the chitin crystallite in arthropod cuticlejcs.biologists.org/content/joces/21/1/73.full.pdf · the...
TRANSCRIPT

J. Cell Set. ai, 73-82 (1976) 73Printed in Great Britain
THE CHITIN CRYSTALLITE IN ARTHROPOD
CUTICLE
A. C.NEVILLEDepartment of Zoology, Woodland Road, Bristol, England
D. A. D. PARRYDepartment of Chemistry, Biochemistry and Biophysics, Massey University,Palmerston North, New Zealand
AND J. WOODHEAD-GALLOWAYMedical Research Council, 20 Park Crescent,London WiN 4AL, England
SUMMARYElectron microscopy has revealed that chitin from a representative selection of insect orders
(plus one crustacean and one arachnid) is localized in crystallites about 2-8 nm across. Further-more, these crystallites are arranged on an hexagonal or pseudo-hexagonal lattice, the lateralorder of which varies considerably. The lattice becomes secondarily reoriented during cuticleexpansion following an ecdysis. The size of the 'unit cell' has been measured both by opticaldiffraction and direct measurements of the micrographs, permitting an estimate of the chitinand resilin content for locust rubberlike cuticle. The number of poly-iV-acetyl-glucosaminechains per sheet and sheets per crystallite can be estimated from the physical dimensions of thecrystallite. Each crystallite is unlikely to comprise more than 3 sheets and 6 chains per sheet. Thecalculated and measured density of a-chitin can be shown to be in close agreement.
INTRODUCTION
Arthropod cuticle is a composite material like fibre-glass, with crystallites or micro-fibrils dispersed in a matrix. In suitably stained electron-microscope sections, it ispossible to resolve the crystallites with particular clarity because their edges arereasonably well defined. It will be shown that the diameter of such crystallites isapproximately 2-8 nm. By contrast it may be noted that plant biophysicists and poly-mer morphologists have difficulty in distinguishing between elementary protofibrilsof similar size and the 15-0-25-0 nm diameter bundles which they form in theirrespective materials (Sarko & Marchessault, 1969; Nieduszynski & Preston, 1970).Some workers prefer to recognize a continuous spectrum of diameters ranging fromthe individual molecular chain through to 100-nm diameter bundles. In arthropodcuticle, Rudall (1965) first illustrated crystallites in electron micrographs of sectionsof Sirex ovipositors. These have subsequently been shown to be arranged in manyinsect cuticles in a helicoid (Neville & Luke, 1969). Each layer of crystallites isarranged in parallel, with successive layers turning progressively through a smallangle in an anti-clockwise direction in layers further away from the observer (Neville& Luke, 1971). In electron-microscope sections of such helicoidal cuticle the

74 A. C. Neville, D. A. D. Parry andj. Woodhead-Galloway
component planes of crystallites can be seen to rotate, appearing alternately intransverse or longitudinal profile with intervening oblique ones. This clearlyestablishes that crystallites in cuticle are not artifacts.
With regard to the chemical composition of cuticle crystallites and matrix, it hasbeen pointed out that this may be demonstrated in a simple qualitative way (Neville,1970). Electron micrographs of pure resilin samples may be compared with those ofrubberlike cuticle in which resilin and chitin are known to be the only 2 componentspresent. With potassium permanganate staining pure resilin is uniformly electrondense; by contrast, in the mixed resilin and chitin system the matrix is electron dense,whereas the crystallites remain unstained because they are impenetrable. Hence, bysubtraction, chitin crystallites are embedded in a protein matrix. It is possible todemonstrate this more convincingly in a quantitative way as we show below, bymaking measurements of volume fractions of crystallites and matrix from electronmicrographs of locust rubberlike cuticle. These gave reasonable agreement withchemical measurements of chitin and resilin content. Furthermore, in sections of thesame material which had been partly digested with subtilisin at neutral pH crystalliteswere clearest in the digested region but could also be traced into the intact regionwhere staining of the protein matrix was densest (Weis-Fogh, 1970).
