffraction, radiation damage effects can be
extreme and biological samples can be especially vulnerable. However, radiation damage effects
are not of great significance for nanodiffraction provided that care is taken to prevent the beam
from staying for an excessive amount of time in any one position. In nanodiffraction, structurally
ordered substances can become amorphous and loss some of its weight limiting the attainable
resolution (Cowley & Spence, 2000).
EDS or x-ray microanalysis was performed inside the TEM as well. Accurate quantitative
analysis requires calibration of the x-ray analysis system using standards of known composition
and thin specimens. Large differences in the self-absorption and fluorescence of emitted x-rays
will limit the precision of quantitative analysis for some combinations of elements.
In vivo crab shell implantation
Female Sprague-Dawley rats ranging in weight from 400 to 450 g were used in this study.
The animals were housed in plastic cages in a room with the temperature in the range of 70-74ºC
and humidity in the range of 35-45%. All animals were fed a rodent chow (Harlan, Teklad
Laboratory) diet and water.

162
Figure 5-5 The implantation study has an aim to assess the histological response to the crab
cuticle and determine its potential as as a bone biomaterial.
In our collaboration with Howard, approximately 48 rats were used to address the question of
crab shell influencing bone formation using an osteoinductive model involving subcutaneous
implantation of crushed crab shell. The focus of my study involved analyzing tissue samples that
were obtained after 28 days of aging. The following protocols were used in implanting,
removing, and preparing tissue samples for analysis.
Each rat was slightly anesthetized with Halothane (Sigma) just for the duration of the crab
shell implantation. The abdominal skin was initially cleaned with ethanol. A subcutaneous
incision was made in the abdominal region of anesthetized twenty eight day old Sprague–
Dawley rats and approximately 10–20 mg of demineralized or untreated crab shell was
Incision Site
Objective
The goal of this work was to assess the histological response
or changes to a crab shell stimuli in a Wistar rat animal model;
Assessment
Consideration of the cellular changes, cell migration, proliferation,
and differentiation;
Determination
The potential of crab shell use as a bone implant.

163
implanted. The implant site was closed with stainless steel staples and rinsed with ethanol. The
samples were retrieved at 1, 3, 6, and 28 days, respectively, after implantation.
The protocol for retrieval of the tissue close to the implant was as follows: the rats were
anesthetized slightly with halothane and maintained with temporarily halothane exposure during
the whole protocol. The rat abdominal cavity was opened and then the diaphragm was also
opened to expose the beating heart. The blood was then flushed out from the whole animal by
perfusing the saline solution through the left ventricle until all organs became pale. The saline
solution was replaced by the fixative solution to fix the whole animal and in particularly the
tissue close to the crab shell implant. This tissue from the subcutaneous area of the implantation
was cut-off from the skin side as presenting implants being encapsulated. Tissue for TEM
analysis were post fixed with 1% Osmium Tetroxide (OsO4) then stained with lead acetate
before being dehydrated using increasing ethanol concentration aqueous solutions until a final
dehydration in 100% Ethanol. The samples were then embedded in epoxy in preparation for
ultramicrotoming. Thin sections about 90 nm thick were cut using a diamond knife and mounted
on C coated Cu transmission electron microscopy (TEM) sample grids (Wilson, Gugssa, Mehl,
& Anderson, 2012)
A rat subcutaneous implantation model was used to evaluate the in vivo tissue response,
figure 5-5. The healing response triggered by placement of bone implant in subcutaneous tissue
include the host inflammatory reaction to trauma associated with surgical procedure and
implantation of the whole crab material, the inflammatory and immune reaction to the material
itself, and the cellular processes such as migration, proliferation, and differentiation. The speed

164
and degree of this response and incorporation of the implant into host tissue is dependent on
local and systemic factors, namely type of implant material and surrounding tissue integrity, and
the systemic physiological state of the host (Di Silvio, 2009)
Some Preliminary Results
A short term assessment of subcutaneous implantation of demineralized crab shell in the
abdominal region of 28 day old Sprague–Dawley rats results are shown in Figures 5-6 and 5-7.
The cellular events occurring at the material/tissue interface and material itself depicts
macarophages or monocyte type cells in close proximity to a demineralized crushed Calinectes
Sapides (Blue Claw) crab shell particle which displays Bouligand structures, figure 5-7.
Figure 5-6 Transmission electron microscopy image of in vivo study of demineralized crushed
Calinectes Sapides (Blue Claw) crab shell implanted subcutaneously in abdominal region of 28
day old Sprague–Dawley rats. Aging time was one day and image displays a crab shell particle
in the bottom of the image with a macrophage in close proximity (Wilson, Gugssa, Mehl, &
Anderson, 2012).

165
Figure 5-7 TEM pictures depicts macrophages surrounding the Bouligand structure of
demineralized crushed Calinectes Sapides (Blue Claw) crab shell (Wilson, Gugssa, Mehl, &
Anderson, 2012).
Figure 5-7 also depicts macrophages surrounding the Bouligand structure. The implantation
time was 1 to 6 days. The nucleus displays marginal and central clumping of chromatin material
indicative of fully differentiated cell in which little active synthesis or energy expenditure is
occurring (Porter & Bonneville, 1973). The macrophage or histiocyte is irregularly shaped, has
considerable cytoplasm, a rounded nucleus and a nucleolus. Characteristic of the macrophage is
the presence of numerous lysosomes, which are related to its phagocytic function. Lysosomes
have a heterogeneous electron-dense content and an abundance of proteolytic enzymes, and are
enclosed by membranes which isolate them from the remainder of the cytoplasm. The lysosomes
participate in the intracellular digestion of phagocytosed material (Laguens & Gomez Dumm,
1969). Macrophages are monocytes that have differentiated after entering tissues at sites of

166
infection or inflammation (Janeway, Travers, Walport, & Shomchik, 2005). In responding to
these important mediators, macrophages infiltrate the wounded area and assist in cleaning and
removing damaged tissue debris and foreign particles. Once in the wound site, activated
macrophages release several important growth factors and cytokines. Considered to be the most
important regulatory cell in the inflammatory reaction, macrophages become the predominant
cell type during the latter part of the inflammatory phase. Macrophages phagocytize, digest, and
kill pathogenic organisms; scavenge tissue debris; and destroy any remaining neutrophils.
Macrophages release chemotactic factors that attract fibroblasts to the wound area (Li, Chen, &
Kirsner, 2007).
Figure 5-8 Transmission electron microscopy image of collagen bundles that formed after
subcutaneous implantation of demineralized crab shell for 6 days. The image shows 67 nm
banding pattern and regions of electron dense mineralized particles on the collagen fiber surface
and interspersed in between collagen fibers (Wilson, Gugssa, Mehl, & Anderson, 2012).

167
The initial results of the potential of implanted crab shell to induce ectopic bone formation
were reported (Wilson, Gugssa, Mehl, & Anderson, 2012) in figure 5-8. Our idea is based on the
work of Urist (1965) in which he demonstrated that demineralized bone matrix (DBM) possessed
the ability to induce new bone formation in non-osseous sites such as subcutaneous zones or
intramuscular regions. Studies on the osteoinductive properties of DBM and calcium phosphate
have been reported: (1) Reddi and Huggins (1974) (1972) and Reddi and Anderson (1976)
showed that the bone induction process proceeded via an endochondral mechanism in which the
implanted demineralized bone matrix was transformed to a cartilage type plaque by the action of
macrophages and fibroblasts which transformed to chondroblasts to facilitate cartilage tissue
formation. The chondroblasts were in turn transformed to osteoblasts and the cartilage
mineralized to complete the endochondral bone formation process. Studies (Reddi, 1981),
(Sampath & Reddi, 1981), (Reddi, 2000) later showed that DBM contained a family of
osteogenic extracellular matrix (ECM) proteins which came to be known as bone morphogenetic
proteins (BMP) which promoted various activities that are necessary for bone formation. Bone
formation induced by BMP or BMP-containing matrix is the most typical example of
osteoinduction (Reis & Weiner, 2004) (2) Ectopic bone formation was exhibited by inorganic
ceramics such as calcium phosphate and calcium carbonate: Yamasaki (Yamasaki & Sakai,
1992) reported bone formation when hydroxyapatite was implanted subcutaneously in dogs; and
Ripamonti (1991) induced bone formation in Baboons with porous hydroxyapatite obtained from
the conversion of calcium carbonate exoskeletons of coral. Crab shell exoskeleton’s main
constituents are chitin and calcium carbonate. The osteoinductive potential of various materials

168
differed based on starting time, amount of bone induced, type of material, geometrical
parameters such as microstructure, the chemistry, and type of host or animal (Reis & Weiner,
2004). As indicated previous, crab cuticle is strikingly similar to bone. Coupled with our initial
assessment of collagen fibers produced with the implantation of demineralized crab shell and
previous studies with calcium phosphate, coral, nacre, and similar materials, the idea for using
crab shell as an osteoinductive bone biomaterial showed tremendous promise. The surface
topography of biomaterials is especially important to guide cellular behavior like cell movement
and adhesion that is regarded as being very important for promoting wound healing and tissue
regeneration (Lamers, et al., 2010).
Analysis
Crab shell or cuticle, replaced at each molt, is secreted by epithelial cells. It varies in
thickness and hardness according to its location. The fully formed cuticle of a crab is a
proteinaceous structure that consist s of an outer later, the epicuticle, and underlying exocuticle
(pigmented layer), endocuticle (calcified layer), and membranous layer (uncalcified layer).
Chitin occurs in all but the epicuticle, and calcium, mainly as CaCO3, but also as Ca3(PO4)2 and
CaSO4, is found to varying degree and, depending on location, in all but the membranous layer.
In the blue crab, calcium occurs in parts of the epicuticle; its presence is species dependent. The
epicuticle is the first line of defense between the animal and its environment; contains lipids,
proteins, and usually calcium; and is relatively impermeable to water and salts (Johnson, 1980).

169
Figure 5-9 (a) SEM micrograph and a schematic drawing of a cross-sectional surface showing
three different layers in the crab exoskeleton: epicuticle, exocuticle, and endocuticle (Chen, Lin,
McKittrick, & Meyers, 2008); (b) TEM micrograph of demineralized crushed Calinectes Sapides
(Blue Claw) crab shell displaying prominent Bouligand nested arc patterns and a schematic
drawing of the Bouligand structure showing the arc pattern on oblique surface.
Bouligand Structure
Epicuticle
Exocuticle
Endocuticle

170
A prominent feature of crustacean exoskeletons is their well-defined hierarchical
organization, which reveals different structural levels, figure 5-9. A twisted plywood structure,
called Bouligand structure, is created by planes that are stacked in a helicoid. This nesteted arc
pattern is illustrated in the TEM and schematic image shown in figure 5-9. These structures
repeat to form the exocuticle and endocuticle. The same Bouligand structure is also characteristic
of collagen networks in compact bone and other fibrous materials (Chen, Lin, McKittrick, &
Meyers, 2008).
Figure 5-10 Transmission electron microscopy image of collagen bundles that formed after
subcutaneous implantation of mineralized crab shell for 28 days. Similar to the image of
(Wilson, Gugssa, Mehl, & Anderson, 2012), it depicts collagen fibers with the characteristic 67
nm banding pattern.

171
Figure 5-11 Transmission electron microscopy image Collagenous fibers, collagenous bundles,
and fibroblasts. Fibers oriented in longitudinal directions exhibit the characteristic 67 nm
banding pattern of collagen. Lying between regions of many collagen bundles and fibers,
fibroblasts often present an elongated form. Fibrocytes are essentially mature fibroblasts.
Fibrocytes can easily be recognized as such by their dense nucleus, long and thin cytoplasmic
processes and scarcity of cell organelles (Ebe & Kobayashi, 1972).
The response produced by subcutaneous implantation of demineralized crab shell in the
abdominal region of 28 day old Sprague–Dawley rats was assessed. The implantation time
spanned 28 days. Transmission electron microscopy image, from the harvested sample, figure 5-
10, displays the characteristic 67 nm banding pattern and collagen bundle. Collagenous fibers,
collagenous bundles, and fibroblasts are depicted in figures 5-10, 5-11, and 5-12. The fibers are
orientated in either cross-section or different degrees of tangential or longitudinal sections. In the
case of the longitudinal sections, the fibers exhibit their characteristic banding pattern.

172
Figure 5-12 Transmission electron microscopy image depicting collagenous fibers, collagenous
bundles, and fibroblasts. Fibers oriented in longitudinal directions exhibit the characteristic 67
nm banding pattern of collagen. Lying between regions of many collagen bundles and fibers,
fibroblasts often present an elongated form. Fibrocytes are essentially mature fibroblasts.
Fibrocytes can easily be recognized as such by their dense nucleus, long and thin cytoplasmic
processes and scarcity of cell organelles (Ebe & Kobayashi, 1972).
Fibroblast are the most common cells in all forms of fibrous connective tissue and they
become active following tissue injuries. Fibrocytes are essentially mature fibroblast that have
ceased their active protein synthesis (Ebe & Kobayashi, 1972). Fibroblasts are typically irregular
in form and present in wide individual variations. Fibroblasts are the elongated cytoplasmic

173
profiles containing a nucleus and lying between the tissue fibers (Laguens & Dumm, 1969),
(Hammersen, 1976). Fibroblasts are attracted by macrophages.
Macrophage presence represent the hallmark of the inflammation process. A number of
chemical substances are involved in the initiation and control of inflammation. These chemicals
work in concert. Remodeling consists of the deposition of the matrix and its subsequent changes
over time. It occurs throughout the entire wound repair process. One of the characteristics of
wound remodeling is the change of extracellular matrix composition. The regulation of collagen
synthesis is controlled by several growth factors, including TGF-β and fibroblast growth factor
(Li, Chen, & Kirsner, 2007). In cell culture, osteoblasts are nearly indistinguishable from
fibroblasts. All the genes expressed in fibroblasts are also expressed in osteoblasts, and,
conversely, only two osteoblast- specific transcripts have been identified: one encoding Cbfa1, a
transcription factor, and the other encoding Osteocalcin, a secreted molecule that inhibits
osteoblast function. Thus, genetically, the osteoblast can be viewed as a sophisticated fibroblast
(Ducy, Schinke, & Karsenty, 2000).
Fibroblasts, which are nonimmunogenic, easily expandable, and readily available through a
minimally invasive harvesting procedure, have been under investigation as a potential source of
cells for bone tissue engineering applications because native osteoblasts are typically difficult to
isolate and expand in vitro (Hee & Nicoll, 2006). Rutherford et al demonstrated that cultured
strains of gingival or dermal fibroblasts secrete biologically active BMP-7 in vitro, and when
absorbed to porous, preformed sponges and implanted in vivo, induce ectopic and orthotopic
bone formation. Both the implanted donor and host cells participate in bone formation suggesting

174
that the BMP-transduced fibroblasts in contact with host tissue converted to osteoblasts in vivo
(Rutherford, et al., 2002).
A multipotent progenitor cell is postulated to give rise to fibroblasts and osteoblasts.
Hormonal, cytokine, and metabolic cues influence the stromal cell progenitor’s final
differentiation pathway (Wu, et al., 2000). Osteoprogenitor cells can develop into osteoblasts or
fibroblasts depending on various factors. Several members of the fibroblast growth factor (FGF)
family have an important role in the development of skeletal tissues. FGF signaling pathways
also have important roles in bone and contribute to increased osteoblast differentiation (Valta, et
al., 2006).
In this investigation, selected area electron diffraction and high spatial resolution electron
probe X-ray microanalysis have been used to characterize chemical and structural features of
possible calcium phosphate mineral phases. Selected area electron diffraction is among the most
powerful techniques for studying the nano-, atomic- and molecular level structure of pure
organic and inorganic substances, minerals, polymers, mixtures and materials. It can be used
effectively to investigate poorly ordered or disordered materials, such as most biological tissues
and composite materials that resist detailed analysis by conventional chemical and biochemical
techniques (Neary, et al., 2010). Figure 5-13 depicts regions in which selected area diffraction
(SAD) were performed. TEM can provide information to help identify material characteristics in
terms of chemistry and crystallography as well as morphology and size distributions (Bang,
Trillo, & Murr, 2003) of minerals present. Biominerals and biomineralization confer beneficial
attributes on biological forms by: (i) physical, or macrocontributions, the production of skeletal

175
structures that provide integrity and specificity on the organism, and (ii) chemical, or micro- or
sub-microcontributions that provide a personal storage system from which ions are mobilized,
used, or sequestered to assist with the distribution of nutrients that regulate growth. Availability
of stored nutrient ions is crucial to skeletal and tissue repair activities (Skinner & Jahren,
Biomineralization, 2003).
Levy showed in 1984 that bone mineral was a calcium phosphate carbonate, and DeJong,
demonstrated in 1926 that bone mineral gave an Xray diffraction (XRD) pattern similar to that of
poorly crystalline apatite (Glimcher, Bonar, Grynpas, Landis, & Roufosse, 1981). Most
investigators now agree that the principal mineral constituent of mature bone has a structure
closely resembling that of poorly crystalline hydroxyapatite (PCHA). There is also general
agreement that the crystallinity of bone mineral increases with the age of the bone mineral and is
subject to variability and heterogeneity (Glimcher, Bonar, Grynpas, Landis, & Roufosse, 1981).
The apatite family of minerals crystallizes into hexagonal rhombic prisms and has unit cell
dimensions a = 0.9432 nm and c = 0.6881 nm (Park & Lakes, 2007).
In vertebrate hard tissue, poorly crystalline hydroxyapatite-like calcium phosphate mineral
predominates (Neary, et al., 2010). Bone mineral contains approximately 5 percent of carbon
dioxide by weight. Carbon dioxide is therefore a major constituent, and its state in the solid has
an important bearing on the foregoing discussions of the nature of the crystalline lattice. No
study has presented proof that the carbon dioxide in bone occurs as carbonate, although this has
been almost universally assumed (Neuman & Neuman, 1953).

