the cell cycle & mitosis 細胞周期與細胞分裂. cell cycle there are so many accelerators...
TRANSCRIPT

The Cell Cycle & Mitosis
細胞周期與細胞分裂


Cell Cycle
There are so many accelerators and brakes.



The Cell Cycle
•During development from stem to fully differentiated, cells in the body alternately divide (mitosis) and "appear" to be resting (interphase). This sequence of activities exhibited by cells is called the cell cycle.
•Interphase, which appears to the eye to be a resting stage between cell divisions, is actually a period of diverse activities. Those interphase activities are indispensible in making the next mitosis possible.


Leland Hartwell used baker's yeast, Saccharomyces cerevisiae, as a model system for genetic studies of the cell cycle. In a series of experiments 1970-71, he isolated yeast cells, in which genes controlling the cell cycle were mutated.
By this approach, he identified genes specifically involved in cell cycle control, CDC genes (cell division cycle genes). One of these genes, designated CDC28, controls the first step in the progression through the
G1 phase of the cell cycle (the function "start").
Hartwell also identified the fundamental role of "checkpoints" in cell cycle control. These checkpoints monitor that all steps in the previous phase have been correctly executed and ensure a correct order between the cell cycle phases.
Leland Hartwell, 1939. Fred Hutchinson Cancer Research Center, Seattle, WA, USA.

.
Leland Hartwell used baker's yeast, Saccharomyces cerevisiae (left). Paul Nurse used another type of yeast, Schizosaccharomyces pombe (middle). Tim Hunt used sea urchin, Arbacia (right).

Paul Nurse identified the key regulator of the cell cycle, the gene cdc2, during the years 1976-80. The product of this gene controls cell division (transition from G2 to M). The gene cdc2 in the fission yeast Schizosaccharomyces pombe had the same function as the gene CDC28 in the baker's yeast.cdc2 controls both the transition from G1 to S and G2 to M. In 1987. He isolated the human gene CDK1. The CDK function has been conserved through evolution.
CDK1 encodes cyclin dependent kinases (CDK). These molecules function by linking phosphate groups to other proteins. Today half a dozen different CDK-mol
ecules have been found in humans.
Paul Nurse,1949. Imperial Cancer Research Fund, Lincoln's Inn Fields, London, UK

CDK and cyclin together form an enzyme that activates other proteins by phosphorylation. The amount of CDK molecules is constant during the cell cycle, but their activities vary because of the regulatory function of the cyclins.
CDK can be compared with an engine and cyclin with a gear box controlling whether the engine will run in the idling state or drive the cell forward in the cell cycle.

Tim Hunt,1943, Imperial Cancer Research Fund, Clare Hall Lab, south Mimms, UK.
Tim Hunt discovered cyclins, proteins that bind to the CDK molecules.
Cyclins regulate the CDK activity and select the target proteins to be phosphorylated. The proteins were named cyclins because of their cyclic variation in amount during the cell cycle.
Cyclins were degraded during mitosis turned out to be another fundamental control mechanism in the cell cycle.
Tim Hunt discovered the first cyclin molecule in 1982, using eggs from sea urchin, Arbacia, as a model system. He also found that cyclins, like CDK, were conserved during evolution. Today around ten different cyclins have been found in humans.

The fundamental molecular mechanisms controlling the cell cycle are highly conserved through evolution and operate in the same manner in yeasts, insects, plants, animals and humans.
Cyclins are proteins formed and degraded during each cell cycle. Periodic protein degradation is an important control mechanism of the cell cycle. (D = cell division.)

Chromosomal instability in cancer cells may be the result of defective cell cycle control. Three pairs of chromosomes (1, 3 and 14) in normal cells, compared with the cancer cells. In cancer cells, the chromosome number may be altered (aneuploidy) and parts of chromosomes may be rearranged



染色體 - genetic information in the form of chromatin, highly folded ribbon-like complexes of deoxyribonucleic acid (DNA) and a class of proteins called histones.

When a cell divides, chromatin fibers are very highly folded, and become visible in the light microscope as chromosomes.
During interphase (between divisions), chromatin is more extended, a form used for expression genetic information.