We may safely conclude that rubberlike cuticle consists of chitin crystallites em-bedded in a resilin matrix and by analogy we may interpret similar crystallites inordinary solid cuticles as chitin also. In support of the latter is the presence ofswollen 7-5-11111 diameter crystallites in preparations of deproteinized cuticle whichwere colloidally dispersed in lithium thiocyanate, sonicated, centrifuged and visualizedby negative staining in the electron microscope (Neville, 1975).
The purpose of the present paper is to report measurements of the size and dis-position of the chitin crystaUites from 8 orders of insects, plus one crustacean and onearachnid, by electron microscopy and optical diffraction. This permits an estimate ofthe number of molecular chains of poly-JV-acetylglucosamine per sheet and of sheetsper crystallite. Furthermore, we have attempted to understand the packing of theunits as related to the stage of development of the exocuticle.
METHODS
Electron microscopy
Material was fixed for 2 h in 25 % glutaraldehyde in 005 M cacodylate buffer at pH 72 at4 °C. It was washed in several changes of buffer at 4 °C for 05 h, then post-osmicated in 1 %aqueous osmium tetroxide for 1 h. After dehydration and embedding in Araldite, thin sectionswere cut and stained in 2 % aqueous potassium permanganate. This stains the protein matrixbut is unable to penetrate the chitin crystallite, so contrast between the 2 phases is improved.
Optical diffraction
Periodicities in the electron micrographs were found by using the latter as a diffraction gratingon the Polaron Optical Diffractometer. The helium-neon laser source generated 1 mW of power(wavelength 6328 nm). Exposures varied but were usually about 1 s at the camera distance used.The optical diffraction patterns were recorded on Kodalith Pan Film 2568 and were developedand fixed in the recommended manner.