176
Figure 5-13 Micrograph depicting regions of selected area diffraction. This image contains
collagenous fibers, fibroblasts.
TEM using a field emission gun (FEG) and HRTEM was used to study the crystal
morphology and the structure on the atomic level and data was compared to published x-ray
diffraction data and patterns obtained from the American Mineralogist Crystal Structure
Database. Figure 5-13 displays the regions in which SAD was performed. Indexing diffraction
pattern was accomplished by labeling diffraction rings and spots with appropriate (hkl)
usinginterplanar spacing (d-spacing). The diffraction patterns of figure 5-14 display diffuse

177
Figure 5-14 (a) Selected area diffraction (SAD) coincides with 0021 SAD region from figure
13. (b) ) Selected area diffraction (SAD) coincides with 0023 SAD region from figure 13. (c)
Selected area diffraction (SAD) coincides with 0025 SAD region from figure 13. (d) Selected
area diffraction (SAD) coincides with 0028 SAD region from figure 13. All of these SAD images
display broad or diffuse rings which indicate amorphous-like phase.
rings. This is indicative of amorphous solids. Amorphous solids have a random orientation of
atoms (Neary, et al., 2010).
Amorphous calcium phosphate (ACP) is believed to be the initial solid phase that precipitates
from a highly supersaturated calcium phosphate solution, and can convert readily to stable

178
crystalline phases such as octacalcium phosphate or apatitic products. Its morphological form,
structural model and X-ray diffraction patterns are typical for noncrystalline substances with
short-range periodic regularity. In X-ray diffraction, it was shown to have broad and diffuse
peaks. This pattern is typical for substances that lack long range periodic regularity. Amorphous
phases have short-range order that varies between organisms and is presumably under molecular
control (Zhao, Liu, Sun, & Zhang, 2011).
For 0021 SAD, two d spacing values (d121= 0.2933 nm, d222=0.1956 nm) were calculated
from measured diameter. The diffraction pattern of figure 5-14 displays diffuse rings. This is
indicative of amorphous solids. For 0023 SAD, two d spacing values (d121= 0.2933 nm,
d222=0.1956 nm) were calculated from measured diameter. For 0025 SAD, two d spacing values
(d121= 0.2933 nm, d222=0.1956 nm) were calculated from measured diameter. For 0028 SAD,
two d spacing values (d121= 0.2933 nm, d310=0.2200 nm) were calculated from measured
diameter. The values were consistent with those of Carbonate-hydroxylapatite obtained from the
American Mineralogist Crystal Structure Database and found to be consistent (Fleet, Liu, &
King, 2004). The diffraction patterns of figure 5-14 all display broad, diffuse rings. This is
indicative of amorphous solids that have random orientation of particles (Neary, et al., 2010) and
short range order (SRO).
In an effort to locate regions of potential mineral formation, dark sections adjacent to the
collagen bundles were evaluated. Figure 5-15 depicts a region that appears to harbor material
with crystalline characteristics. To assess its crystalline nature, SAD was performed. The SAD
pattern, figure 5-16, displayed diffuse rings. This is typically indicative of amorphous solids.

179
Figure 5-15 (a) Transmission electron microscopy image of dark region adjacent to
collagenase matrix and (b) its SAD pattern which shows broad diffuse rings that are indicative
of a amorphous solid.

180
Figure 5-16 SAD pattern of dark region adjacent to collagen like matrix depicted in figure 5-15
which shows broad diffuse rings that are indicative of a amorphous solid.
However, nanosize particles can contribute to misleading SAD results. The particles are
definitely in the nanosized range. Is it amorphous, too?

181
Size of the crystalline domains can lead to peak broadening (Weidenthaler, 2011). At first
glance, the sample appears to be amorphous. High resolution-transmission electron microscopy
(HRTEM) was used to provide a complementary characterization technique and a more accurate
picture. The HRTEM image, figure 5-17, disclosed lattice spacing with long range order despite
the fact the reflections observed in selected area electron diffraction pattern exhibited broad
peaks or diffuse rings (figure 5-16). The broadening of reflections and weakening of the
scattered intensity was probably due to the size effect of the nanocrystals. The HRTEM studies
confirm the crystalline character of the nanoparticles depicting long range order. Amorphous or
non-crystalline solids possess short range or local order (Termine & Posner, 1967). Suvorova
and Buffat demonstrated that the XRD method has limitations in the phase analysis of small
particles. Even selected area electron diffraction patterns taken from relatively large samples can
be interpreted erroneously due to size effects (Suvorova & Buffat, 2001).
When diffraction patterns give broad and diffuse rings, it is assessed as amorphous. However,
it remains to be seen whether the material is amorphous or nanocrystalline. The use of electron
probes of nanometer-size scale unleashes the possibility to determine the extent of crystallinity
of nanoparticles when the incident beam focuses and illuminates only a small and thin area of the
specimen (Suvorova & Buffat, 1999). When only this area contributes to the diffraction pattern,
electron nanodiffraction (END) is possible (Cowley, 2004). Figure 5-17 depicts HRTEM of
potential carbonated HAP exhibiting long range order.
Diffraction studies on the detailed structure of biological apatite have been limited by its very
small crystal size, and most of our present understanding is inferred from studies on synthetic

182
material (Fleet & Liu, 2007). Carbonate substitution can cause crystal disorder, resulting in
decrease of apatite crystallinity that is one of the reasons why biological apatites are poorly
crystallized (Wang, Zuo, Huang, Hou, & Li, 2010). Combining the information from the SAD
and HRTEM methods, it was determined that the sample was crystalline with long-range order.
Digitally processing the HRTEM micrographs would facilitate better interpretation of image
contrast.
Particle Size
Bone consists of homogeneous plate-like crystals of biological apatite of 15 – 30 nm wide and
30 – 50 nm long (Dorozhkin S. , 2012). Researchers have investigated calcium phosphate, from
precipitation experiments, with HRTEM (Rangavittal, Landa-Canovas, Gonzalez-Calbet, &
Vallet-Regı, 2000), (Suvorova & Buffat, 2001), (Li, et al., 2007), (Neira, et al., 2009),
(Nudelman, et al., 2010), (Ospina, et al., 2012). However, Biggemann et al determined that
nanosized particles of different size preserve stoichiometric HA-like crystal. In their study, they
synthesized HAP by precipitation and then observed with HRTEM and classified particles based
on size: large (between approximately 25 and 150 nm), medium (between approximately 5 and
20 nm), and small (less than 2.5 nm). The small crystals had a rectangular domain which were
approximately two times the HA unit cell parameters. The particles were observed within
amorphous regions of their sample (Biggemann, Prado da Silva, Rossi, & Ramirez, 2008). The
particles of Biggemann, figure 5-18, represent the smallest synthetic nanostructured HAP
crystals, between 2.1 and 2.3 nm, observed with HRTEM coupled with focal-series restoration. It

183
appears that our study is the first of its kind to find biologically produced HAP crystals ~2 nm in
size while investigating a harvested sample for osteogenesis.
Nanodiffraction
X-ray crystallography is a technique for determining the molecular and atomic structure crystals.
Crystalline materials cause X-ray beams to diffract in many directions (Smyth & Martin, 2000).
X-rays are diffracted in crystals when the Bragg’s law (nλ = 2d sin θ ) conditions
are satisfied (Kasai & Kakudo, 2005). Electron diffraction has become a vital part of
transmission electron microscopy. Diffraction patterns obtained in the TEM can provides
information about crystallinity, lattice parameters, symmetry, grain morphology, orientation, and
amount of phases present. Crystal structure is a prominent characteristic in determining and
controlling properties of a material (Williams & Carter, 2009).
The physical structure of solid materials can be classified as amorphous or crystalline and
depends mainly on the arrangements of the atoms, ions, or molecules. A crystal structure has
atoms or ions arranged in a pattern that repeats itself in three dimensions and it is then referred as
a crystalline solid or crystalline material. Atomic arrangements in crystalline solids can be
described by a three dimensional lattice (the unit cell) with three vectors (a, b, c) originating
from one corner of the cell. The axial lengths a, b, and c and the inter-axial angles α, β, and γ are
the lattice constants of the unit cell. Unit cells of different types can be constructed by assigning

184
Figure 5-17 HRTEM image of potential Carbonated HAP depicting the short range or local
order.

185
Figure 5-18 HRTEM image of small-size particle. Determined that nanosized particles of
different size preserve stoichiometric HA-like crystal structure (Biggemann, Prado da Silva,
Rossi, & Ramirez, 2008).
specific values for axial lengths and inter-axial angles. The Bravais lattices consist of seven
crystal systems: cubic, tetragonal, orthorhombic, rhombohedral, hexagonal, monoclinic, and
triclinic. These seven crystal systems have a total of 14 space lattices (unit cells) based on the
internal arrangements of atomic sites within the unit cells. These arrangements can be describes
as simple, body-centered, or face-centered (Kasai & Kakudo, 2005).
A family of crystallographic planes is: (1) are parallel to each other and equally spaced; (2) is
defined as a set of planes that intersect all lattice points; and (3) fully described using three
integer indices h, k, and l, which are called crystallographic or Miller indices. Miller indices
indicate that the planes belonging to the family (hkl) divide lattice vectors (unit cell edges) a, b,
and c into h, k, and l equal parts, respectively. The distance between neighboring crystallographic

186
planes is called the interplanar distance or d-spacing which is dependent on the Miller indices of
a family and the lengths of the three-unit cell edges (Pecharsky & Zavalij, 2009).
There are several different electron diffraction techniques. Techniques associated with the
TEM, such as selected area diffraction (SAD or SEAD) and convergent beam electron diffraction
(CBED), are the focus of this discussion. CBED developed by Kossel and .Möllenstedt, is the
oldest technique used in TEM. Although SAD is the classic way of relating diffraction contrast
information and provides useful information about crystal, ~0.5 µm is the area of the smallest
selectable diameter. CBED overcomes this obstacle and easily penetrates the nanotechnology
domain (< 100 nm). Over the past 40 years, several micro and naodiffraction have been
developed to overcome the spatial resolution limitations of SAD in a TEM (Williams & Carter,
2009). Electron nanodiffraction (END) (Cowley, 2004), a special form of convergent beam
electron diffraction (CBED), involves obtaining diffraction patterns with a nanometer-scale
beam. Nanometer beams are nosiy due to the picoamp current and require a field-emission gun
(FEG) or a cold FEG source to provide a coherent beam. END affords the opportunity to analyze
individual nanoparticles (Williams & Carter, 2009).
Nanodiffraction or END was pioneered by Cowley (1981), (1984), (1996). The technique of
electron micro- and nanodiffraction was extended to the study of near-amorphous materials for
the study of particles having diameters in the 0.10-0.20 nm range. In a preliminary survey, it was
assumed that atom positions were representative of the state of order of the whole sample
(Monosmith & Cowley, 1983). It was demonstrated that as an electron beam with a diameter of
0.4-0.5 nm is moved within a projected unit cell of a crystal changes can be observed in the

187
nanodiffraction pattern (Cowley, 1981). END has been applied most extensively in the
investigation of metal nanoparticles (Cowley, Janney, Gerkin, & Buseck, 2000), (Quintana,
Cowley, & Marhic, 2004) and carbon nanotubes (Qin, et al., 1997), (Cowley & Sundell, 1997).
Nanodiffraction was employed in the same region where the nanocrystals occurred that
produce the diffuse ring pattern and the result is shown in figure 5-19. This establishes that there
are small particles single crystals with no obvious defects in this sample. Most nanodiffraction
work has been concerned with finding the crystal structure, crystal defects, and sometimes
crystal shapes for nanoparticles in the size range of 1–2 nm. In each case, the regular pattern of
widely spaced strong spots comes from a HAP crystallite. The more closely spaced patterns of
weaker spots come from the HAP. The results from many nanodiffraction patterns from
randomly selected areas may be suggestive, and a great amount of statistical information would
be required for significant results. However, in this case the presence of a crystal lattice can be
used as a reference point for orientations (Cowley, 1999).
The (000), (002), (112), and (222) diffraction spots are indexed and displayed in this
nanodiffraction pattern in figure 5-19. The model proposed here of the hexagonal lattice, as an
explanation of the experimentally obtained diffraction pattern dataset.
The chronicle of ACPs began in 1955 with Robinson and Watson (Robinson & Watson,
1955) suggesting that a considerable portion of newly formed mineral in young bone was not
crystalline but instead similar to an amorphous-like precipitate. By 1960s, Posner and
collaborators (Eanes, Gillessen, & Posner, 1965), (Termine & Posner, 1967) used the data
obtained from x-ray diffraction and electron microscopy to point to the fact that bone mineral is a

188
mixture of amorphous calcium phosphate and crystalline apatite. Futhermore, Poser and Betts
proposed a possible structure for ACP, a Ca9(PO4)6 cluster known as Posner’s cluster, that is
present in crystalline HAP (Betts & Posner, 1974), (Posner & Betts, 1975).
Presently, biomineralization processes considers that every biogenic mineral has a precursor
phase that can be amorphous and/or composed of a different mineral phase. In the case bone
Figure 5-19 HRTEM nanodiffraction image of potential Carbonated HAP depicting a single
crystal.

189
apatite, it has been found that minute round particles of ∼1–2 nm in diameter are present in
conjugation with type I collagen molecule and aggregated small round particles are present in
direct association with early nanocrystal formation. A significant feature observed was that
particle crystallinity was extremely variable. Some particles were crystalline over their entire
extent, while others presented crystalline and amorphous zones of different sizes (Ospina, et al.,
2012). Combining the information from the SAD, HRTEM, and nanodiffraction methods and
supplying a more comprehensive assessment, it was determined that the sample was crystalline
with long-range order.
While hydroxyapatite forms needle-like crystals, the amorphous calcium phosphate phase is
in the form of spherical grains of diameter ~ 300 – 1000 Å. Posner’s clusters are roughly
spherical with a diameter of 9.5 Å and is speculated to be the transient precursor to ACP
formation. This Ca9(PO4)6 cluster allows the ACP to dissociate into these clusters instead of
undergoing complete ionic solvation in ACP to HAP transformation (Yin & Stott, 2003).
Indexing the nanodiffraction pattern
There are conventions for labeling, or indexing, diffraction patterns using the numbers (hkl).
One approach is termed “fingerprinting.” The idea is to match d spacings in the observed
diffraction pattern to known patterns from a standard sample, from calculation, or from a
database. The International Center for Diffraction Data, ICDD, (Fultz, 2013); the Joint
Committee for Powder Diffraction Standards, JCPD (Kasai & Kakudo, 2005); and the American
Mineralogist Crystal Structure Database are example of databases that maintain diffraction

190
patterns from hundreds of thousands of inorganic and organic materials. For each material the
data fields include the observed interplanar spacings for all observed diffraction peaks, their
relative intensities, and their hkl indexing. Software packages are also available to identify peaks
or d spacings in the experimental diffraction pattern, and then search databases to find candidate
materials. The task of indexing a diffraction pattern can be helped with information about
chemical compositions and candidate crystal structures (Fultz, 2013). Thus, diffraction patterns
can be indexed using graphical, mathematical, and analytical techniques as well as databases and
computer software or a combination of these techniques. Sometimes diffraction patterns do not
have all the peaks that they should because the sample specimen could be textured or particle
size could be too small. It should be noted that electron patterns can easily be obtained from very
minute volumes of calcium phosphate solid phases containing only a few solid phase particles
(Landis & Glimcher, 1978).
Mathematical Method for Hexagonal
Mathematical methods require some knowledge of the crystal structure and resulting lattice
parameter ratios such as c/a. This information can be determined graphically using a method
developed by Hull and Davey (Hull & Davey, 1921), (Schneer, 1957). This method allows the
determination of structure even if lattice parameters are unknown.
A diffraction pattern from a material typically contains many distinct peaks, each
corresponding to a different interplanar spacing, d. For hexagonal crystals with lattice parameter
a, the interplanar spacing, d, is denoted by:

191
1
𝑑2=
4
3(
ℎ2 + ℎ𝑘 + 𝑘2
𝑎2) +
𝑙2
𝑐2
Equation 1
Combine with Bragg’s law ( = 2dsin)
1
𝑑2=
4
3(
ℎ2 + ℎ𝑘 + 𝑘2
𝑎2) +
𝑙2
𝑐2=
4 𝑠𝑖𝑛2 𝜃
2
Equation 2
Rewrite as
𝑠𝑖𝑛2𝜃 =2
4[4
3(
ℎ2 + ℎ𝑘 + 𝑘2
𝑎2) +
𝑙2
𝑐2]
Equation 3
Or
𝑠𝑖𝑛2𝜃 =2
4𝑎2[4
3(ℎ2 + ℎ𝑘 + 𝑘2) +
𝑙2
(𝑐𝑎⁄ )2
]
Equation 4

192
The lattice parameter a and the ratio of lattice parameters c/a are constant for a given diffraction
pattern. These parameters are highlighted in red.
𝑠𝑖𝑛2𝜃 =2
4𝑎2[4
3(ℎ2 + ℎ𝑘 + 𝑘2) +
𝑙2
(𝑐𝑎⁄ )2
]
Equation 5
=2
4𝑎2[𝐴] +
2
4𝑎2[𝐵]
The pattern can now be indexed by considering the parameters [A] and [B] individually.
4
3(ℎ2 + ℎ𝑘 + 𝑘2) ……………………………………………………[A]
and
𝑙2
(𝑐𝑎⁄ )2…………………………………………………………………[B]

193
Table 5-2 Term [A] calculation for various values of hk
k
0 1 2 3
h
0 0.000 1.333 5.333 12.000
1 1.333 4.000 9.333 17.333
2 5.333 9.333 16.000 25.333
3 12.000 17.333 253.333 36.000
Starting with [A], this term only depends on the indices h and k. This value can be calculated by
modulating h and k.
Term [A] is calculated for various values of hk, Table 5-2.
Term [B] can be determined by substituting the known value of c/a ratio. This ratio can be
determined graphically by Hull-Davey method discussed earlier; mathematically; or can be an
estimate based on other experimental data. Based on other experimental data such as TEM
micrographs, Ca/P ratios, it is believed that our diffraction pattern depicts a member of the
hydroxyapatite family. For illustrative purposes, we will use c/a value of 0.728 for carbonate-
hydroxyapatite in the table below:
Term [B] calculated for carbonate-hydroxyapatite (c/a = 0.728).