The DNA of chromatin is wrapped around a complex of histones appears in the electron microscope as "beads on a string" or nucleosomes. Changes in folding between chromatin and the mitotic chromosomes is controlled by the packing of the nucleosome complexes.
.
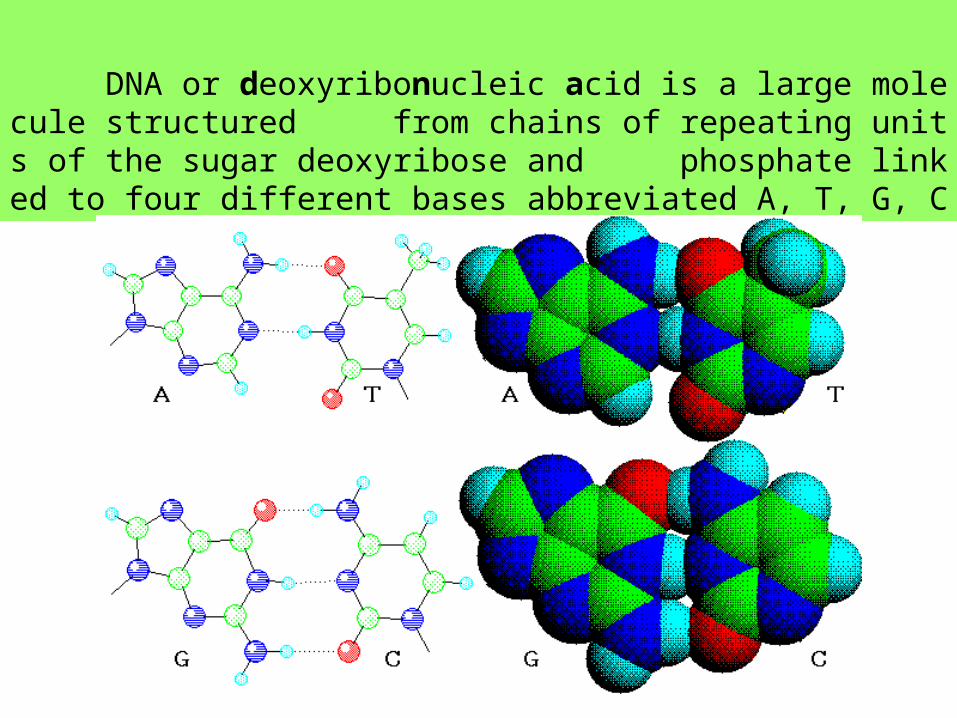
DNA or deoxyribonucleic acid is a large molecule structured from chains of repeating units of the sugar deoxyribose and phosphate linked to four different bases abbreviated A, T, G, C

The Cell Cycle
•Stages of the cell cycle The cell cycle is an ordered set of events, culminating in cell growth and division into two daughter cells. Non-dividing cells not considered to be in the cell cycle.
•The G1 stage stands for "GAP 1".
•The S stage stands for "Synthesis". This is the stage when DNA replication occurs.
•The G2 stage stands for "GAP 2".
•The M stage stands for "mitosis", and is when nuclear (chromosomes separate) and cytoplasmic (cytokinesis) division occur.
Mitosis is further divided into 4 phases

Eukaryotic Cell Cycle

Regulation of the cell cycle How cell division (and thus tissue growth) is controlled is very complex.
•Cdk (cyclin dependent kinase, adds phosphate to a protein), along with cyclins, are major control switches for the cell cycle, causing the cell to move from G1 to S or G2 to M.
•MPF (Maturation Promoting Factor) includes the CdK and cyclins that triggers progression through the cell cycle.
•p53 is a protein that functions to block the cell cycle if the DNA is damaged. If the damage is severe this protein can cause apoptosis (cell death).

p21

1. p53 levels are increased in damaged cells. This allows time to repair DNA by blocking the cell cycle.
2. A p53 mutation is the most frequent mutation leading to cancer. An extreme case of this is Li Fraumeni syndrome, where a genetic a defect in p53 leads to a high frequency of cancer in affected individuals.
3. p53 protein binds DNA and stimulates another gene to produce a p21 that interacts with cdk2. Stop the cell cycle.
4. p27 binds to cyclin and CdK blocking entry into S phase. Breast cancer prognosis is determined by p27 levels. Reduced levels of p27 predict a poor outcome.

Cyclins
•a G1 cyclin (cyclin D)
•S-phase cyclins (cyclins E & A)
•mitotic cyclins (cyclins B & A)
Their levels in the cell rise and fall with the stages of the cell cycle.
Cyclin-dependent kinases (Cdks)
• G1 Cdk (Cdk4)
• S-phase Cdk (Cdk2)
• M-phase Cdk (Cdk1) Their levels in the cell remain fairly stable, but each must bind the appropriate cyclin in order to be activated. They add phosphate groups to a variety of protein substrates that control processes in the cell cycle.