Chitin crystallite in arthropod cuticle 75
X-ray diffraction
X-ray diffraction patterns were taken on a Guinier focusing camera of specimen to film distanceup to 60 cm. The Elliott rotating anode X-ray generator was run at 50 kV x 50 mA. Exposureswere recorded with Ilfex Industrial G X-ray film on a flat plate holder.
RESULTS
Crystallite sizeChitin-containing samples from 8 orders of insects, one crustacean and one arachnid
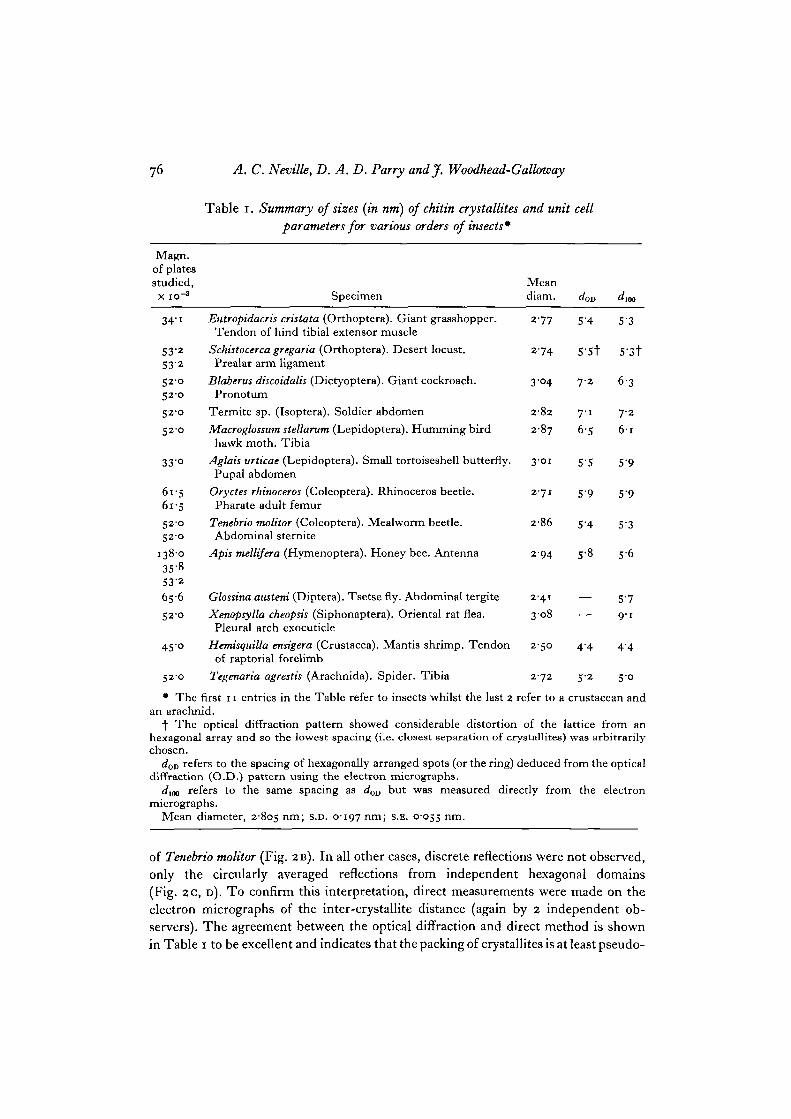
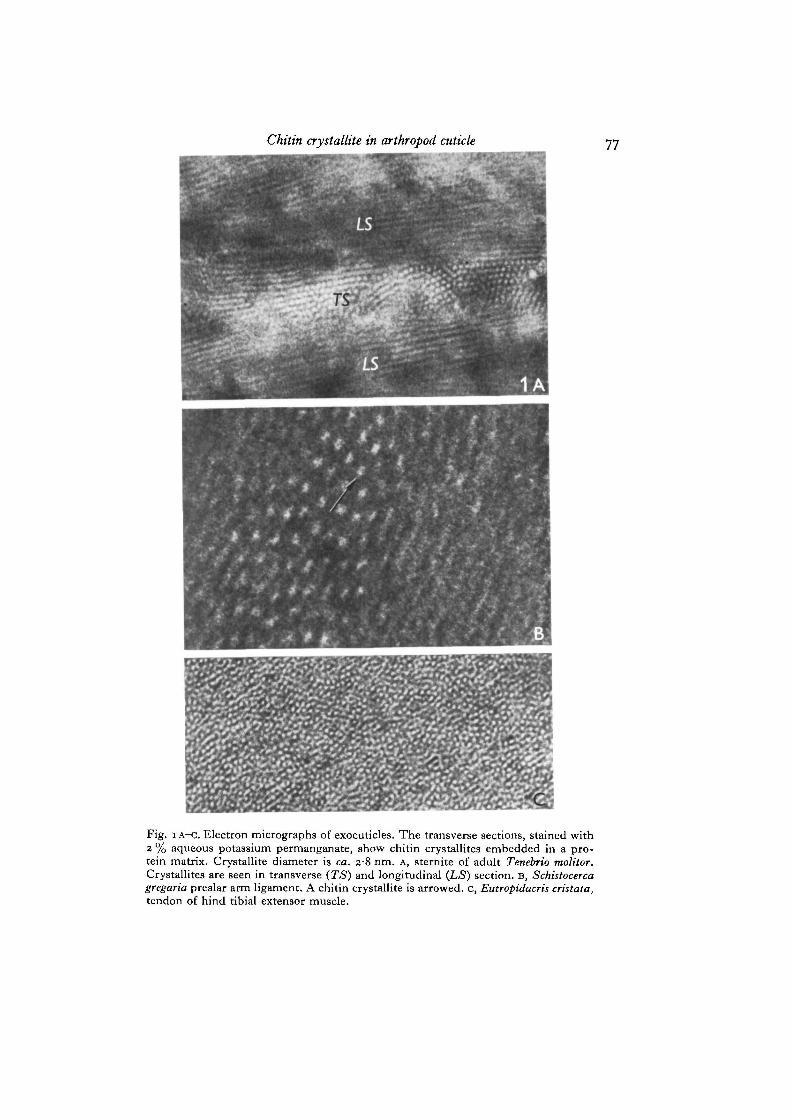
have been studied by electron microscopy and optical diffraction techniques. Theresults obtained are summarized in Table 1. The mean diameter of the crystallitesseen in the electron micrographs (Fig. 1) was measured directly using a Nikon ProfileProjector. Each electron micrograph was measured independently by 2 observers. Theresults were averaged, as no significant differences were noticed between the sets ofmeasurements. Over 600 values of the crystallite diameter were obtained, the extremevalues being 2-3 and 3-5 nm. The spread of mean diameters recorded in Table 1 issmall, bearing in mind the difficulty of measuring units of this size and also thepossible error (as much as 5-10%) in the electron-microscope magnification. Theinherent errors of the measurements are sufficient to disguise any real but smalldifferences in the diameters of chitin crystallites for these thirteen specimens. Wemay therefore consider that the diameter of the crystallites is constant and has a valueclose to 2-8 nm. If we assume that all the chitin-containing specimens studied docontain crystallites of constant diameter then there is about 1 chance in 1000 that thetrue diameter lies outside the range 2-8 + 0-25 nm.