194
Table 5-3 Calculation of l2 and l2/(c/a)2
l l2 l2/(c/a)2
0 0 0.000
1 1 1.885
2 4 7.540
3 9 16.965
4 16 30.160
5 25 47.126
6 36 67.861
Next, the two terms [A] and [B] are permitted by the allowed hkl values are added and ranked
din increasing order. Due to symmetry relationships, h and k are interchangeable. For a
hexagonal system, the structure factor calculation follows the rule: If h + 2k = 3n and l is odd,
then there is no XRD peak. Therefore, remove forbidden peaks and eliminate equivalent ones.
Table 5-4 Evaluation of Peaks.
Indices (hkl) l h + 2k Peak
111 ODD 3 NO
105 ODD 1 3n YES
Etc….

195
The calculated values in the table represent a partial list of the allowed (hkl) reflections and the
interplanar spacing.
Table 5-5 Allowed hkl and calculated d spacings
Allowed hkl Calculated d spacing (nm)
100 0.81811
101 0.52658
110 0.47234
200 0.40905
111 0.38941
201 0.35161
002 0.34403
102 0.31713
120 0.30921
211 0.28205
121 0.28205
112 0.27809

196
The parameters a and c can be calculated from the interplanar spacing equation. The
parameter a can be calculated where l = 0 (i.e., hk0) and similarly the parameter c can be
determined where h=k=0 (i.e., 00l) (Suryanarayana & Norton, 1998). These steps can be used to
generate calculated d spacings for both powder diffraction and nanodiffraction patterns.
Assignments can be made for observed d spacing values for powder diffraction patterns. For
nanodiffraction patterns, indexing begins with the identifying the brightest spot in the center of
the diffraction pattern. This represents the transmitted beam, or the (000) diffraction. Next two
independent diffraction spots nearest to the (000) spot require indexing. Once these
two vectors are determined, linear combinations of them can be used to obtain the positions and
indices of all the other diffraction spots. Much of the work in indexing nanodiffraction patterns
include measuring angles and distances between diffraction spots and then comparing these
measurements to geometrical calculations of angles and distances. The structure rule applies and
certain diffraction spot must be eliminated (Fultz, 2013). Computer programs are useful to help
elucidate the combinations of interplanary spacings.
In nanodiffraction, there are different approaches to indexing patterns. All carry frustrations.
One approach is referred to as “by hand.” This involves guessing the zone axis and its diffraction
pattern. This method is usually perform on diffraction patterns with obvious symmetry, such as a
square or hexagonal array of spots for a cubic crystal. The camera equation, r d = λ L, can be
used to obtain the interplanar distance, d, if the camera constant is known. Once the diffraction
pattern has been identified, all linear combinations of the reciprocal lattice vectors must give

197
Figure 5-20 TEM image of possible polycrystalline HAP with interplanar or lattice spacings
indices of all other spots in the diffraction pattern and the zone axis should be consistent with a
right-handed coordinate system (Fultz, 2013).
Another approach involves indexing the spots first and the zone axis can be obtained at the
end of the procedure. Ratios from the allowed hkl are sought and listed. These ratios are equal to
the ratios of the spot separations. Search for diffractions whose spot spacings are in the ratio of
the measured distances. Choose specific vector families that provide the correct angles in the
Interplanar spacing = 0.95 nm

198
Figure 5-21 SAD pattern of a polycrystalline HAP
diffraction pattern. Computer programs can provide ratio data and simplify the task of indexing.
Fultz presents a more detailed explanation (Fultz, 2013).
To index our nanofiffraction pattern, yet another approach was used. In this case, the scale bar
was used to measure and calculate interplanar spacings. This approach involved measuring
angles and distances between diffraction spots, followed by, matching the observed interspacing
with published data. The d spacings observed were: d002 = 0.3451 nm; d112 = 0.2711 nm; and d222

199
= 0.1921 nm. The camera equation, r d = λ L could be used to determine d when the camera
length and λ are known. Another method that could have been utilized involve using ratios of r’s.
The selection of a basic parallelogram can be made and distance between the diffraction spots, r,
measured. A list of all the allowed reflections along their relative spacing can be generated. The
ratios can be compared to find the appropriate hkl. After indexing a couple diffraction spots, the
diffraction pattern can be completely labeled by vector addition. Computer program can help in
various ways to ease the tasks.
SAD was taken in another region in close proximity to the nanocrystals. Characteristic
interplanar or lattice spacing were displayed, figure 5-20. The crystallites are plate shaped nano -
sized particles with lengths and widths of approximately 40 x 20 nm. The measures lattice
spacing was 0.95 nm. This is representative of resolution of the 100 planes of the hydroxyapatite
crystal lattice. This is in line with previous studies (Selvig, 1970), (Suvorova & Buffat, 2001).
The selected area diffraction is more distinct, indicating either a higher degree of crystallinity,
an increase in particle size, or both. Crystalline solids are arranged in fixed geometric patterns or
lattices. They have an ordered arrangement of units maximizing the space they occupy, and are
practically incompressible. (Neary, et al., 2010). Bone mineral has been shown to contain both
an amorphous and a crystalline solid (Termine & Posner, 1967), (Eanes, Termine, & Posner,
1967).
The diffraction pattern of figure 5-21 displays speckled rings made up of discrete spots. This
is indicative of crystalline solids with a continuum of states from nanocrystalline to poly-
crystalline. This suggest that there are nanosized crystals of different sizes represented in this

200
SAD pattern. In essence the smaller HAP nanocrytsals aggregated to form larger HAP crystals.
This implies that the process of HAP formation in this case is based on the mesocrystal model.
This nanocrystal mediated crystallization involves nucleation events of nanocrystals which
engage self-organization to form an ordered HAP agglomerated crystal superstructure. In
classical models, crystalline materials were classified into single crystal and polycrystal, but
mesocrystals introduce a concept of intermediate states --- between single crystal and polycrystal
(Oaki & Imai, 2013). Nanocrystals 20-50 nm in size were observed. The nanocrystal specimen
can be classified in accordance with crystal sizes. The crystals of ~ 2nm gave two diffuse rings in
SAD patterns which look like ‘amorphous’ patterns but produced a nanodiffraction pattern that
indicated a single crystal. The larger nanocrystals with width ~ 20 nm gave spotted SAD
patterns. Nonetheless, all the nanocrystals were determined to be crystalline whatever the particle
size (Suvorova & Buffat, 1999). The nanodiffraction technique indicate that the nanocrystals can
be recognized as a single crystal and are oriented in the same crystallographic directions. The
nanocrystals and biological macromolecules can be regarded as the nanoscale bricks and mortar,
respectively (Oaki & Imai, 2013). This brick and mortar building describes the behavior of the
inorganic nano-sized crystals in the mesocrystal model. The nanocrystals aggregate into ordered
solid phases via oriented attachment to control the reactivity of nanophase materials in nature
(Dorozhkin S. , 2012). These series of SAD, HRTEM, and nanodiffraction results provide
evidence of growth of our HAP crystal via the mesocrystal model.
For SAD, numerous d spacings were calculated from measured diameter shown in Table 5-6.
The values were more consistent with those of Carbonate-hydroxylapatite obtained from the

201
Table 5-6 d spacing values calculated for crystalline harvested sample. a (Fleet, Liu, & King,
2004). b (Wilson, Elliot, & Dowker, 1999)
Carbonate
Hydroxyapatitea
Hydroxyapatiteb Harvested Specimen
hkl dnm dnm dnm
200 0.40906 0.40738 0.40714
111 0.38941 0.38847 0.38000
002 0.34403 0.34444 0.33529
102 0.31713 0.31725 0.31667
211 0.28205 0.28114 0.28500
300 0.27271 0.27159 0.27143
202 0.26329 0.26302 0.26430
301 0.25352 0.25266 0.25333
130 0.22690 0.22598 0.22800
131 0.21549 0.21472 0.21923
222 0.19471 0.19424 0.19000
213 0.18421 0.18408 0.18387
004 0.17201 0.17222 0.17273
151 0.14370 0.14200

202
American Mineralogist Crystal Structure Database and found to be consistent (Fleet, Liu, &
King, 2004).
A simple two-phase system in which one phase gradually changes to, or is replaced by,
another can explain the diffuse and poorly defined selected area diffraction pattern that is
obtained from bone mineral (Glimcher, Bonar, Grynpas, Landis, & Roufosse, 1981).
The change from a poorly crystalline hydroxyapatite diffraction pattern to a more highly
crystalline one does not occur abruptly and varies only slightly at different locations within a
single section (Landis & Glimcher, Electron Diffraction and Electron Probe Microanalysis of the
Mineral Phase of Bone Tissue Prepared by Anhydrous Techniques, 1978).
A semiquantitative nondispersive electron probe x-ray microanalysis was used to examine the
region of the sample that displayed lattice spacing of 0.95 nm. This revealed an average
calcium:phosphorus (Ca:P) ratio of 1.81 ± 0.37, figures 5-22 and 5-23. Bone-apatite is
characterized by calcium, phosphate and hydroxyl deficiency and Ca:P ratios reported between
1.37–1.87 (Hing, 2004). While many investigators of calcification agree that mineralization
originates within matrix vesicles (Anderson, 1984), there is some disagreement about the exact
mechanisms of the process. It seems likely that both cellular and physiochemical factors are
involved. Generally amorphous calcium phosphates with a Ca:P ratio varying between 1.44 and
1.55 are believed to be deposited under the control of osteoblasts (Hing, 2004).
In a study conducted on human rib bones, the median reference value for the Ca/P was 2.19.
The individual variation for the Ca/P ratio in rib bone from healthy humans was lower than for

203
Element Weight% Atomic%
O K 47.43 66.88
P K 21.31 15.52
Ca K 31.26 17.60
Totals 100.00
Figure 5-22 Elemental Analysis with Weight and Atomic Percentages of oxygen, phosphorus
and calcium.

204
Element Weight% Atomic%
O K 37.88 58.28
P K 19.75 15.70
Ca K 42.37 26.02
Totals 100.00
Figure 5-23 Elemental Analysis with Weight and Atomic Percentages of oxygen, phosphorus and
calcium.
Ca and P values taken separately; therefore, Ca/P ratio provides greater reliability (Tzaphlidou &
Zaichick, 2003).
This preliminary investigation has demonstrated the potential of characterizing crystals in
the nanometer scale, and identifying the components via combined analysis of TEM, HRTEM,
SAD, and EDS. With diffraction pattern of broad and diffuse rings and speckled rings made up

205
of discrete spots, and d spacing values, and EDS findings, this sample taken from the
implantation of blue claw crab in rats is surmised to contain mineralized collagen matrix. This
sample has regions with a variety of crystalline states or variations in crystallinity. The crystal
structure of apatite allows the incorporation of a wide variety of cations and ions beyond the
essential calcium and phosphate needed for mineral and apatitic bone formation (Skinner &
Jahren, 2003).
Amorphous calcium phosphate was first described by Aaron S. Posner (Posner, Perloff, &
Diorio, 1958). Leng obtained a similar pattern from a HA single crystal in transmission electron
microscopy (TEM) when examining a sample for heterotopic osteogenesis of calcium phosphate
(Leng & Qu, 2002). Posner’s structural studies led to the highly publicized debate over the
nature of the carbonate found in bone and ignited the story of amorphous calcium phosphate.
Termine and Eanes also identified amorphous calcium phosphate (Eanes, Gillessen, & Posner,
1965). Posner and Termine quantified how much ACP was in bone and also determined how
ACP could be stabilized (Termine & S, 1967). Blumenthal, Betts and Lehninger identified ACP
as the mineral in the hepatopancreas of the blue crab (Betts, Blumenthal, Posner, Becker, &
Lehninger, 1975). Boskey studied the mechanism of hard tissue and became convinced that ACP
was not a mandatory precursor to apatite (Boskey A. L., 1997). Lowenstam asserted that in many
non-vertebrates, the initial mineral (whether phosphate or carbonate) is amorphous (Lowenstam
H. A., 1981), and that these amorphous minerals transform to crystalline phases as the animals
mature (Lowenstam & Weiner, 1989).

206
Figure 5-24 Schematic diagrams representing the HAP unit cell (Menéndez-Proupin, et al.,
2011)
Arzate et. al. studied the cementoblastoma cell line by transmission electron, high resolution,
scanning, and atomic force microscopy. Cemtoblasts are thought to give rise to progenitors of
each cell type that comprise the periodontium, such as osteoblasts and periodontal ligament

207
fibroblasts. This study revealed that the diffraction patterns obtained from the intracellular
material and human cellular cementum were similar, with D-spacings of 3.36 and 2.8, consistent
with those of hydroxyapatite (3.440 and 2.814). To reveal the formed mineral phase, selected
area diffraction and nanodiffraction techniques were used and because of the nanosize of the
formed crystals, patterns of concentric double rings emerged (Arzate, et al., 2000). Another study
by Leng and Qu examined a single crystal of hydroxyapatite from an implanted porous
hydroxyapatite/tricalcium phosphate from dorsal pouches of dogs after 2-months. The diffraction
pattern lead them to identify the particle as an HA single crystal after comparing the diffraction
pattern with the HA lattice structure (Leng & Qu, 2002). To date, this research is the first to
describe the use of nanodiffraction to confirm mineral hydroxyapatite formation from an implant
of crab shell in a Wistar rat. The goal of this paper is not to give a detailed review of the growth
of HAP mineral crystal or the biomineralization process. This is a report of our investigation
with HRTEM and electron nanodiffraction of hydroxyapatite formation from an implant of crab
shell in a Wistar rat.
Mineralization of a tissue refers to the deposition of calcium phosphate mineral phase. This
mineral, often called ‘apatite-like, closely resembles bone mineral in composition and structure
(Ito & Onuma, 2003). The chemical equation for the formation of hydroxyapatite is 10 Ca2+ + 6
PO43- + 2 OH Ca10(PO4)6(OH)2. Stoichiometrically this reaction could be simplified by
dividing by two, it is written to emphasize that Ca10(PO4)6(OH)2 is the formula for a single unit
cell of HAP (Simmer & Fincham, 1995). Figure 5-24 depicts different ways of representing the
HAP unit cell. The primary difference between bone mineral and hydroxyapatite manifest in

208
crystallinity and impurity content (Ito & Onuma, 2003). Perfect and pure Hap crystals are
extremely difficult to form in both synthetic processes as well as in vivo in biological systems.
Biological HAP contains a variety of vacancies, interstitials, and trace elements (Matsunaga,
2010) and is subjected to common ion substitutions, PO43- by CO3
2- and OH- by CO32-. These
two are the most common ion substitutions and are called B-type and A-type, respectively. B-
type substitution, the preferential carbonate substitution found in the bone of a variety of species,
is believed to improve bioactivity and greater strength (Chappell & Bristowe, 2007), (Ibrahim,
Mostafa, & Korowash, 2011). B-type carbonate substitution, where carbonate substitutes for the
phosphate ion, is of particular importance because biological apatites contain 4–6% carbonate by
weight and the connection between carbonate concentration and physical properties (Leventouri,
2006). Thus, biological apatites are often described as a calcium deficient HAP modified with
different ion substitutions (Mostafa & Brown, 2007). Hydroxyapatite is among the most stable
calcium phosphate solid phase and can precipitate from a solution having a relatively low
concentration of calcium and phosphate ions (Simmer & Fincham, 1995).
Bone apatite crystals are plate-like shaped with average length, width and thickness of
50×25×4 nm, although a great range of values can be found in the literature (Rossi, et al., 2012).
The Hap crystal structure is comprised of two inequivalent Ca2+ sites: (1) Ca cations at site I
forms triangles coordinated by nine oxygens belonging to six PO43-
that displays a columnar
arrangement; (2) Ca cations at site II are coordinated by six oxygen atoms belonging to five
PO43- anions and one OH- anion (Li, et al., 2007), figures 5-24, 5-25, and 5-26. Substitutions can

209
result in changes in the crystal lattice dimension, with a decrease in the a-axis and an increase in
the c-axis (Hench & Best, 2013).
Having similar general atomic arrangements, HAp has two kinds of polymorphism, a more
complex monoclinic (P21/b), and hexagonal (P63/m). The hexagonal form is encountered most
frequently and is involved in bone formation because it allows for much easier exchange of OH-
groups with other anions. However, the monoclinic form is more stable, from the thermodynamic
prospective (Corno, Busco, Civalleri, & Ugliengo, 2006). The hexagonal primitive unit cell of
HAP has a total 44 atoms and the lattice parameters of a=b=0.94081 nm, c=0.68887 nm, with a =
β = 90 and γ = 120 (de Leeuw, 2010). To form the OH group along the c axis, there are two
inequivalent Ca sites (Ca-1 and Ca-2), one P site, four different oxygen sites (O-1 to O-4), and
one H site bonded to O-4. This structure has four possible sites for the OH groups along the c
axis and may undergo disorder in the column (Matsunaga, 2010). The monoclinic unit cell is
obtained from the hexagonal one by doubling the b lattice parameter with an alternating
arrangement of the anion chain (Calderin & Stott, 2003). Singh and Jonnalagadda studied metal
substituted hydoxy apatites’ ability to maintain their structure and temperature at temperatures
reaching 900°C. Using Brunauer–Emmett–Teller (BET), they determined that HAP had a surface
area of 72.5 m2/g and a pore volume of 0.2385 m2/g. Additionally, the degree of crystallinity, Xc,
was calculated by Scherer’s equation and was determined to be 0.795 (Singh & Jonnalagadda,
2013).
Harries, Hukins and Hasnain (1986) analyzed a spectrum recorded above the calcium edge of
hydroxyapatite using Extended X-ray Absorption Fine Structure (EXAFS) to gain structural

210
information on the local atomic environment surrounding the calcium ion out to a distance of 0.6
nm. EXAFS spectra have been used to provide quantitative information on relatively simple
crystal structures, distances between atoms, and the near neighbors of absorbing atoms in more
complex systems. Crystallographic data was essential to interpret the EXAFS of hydroxyapatite
providing the initial EXAFS parameters used in simulating the spectrum. In addition, phase
shifts calculated using the ab initio method and local atomic potentials were constructed using
the muffin-tin approximation, choosing a local atomic arrangement consistent with the known
Figure 5-25 Schematic depiction of one hexagonal building unit of HAP (Li, et al., 2007).