Cdk
cyclin

The anaphase-promoting complex (APC) •The APC is also called the cyclosome, and the complex is often designated as the APC/C.
The APC/C •triggers the events leading to destruction of the cohesins thus allowing the sister chromatids to separate
•degrades the mitotic cyclin B.

Steps in the cycle
•A rising level of G1-cyclins bind to their Cdks and signal the cell to pre
pare the chromosomes for replication.
•A rising level of S-phase promoting factor (SPF) — which includes cyclin A bound to Cdk2 — enters the nucleus and prepares the cell to duplicate its DNA (and its centrosomes).
•As DNA replication continues, cyclin E is destroyed, and the level of mitotic cyclins begins to rise (in G2).

One example of a well-known transcription factor activation is the Rb/E2F protein regulation of the G1 to S phase transition in mammals cells
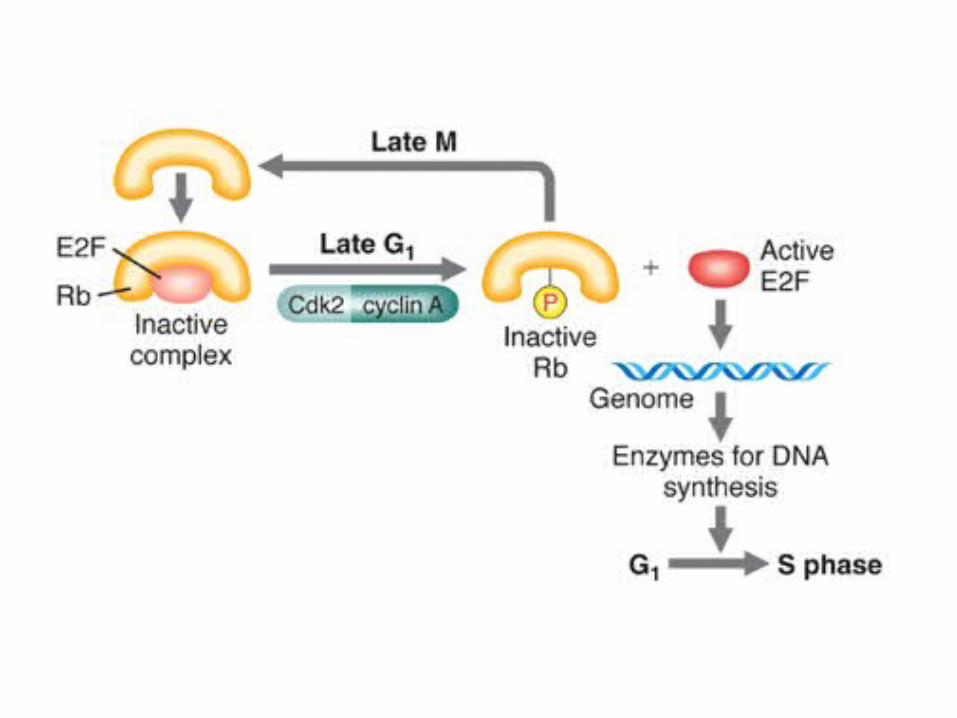

M-phase promoting factor (the complex of mitotic cyclins with the M-phase Cdk) initiates
• assembly of the mitotic spindle
•breakdown of the nuclear envelope
•condensation of the chromosomes
These events take the cell to metaphase of mitosis.
At this point, the M-phase promoting factor activates the anaphase-promoting complex (APC/C)

APC/C
•allows the sister chromatids at the metaphase plate to separate and move to the poles (= anaphase), completing mitosis;
•destroys cyclin B. It does this by attaching it to the protein ubiquitin which targets it for destruction by proteasomes.
• turns on synthesis of G1 cyclin for the next turn of the cycle;
•degrades geminin, a protein that has kept the freshly-synthesized DNA in S phase from being re-replicated before mitosis.
This is only one mechanism by which the cell ensures that every portion of its genome is copied once — and only once —during S phase.