Crystallite lattice
Studying the electron micrographs on the optical diffractometer, either 6 hexagon-ally arranged spots (Fig. 2 A, B) or a ring of density (Fig. 2 c, D) were observed. Thelateral width of the ring and the spread of the spots varied considerably betweendifferent specimens and illustrated the differing extent of the lateral range of orderin the hexagonally packed array. The radial distance of either the spots or the ringfrom the centre is related to the mean separation of the crystallites. Visual inspectionof the electron micrographs confirmed that the crystallites were packed in hexagonalarray but the extent and degree of perfection of the lattice varied considerably. Thosespecimens which showed the least distortion, i.e. the clearest example of an hexagonalpattern, were from the Oryctes rhinoceros pharate adult femur (Fig. 2 A). In this case,the pharate adult beetle had been sectioned before it had emerged from the pupa andhence would not be expected to have the exocuticle deformed by expansion of thebody, as occurs at ecdysis. This beetle is unusual in that it tans parts of the exocuticlefor up to 3 days before ecdysis (Zelazny & Neville, 1972). Tanned cuticle takes upstain heavily, giving improved contrast between crystallites and matrix. Evidence ofhexagonality in the optical-diffraction patterns was obtained from some specimens ofApis mellifera antenna, the abdomen of the soldier termite and the abdominal sternite
76 A. C. Neville, D. A. D. Parry andjf. Woodhead-Gallmoay
Table i. Summary of sizes (in nm) of chitin crystallites and unit cellparameters for various orders of insects*
Magn.of platesstudied,
x io"3 Specimen
34'i Eutropidacris cristata (Orthoptera). Giant grasshopper.Tendon of hind tibial extensor muscle
53-2 Schistocerca gregaria (Orthoptera). Desert locust.532 Prealar arm ligament
520 Blaberus discoidalis (Dictyoptera). Giant cockroach.52'O Pronotum
52'O Termite sp. (Isoptera). Soldier abdomen
520 Macroglossum stellarum (Lepidoptera). Humming birdhawk moth. Tibia
33 o Aglais urticae (Lepidoptera). Small tortoiseshell butterfly.Pupal abdomen
61-5 Oryctes rhinoceros (Coleoptera). Rhinoceros beetle.61-5 Pharate adult femur
520 Tenebrio molitor (Coleoptera). Mealworm beetle.520 Abdominal sternite
138-0 Apis mellifera (Hymenoptera). Honey bee. Antenna35-8532656 Glossina austeni (Diptera). Tsetse fly. Abdominal tergite
52-0 Xenopsylla cheopsis (Siphonaptera). Oriental rat flea.Pleural arch exocuticle
450 Hemisquilla ensigera (Crustacea). Mantis shrimp. Tendon 250 4 4 44of raptorial forelimb
520 Tegenaria agrestis (Arachnida). Spider. Tibia 272 52 5-0
• The first 11 entries in the Table refer to insects whilst the last 2 refer to a crustacean andan arachnid.
f The optical diffraction pattern showed considerable distortion of the lattice from anhexagonal array and so the lowest spacing (i.e. closest separation of crystallites) was arbitrarilychosen.
dOv refers to the spacing of hexagonally arranged spots (or the ring) deduced from the opticaldiffraction (O.D.) pattern using the electron micrographs.
rf]00 refers to the same spacing as d0D but was measured directly from the electronmicrographs.