211
Figure 5-26 Black lines connect Ca(I) columns in hexagonal networks. Cyan and magenta
triangles connect staggered Ca(II) atoms (Xia, Lindahl, Lausmaa, & Engqvist, 2011), (Boanini,
Gazzano, & A, 2010).
Figure 5-27 The HAP unit cell has symmetry and order. Table of average distances of atom shell
from calcium atom via x-ray and EXAFS (Harries, Huskins, & Hasnain, 1986).
Hydroxyapatite
(HAP)
Chemical Formula Ca10(PO4)6(OH)2
Ideal Ca/P ratio 10/6
Density 3.2 g/cm3
Atomic Mass 1004.6 g/mol
Mass of unit cell ~1.67 x 10-21 g
Volume of unit cell ~5.21 x 10-22 cm3
Atom Type Averaged
Coordination
Number
Average
distance of atom
shell from
calcium atom
(nm) (x-ray)
Average
distance of atom
shell from
calcium atom
(nm) (EXAFS)
Oxygen 5.4 0.239 0.237
Oxygen 1.0 0.251 0.256
Oxygen 2.0 0.279 ---
Phosphorus 2.4 0.320 0.311
Calcium 0.8 0.344 0.346
Phosphorus 3.0 0.360 ---
Oxygen 6.6 0.399 ---
Calcium 8.4 0.405 0.395
Oxygen 4.8 0.445 0.445
Oxygen 6.5 0.481 0.469
Calcium 1.5 0.545 ---
Calcium 2.0 0.584 0.574
Calcium 6.0 0.618 0.622

212
crystal structure of hydroxyapatite. The inter-atomic distance and the Debye-Waller terms were
refined to obtain the best agreement with experimental data. Their analysis of the average
calcium environment for hydroxyapatite as determined by their x-ray crystallography and
EXAFS is provided in figure 5-27. With image processing to reveal a clearer picture, the
distances between the atoms in figure 5-17 can be compared with the data from figure 5-27.
Possible mechanism
Bone formation induced by osteoinductive biomaterials occur in several steps: (1) attachment
of mesenchymal cells on material surface; (2) proliferation and differentiation of mesenchymal
cells; (3) bone matrix formation by induced osteogenic cells; (4) mineralization of bone matrix;
an (5) bone remodeling to form mature bone. The time for different steps to happen varies with
the osteoinductive potentials of the biomaterial. Within two weeks of implantation, cells
typically attach to biomaterial; within three weeks, cells proliferate and differentiate; within 30
days, bone matrix form; within 45 days, mineralized bone forms; and within 60 days,
remodeling. The processes of osteoinduction will never happen if a non-osteoinductive
biomaterial is implanted in tissue (Reis & Weiner, 2004).
Similar steps are always followed for bone formation induced by different biomaterials
despite the varying starting times. In skeletal development, bone naturally occurs either by direct
replacement of pre-existing connective tissues (intramembranous ossification) or by replacement
of pre-existing cartilage (endochondral ossification). Cartilage formation has not been found in

213
Figure 5-28 Diagram for wound healing response to crushed crab carapace (Falanga, 2005),
(Salgado, et al., 2011).
Differentiation
Chemotaxis,
Fibroblastic to
Osteoblast phenotype
Resolution Osteogenic cells,
Collagen matrix release,
Mineralization
Monocytes, Neutrophils
migrate to implant site
after injury
Monocytes become
macrophages and attach to
crab carapace
Fibroblastic cells of the
osteoprogenitor cell line
migrate to site
Signal for phenotype
switch: fibroblast to
osteoblast phenotype
Cells differentiate to
Osteogenic-like cells
Cytokine and other
metabolic cues influence
cells pathways
Osteogenic cells release
collagen matrix and begin
to mineralize.

214
bone formation induced by osteoinductive biomaterials; therefore it is supposed that bone
formation induced by osteoinductive materials resembles the intramembranous ossification
development. Starting directly as bone, secondary bone formation occurs when bone formation
takes place at the resorbed bone bed during remodeling. In secondary bone formation, osteogenic
precursor cells attach on the resorbed bone bed, proliferate, differentiate, and form new bone.
Bone formation induced by osteoinductive biomaterials has sequences similar to secondary
bone formation. When bone formation induced by biomaterials begins, osteogenic precursor
cells (mesenchymal cells) attach on osteoinductive biomaterials surface, proliferate and
differentiate to osteogenic cells which then form bone. Therefore, it is more likely that bone
formation induced by osteoinductive biomaterials is a secondary bone formation. Bone repair
happens faster with implants with a high osteoinductive potential. Osteoinductive biomaterials
provide a good environment for osteogenic cells to form bone and induce non-osteogenic cells to
differentiate into osteogenic cells that then form bone (Reis & Weiner, 2004).
In the case of Calinectes Sapides (Blue Claw) crab shell, we postulate that monocytes, closely
following the neutrophils, migrate to the implant. Once in the tissue, the monocytes become
macrophages and attach to the crab shell implant. These attached macrophages release a wide
range of biochemical factors that can mediate the activity of many other cells, including
fibroblasts, osteoblast, and osteoclasts (Ziats, Miller, & Anderson, 1988). It has been shown that
macrophages express osteopontin, a protein needed in bone formation (Freeman & Otto, 2005).
Macrophages influence the migration of multipotent progenitor fibroblastic cells that are of
osteoprogenitor precursor cell line or osteoblast phenotype. These multipotent progenitor

215
fibroblastic cells give rise to osteoblasts (Wu, et al., 2000). Several members of the fibroblast
growth factor (FGF) family have an important role in the development of skeletal tissues and
increased osteoblast differentiation (Yun, et al., 2010) (Valta, et al., 2006). These fibroblastic
cells in contact with the crushed crab cuticle proliferate and differentiate to osteogenic cells.
Hormonal, cytokine, and metabolic cues influence the multipotent progenitor cell’s final
differentiation pathway (Wu, et al., 2000). The osteogenic cells release collagen matrix and
begin to mineralize. The macrophage, fibroblastic cells, and the crab cuticle signal each other
releasing biochemical factors that create a microenvironment conducive to bone formation.
Termine and Posner suggested that amorphous calcium phosphate is the first mineral deposited
during the calcification process and that the amorphous bone mineral fraction can act as a
metabolically active, metastable precursor of crystalline bone apatite (Termine & Posner,
Amorphous / Crystallinne Interrelationship in Bone Mineral, 1967). The beginning stages of
matrix mineralization were evidenced in this study.
Other factors may be involved in bone formation induced by biomaterials, including growth
factors other than BMP’s, the surface energy (zeta-potentials), low oxygen, asymmetric cell
division which may occur on a rough surface and cause differentiate into osteogenic cells (Reis
& Weiner, 2004).
Additionally, the importance of the inflammation phase is noteworthy. Inflammation is a
crucial phase in both normal wound healing and bone repair and remodeling. A growing body of
evidence has emerged that suggest that a complex balance exists between regenerative and
damaging inflammatory processes and tissue healing involves overlapping phases of

216
inflammation, renewal, and remodeling (Mountziaris, Spicer, Kasper, & Mikos, 2011).
Inflammation is necessary early in the regenerative process to initiate the repair cascade. If
healing occurs normally, then the inflammatory phase is resolved promptly and tissue
regeneration can occur (Thomas & Puleo, 2011). In reference to bone, the study of the
relationship between inflammatory mediators and bone has been termed “osteoimmunology”
(Graves, 2008).
There are several pathways through which tissue can heal, yet the unique attribute of bone
repair is that it occurs without the development of a fibrous scar. Inflammation, repair, and
regeneration are well- orchestrated, carefully choreographed processes with many interrelated
anatomical, biomechanical and biochemical processes (Marsell & Einhorn, 2011).
We believe that the phases of healing regarding the implanted crushed crab shell resembles a
cross between normal wound healing and bone tissue repair. This process, with overlapping
phases, begins with the inflammation phase after injury, followed by proliferation phase, then a
differentiation phase based on the modification of fibroblast behavior in the presence of the crab
carapace, and finally resolution phase involving the deposition of collagen matrix followed by
mineralization of newly formed fibers. The cascading events are depicted in figure 5-28.
Concluding Remarks
Biology and technology/engineering use very different approaches to the solution of similar
problems or functions of design and integration. However, many of nature’s systems use the
same exquisite and superior design principles. As hard tissue systems are studied, the similarities

217
between systems such as bone and crab cuticle become obviously clear. Exploitation of these
similarities, biorecyclying of crab cuticle takes advantage of the economical and efficient ways
nature achieves its objective in terms of energy, cost, and materials. In addition, most of the
design prototyping and fabrication has been done.
The hypothesis that crab shell may possess the inherent ability to induce bone formation in is
based on unique similarities between the hard tissue bone and crab shell integument. The results
from TEM, HRTEM, XRD, SAD, END, and EDS, in this study indicate that crushed shell from
C. sapidus exhibits some unique interactions in vivo using Sprague–Dawley rats. These results
further suggest that crab shell may be suitable for use in developing functionally advanced bone
implants and this material warrants further study to ascertain its potential osteoinductive
properties.
Thus a few points are highlighted:
1. Degree of crystallinity changes as bone matrix matures
2. Degree of crystallinity can be masked by the size of the crystal
3. Degree of crystallinity varies with location
4. Our study suggests that crab shell cuticle is osteoinductive.
5. Our study indicates that crab cuticle may be an effective material for stimulating bone
growth.
6. Our findings lays the groundwork for further studies.
Assessing crab shell osteoinductive attributes and providing the first general biological
performance profile about the suitability of crab cuticle as a bone biomaterial is a novel

218
contribution to the fields of biomimetics, biological material science, orthopedic medicine,
microbiology, and bone tissue engineering. Furthermore, this process will enhance treatments
options for repair, remodeling, and reestablishing health of bone.

219
Chapter 6 Final Remarks
Biomimetics involves examining the manner in which biological organisms solve problems to
observe how that compares with solutions of the same or similar problems within technical
environment. Using biology as a living database of the solution of technical problems by
organism, TRIZ is a method that allows interpretation or translation from the biological to
technical world by providing focused ideas for the solutions of recognized problems. Biology
can be analyzed and described by TRIZ to complete the concept of biomimetics assisting in the
transfer of nature’s technology and its implementation in the technical world. By comparing the
functionality of biological a man-made polymers, proteins, and polysaccharides, man-made
polymers are not as versatile or responsive as the two biological polymers, protein and
polysaccharide. When considering biological hard tissue, calcium derivatives, carbonates and
phosphates are predominant. A limited range of chemicals suffices for nearly all biological hard
materials (Vincent J. F., Biomimetic materials, 2008). Although biological/engineering TRIZ in
its infancy, it can be used to feature that crab cuticle and bone tissue are the solution to similar
problems. Biomimetics and TRIZ can illuminate and build on the similarities between bone and
crab cuticle sanctioning biology’s ability to transform engineering.
Bone tissue and crab cuticle are nanocomposite that exhibit hierarchical structuring and
contain two distinct constituent phases. Bone tissue has an organic and inorganic phase, collagen
I and calcium phosphate, respectively. Crab cuticle also has an organic and inorganic phase, α-
chitin and calcium carbonate, respectively. Bone and crab cuticle are the sum of their parts.

220
When inspecting the organic constituents (collagen and chitin) of these hard tissues, many
uncanny observations emerge. Collagen and chitin have a similar and engaging life story. They
do not act alone; collagen is a triple helix embedded in an inorganic matrix and chitin is
embedded in a protein helix. Compared to typical structures of their kind, both collagen and
chitin are unusual. Although chitin is a polysaccharide and collagen is a protein, collagen and
chitin associate with proteoglycans. Collagen and chitin complex composites are hierarchically
structured and multifunctional Microfibril: Fribil Aggregation (Fiber): Fibrous Bundle:
‘Twisted Plywood”. Chitin and collagen are lyotropic liquid crystals that are both mobile, have
similar ordered geometries, can self-assemble, and have similar optical properties. The
biosynthesis and degradation of both collagen and chitin are stepwise procedures governed by
enzymatic activity. Both collagen and chitin are synthesized by sequential changes in the
cytoplasmic matrix, RER, processing in the Golgi apparatus and transport through vesicles
culminating in exocytosis across the membrane where chemical reactions occur and association
into fibril, fibers, and fibrous bundle. Collagen and chitin share properties are both
biocompatible, biodegradable, non-toxic, highly insoluble, and birefringement. Both chitin and
collagen have a wide variety of medical applications.
Bone and crab cuticle are naturally fabricated bioceramic composites that are assembled from
readily available materials, typically in aqueous media, at ambient conditions, and to net ordered
shape. The production of hard tissues involves an exquisite level of control of the nucleation and
growth of mineral. The most common mineral being phosphate and carbonate salts of calcium

221
that are used in conjunction with organic polymers such as collagen and chitin to give structural
support to bones and shells. The minerals are formed by normal cellular processes and replicate
to produce precisely organized structures in cells ranging from bacteria to osteoblasts of bone
(Mann, Molecular recognition in biomineralization, 1988).
Biomineralization is not a single process. Every organism has adapted certain strategic
principles to optimize the specific function of its hard tissue to the specific environment in which
it lives (Heuer, et al., 1992). The tools for biomineralization of crab cuticle and bone are calcium
carbonate and calcium phosphate, respectively. Biological systems are teeming with examples of
organic supramolecular assemblies (double and triple helices, multisubunit proteins, membrane
bound reaction centers, vesicles, tubules, etc.), some of which (collagen, cellulose, and chitin)
extend to microscopic dimensions in the form of hierarchical structures. This hierarchical
processing is important in the application of molecular tectonics in biomineralization: organic
architectures produced by molecular processes are used in turn as the building framework for the
synthesis of organized materials, which in turn provide the prefabricated units of complex
microstructures (Mann, Molecular tectonics in biomineralization and biomimetic materials
chmeistry, 1993).
Several theories have been proposed for the mechanism of mineralization of hard tissue. There is
at present no unified theory of the mechanism of mineralization. Theories of mineralization
include, but not limited to, biologically induced, organic matrix-mediated, mesocrystal model,
cell-mediated, matrix vesicles initiation, and a multistep phase transformation involving an
amorphous transient phase.