Checkpoints: Quality Control of the Cell Cycle
The cell has several systems for interrupting the cell cycle if something goes wrong.
A check on completion of S phase. The cell seems to monitor the presence of the Okazaki fragments on the lagging strand during DNA replication. The cell is not permitted to proceed in the cell cycle until these have disappeared.
DNA damage checkpoints. These sense DNA damage
•before the cell enters S phase (a G1 checkpoint);
•during S phase, and
•after DNA replication (a G2 checkpoint).
Spindle checkpoints
•detect any failure of spindle fibers to attach to kinetochores and arrest the cell in metaphase (M checkpoint);
•detect improper alignment of the spindle itself and block cytokinesis
•trigger apoptosis if the damage is irreparable


Cancer

p53
•The p53 protein senses DNA damage and can halt progression of the cell cycle in G1.
•Both copies of the p53 gene must be mutated for this to fail so mutations in p53 are recessive, and p53 qualifies as a tumor suppressor gene.
•.The p53 protein is also a key player in apoptosis, forcing "bad" cells to commit suicide.
•Mutant p53 develops into a cancer.
•More than half of all human cancers harbor p53 mutations and have no functioning p53 protein.


The p53 gene is found in chromosome 17.
In the cell, p53 protein binds DNA and stimulates p21 that interacts
with cdk2.
When p21 is complexed with cdk2 the cell cannot pass through to the next stage of cell division.
Mutant p53 can no longer bind DNA in an effective way, and the p21 protein is not made available to act as the 'stop signal' for cell division.
Thus cells divide uncontrollably, and form tumors.


A genetically-engineered adenovirus, called ONYX-015, can only replicate in human cells lacking p53.
Thus it infects, replicates, and ultimately kills many types of cancer cells in vitro.
Clinical trials are now proceeding to see if injections of ONYX-015 can shrink a variety of types of cancers in human patients.

p53 seems to evaluate the extent of damage to DNA, at least for damage by radiation.
•At low levels of radiation, producing damage that can be repaired, p53 triggers arrest of the cell cycle until the damage is repaired.
•At high levels of radiation, producing hopelessly damaged DNA, p53 triggers apoptosis. Possible mechanism:
•Serious damage, e.g., double-stranded breaks (DSBs), causes a linker histone (H1) to be released from the chromatin.
H1 leaves the nucleus and enters the cytosol where it triggers the release of cytochrome c from mitochondria leading to apoptosis


ATM
ATM (ataxia telangiectasia mutated) gets its name from a human disease of that name, whose patients — among other things — are at increased risk of cancer.
The ATM protein is involved in
•detecting DNA damage, especially double-strand breaks
•interrupting (with the aid of p53) the cell cycle when damage is found;
•maintaining normal telomere length

MADMAD (mitotic arrest deficient) genes (there are two) encode proteins that bind to each kinetochore until a spindle fiber (one microtubule will do) attaches to it. If there is any failure to attach, MAD remains and blocks entry into anaphase.
Mutations in MAD produce a defective protein and failure of the checkpoint. The cell finishes mitosis but produces daughter cells with too many or too few chromosomes (aneuploidy).
Aneuploidy is one of the hallmarks of cancer cells suggesting that failure of the spindle checkpoint is a major step in the conversion of a normal cell into a cancerous one.



Infection with the human T cell leukemia virus-1 (HTLV-1) leads to a cancer (ATL = adult T cell leukemia) in about 5% of its victims.
HTLV-1 encodes a protein, called Tax, that binds to MAD protein causing failure of the spindle checkpoint.
The leukemic cells in these patients show many chromosome abnormalities including aneuploidy

G0
Many times a cell will leave the cell cycle, temporarily or permanently.
It exits the cycle at G1 and enters a stage designated G0 (G zero).
A G0 cell is often called "quiescent", but that is probably more a
reflection of the interests of the scientists studying the cell cycle than the cell itself.
Many G0 cells are anything but quiescent. They are busy carrying
out their functions in the organism. e.g., secretion, attacking pathogens.
Often G0 cells are terminally differentiated: they will never reent
er the cell cycle but instead will carry out their function in the organism until they die.


For other cells, G0 can be followed by reentry into the cell cycle.
Most of the lymphocytes in human blood are in G0. However, with prope
r stimulation, such as encountering the appropriate antigen, they can be stimulated to reenter the cell cycle (at G1) and proceed on to new rounds o
f alternating S phases and mitosis.
G0 represents not simply the absence of signals for mitosis but an active r
epression of the genes needed for mitosis.
Cancer cells cannot enter G0 and are destined to repeat the cell cycle inde
finitely


Cell Cycle Pathway


Retinoblastoma

The retinoblastoma gene was isolated in 1986. It was the first tumor suppressor gene that was isolated based on knowledge of its chromosomal location: chrom 13 band q14. Germ line mutations in the Rb gene, predispose to a pediatric malignancy of the eye: retinoblastoma.
Loss of Rb predisposes to a variety of other tumors later in life, with osteosarcoma being the most prominent secondary tumor.
Loss of Rb function: lung cancer, lymphoma & breast cancer.
The Rb gene encodes a 110 kDa phosphoprotein (pRb) that is expressed in almost every cell of the human body and contributes to growth regulation in these cells. Reintroduction of a functional Rb gene in retinoblastoma tumor cells results in growth arrest, indicating that the function of the gene is to restrict proliferation.