Mean diameter, 2805 nm; s.D. 0197 nm; s.E. 0055 nm.
of Tenebrio molitor (Fig. 2 B). In all other cases, discrete reflections were not observed,only the circularly averaged reflections from independent hexagonal domains(Fig. 2 c, D). TO confirm this interpretation, direct measurements were made on theelectron micrographs of the inter-crystallite distance (again by 2 independent ob-servers). The agreement between the optical diffraction and direct method is shownin Table 1 to be excellent and indicates that the packing of crystallites is at least pseudo-
Meandiam.
2-77
2 7 4
3-04
2 8 2
2-87
3 0 1
2 7 1
2 8 6
2 9 4
2 4 1
3 0 8
doo
5-4
s-st
7-2
7 ' i
6-5
5'5
5 9
5'4
5 8
.—
^100
5 3
5-3t
6 3
7-2
6 1
5 9
5 9
5'3
56
5 79 1
Chitin crystallite in arthropod cuticle 77
Fig. i A-C. Electron micrographs of exocuticles. The transverse sections, stained with2 % aqueous potassium permanganate, show chitin crystallites embedded in a pro-tein matrix. Crystallite diameter is ca. 2-8 nm. A, sternite of adult Tenebrio molitor.Crystallites are seen in transverse (TS) and longitudinal (LS) section, B, Schistocercagregaria prealar arm ligament. A chitin crystallite is arrowed, c, Eutropidacris cristata,tendon of hind tibial extensor muscle.

A. C. Neville, D. A. D. Parry andjf. Woodhead-Galloway
PFig. 2A-D. Optical diffraction patterns from the electron micrographs showing thechitin crystallites in exocuticle. A, Oryctes rhinoceros, pharate adult femur. Sixhexagonally arranged spots may be seen showing the near-perfect hexagonal packingof the chitin crystallites, B, Tenebrio molitor, abdominal sternite. A ring of densitycan be seen, the spacing of which corresponds to the d100 spacing of an hexagonalarray. Evidence of the ring of density being broken into 6 maxima can be seen lessclearly than in Fig. 2 A. C, Apis mellifera, antenna. The ring of density as in B corre-sponds to dlm of an hexagonal array but the orientation of hexagonal unit cells variesin a random way. D, Eutropidacris cristata, as in c.

Chitin crystallite in arthropod cuticle 79
hexagonal on a nearest-neighbour basis. Some specimens showed an extremely dis-torted lattice, e.g. Schistocerca gregaria. In this case, the hexagonal lattice had in someplaces been greatly deformed. Only very small areas of crystallites could be seen inthese electron micrographs. In all these cases which have been studied by electronmicroscopy, the exocuticles were secreted before an ecdysis and therefore distortedduring expansion at ecdysis. Endocuticle which is secreted after ecdysis is not sub-jected to expansion distortion and one would expect considerably more order in thechitin structure at this level. However, exocuticles appear to have chitin which is morecrystalline than the endocuticles on the basis of the electron micrographs whichshow a clearer phase separation between chitin and the protein matrix.
Low angle X-ray patterns of Eutropidacris cristata hind leg tendons showedequatorial reflections at spacings of 5-9 and 3"3~3"4 nm. These 2 reflections index asthe first 2 orders of a hexagonal array of lattice side about 6-75 nm which is larger(by 10%) than the lattice spacing observed in electron-microscope sections. Thismight be explained by shrinkage of the specimen during preparation, although it mustbe remembered that the X-ray pattern is derived from both exo- and endocuticle.The reflections are broad, which means that there is difficulty in making the measure-ments very precisely. Rudall observed spacings of 6-3 and 3-5 nm for ovipositor wall(Megarhyssa), 3-3 nm for lobster cuticle and 3-7 nm for stretched blowfly larvalcuticle.
Fraction of chitin present
Rubberlike cuticle presents an opportunity to measure volume fractions of crystal-lites and matrix, for comparison with chemical measurements of resilin and chitincontent, since these are the only 2 components present. For this, selected areas werechosen on electron micrographs of locust prealar arm resilin in which the crystallitesare distributed in an approximately hexagonal lattice.