222
Biomineralization deals with ratios of Ca/P that change as mineralization matures.
Crystallinity varies with bone maturity (Landis & Glimcher, Electron Diffraction and Electron
Probe Microanalysis of the Mineral Phase of Bone Tissue Prepared by Anhydrous Techniques,
1978). Degree of crystallinity can be masked by the size of the crystal and varies with location.
Studies have indicated that the earliest deposits of mineral generate an amorphous solid phase
pattern while more mature deposits generate patterns of very poorly crystalline hydroxyapatite
which gradually approach those of crystalline hydroxyapatite (Landis & Glimcher, Electron
Diffraction and Electron Probe Microanalysis of the Mineral Phase of Bone Tissue Prepared by
Anhydrous Techniques, 1978).
Our study suggests that crab shell cuticle is osteoinductive and may be an effective material
for stimulating bone growth similar to demineralize bone matrix. Clinical outcome assessment
and comparative effectiveness to determine the efficacy of the tissue-engineered bone are
limited. Many of the preclinical models to test efficacy use methods that are invasive or not
readily translated to the assessment of human engineered tissues. Outcomes such as histology
and biomechanical evaluation require sacrifice of the animal and tissue harvest. Improved
noninvasive assessment tools are needed to monitor bone healing to allow effective comparisons
of the relative efficacy of different materials and composites.
The old adage that life is merely advanced aqueous organic chemistry has been superseded
for the most part by a view that embraces the interconnectivity of metal and non-metals in the
biochemical processes (Mann, Molecular tectonics in biomineralization and biomimetic
materials chmeistry, 1993). Embracing the interconnectivity of biological systems can be

223
achieved using biomimetics, TRIZ, and robust information technology tools involving
monitoring, tracking, and reporting for all aspects and levels of the biological system.
Designing biomimetics bone implants using involves transforming needs and requirements
into an integrated system design solution through concurrent consideration of all life-cycle needs
from development, testing, evaluation, tracking, reporting, and disposal. While bone tissue
engineering has capitalized on a number of breakthrough technologies, none have been able to
produce an ideal bone biomaterial. As our ability improves to collect, store, and analyze
information about all the elements of a biological system, more cues will emerge about the true
mechanisms of biological processes such as bone formation. This will undoubtedly lead to
advances in understanding and replicating the best material scientist, nature.

224
References
Abdou, E. S., Nagy, K. S., & Ellsabee, M. Z. (2008). Extraction and characterization of chitin
and chitosan from local sources. Bioresource Technology, 99, 1359-1367.
Abraham, L. C., Zuena, E., Perez-Ramirez, B., & Kaplan, D. (2008). Review: Guide to Collagen
Characterization for Biomaterial Studies. J Biomed Mater Res Part B: Appl Biomater,
264–285.
Addadi, L., Sefi, R., & Weiner, S. (2003). Taking Advantage of Disorder: Amorphous calcium
carbonate and Its Roles in Biomineralization. Advanced Materials, 15(12), 959-969.
Albrektsson, T., & Johansson, C. (2001). Osteoinduction, osteoconduction and osseointegration.
European Spine Journal, 10, S96–S101.
Allen, R. (2010). Bulletproof Feathers: How Science Uses Nature's Secrets to Design Cutting-
Edge Technology. Chicago: The University of Chicago Press.
Anderson, H. C. (1967). Electron Microscopic Studies of Induced Cartilage Development and
Calcification. Journal of Cell Biology, 35(1), 81-101.
Anderson, H. C. (1984). Mineralization by Matrix Vesicles. Scanning Electron Microscopy, 953-
964.
Anderson, H. C. (1984). Mineralization by Matrix Vesicles. Scanning Electron Microscopy, 953-
964.
Anderson, H. C., Mulhall, D., & Garimella, R. (2010). Role of extracellular membrane vesicles
in the pathogenesis of various diseases, including cancer, renal diseases, atherosclerosis,
and arthritis. Laboratory Investigation, 90, 1549-1557.
Andrade, A. L., Ferreira, J. M., & Domingues, R. Z. (2004). Zeta Potential Measurement in
Bioactive Collagen. Materials Research, 7(4), 631-634.
Aranaz, I., Harris, R., & Heras, A. (2010). Chitosan Amphiphilic Derivatives. Chemistry and
Applications. Current Organic Chemistry, 14, 308-33.
Arzate, H., Alvarez-Perez, M. A., Alvarez-Fregoso, O., Wusterhaus-Chavez, A., Reyes-Gasga,
J., & Ximenez-Fyvie, L. A. (2000). Electron Microscopy, Micro-analysis, and X-ray

225
Diffraction Characterization of the Mineral-like Tissue Deposited by Human Cementum
Tumor-derived Cells. Journal of Dental Research, 79(1), 28-34.
Atala, A., Lanza, R., Thomson, J. A., & Nerem, R. M. (2008). Principles of Regenerative
Medicine. Boston: Academic Press.
Aubin, J. E. (1998). Bone Stem Cells. Journal of Cellular Biochemistry Supplement, 30/31, 73-
82.
Austin, P. R., Brine, C. J., Castle, J. E., & Zikakis, J. P. (1981). Chitin: New Facets of Research.
Science, 212, 749-753.
Bang, J. J., Trillo, E. A., & Murr, L. E. (2003). Utilization of Selected Area Electron Diffraction
Patterns for Characterization of Air Submicron Particulate Matter Collected by a
Thermophoretic Precipitator. Journal of the Air & Waste Management Association, 53,
227-236.
Barradas, A. M., Yuan, H., van Blitterswijk, C. A., & Habibovic, P. (2011). Osteoinductive
Biomaterials: Current Knowledge of Properties, Experimental Models and Biological
Mechanisms. European Cells and Materials, 21, 407-429.
Barthelat, F. (2007). Biomimetics for next generation materials. Philosophical Transactions of
The Royal Society A, 365, 2907-2919.
Bauer, T. W., & Muschler, G. F. (2000). Bone Graft Materials: An Overview of the Basic
Science. Clinical Orthopaedics and Related Research, 371, 10-27.
Belamie, E., Mosser, G., Gobeaux, F., & Giraud-Guille, M. M. (2006). Possible transient liquid
crystal phase during the laying out of connective tissues: α-chitin and collagen as models.
JOURNAL OF PHYSICS: CONDENSED MATTER, 18, S115-S129.
Bertelsen, A. (1944). Experimental Investigations into Post-Foetal Osteogenesis. Acta
orthopaedica Scandinavica, 15, 139-144.
Betts, F., & Posner, A. S. (1974). An x-ray radial distribution study of amorphous calcium
phosphate. Mater. Res. Bull., 9, 353-360.
Betts, F., Blumenthal, N. C., Posner, A. S., Becker, G. L., & Lehninger, A. L. (1975). Atomic
Structure of Intracellular Amorphous Calcium Phosphate Deposits. Procceedings of the
National Acadeny of Sciences, 72(6), 2088-2090.

226
Biggemann, D., Prado da Silva, M. H., Rossi, A. M., & Ramirez, A. J. (2008). High-Resolution
Transmission Electron Microscopy Study of Nanostructured Hydroxyapatite. Microscopy
and Microanalysis, 14, 433-438.
Black, J. (1999). Biological Performance of Materials: Fundamentals of Biocompatibility (3rd
edition ed.). New York: Marcel Dekker, Inc.
Blackwell, J., & Weih, M. A. (1980). Structure of Chitin-Protein Complexes: Ovipositor of the
Ichneumon Fly Megarhyssa. J Mol. Biol., 137, 49-60.
Boanini, E., Gazzano, M., & A, B. (2010). Ionic substitutions in calcium phosphates synthesized
at low temperature. Acta Biomaterialia, 6, 1882–1894.
Bonucci, E. (1967). Fine structure of early cartilage calcification. Journal of Ultrastructure
Research, 20, 33-50.
Bonucci, E. (1992). Calcification in Biological Systems. Boca Raton: CRC Press.
Boonrungsiman, S., Gentleman, E., Carzaniga, R., Evans, N. D., McComb, D. W., Porter, A. E.,
& Stevens, M. M. (2012). The role of intracellular calcium phosphate in osteoblast-
mediated bone apatite formation. PNAS, 109(35), 14170-14175.
Boskey, A. L. (1997). Amorphous Calcium Phosphate: The Contention of Bone. Journal of
Dental Research, 76(8), 1433-1436.
Boskey, A. L., & Posner, A. S. (1976). Extraction of a Calcium-Phospholipid-Phosphate
Complex from Bone. Calcified Tissue Research, 19, 273-283.
Bouligand, Y. (1972). Twisted Fibrous Arrangements in Biological Materials and Cholesteric
Mesophases. Tissue & Cell, 4, 189-217.
Bouligand, Y. (1972). Twisted Fibrous Arrangemnts in Biological Materials and Cholesteric
Mesophases. Tissue & Cell, 4(2), 189-217.
Bouligand, Y. (2008). Liquid crystals and biological morphogenesis: Ancient and new questions.
C. R. Chimie, 11, 281-296.
Braddock, M., Houston, P., Campbell, C., & Ashcroft, P. (2001). Born Again Bone: Tissue
Engineering for Bone Repair. News Physiol. Sci., 16, 208-213.

227
Brodsky, B., Werkmeister, J. A., & Ramshaw, J. A. (2005). Collagens and Gelatins. Biopolymers
Online, 119-128.
Cai, Y., & Tang, R. (2008). Calcium Phosphate nanoparticles in biomineralization and
biomaterials. Journal of Materials Chemistry, 18, 3775-3787.
Calderin, L., & Stott, M. J. (2003). Electronic and crystallographic structure of apatites. Physical
Review B, 67, 134106.
Cardenas, G., Cabrera, G., Taboada, E., & Miranda, S. P. (2004). Chitin Characterization by
SEM, FTIR, XRD, and 13C Cross Polarization Mass Angle Spinning NMR. Journal of
Applied Polymer Science, 93(4), 1876 - 1885.
Chai, Y. C., Carlier, A., Bolander, J., Roberts, S. J., Geris, L., Schrooten, J., . . . Luyten, F. P.
(2012). Current views on calcium phosphate osteogenicity and the translation into
effective bone regeneration strategies. Acta Biomaterialia, 8, 3876-3887.
Chandrasekhar, S., Sadashiva, B. K., & Suresh, K. A. (1977). Liquid crystal of disc-like
molecules. Pramana, 9(5), 471-480.
Chappell, H. F., & Bristowe, P. D. (2007). Density functional calculations of the properties of
silicon-substituted hydroxyapatite. J Mater Sci: Mater Med, 18, 829-837.
Chen, P. Y., Lin, A. Y., McKittrick, J., & Meyers, M. A. (2008). Structure and mechanical
properties of crab exoskeletons. Acta Biomaterialia, 4, 587-596.
Chen, P.-Y., A.Y.M., L., Y.-S., L., Seki, Y., Stokes, A., Peyras, J., . . . McKittrick, J. (2008).
Structure and mechanical properties of selected biological materials. Journal of the
Mechanical Behavior of Biomedical Materials I, 208-226.
Chichester, C. O. (1982). Advances in Food Research. New York: Academic Press, Inc.
Ciftcioglu, N., & McKay, D. S. (2010). Pathological Calcification and Replicating Calcifying-
Nanoparticles: General Approach and Correlation. Pediatric Research, 67(5), 490-499.
Cohen, E. (1993). Chitin Synthesis and Degradation as Targets for Pesticide Action. Archives of
Insect Biochemistry and Physiology, 22, 245-261.
Cohen, I. K., & Mast, B. A. (1990). Models of Wound Healing. The Journal of Trauma, 30(12),
S149-S155.

228
Colfen, H., & Mann, S. (2003). Higher-Order Organization by Mesoscale Self-Assembly and
Transformation of Hybrid Nanostructures. Angew. Chem. Int. Ed., 42, 2350-2365.
Collings, P. J. (2002). Liquid crystals: nature's delicate phase of matter (2nd ed.). Princeton:
Princeton University Press.
Corno, M., Busco, C., Civalleri, B., & Ugliengo, P. (2006). Periodic ab initio study of structural
and vibrational features of hexagonal hydroxyapatite Ca10(PO4)6(OH)2. Physical
Chemistry Chemical Physics, 8, 2464–2472.
Costa-Pinto, A. R., Reis, R. L., & Neves, N. M. (2011). Scaffolds Based Bone Tissue
Engineering: The Role of Chitosan. Tissue Engineering: Part B, 17(5), 1-18.
Cowley, J. M. (1981). Coherent Interference Effects in SIEM and CBED. Ultramicroscopy, 7,
19-26.
Cowley, J. M. (1984). Microdiffraction and STEM of Interfaces. Ultramicroscopy, 14, 27-36.
Cowley, J. M. (1996). Electron Nnaodiffraction: Progress and Prospects. Journal of Electron
Microscopy, 45, 3-10.
Cowley, J. M. (1999). Electron Nanodiffraction. Microscopy Research and Technique, 46, 75-
97.
Cowley, J. M. (2004). Applications of electron nanodiffraction. Micron, 35, 345-360.
Cowley, J. M., & Spence, J. C. (2000). Nanodiffraction. In H. S. Nalwa, Handbook of
Nanostructured Materials and Nanotechnology: Spectroscopy and Theory (Vol. 2, pp. 2-
83). San Diego: Academic Press.
Cowley, J. M., & Sundell, F. A. (1997). Nanodiffraction and dark-field STEM characterization
of single-walled carbon nanotube ropes. Ultramicroscopy, 68, 1-12.
Cowley, J. M., Janney, D. E., Gerkin, R. C., & Buseck, P. R. (2000). The Structure of Ferritin
Cores Determined by Electron Nanodiffraction. Journal of Structural Biology, 131, 210-
216.
Cui, F., Li, Y., & Ge, J. (2007). Self-assembly of mineralized collagen composites. Materials
Science and Engineering R, 57, 1-27.

229
Daamen, W. F., van Moerkerk, H. T., Hafmans, T., Buttafoco, L., Poot, A. A., H, V. J., & van
Kuppevelt, T. H. (2003). Preparation and avaluation of molecularly-defined collagen-
elastin-glycosaminoglycan scaffolds fr tissue engineering. Biomaterials, 24, 4001-4009.
de Leeuw, N. H. (2010). Computer simulations of structures and properties of the biomaterial
hydroxyapatite. Journal of Materials Chemistry, 20, 5376-5389.
Dee, K. C., Puleo, D. A., & Bizios, R. (2002). An Introduction to Tissue-Biomaterial
Interactions. Hoboken: John Wiley & Sons, Inc.
Demers, C., Hamdy, C. R., Corsi, K., Chellat, F., Tabrizian, M., & Yahia, L. (2002). Natural
coral exoskeleton as a bone graft substitute: A review. Bio-Medical Materials and
Engineering, 12, 15-35.
Desbrieres, J., & Babak, V. G. (2008). Interfacial Properties of Amphiphilic Systems on the
Basis of Natural Polymers - Chitin Deratives. Russian Journal of General Chemistry,
78(11), 2230-2238.
Di Silvio, L. (2009). Cellular response to biomaterials. Boca Raton: CRC Press.
Diaz-Rojas, E. I., Arguelles-Monal, W., Higuera-Ciapara, I., Hernandez, J., Lizardi-Mendoza, J.,
& Goycoolea, F. M. (2006). Determination of Chitin and Protein Contents During the
Isolation of Chitin from Shrimp Waste. Macromolecular Bioscience, 6, 340–347.
Dorozhkin, S. (2012). Nanodimensional and Nanocrystalline Calcium Orthophosphates.
American Journal of Biomedical Engineering, 2(3), 48-97.
Dorozhkin, S. V., & Epple, M. (2002). Biological and Medical Significance of Calcium
Phosphates. Angewandte Chemie International Edition, 41, 3130-3146.
Dorsett-Martin, W. A., & Wysocki, A. B. (2008). Rat Models of Skin Wound Healing. In P. M.
Conn, Sourcebook of Models for Biomedical Research (pp. 631-638). Totowa: Humana
Press Inc.
Ducy, P., Schinke, T., & Karsenty, G. (2000). The Osteoblast: A Sophisticated Fibroblast under
Central Surveillance. Science, 289, 1501-1504.
Dunlop, J. W., & Fratzl, P. (2010). Biological Composites. The Annual Review of Materials
Research, 40, 1-24.

230
Dutta, P. K., Ravikumar, M. N., & Dutta, J. (2002). Chitin and Chitosan for Versatile
Applications. Journal of Macromolecular Science, C42(3), 307-354.
Dutta, P. K., Ravikumar, M., & Joydeep, D. (2002). Chitin and Chitosan for versatile
Applications. Journal of Mcromolecular Science Part C-Polymer Reviews, C42(3), 307-
354.
Eanes, E. D., Gillessen, I. H., & Posner, A. S. (1965). Intermediate states in the precipitation of
hydroxyapatite. Nature, 208, 365-367.
Eanes, E. D., Termine, J. D., & Posner, A. S. (1967). Amorphous Calcium Phosphate in Skeletal
Tissues. Clinical Orthopaesics and Related Research, 53, 223-235.
Ebe, T., & Kobayashi, S. (1972). Fine Structure of Human Cells and Tissues. New York: John
Wiley & Sons Inc.
Ehrlich, H. (2010). Biological Materials of Marine Origin. New York: Springer.
Ehrlich, H. (2010). Biological Materials of Marine Origin, Biologically-Inspired Systems 1. New
York: Springer.
Ehrlich, H. (2010). Chitin and collagen as universal and alternative templates in
biomineralization. International Geology Review, 52(7-8), 661-699.
Ehrlich, H., Koutsoukos, P. G., Demadis, K. D., & Pokrovsky, O. S. (2008). Principles of
demineralization: Modern strategies for the isolation of organic frameworks Part I.
Common definitions and history. Micron, 39, 1062-1091.
Enderle, J., Blanchard, S., & Bronzino, J. (2005). Introduction to Biomaedical Engineering (2nd
Edition ed.). Amsterdam: Elsevier Academic Press.
Falanga, V. (2005). Wound healing and its impairment in teh diabetic foot. The Lancet, 366,
1736-1743.
Falini, G., & Fermani, S. (2004). Chitin Mineralization. Tissue Engineering, 10(1/2), 1-6.
Ferrell Jr., R. E., & Paulson, G. G. (1977). Practical Bmitations of selected area electron
diffraction technlques for identifying mineral particles. Micron, 8, 47-55.