ATM acts as a tumor suppressor. ATM activation, via IR damage to DNA, stimulates DNA repair and blocks cell cycle progression. p53 can cause growth arrest of the cell at a checkpoint to allow for DNA damage repair or can cause the cell to undergo apoptosis. p53 is mutated in over 50% of cancers.


•The p53 tumor suppressor protein can bind to specific DNA elements and in free from can activate transcription of genes that harbor p53 response elements in their promoters.
•The activity of p53 is counteracted by a protein called Mdm2. •Mdm2 can bind to the transactivation domain of p53 and thereby prevent transactivation by p53. In addition, Mdm2 acts as a “ubiquitin ligase” that can target p53 for degradation by the proteasome. Thus, p53 in complex with Mdm2 is both inactive and unstable.
•Binding of Mdm2 to p53 can be disrupted by a protein named p19ARF (or ARF for short). ARF can bind Mdm2 which renders it unable to interact with p53. •Expression of ARF therefore activates p53 by releasing Mdm2.





Proliferation vs. Apoptosis Controls
•These are interrelated, and may induce apoptosis in cells that fail to successfully complete some phase of cell cycle.
•Intracellular signals •cell cycle negative controls: inhibition of CDK-cyclin •cell cycle positive controls: activation of CDK-cyclin
•MITOGENS are polypeptide ligands•Some of these are growth factors that activate receptor tyrosine kinases (RTK proteins). This initiates a signal cascade that affects the configuration of many different transcription factors, affecting the gene activity in the cell
•apoptosis positive controls: leakage of cytochrome c from defective mitochondria acts as a trigger for apoptosis •apoptosis negative controls: proteins such as Bcl-2 & Bcl-x block the release of cytochrome c from mitochondria, possibly stabilizing the mitochondrial membrane and preventing its rupture).
•Extracellular signals •based on cell-cell communication





節食減少代謝 使 NAD, Sir2 增加


What is (and is not) mitosis?
Mitosis is nuclear division plus cytokinesis, and produces two identical daughter cells during prophase, prometaphase, meta
phase, anaphase, and telophase.
Interphase is often included in discussions of mitosis, but interphase is technically not part of mitosis, but rather encompasses stages G1, S, and G2 of the cell cycle.

The cell is engaged in metabolic activity and performing its prepare for mitosis. Chromosomes are not clearly discerned in the nucleus, although a dark spot called the nucleolus may be visible. The cell may contain a pair of centrioles both of which are organizational sites for microtubules.
Interphase

Chromatin in the nucleus begins to condense and becomes visible in the light microscope as chromosomes. The nucleolus disappears. Centrioles begin moving to opposite ends of the cell and fibers extend from the centromeres. Some fibers cross the cell to form the mitotic spindle.
Prophase

The nuclear membrane dissolves, marking the beginning of prometaphase. Proteins attach to the centromeres creating the kinetochores. Microtubules attach at the kinetochores and the chromosomes begin moving.
Prometaphase

Spindle fibers align the chromosomes along the middle of the cell nucleus. This line is referred to as the metaphase plate. This organization helps to ensure that in the next phase, when the chromosomes are separated, each new nucleus will receive one copy of each chromosome.
Metaphase

The paired chromosomes separate at the kinetochores and move to opposite sides of the cell. Motion results from a combination of kinetochore movement along the spindle microtubules and through the physical interaction of polar microtubules.
Anaphase

Chromatids arrive at opposite poles of cell, and new membranes form around the daughter nuclei. The chromosomes disperse and are no longer visible under the light microscope. The spindle fibers disperse, and cytokinesis or the partitioning of the cell may also begin during this stage.
Telophase

In animal cells, cytokinesis results when a fiber ring composed of a protein called actin around the center of the cell contracts pinching the cell into two daughter cells, each with one nucleus. In plant cells, the rigid wall requires that a cell plate be synthesized between the two daughter cells.
Cytokinesis