From the measurements of the unit cell and the diameter of the crystallite, it ispossible to calculate the percentage of chitin present in the samples of rubberlikecuticle. The exact amount of chitin will depend on 3 assumptions: (1) the shape of thecrystallite in cross-section - round, square or rectangular; (2) the degree of shrinkageof the specimen during preparation; and (3) the amount of chitin which is 'dis-organized' and outside the periphery of the observed crystallite. In this latter case(Neville, 1970, 1975), it seems likely that little chitin is found outside the crystallite,though the possibility that some may be present cannot be completely eliminated.
If s2 is the cross-sectional area and d the centre-to-centre spacing of thecrystallites, then the volume fraction of crystallites is s2j{(KSJT)J2)d2 — s2) (assumingcrystallites of square cross-section). In Table i, 2 measurements are given which areused for calculating d\ one is obtained directly from the electron micrographs (d100)and the other from optical diffraction (d0T)). Then let A be the product of crystallitevolume fraction and density of dry chitin (1-425 g cm"3: Carlstrom, 1957); and let Bbe the product of matrix volume fraction and density of dry resilin (1-3 gem"3:Weis-Fogh, 1961). Then AjA + B may be compared with the chitin and resilin contentobtained from chemical analysis (Neville, 1963). We have used values measured from

80 A. C. Neville, D. A. D. Parry andj. Woodhead-Gallotoay
electron micrographs oi s = 2-8 nm and d = 5-4 nm x I-I x 2/̂ /3 (allowing for about10% shrinkage of the matrix, which is indicated by the reduced X-ray value forintercrystallite spacing reported in the results above). This gives 20-6% chitin and79-4% resilin for dry prealar arm rubberlike cuticle of locusts (Schistocerca gregaria),in reasonable agreement with direct measurement before and after acid hydrolysis,which showed that the proportion of chitin increases from 13 to 24% of the totalvacuum dry weights of samples measured between 2 days prior to adult ecdysis and20 days after it (Neville, 1963). If, on the other hand, it is assumed that the crystallitesare circular in cross-section then a lower estimate for chitin content would result.Table 1 shows that since crystallite size is relatively constant, chitin content wouldbe expected to vary with lattice spacing in the range of species measured.
Crystallite structure
The physical dimensions of the crystallite enable us to estimate the maximumnumber of poly-iV-acetyl-glucosamine chains per sheet and sheets per crystallite.Arnott, Dover & Neville (unpublished work) have shown from the high-angle X-raydiffraction pattern of chemically untreated a-chitin from Eutropidacris cristata thatthe unit cell has axes {a, b and c) of dimensions 1-884 x 0*476 x 1-028 nm. The i-axiscorresponds to the hydrogen bond distance between similarly directed chains, whilstthe a-axis corresponds to the intersheet period (i.e. between nearest sheets of thesame direction). It follows from the X-ray measurements that there are 4 monomersper unit cell, i.e. mean volume per residue is 0-23 nm3. Using Partington's method ofadding the volumes of the atomic components (based on data from proteins in water),the poly-iV-acetyl-glucosamine (of residue weight 203) would have a volume of about0-234 nm3. This confirms Meyer & Pankow's (1935) observation that there are 4 resi-dues per unit cell. It also confirms that Partington's method works for polysaccharides.Assuming that the crystallite is square in cross-section, there could be up to 3 sheets(3x0-942 = 2-826 nm) and 6 chains per sheet (6x0-476 = 2-856 nm) within themeasured boundaries. The volume of a block 0-514 nm high (the axial height per unit)and 2-8 by 2-8 nm is 4-030 nm3 and taking the mean volume per unit as approximately0-230 nm3, there could be as many as 18 chains passing through a crystallite at anyone level. This result is in reasonable agreement with the model proposed by Rudall& Kenchington (1973) on the basis of X-ray diffraction studies. Their model consistsof 3 sheets, each of 7 chains.