231
Fleet, M. E., & Liu, X. (2007). Coupled substitution of type A and B carbonate. Biomaterials,
28, 916-926.
Fleet, M. E., Liu, X., & King, P. L. (2004). Carbonate-hydroxylapatite. American Mineralogist,
89, 14221432.
Franchi, M., Raspanti, M., Dell'Orbo, C., Quaranta, M., De Pasquale, V., Ottani, V., & Ruggeri,
A. (2008). Different Crimp Patterns in Collagen Fibrils Relate to the Subfibrillar
Arrangement. Connective Tissue Research, 49, 85-91.
Fratzl, P. (2007). Biomimetic materials research: what can we really learn from nature’s
structural materials? Journal of The Royal Society Interface, 4, 637–642.
Freeman, R. V., & Otto, C. M. (2005). Spectrum of Calcific Aortic Valve Disease :
Pathogenesis, Disease Progression, and Treatment Strategies. Circulation, 111, 3316-
3326.
Fultz, B. (2013). Transmission Electron Microscopy and Diffractometry. Berlin: Springer.
Furuhashi, T., Schwarzinger, C., Miksik, I., Smrx, M., & Beran, A. (2009). Molluscan shell
evolution with review of shell calcification hypothesis. Comparative Biochemistry and
Physiology, Part B, 154, 351-371.
Gajjeraman, S., Narayanan, K., Hao, J., Qin, C., & George, A. (2007). Matrix Macromolecules
in Hard Tissues Control the Nucleation and Hierarchical Assembly of Hydroxyapatite.
Journal of Biological Chemistry, 282(2), 1193-1204.
Ge, Z., Baguenard, S., Lim, L. Y., Wee, A., & Khor, E. (2004). Hydroxyapatite–chitin materials
as potential tissue engineered bone substitutes. Biomaterials, 25, 1049-1058.
Giordano, C., Sanginario, V., Ambrosio, L., Silvio, L. D., & Santin, M. (2006). Chemical-
Physical Characterization and in vitro Preliminary Biological Assessment of Hyaluronic
Acid Benzyl Ester-hydroxyapatite Composite. Journal of biomaterials applications,
20(3), 237-252.
Giraud Guille, M. M., Mosser, G., Helary, C., & Eglin, D. (2005). Bone matrix like assemblies
of collagen: From liquid crystals to gels and biomimetic materials. Micron, 36, 602-608.
Giraud-Guille, M. M. (1984). Calcification initiation sites in the crab cuticle: The interprismatic
septa. Cell and Tissue Research, 236, 413-420.

232
Giraud-Guille, M. M. (1996). Twisted Liquid Crystalline Supramolecular Arrangements in
Morphogenesis. lnrernarional Review of Cytology, 166, 59-101.
Giraud-Guille, M. M., Belamie, E., & Mosser, G. (2004). Organic and mineral networks in
carapaces bones and biomimetic materials. C. R. Palevol, 3, 503–513.
Giraud-Guille, M.-M. (1992). Liquid Crystallinity in Condensed Type I Collagen Solutions: A
Clue to the Packing of Collagen in Extracellular Matrices. J. Mol. Biol., 861-873.
Giraud-Guille, M.-M. (1994). Liquid Crystalline Order of Biopolymers in Cuticles and Bones.
Microscopy Research and Technique, 27, 420-428.
Giraud-Guille, M.-M., & Besseau, L. (1998). Banded Patterns in Liquid Crystalline Phases of
Type I Collagen: Relationship with Crimp Morphology in Connective Tissue
Architecture. Connective Tissue Research, 37(3), 183-193.
Glimcher, M. J., Bonar, L. C., Grynpas, M. D., Landis, W. J., & Roufosse, A. H. (1981). Recent
Studies of Bone Mineral: Is the Amorphous Calcium Phospahte Theory Valid? Journal of
Crystal Growth, 53, 100-119.
Glimcher, M. J., Bonar, L. C., Grynpas, M. D., Landis, W. J., & Roufosse, A. H. (1981). Recent
Studies of Bone Mineral: Is the Amorphous Calcium Phosphate Theory Valid? Journal of
Crystal Growth, 53, 100-119.
Gobeaux, F., Mosser, G., Anglo, A., Panine, P., Davidson, P., Giraud-Guille, M., & Belamie, E.
(2008). Fibrillogenesis in Dense Collagen Solutions: A Physicochemical Study. J. Mol.
Biol., 1509–1522.
Godin, B., & Touitou, E. (2007). Transdermal skin delivery: Predictions for humans from in
vivo, ex vivo and animal models. Advanced Drug Delivery Reviews, 59(11), 1152-1161.
Gower, L. B. (2008). Biomimetic Model Systems for Investigating the Amorphous Precursor
Pathway and Its Role in Biomineralization. Chemical Reviews, 4551–4627.
Graham, C., Waitkoff, H. K., & Hier, S. W. (1948). The amino acid content of Some
Scleroproteins. The Journal of Biological Chemistry, 529-532.
Graves, D. (2008). Cytokines that promote periodontal tissue destruction. Journal of
periodontology, 79(8 Suppl), 1585S-1591S.

233
Greco, R. S. (1994). Implantation Biology: The Host Response and Biomedical Devices. Boca
Raton: CRC Press, Inc.
Gruskin, E., Doll, B. A., Futrell, F. W., Schmitz, J. P., & Hollinger Jeffrey, O. (2012).
Demineralized bone matrix in bone repair: History and use. Advanced Drug Delivery
Reviews, 64, 1063-1077.
Habibovic, P., & de Groot, K. (2007). Osteoinductive biomaterials – properties and relevance in
bone repair. Journal of tissue engineering and regenerative medicine, 1(1), 25-32.
Hackshaw, A. (2008). Small studies: strengths and limitations. The European Respiratory
Journal, 32(5), 1141-1143.
Hammersen, F. (1976). Histology: A Colour Atlas of Cytology, Histology, and Microscopic
Anatomy. Philadelphia: Lea & Febiger.
Harries, J. E., Huskins, D. W., & Hasnain. (1986). Analysis of the EXAFS spectrum of
hydroxyapatite. J. Phys. C: Solid State Phys, 19, 6859-6872.
Hee, C. K., & Nicoll, S. B. (2006). Induction of Osteoblast Differentiation Markers in Human
Dermal Fibroblasts: Potential Application to Bone Tissue Engineering. Proceedings of
the 28th IEEE Engineering in Medicine and Biology Science Annual International
Conference, 521-524.
Hench, L. L. (1998). Bioactive Materials: The Potential for Tissue Regeneration. Biomedical
Materials Research, 41, 511-518.
Hench, L. L., & Best, S. M. (2013). Ceramics, Glasses, and Glass-Ceramics: Basic Principles. In
B. D. Ratner, H. A. S, F. J. Schoen, & J. E. Lemons, Biomaterial Science: An
Introduction to Materials in Medicine (pp. 130-151). Amsterdam: Elsevier.
Hertz, A., & Bruce, I. J. (2007). Inorganic materials for bone repair or replacement applications.
Nanomedicine, 2(6), 899-918.
Heuer, A. H., Fink, D. J., Laraia, V. J., Arias, J. L., Calvert, P. D., Kendall, K., . . . Caplan, A. I.
(1992). Innovative Materials Processing Strategies: A Biomimetic Approach. Science,
255(5048), 1098-1105.

234
Higman, E. B., Schmeltz, I., & Schlotzhauer, W. S. (1970). Products from the Thermal
Degradation of Some Naturally Occurring Materials. Journal of Agricultural and Food
Chemistry, 18(4), 636-639.
Hin, T. S. (2004). Engineering Materials for Biomedical Applications. London: World Scientific.
Hing, K. A. (2004). Bone repair in the twenty-first century: biology, chemistry or engineering?
Philosophical Transaction of the Royal Society A, 362, 2821-2850.
Hinman, M., Markarewicz, C., asara, J., Cody, G., & Tuross, N. (2008). Degradation Patterns in
Archaeological Collagen: New Evidence from Pyrolysis GC-MS and Solid Satae 13C
NMR. Retrieved September 21, 2009, from Degradation Patterns in Archaeological
Collagen: New Evidence from Pyrolysis GC-MS and Solid Satae 13C NMR:
http://www.socarchsci.org/poster/Hinman_SAA_poster_2008.pdf
Hirano, S., Nakahira, T., Nakagawa, M., & Kim, S. K. (1999). The preparation and applications
of functional fibres from. Journal of Biotechnology, 70, 373–377.
Holzapfel, B. M., Chhaya, M. P., Melchels, F. P., Holzapfel, N. P., Prodinger, P. M., von
Eisenhart-Rothe, R., . . . Hutmacher, D. (2013). Can Bone Tissue Engineering Contribute
to Therapy Concepts after Resection of Musculoskeletal Sarcoma? Sarcoma, 1-10.
Horst, M. N. (1989). Association between Chitin Synthesis and Protein Synthesis in the Shrimp
Penaeus vannamei. Journal of Crustacean Biology, 9(2), 257-265.
Huggins, C. B. (1931). The formation of bone under the influence of epithelium of the urinary
tract. Archives of Surgery, 22(3), 377-408.
Hull, A. W., & Davey, W. P. (1921). Graphical Determination of Hexagonal and Tetragonal
Crystal Structures from X-Ray Data. 17(5), 549-570.
Ibrahim, D. M., Mostafa, A. A., & Korowash, S. I. (2011). Chemical characterization of some
substituted hydroxyapatites. Chemistry Central Journal, 5, 1-11.
Iconomidou, V. A., Willis, J. H., & Hamodrakas, S. J. (2005). Unique features of the structural
model of ‘hard’cuticle proteins: implications for chitin–protein interactions and cross-
linking in cuticle. Insect Biochemistry and Molecular Biology, 35, 553–560.

235
Ito, A., & Onuma, K. (2003). Growth of Hydroxyapatite Crystals. In K. Byrappa, T. Ohachi, W.
Michaeli, H. Warlimont, & E. Weber, Crystal Growth Technology (pp. 525-559). Kyoto:
William Andrew Inc.
Jahnen-Dechent, W. (2004). Lot’s Wife’s Problem Revisited: How We Prevent Pathological
Calcification. In E. Bäuerlein, Biomineralization: From Biology to Biotechnology and
Medical Application (pp. 245-268). Weinheim: WILEY-VCH Verlag GmbH & Co.
Janeway, C. H., Travers, P., Walport, M., & Shomchik, M. J. (2005). Immuno Biology: the
immune system in health and disease (6th Edition ed.). New York: Garland Science.
Janicki, P., & Schmidmaier, G. (2011). What should be the characteristics of the ideal bone graft
substitute? Combining scaffolds with growth factors and/or stem cells. Injury, 42, S77-
S81.
Jayakumar, R., & Tamura, H. (2008). Synthesis, characterization and thermal properties of
chitin-g-poly(e-caprolactone) copolymers by using chitin gel. International Journal of
Biological Macromolecules, 32–36.
Jayakumar, R., Prabaharan, M., Nair, S. V., & Tamura, H. (2010). Novel chitin and chitosan
nanofibers in biomedical applications. Biotechnology Advances, 28, 142–150.
Johnson, P. T. (1980). Histology of the Blue Crab, Callinectes sapidus: A Model for the
Decapoda. New York: Praeger.
Jolles, P., & Muzzarelli, R. (1999). Chitin and Chitinases. Boston: Birkhauser Verlag.
Junqueira, L. C., & Montes, G. S. (1983). Biology of Collagen-Proteoglycan Interaction. Arch.
histol. jap., 46(5), 589-629.
Kalfas, I. H. (2001). Principles of bone healing. Neurosurg Focus, 10(4).
Kasai, N., & Kakudo, M. (2005). X-ray Diffraction by Macromolecules. New York: Springer.
Kaszuba, M. (2008, December 17). Why measure zeta potential. Retrieved May 2009, from
Malvern: http://www.brainshark.com/malvern/zetawebinar
Kierszenbaum, A. L. (2007). Histology and Cell Biology: An Introduction to Pathology (2nd
ed.). New York: Mosby, Inc.

236
Kneser, U., Schaefer, D. J., Munder, B., Klemt, C., Andree, C., & Stark, G. B. (2002). Tissue
engineering of bone. Minimally Invasive Therapy & Allied Technologies, 11(3), 107-116.
Kojima, K., Okamoto, Y., Kojima, K., Miyatake, K., Fujise, H., Shigemasa, Y., & Minami, S.
(2004). Effects of Chitin and Chitosan on Collagen Synthesis in Wound Healing. J. Vet.
Med. Sci, 1595-1598.
Kokkoli, E., Mardilovich, A., Wedekind, A., Rexeisen, E. L., Garg, A., & Craig, J. A. (2006).
Self-assembly and applications of biomimetic and bioactive peptide-amphiphiles. The
Royal Society of Chemistry, 2, 1015–1024.
Kolacna, L., Bakesova, J., Varga, F., Kostakova, E., & Planka, L. (2007). Biochemical and
Biophysical Aspects of Collagen Nanostructure in the Extracellular Matrix. Physiol. Res.,
S51-S60.
Köster, S., Evans, H. M., Wong, J. Y., & Pfohl, T. (2008). An In Situ Study of Collagen Self-
Assembly Processes. Biomacromolecules, 9, 199–207.
Krebs, M. R., MacPhee, C. E., Miller, A. F., Dunlop, I. E., Dobson, C. M., & Donald, A. M.
(2004). The formation of spherulites by amyloid fibrils of bovine insulin. PNAS, 101(40),
14420–14424.
Kuhn, K., & Glanville, R. W. (1980). Molecular Structure and Higher Organization of Different
Collagen Types. In A. Vidik, & J. Vuust (Eds.), Biology of Collagen. London: Academic
Press.
Laguens, R. P., & Dumm, C. L. (1969). Atlas of Human Electron Microscropy. Saint Louis: The
C.V. Mosby Company.
Laguens, R. P., & Gomez Dumm, C. L. (1969). Atlas of Human Electron Microscopy. Saint
Louis: The C.V. Mosby Company.
Lamers, E., van Horssen, R., te Riet, J., van Delft, F. C., Luttge, R., Walboomers, X. F., &
Jansen, J. a. (2010). The Influence of Nanoscale Topographical Cues on Initial Osteoblast
Morphology and Migration. European Cells and Materilas, 20, 329-343.
Landis, W. J., & Glimcher, M. J. (1978). Electron Diffraction and Electron Probe Microanalysis
of the Mineral Phase of Bone Tissue Prepared by Anhydrous Techniques. Journal of
Ultrastructure Research, 63, 188-223.

237
Langer, R., & Vacanti, J. P. (1993). Tissue Engineering. Science, 260(5110), 920-926.
Laurent, G. J. (1987). Dynamic state of collagen: pathways of collagen degradation in vivo and
their possible role in regulation of collagen mass. Am. J. physiol, C1-C9.
Leikina, E., Mertts, M. V., Kuznetsova, N., & Leikin, S. (2002). Type I collagen is thermally
unstable at body temperature. Proceedings of the National Academy of Sciences, 99(3),
1314–1318.
Leng, Y., & Qu, S. (2002). TEM examination of single crystal hydroxyapatite diffraction.
Journal of Materials Science Letters, 21, 829-830.
Levander, G. (1934). On the formation of new bone in bone transplantation. Acta chirurgica
Scandinavica, 74, 425-426.
Leventouri, T. (2006). Synthetic and biological hydroxyapatites: Crystal structure questions.
Biomaterials, 27, 3339-3342.
Li, J., Chen, J., & Kirsner, R. (2007). Pathophysiology of acute wound healing. Clinics in
Dermatology, 25, 9-18.
Li, J., Revol, J.-F., & Marchessault, R. H. (1996). Rheological Properties of Aqueous
Suspensions of Chitin Crystallites. Journal of Colloid and Interface Science, 183, 365-
373.
Li, X., & Huang, Z. (2009). Unveiling the Formation Mechanism of Pseudo-Single-Crystal
Aragonite Platelets in Nacre. Physical Review Letters, 102(7), (075502)1-4.
Li, Z. Y., Lam, W. M., Yang, C., Xu, B., Ni, G. X., Abbah, S. A., . . . W, L. W. (2007). Chemical
composition, crystal size and lattice structural changes after incorporation of strontium
into biomimetic apatite. Biomaterials, 28, 1452–1460.
Lin, Y. K., & Liu, D. C. (2006). Comparison of physical–chemical properties of type I collagen
from different species. Food Chemistry, 99, 244–251.
Linton, S. M., & Greenaway, P. (1997). Urate Deposits in the Gecarcinid Land Crab
Gecargoidea Natalis Are Synthesized DE NOVO From Excess Dietary Nitrogen. The
Journal of Experimental Biology, 200(17), 2347-2354.

238
Linton, S. M., & Greenaway, P. (2000). The nitrogen requirements and dietary nitrogen
utilization for the gecarcinid land crab Gecarcoidea natalis. Physiological and
Biochemical Zoology, 73(2), 209-218.
Lowenstam, H. A. (1981). Minerals Formed by Organisms. Science, 211(4487), 1126-1131.
Lowenstam, H. A., & Weiner, S. (1989). On biomineralization. New York: Oxford University
Press.
Lowenstam, H. A., & Weiner, S. (1989). On Biomineralization. New York: Oxford University
Press.
Ma, P. X., & Elisseeff, J. (2006). Saffolding in Tissue Engineering. Boca Raton: Taylor &
Francis.
Mainil-Varlet, P., Gogolewski, S., & Nieuwenhuis, P. (1996). Long-term soft tissue reaction to
various polyactides and their in vivo degradation. Journal of Materials Science:
Materials in Medicine, 7, 713-721.
Mann, S. (1983). Mineralization in Biological Systems. Structure and Bonding, 54, 125-174.
Mann, S. (1988). Molecular recognition in biomineralization. Nature, 332, 119-124.
Mann, S. (1993). Molecular tectonics in biomineralization and biomimetic materials chemistry.
Nature, 365, 499-505.
Mann, S. (1993). Molecular tectonics in biomineralization and biomimetic materials chmeistry.
Nature, 365, 499-505.
Mann, S. (1995). Biomineralization and Biomimetic Materials Chemistry. Journal of Materials
Chemistry, 5(7), 935-946.
Mann, S., Archibald, D. D., Didymus, J. M., Douglas, T., Heywood, B. R., Meldrum, F. C., &
Reeves, N. J. (1993). Crystallization at Inorganic-Organic Interfaces: Biominerals and
Biomimetic Synthesis. Science, 261(5126), 1286-1292.
Marsell, R., & Einhorn, T. A. (2011). The Biology of Frcature Healing. Injury, 42(6), 551-555.
Matsunaga, K. (2010). Theoretical Defect Energetics in Calcium Phosphate Bioceramics.
Jpurnal of American Ceramic Society, 93(1), 1-14.

239
Mendelson, K., & Schoen, F. J. (2006). Heart Valve Tissue Engineering: Concepts, Approaches,
Progress, and Challenges. Annals of Biomedical Engineering, 34(12), 1799–1819.
Menéndez-Proupin, E., Cervantes-Rodríguez, S., Osorio-Pulgar, R., Franco-Cisterna, M.,
Camacho-Montes, H., & Fuentes, M. E. (2011). Computer simulation of elastic constants
of hydroxyapatite and fluorapatite. Journal of the Mechanical Behavior of Biomedical
Materials, 4, 1011-1020.
Meyers, M. A., Chen, P.-Y., Lin, A. Y.-M., & Seki, Y. (2008). Biological materials: Structure
and mechanical properties. Progress in Materials Science, 1–206.
Meyers, M. A., Lin, A. Y., Seki, Y., Chen, P.-Y. K., & Bodde, S. (2006). Structural Biological
Composites: An Overview. JOM, 58(7), 35-41.
Minami, S., Okamoto, Y., Miyatake, K., Matsuhashi, A., Kitamura, Y., Taniqawa, . . . Y, S.
(1996). Chitin induces type IV collagen and elastic fiber in implanted non-woven fabric
of polyester. Carbohydrate Polymers, 298-299.
Monosmith, W. B., & Cowley. (1983). Pattern Recognition Techniques for the Analysis of
Electron Microdiffraction Patterns. Ultramicroscopy, 12, 51-58.
Monzack, E. L., Rodriguez, K. J., McCoy, C. M., Gu, X., & Masters, K. (2011). Natural
Materials in Tissue Engineering Applications. In J. A. Burdick, & R. L. Mauck,
Biomaterials for Tissue Engineering Applications: A Review of the Past and Future
Trends (pp. 209-242). New York: SpringerWien.
Monzack, E. L., Rodriguez, K. J., McCoy, C. M., Gu, X., & Masters, K. S. (2011). Natural
Materials in Tissue Engineering Applications. In J. A. Burdick, & R. L. Mauck,
Biomaterials for Tissue Engineering Applications: A Review of the Past and Future
Trends (pp. 209-241). New York: SpringerWien.
Moss, M. L. (1958). Extraction of an Osteogenic Inductor Factor from Bone. Science, 127, 755-
756.
Mosser, G., Anglo, A., Helary, C., Bouligand, Y., & Giraud-Guille, M. M. (2006). Dense tissue-
like collagen matrices formed in cell-free conditions. Matrix Biology, 25, 3-13.
Mostafa, N. Y., & Brown, P. W. (2007). Computer simulation of stoichiometric hydroxyapatite:
Structure and substitutions. Journal of Physics and Chemistry of Solids, 68, 431–437.

240
Mountziaris, P. M., Spicer, P. P., Kasper, F. K., & Mikos, A. G. (2011). Harnessing and
Modulating Inflammation in Strategies for Bone Regeneration. Tissue Engineering: Part
B, 17, 393-402.
Murray, S., & Neville, A. (1997). The role of the electrostatic coat in the formation of cholestric
liquid crystal spherulites from Alpla-chitin. International Journal of Biological
Macromolecules, 20, 123-130.
Murugan, R., & Ramakrishna, S. (2007). Nanoengineered Biomimetic Bone-Building Blocks. In
G. A. Mansoori, T. F. George, L. Assoufid, & G. Zhang, Molecular Building Blocks for
Nanotechnology: From Diamondoids to Nanoscale Materials and Applications (Vol.
109, pp. 301-352). Springer.
Muzzarelli, R. A. (1999). Native, industrial and fossil chitins. In P. Jolles, & R. A. Muzzarelli
(Eds.), Chitin and Chitinases. Boston: Birkhauser Verlag.
Nader, H. B., Ferreira, T. M., Paiva, J. F., Medeiros, M. G., Jeronimo, S. M., Paiva, V. M., &
Dietrich, C. P. (1984). Isolation and Structural Studies of Heparan Sulfates and
Chondroitin Sulfates from Three Species of M olluscs. The Journal of Biological
Chemistry, 259(3), 1431-1435.
Nandi, S. K., Roy, S., Mukherjee, P., Kundu, B., De, D. K., & Basu, D. (2010). Orthopaedic
applications of bone graft & graft substitutes: a review. Indian J Med Res, 132, 15-30.
Neary, M. t., Reid, D. G., Mason, M. J., Friscic, T., Duer, M. J., & Cusack, M. (2010). Contrasts
between organic participation in apatite biomineralization in brachiopod shell and
vertebrate bone identified by nuclear magnetic resonance spectroscopy. Journal of The
Royal Society Interface, 8, 282–288.
Neira, I., Kolen'ko, Y. V., Lebedev, O. I., Van Tendeloo, G., Gupta, H. S., Matsushita, N., . . .
Guitián, F. (2009). Rational synthesis of a nanocrystalline calcium phosphate cement
exhibiting rapid conversion to hydroxyapatite. Materials Science and Engineering C, 29,
2124–2132.
Neues, F., Ziegler, A., & Epple, M. (2007). The composition of the mineralized cuticle in marine
and terrestrial isopods: A comparative study. CrystEngComm, 9, 1245-1251.
Neuman, W. F., & Neuman, M. W. (1953). The Nature of the Mineral Phase of Bone. Chemical
Reviews, 53(1), 1-45.

241
Neville, A. (1993). Biology of Fibrous Composites: Development Beyond the Cell Membrane.
Cambridge: Cambridge Unversity Press.
Neville, A. C. (1976). The Chitin Crystallite in Arthropod Cuticle. Journal of Cell Science, 21,
73-82.
Neville, A. C. (1993). Biology of fibrous composites: Development beond the cell membrane.
New York: Cambridge University Press.
Neville, A. C., & Luke, B. M. (1971). A Biological System Producing a Self-Assembling
Cholesteric Protein Liquid Crystal. J Cell Sci, 93-109.
Nicoll, S. B. (2011). Materials for Bone Graft Substitutes and Osseous Tissue Regeneration. In J.
A. Burdick, & R. L. Mauck, Biomaterials for Tissue Engineering Applications (pp. 343-
362). New York: Springer-Verlag/Wien.
Nilsen, R. (1980). Electronmicroscopic study of mineralization in induced heterotopic bone
formation in guinea pig. Journal of Dental Research, 88, 340-347.
Nudelman, F., Pieterse, K., George, A., Bomans, P. H., Friedrich, H., Brylka, L. J., . . .
Sommerdijk, A. J. (2010). The role of collagen in bone apatite formation in the presence
of hydroxyapatite nucleation inhibitors. Nature Materials, 9, 1004-1009.
Oaki, Y., & Imai, H. (2013). Recent advances in mesocrystal and their related structures. In P.
O'Brien, Nanoscience: Nanostructures Through Chemistry (pp. 1-28). Cambridge: The
Royal Society of Chemistry.
Omelon, S., Georgiou, J., Henneman, Z. J., Wise, L. M., Sukhu, B., Hunt, T., . . . Grynpas, M. D.
(2009). Control of Vertebrate Skeletal Mineralization by Polyphosphates. PLOS ONE,
4(5), e5634 (1-16).
Ospina, C. A., J, T., Ramirez, A. J., Farina, M., Ellis, D. E., & Rossi, A. M. (2012). Experimental
evidence and structural modeling of nonstoichiometric (0 1 0) surfaces coexisting in
hydroxyapatite nano-crystals. Colloids and Surfaces B: Biointerfaces, 89, 15-12.
Ottani, V., Matin, D., Franchi, M., Ruggeri, A., & Raspanti, M. (2002). Hierarchial structures in
fibrillar collagens. Micron, 33, 587-596.
Ottani, V., Raspantib, M., & Ruggeri, A. (2001). Collagen structure and functional implications.
Micron, 251-260.

242
Park, J., & Lakes, R. S. (2007). Biomaterials: An Introduction. New York: Springer
Science+Business Media, LLC.
Paul, W. E. (2008). Fundamental Immunology (6th Edition ed.). Philadelphia: Wolters Kluwer
Health | Lippinott Williams & Wilkins.
Pearce, A. I., Richards, R. G., Milz, S., Schneider, E., & Pearce, S. G. (2007). ANIMAL
MODELS FOR IMPLANT BIOMATERIAL RESEARCH IN BONE: A REVIEW.
European Cells and Materials, 13, 1-10.
Pecharsky, V. K., & Zavalij, P. Y. (2009). Fundamentals of Powder Diffraction and Structural
Characterization of Materials (2nd ed.). New York: Springer.
Poole, A. R. (1986). Proteoglycans in health and disease: structures and functions. Biochemical
Journal, 236, 1-14.
Porter, K. R., & Bonneville, M. A. (1973). Fine Structure of Cells and Tiissues (4th Edition ed.).
Philadelphia: Lea & Febiger.
Posner, A. S. (1985). The Mineral Bone. Clinical Orthopaedics and Related Research, 200, 87-
99.
Posner, A. S., & Betts, F. (1975). Synthetic Amorphous Calcium Phosphate and Its Relation to
Bone Mineral Structure. Accounts of Chemical Research, 8, 273-281.
Posner, A. S., Perloff, A., & Diorio, A. F. (1958). Refinement of the hydroxyapatite structure.
Acta Crystallographica, 308-309.
Prashanth, K. V., & Tharanathan, R. N. (2007). Chitin/chitosan: modifications and their
unlimited application potential --- an overview. Trends in Food Science & Technology,
18, 117-131.
Qin, L.-C., Iijima, S., Kataura, H., Maniwa, Y., Suzuki, S., & Achiba, Y. (1997). Helicity and
packing of single-walled carbon nanotubes studied by electron nanodiffraction. Chemical
Physics Letters, 268, 101-106.
Quintana, C., Cowley, J. M., & Marhic, C. (2004). Electron nanodiffraction and high-resolution
electron microscopy studies of the structure and composition of physiological and
pathological ferritin. Journal of Structural Biology, 147, 166-178.

243
Ramsden, J. (2008). Bomedical Surfaces. Boston: Artech House.
Rand, M. S. (2008). Selection of Biomedical Animal Models. In P. M. Conn, Sourcebook of
Models for Biomedical Research (pp. 9-15). Totowa: Humana Press Inc.
Rangavittal, N., Landa-Canovas, A. R., Gonzalez-Calbet, J. M., & Vallet-Regı, M. (2000).
Structural study and stability of hydroxyapatite and b-tricalcium phosphate: Two
important bioceramics. Journal of Biomedical Materials Research, 51, 660–668.
Ratner, B. D. (2001). Replacing and Renewing: Synthetic Materials, Biomimetics, and Tissue
Engineering in Implant Dentistry. Journal of Dental Education, 65(12), 1340-1347.
Ratner, B. D., Hoffman, A. S., Schoen, F. J., & Lemons, J. E. (1996). Biomaterials Science: An
Introduction to Materials in Medicine. San Diego: Academic Press.
Reddi, A. H. (1981). Cell biology and biochemistry of endochondral bone development.
Collagen and related research, 1(2), 209-226.
Reddi, A. H. (2000). Morphogenesis and tissue engineering of bone and cartilage: inductive
signals, stem cells, and biomimetic biomaterials. Tissue Engineering, 6(4), 351-359.
Reddi, A. H. (2000). Morphogenesis and Tissue Engineering of Bone and Cartilage: Inductive
Signals, Stem Cells, and Biomimetic Biomaterials. Tissue Engineering, 6(4), 351-359.
Reddi, A. H., & Anderson, W. A. (1976). Collagenous bone matrix-induced endochondral
ossification hemopoiesis. Journal of Cell Biology, 69(3), 557-572.
Reddi, A. H., & Huggins, C. (1972). Biochemical Sequences in the Transformation of Normal
Fibroblasts in Adolescent Rats. Proceedings of the National Academy of Sciences of the
United States of America , 69(6), 1601-1605.
Reddi, A. H., & Huggins, C. B. (1974). Cyclic Electrochemical Inactivation and Restoration of
Competence of Bone Matrix to Transform Fibroblasts. Proceedings of the National
Academy of Sciences of the United States of America, 71(5), 1648-1652.
Reis, R. L., & Weiner, S. (2004). Learning from Nature How to Design New Implantable
Biomaterials: From Biomineralization Fundamentals to Biomimetic Materials and
Processing Routes. Dordrecht: Kluwer Academic Publishers.

244
Reis, R. L., & Weiner, S. (2004). Learning from Nature How to Design New Implantable
Biomaterials: From Biomineralization Fundamentals to Biomimetic Materials and
Processing Routes. Dordrecht: Kluwer Academic Publishers.
Rho, J.-Y., Kuhn-Spearing, L., & Zioupos, P. (1998). Mechanical properties and the hierarchical
structure of bone. Medical Engineering & Physics, 20(2), 92-102.
Ripamonti, U. (1991). The morphogenesis of bone in replicas of porous hydroxyapatite obtained
from conversion of calcium carbonate exoskeletons of coral. The Journal of Bone and
Joint Surgery, 73(5), 692-703.
Ripamonti, U., & Roden, L. (2010). Biomimetics for the induction of bone formation. Expert
Rev. Med. Devices, 7(4), 469-479.
Robey, P. G. (2011). Cell Sources for Bone Regeneration: The Good, the Bad, and the Ugly (But
Promising). Tissue Engineering: Part B, 423-430.
Robinson, R. A., & Watson, M. L. (1955). Crystal-collagen relationships in bone as observed in
the electron microscope: III. Crystals and collagen morphology. Annals of the New York
Academy of Sciences, 60, 596-628.
Roer, R., & Dillaman, R. (1984). The Structure and Calcification of the Crustacean Cuticle.
American Zoologist, 24(4), 893-909.
Rossi, A. L., Barreto, I. C., Maciel, W. Q., Rosa, F. P., Rocha-Leao, m. H., Werckmann, J., . . .
Farina, M. (2012). Ultrastructure of regenerated bone mineral surrounding
hydroxyapatite–alginate composite and sintered hydroxyapatite. Bone, 50, 301-310.
Rousseau, M. (2011). Nacre, a Natural Biomaterial . In r. Pignatello, Biomaterials Applications
for Nanomedicine (pp. 281-298). Rijeka: Intech.
Rutherford, R. B., Moalli, M., Franceschi, R. T., Wang, D., Gu, K., & Krebsbach, P. H. (2002).
Bone Morphogenetic Protein-Transduced Human Fibroblasts Convert to Osteoblasts and
Form Bone in Vivo. Tissue Engineering, 8(3), 441-452.
Salgado, P. C., Sathler, P. C., Castro, H. C., Alves, G. G., de Oliveira, A. M., de Oliveira, R. C., .
. . Granjeiro, J. M. (2011). Bone Remodeling, Biomaterials and Technological
Applications: Revisiting Basic Concepts. Journal of Biomaterials and
Nanobiotechnology, 2, 318-328.