DISCUSSION
Examples in which fibres are embedded in a matrix are known from both organicand synthetic systems. Chitin and collagen perform similar structural roles in animals.Collagen often exists in vertebrates as a complex of ordered collagen (protein) mole-cules in a matrix of water and polysaccharide, for example as cartilage. On the otherhand, chitin consists of poly-iV-acetyl-glucosamine (polysaccharide) chains whichaggregate into crystallites, these crystallites being embedded in a matrix of water andprotein. It has been shown by Rudall (1963, 1965) that chitin units may be seen in the

Chitin crystallite in arthropod cuticle 81
electron microscope from transverse sections. We have measured the size of theseunits from 8 orders of insects plus one crustacean and one arachnid. Furthermore,we have attempted to understand the packing of the units as related to the stage ofdevelopment of the exocuticle. The originally hexagonal lattice of crystallites becomesdistorted during expansion of the exocuticle following an ecdysis.
The uniformity of diameter of chitin crystallites in a wide range of samples is aproperty which is shared with keratin microfibrils (Fraser, MacRae & Rogers, 1962).Such uniformity is also found in synthetic systems (e.g. Thorium UO2; Gerdes,Chapman & Clark, 1970). The latter system also demonstrates that precise hexagonallattices such as are found in undistorted cuticle can arise solely by crystallization. Theconcentration of chitin crystallites is low compared with that in a system with close-packed rods in a matrix. There appears to be a phase separation between the chitincrystallites and the protein matrix. Examples are known from both organic and syn-thetic systems in which different regions of a chain molecule form fibres and matrix,so as to mimic the mechanical properties of 2-phase systems (e.g. partially denaturedfeather keratin: Fraser, MacRae, Parry & Suzuki, 19716; a-keratin: Fraser et al.1971a; and polystyrene-polybutadiene-polystyrene 3-block copolymer: Keller et al.1971). For cuticle to behave as a composite material it would be expected that thechitin crystallites are strongly bonded to the protein matrix, and there is somechemical evidence for this (Hackman, i960). Such a composite model for cuticlereplaces the old model of Fraenkel & Rudall (1947), who suggested that monomole-cular sheets of chitin and protein occurred in alternation. The latter model has alreadybeen criticized by Jensen & Weis-Fogh (1962) on the grounds that it would be toobrittle.
We prefer to use the term crystallite for chitin systems rather than miicrofibril.There are examples of microfibrils in biological systems, e.g. a-keratin (Fraser,MacRae & Miller, 1965), feather keratin (Fraser et al. 1971 b) and it has been suggestedseveral times that collagen exists as microfibrils. In all these cases the microfibrilappears to have either a rotation or screw axis coincident with its own axis relatingthe molecules or molecular aggregates to one another. In each of these cases, the X-raydiffraction pattern shows evidence of near equatorial reflections characteristic of asupercoiling in the molecules and/or molecular aggregate. In the case of a-chitin, thehigh-angle X-ray pattern shows no such evidence of satellite layer lines appearingclose to the equator and other main layer lines. It can be concluded that the axes ofthe chains of ./V-acetyl-glucosamine are straight.
Rudall (1963) has suggested (from unit cell measurements) that the density ofa-chitin is not consistent with the calculated values. We find no such difficulty. Themeasured density of dehydrated a-chitin is 1-400-1-425 g cm"3. The calculated densityis given by i-66 x 203/230-5 = 1-462 g cm"3. This is a discrepancy of the same orderas that between the calculated and measured density of fibrous proteins and syntheticpolypeptides.
We want to acknowledge the skilful assistance of Mrs B. M. Luke with the electron micro-scope, Mr D. J. S. Hulmes with the optical diffraction and Mr C. D. Rodger with the X-raystudy. The Agricultural Research Council provided full finance for one of us (A.C.N.).

82 A. C. Neville, D. A. D. Parry andj. Woodhead-Galloway
The research is part of the programme of the M.R.C. Research Group in Molecular Bio-physics, University of Oxford, and was carried out in the Department of Zoology, Universityof Oxford.
REFERENCES
CARLSTROM, D. (1957). The crystal structure of a-chitin (poly-AT-acetyl-D-glucosamine).J. biophys. biochem. Cytol. 3, 669-683.
FRAENKEL, G. & RUDALL, K. M. (1947). The structure of insect cuticles. Proc. R. Soc. B 134,111-143.
FRASER, R. D. B., MACRAE, T. P. & MILLER, A. (1965). X-ray diffraction patterns of a-fibrousproteins. J. molec. Biol. 14, 432-442.
FRASER, R. D. B., MACRAE, T. P., MILLWARD, G. R., PARRY, D. A. D., SUZUKI, E. & TULLOCH,P. A. (1971a). The molecular structure of keratins. Appl. Polym. Symp. 18, 65-83.
FRASER, R. D. B., MACRAE, T. P., PARRY, D. A. D. & SUZUKI, E. (1971 b). The structure of featherkeratin. Polymer 12, 35-56.
FRASER, R. D. B., MACRAE, T. P. & ROGERS, G. E. (1962). Molecular organization in alpha-keratin. Nature, hand. 193, 1052-1055.
GERDES, R. J., CHAPMAN, A. T. & CLARK, G. W. (1970). Refractory oxide-metal composites:scanning electron microscopy and X-ray diffraction of uranium dioxide-tungsten. Science,N. Y. 167, 979-980.
HACKMAN, R. H. (i960). Studies on chitin. 4. The occurrence of complexes in which chitin andprotein are covalently linked. Aust. J. biol. Sci. 13, 568-577.
JENSEN, M. & WEIS-FOGH, T. (1962). Biology and physics of locust flight. V. Strength andelasticity of locust cuticle. Phil. Trans. R. Soc. Ser. B Z45, 137-169.
KELLER, A., DLUGOSZ, J., FOLKES, M. J., PEDEMONTE, E., SCALISI, F. P. & WILLMOUTH, F. M.(1971). Macroscopic 'single crystals' of an S-B-S three block copolymer. J. Physique 33,Csa, 295-300.
MEYER, K. H. & PANKOW, G. W. (1935). Sur la constitution et la structure de la chitine. Helv.chitn. Ada 18, 589-598.
NEVILLE, A. C. (1963). Growth and deposition of resilin and chitin in locust rubberlike cuticle.J. Insect Physiol. 9, 265-278.
NEVILLE, A. C. (1970). Cuticle ultrastructure in relation to the whole insect. In Insect Ultra-structure (ed. A. C. Neville), Symp. R. ent. Soc. Lond. No. 5, pp. 17-39.
NEVILLE, A. C. (1975). Biology of the Arthropod Cuticle, p. 448. Berlin: Springer-Verlag.NEVILLE, A. C. & LUKE, B. M. (1969). A two-system model for chitin-protein complexes in
insect cuticles. Tissue & Cell 1, 689-707.NEVILLE, A. C. & LUKE, B. M. (1971). Form optical activity in crustacean cuticle. J. Insect
Physiol. 17, 519-526.NIEDUSZYNSKI, I. A. & PRESTON, R. D. (1970). Crystallite size in natural celluloses. Nature,
Lond. 225, 273-274.RUDALL, K. M. (1963). The chitin/protein complexes of insect cuticles. Adv. Insect Physiol. 1,
257-3I3-RUDALL, K. M. (1965). Skeletal structure in insects. In Aspects of Insect Biochemistry, Biochem.
Soc. Symp., pp. 83—92. Cambridge University Press.RUDALL, K. M. & KENCHINGTON, W. (1973). The chitin system. Biol. Rev. 49, 597-636.SARKO, A. & MARCHESSAULT, R. H. (1969). Supermolecular structure of polysaccharides.
J. Polymer Sci. (C) 28, 317-331.WEIS-FOGH, T. (1961). Molecular interpretation of the elasticity of resilin, a rubberlike protein.
J. molec. Biol. 3, 648-667.WEIS-FOGH, T. (1970). Structure and formation of insect cuticle. In Insect Ultrastructure
(ed. A. C. Neville), Symp. R. ent. Soc. Lond. No. 5, pp. 165-185.ZELAZNY, B. & NEVILLE, A. C. (1972). Endocuticle layer formation controlled by non-circadian
clocks in beetles. J. Insect Physiol. 18, 1967-1979.(Received 6 October 1975)