245
Sampath, T. K., & Reddi, A. H. (1981). Dissociative extraction and reconstitution of
extracellular matrix components involved in local bone differentiation. Proceedings of
the National Academy of Sciences of the United States of America, 78(12), 7599-7603.
Sarikaya, M. (1994). An Introduction to Biomimetics: A Structural Viewpoint. Microscopy
Research and Technique, 27, 360-375.
Sarikaya, M., Liu, J., & Aksay, I. A. (1995). Nacre: Properties, Crystallography, Morphology,
and Formation. In M. Sarikaya, & I. A. Aksay, Biomimetics: Design and Processing of
Materials (pp. 35-90). Woodbury: AIP Press.
Sato, K. (2007). Mechanism of Hydroxyapatite Mineralization in Biological Systems. Journal of
the Ceramic Society of Japan, 115(2), 124-130.
Schneer, C. J. (1957). A Modified Hull-Davey Chart for High Values of c/a. American
Mineralogist, 4, 282-285.
Schoen, F. J. (2013). Introduction: Rebuilding Humans Using Biology and Biomaterials. In B. D.
Ratner, A. S. Hoffman, F. J. Schoen, & J. E. Lemons, Biomaterials Science: An
Introduction to Materials in Medicine (pp. 1119-1122). Amsterdam: Elsevier.
Scott, J. E. (1975). Composition and Structure of the Pericellular Environment: Physiological
Function and Chemical Composition of Pericellular Proteoglycan (an evolutionary view).
Phil Trans R Soc Land B , 271, 235-242.
Scott, J. E. (1988). Proteoglycan-fibrillar collagen interactions. Biochem. J., 252, 313-323.
Scott, J. E., & Tigwell, M. J. (1978). Periodate Oxidation and the Shapes of
Glycosaminoglycuronans in Solution. Biochemical Journal, 173, 103-114.
Scott, J. E., Orford, C. R., & W, H. E. (1981). Proteoglycan-collagen arrangements in developing
rat tail tendon. Biochem. J., 195, 573-581.
Selvig, K. A. (1970). Periodic Lattice Images of Hydroxyapatite Crystals in Human Bone and
Dental Hard Tissues. Calcified Tissue Research, 6, 227-238.
Senn, N. (1889). On the healing of aseptic bone cavities by implantation of antiseptic decalcified
bone. Annals of Surgery, 10(5), 352-368.

246
Shanmugasundaram, N., Ravikumar, T., & Babu, M. (2004). Comparative Physico-chemical and
in Vitro Properties of Fibrillated Collagen Scaffolds from Different Sources. Journal of
Biomaterials Applications, 247-264.
Shehab, D., Elgazzar, A. H., & Collier, D. (2002). Heterotopic Ossification. THE JOURNAL OF
NUCLEAR MEDICINE, 43(3), 346-353.
Shimomura, M. (2010). The New Trends in Next Generation Biomimetics Material Technology:
Learning from Biodiversity. Science & Technology Trends, 37, 53-75.
Simmer, J. P., & Fincham, A. G. (1995). Molecular Mechanisms of Dental Enamel Formation.
Critical Reviews in Oral Biology & Medicine, 6(2), 84-108.
Singh, D. K., & Ray, A. R. (2000). Biomedical Applications of Chitin, Chitosan, and Their
Derivatives. J.M.S-Rev. Macrolmol Chem Phys, 69-83.
Singh, D., & Ray, A. R. (2000). Biomedical Applications of Chitin, Chitosan, and Their
Derivatives. Journal of Macromolecular Science, Part C: Polymer Reviews, 40, 69-83.
Singh, S., & Jonnalagadda, S. B. (2013). Synthesis of Thermally Stable Metal Substituted
Hydroxy Apatites for the Selective Oxidation of Light Paraffins. Bull. Chem. Soc.
Ethiop., 27(1), 57-68.
Skinner, H. C., & Jahren, A. H. (2003). Biomineralization. Treatise on Geochemistry, 8, 117-
184.
Skinner, H. C., & Jahren, A. H. (2003). Biomineralization.
Slack, H. G. (1955). A Short Review of Connective Tissue Metabolism. Ann. Rheum. Dis, 14,
238-242.
Smith, & Wood. (1991). Molecular and Cell Biochemistry: Biological Molecules. London:
Chapman & Hall.
Smith, C. A., & Wood, E. J. (1991). Biological Molecules: Molecular and Cell Biochemistry.
New York: Chapman & Hall.
Smyth, M. S., & Martin, J. H. (2000). x Ray Crystallography. Journal of Clinical Pathology-
Molecular Pathology, 8-14.

247
Stanley, L., Zlotin, B., Bolckmans, K., & Zusman, A. (2005, June 16). Basic I-TRIZ Course.
Retrieved from www.ideabrain.co.kr/Basic%20I-TRIZ%20Course/index.html
Stephen, A. M. (1995). Food polysaccharides and their applications. New York: Marcel Dekker.
Stern, R., Asari, A. A., & Sugahara, K. N. (2006). Hyaluronan fragments: An information-rich
system. European Journal of Cell Biology, 85, 699-715.
Stevens, M. M., & George, J. H. (2005). Exploring and Engineering the Cell Surface Interface.
Science, 310, 1135-1138.
Stocker, S., & Wolinetz, C. D. (2009). Bone Builders: The Science of Grafts, Biomaterials, and
Bone Engineering. Breakthroughs in Bioscience, 1-13.
Summerlee, A. J. (2002). Bone formation and development. In G. Sumner-Smith, Bone in
clinical orthopedics (pp. 1-21). Switzerland: AO Publishing.
Suneelkumar, C., Datta, K., Srinivasan, M. R., & Kumar, S. T. (2008). Biphasic calcium
phosphate in periapical surgery. Journal of Conservative Dentistry, 11(2), 92-96.
Suryanarayana, C., & Norton, M. G. (1998). X-Ray diffraction : a practical approach. New
York: Plenum Press.
Suvorova, E. I., & Buffat, P. A. (1999). Electron diffraction from micro- and nanoparticles of
hydroxyapatite. Journal of Microscopy, 196(1), 46-58.
Suvorova, E. I., & Buffat, P. A. (1999). Electron diffraction from micro- and nanoparticles of
hydroxyapatite. Journal of Microscopy, 196(1), 46-58.
Suvorova, E. I., & Buffat, P. A. (2001). Electron Diffraction and High Resolution Transmission
Electron Microscopy in the Characterization of Calcium Phosphate Precipitation from
Aqueous Solutions under Biomineralization Conditions. European Cells and Materials,
1, 27-42.
Suvorova, E. I., & Buffat, P. A. (2001). Size Effect in X-ray and Electron Diffraction Patterns
from Hydroxyapatite Particles. Crystallography Reports, 46(5), 722–729.
Takeuchi, Y., Matsumoto, T., Ogata, E., & Shishiba, Y. (1990). Isolation and characterization of
proteoglycans synthesized by mouse osteoblastic cells in culture during the
mineralization process. Biochem J, 266, 15-24.

248
Tam, K. F., Wang, K., & Fan, W. C. (2002). Metastatic Calcification. Journal of Hong Kong
College of Radiologists, 5, 186-189.
Tanodekaewa, S., Prasitsilpa, M., Swasdison, S., Thavornyutikarna, B., Pothsreea, T., &
Pateepasen, R. (2004). Preparation of acrylic grafted chitin for wound dressing
application. Biomaterials, 25, 1453–1460.
Tao, J., Pan, H., Zeng, Y., Xu, X., & Tang, R. (2007). Roles of amorphous calcium phosphate
and biological additives in the assembly of hydroxyapatite nanoparticles. The Journal of
Physical Chemistry B, 111(47), 13410-13418.
Tao, J., Zhou, D., Zhang, Z., Xu, X., & Tang, R. (2009). Magnesium-aspartate-based
crystallization switch inspired from shell molt of crustacean. PNAS, 106(52), 22096-
22101.
Termine, J. D. (1972). Mineral Chemistry and Skeletal Biology. Clinical Orthopaedics and
Related Research, 85, 207-241.
Termine, J. D., & Posner, A. S. (1967). Amorphous / Crystallinne Interrelationship in Bone
Mineral. Calcified Tissue Research, 1, 8-23.
Termine, J. D., & S, P. A. (1967). Amorphous/Crystalline Interrelationships in Bone Mineral.
Calcified Tissue Research, 1, 8-23.
Thomas, M. V., & Puleo, D. A. (2011). Infection, Inflammation, and Bone Regeneration: a
Paradoxical Relationship. Critical Reviews in Oral biology & Medicine, 90(9), 1052-
1061.
Travis, D. F. (1963). Structural features of mineralization from tissue to macromolecular levels
of organization in Decapod Crustacea. Annuals of New York Academy of Sciences, 109,
177-245.
Trim, A. R. (1941). Studies in the chemistry of the insect cuticleI. Some general observations on
certain arthropod cuticles with special reference to the characterization of the proteins.
Biochemical Journal, 35, 1088–1098.
Tsai, G.-J., Su, W.-H., Chen, H.-C., & Pan, C.-L. (2002). Antimicrobial acivity of shrimp chitin
and chitosan from different treatments and applications of fish preservation. Fisheries
Science, 68, 170-177.

249
Tsuji, T., Onuma, K., Yamamoto, A., Iijima, M., & Shiba, K. (2008). Direct transformation from
amorphous to crystalline calcium phosphate facilitated by motif-programmed artificial
proteins. PNAS, 105(44), 16866-16870.
Turing, A. M. (1952, August 14). The Chemical Basis of Morphogenesis. Philosophical
Transactions of the Royal Society of London. Series B, Biological Sciences, 237(641), 37-
72.
Tzaphlidou, M., & Zaichick, V. (2003). Calcium, Phosphorus, Calcium–Phosphorus Ratio in Rib
Bone of Healthy Humans. Biological Trace Element Research, 93, 63-74.
Urist, M. R. (1965). Bone: Formation by Autoinduction. Science, 150, 893-899.
Urist, M. R., & McLean, F. C. (1952). Osteogenetic potency and new-bone formation by
induction in transplants to the anterior chamber of the eye. The Journal of bone and joint
surgery. American, 34-A(2), 443-476.
Urist, M. R., & Strates, B. S. (1971). Bone morphogenetic protein. Journal of Dental Research,
50, 1392-1406.
Valta, M. P., Hentunen, T., Qu, Q., Valve, E. M., Harjula, A., Seppanen, J. A., . . . Harkonen, P.
L. (2006). Regulation of Osteoblast Differentiation: A Novel Function for Fibroblast
Growth Factor 8. Endocrinology, 147(5), 2171–2182.
van Blitterswijk, C. (2007). Tissue Engineering. Amsterdam: Elsevier.
Veis, A. (2003). Mineralization in organic matrix frameworks. Rev Mineral Geochem, 54, 249-
289.
Vincent, J. F. (2002). Arthropod cuticle: a natural composite shell system. Composites: Part A,
33, 1311–1315.
Vincent, J. F. (2005). Deconstructing the design of a biological material. Journal of Theoretical
Biology, 236, 73-78.
Vincent, J. F. (2008). Biomimetic materials. Journal of Materials Research, 23(12), 3140-.
Vincent, J. F., & Mann, D. L. (2002). Systematic technology transfer from biology to
engineering. Phil. Trans. R. Soc. Lond. A, 360, 159-173.

250
Vincent, J. F., Bogatyreva, O. A., Bogatyrev, N. R., Bowyer, A., & Pahl, A.-K. (2006).
Biomimetics: its practice and theory. Journal of the Royal Society Interface, 3, 471-482.
Vincent, J. F., Bogatyreva, O., Pahl, A.-K., Bogatyrev, N., & Bowyer, A. (2005). Putting
Biology into TRIZ: A Database of Biological Effects. Creativity and Innovation
Management, 14(1), 66-72.
Vincent, S., & Perrimon, N. (2001, May 31). Developmental biology: Fishing for morphogens.
Nature, 411, 533-536.
Wahl, D. A., & Czernuszka, J. T. (2006). Collagen-Hydroxyapatite Composites for Hard Tissue
Repair. European Cells and Materials, 11, 43-56.
Wainwright, S. A. (1982). Mechanical design in organisms. Princeton: Princeton University
Press.
Wanchoo, R. K., Thakur, A., & Sweta. (2008). Viscometric and Rheological Behaviour of
Chitosan-Hydrophilic Polymer Blends. Chem. Biochem. Eng. Q., 22(1), 15-24.
Wang, X.-Y., Zuo, Y., Huang, D., Hou, X.-D., & Li, Y.-B. (2010). Comparative Study on
Inorganic Composition and Crystallographic Properties of Cortical and Cancellous Bone.
Biomedical and Environmental Sciences, 23, 473-480.
Watabe, N. (1974). Crystal growth of calcium carbonate in biological systems. Journal of
Crystal Growth, 24-25, 116-122.
Weidenthaler, C. (2011). Pitfalls in the characterization of nanoporous and nanosized materials.
Nanoscale, 3, 792-810.
Weiner, S., & Dove, P. M. (2003). An overview of biomineralization processes and the problem
of the vital effect. Reviews in Mineralogy and Geochemistry, 54, 1-29.
Weiner, S., & Wagner, H. D. (1998). The Material Bone: Structure-Mechanical Function
Relations. Annual Review of Materials Science, 28, 271-298.
Westbroek, P., & Marin, F. (1998). A marriage of bone and nacre. Nature, 392, 861-862.
White, E. W., Weber, J., Roy, D. M., Owen, E. L., Chiroff, R. T., & White, R. A. (1975).
Replamineform porous biomaterials for hard tissue implant applications. Journal of
Biomedical Materials Research Part A, 9(4), 23-27.

251
Williams, D. B., & Carter, C. B. (2009). Transmission Electron Microscopy (2nd ed.). New
York: Springer.
Williams, D. B., & Carter, C. B. (2009). Transmission Electron Microscopy (2nd ed.). New
York: Springer.
Williamson, W. C. (1860). On some histological features in the shells of the crustacea. Quarterly
Journal of Microscopical Science, 8, 35-47.
Wilson Jr., O. C. (2008). Bone Inspired Nanocomposites. In L. M. Krause, & J. T. Walter, New
Research on Nanocomposites (pp. 57-78). New York: Nova Science Publishers, Inc.
Wilson, O. C., Gugssa, A., Mehl, P., & Anderson, W. (2012). An initial assessment of the
biocompatibility of crab shell for bone tissue engineering. Materials Science and
Engineering C, 32, 78-82.
Wilson, R. M., Elliot, J. C., & Dowker, S. E. (1999). Hydroxylapatite. American Mineralogist,
84, 1406-1414.
Winter, G. D., & Simpson, B. J. (1969). Heterotopic bone formed in a synthetic sponge in the
skin of young pigs. Nature, 223(201), 88-90.
Wise, D. L., Trantolo, D. J., Lewandrowski, K.-U., Gresser, J., & Cattaneo, M. V. (2002).
Biomaterials Engineering and Devices: Human Applications, Volume 2: Orthopedic,
Dental, and Bone Graft Applications. Totowa: Humana Press.
Wright, J. E., & Arthur, R. (1987). Chitin and benzoylphenyl ureas. Boston: W. Junk.
Wu, L. N., Genge, B. R., Dunkelberger, D. G., LeGeros, R. Z., Concannon, B., & Wuthier, R. E.
(1997). Physicochemical Characterization of the Nucleational Core of Matrix Vesicles.
The Journal of Biological Chemistry, 272(7), 4404-4411.
Wu, X., Peters, J. M., Gonzalez, F. J., Prasad, H. S., Rohrer, M. D., & Gimble, J. M. (2000).
Frequency of Stromal Lineage Colony Forming Units in Bone Marrow of Peroxisome
Proliferator-Activated Receptor-alpha-Null Mice. Bone, 26(1), 21-26.
Xia, W., Lindahl, C., Lausmaa, J., & Engqvist, H. (2011). Biomimetic Hydroxyapatite
Deposition on Titanium Oxide Surfaces for Biomedical Application. In M. Cavrak,
Advances in Biomimetics (pp. 429-452). Shanghai: InTech.

252
Xu, A.-W., Ma, Y., & Colfen, H. (2007). Biomimetic mineralization. Journal of Materials
Chemistry, 17(5), 415-449.
Yamasaki, H., & Sakai, H. (1992). Osteogenic response to porous hydroxyapatite ceramics under
the skin of dogs. Biomaterials, 13(5), 308-312.
Yeh-Liang, H., Hung, Y.-C., & Yin, J.-Z. (2006). Design of a Novel Total Knee Prosthesis Using
TRIZ. Journal of Medical and Biological Engineering, 26(4), 177-185.
Yen, M.-T., Yang, J.-H. Y., & Mau, J.-L. (2009). Physicochemical characterization of chitin and
chitosan from crab shells. Carbohydrate Polymers, 75, 15–21.
Yin, X., & Stott, M. J. (2003). Biological calcium phosphates and Posner’s cluste. Journal of
Chemical Physics, 118(8), 3717-3723.
Young, R. (1985). The Ultrastructural organization of proteglycans and collagen in human and
rabbit matrix. J Cell Sci, 74, 95-104.
Yuan, H., & Groot, K. D. (2004). Calcium Phosphate Biomaterials: An Overiview. In R. L. Reis,
& S. Weiner, Learning from Nature How to Design New Implantable Biomaterials (pp.
37-57). Netherlands: Kluwer Academic Publishers.
Yun, Y.-R., Won, J. E., Jeon, E., Lee, S., Kang, W., Jo, H., . . . Kim, H.-W. (2010). Fibroblast
Growth Factors: Biology, Function, and Application for Tissue Regeneration. Journal of
Tissue Engineering, 1, 1-18.
Zhao, J., Liu, Y., Sun, W.-b., & Zhang, H. (2011). Amorphous calcium phosphate and its
application in dentistry. Chemistry Central, 5(40).
Ziats, N. P., Miller, K. M., & Anderson, J. M. (1988). In vitro and in vivo interactions of cells
with biomaterials. Biomaterials, 9, 5-13.