the dynamics of active categorical perception in an ...meeden/developmentalrobotics/beer03.pdf209...
TRANSCRIPT

209
The Dynamics of Active Categorical Perception in an
Evolved Model Agent
Randall D. BeerDepartment of Electrical Engineering and Computer Science, and Department of Biology Case Western Reserve University
Notions of embodiment, situatedness, and dynamics are increasingly being debated in cognitive sci-ence. However, these debates are often carried out in the absence of concrete examples. In order to
build intuition, this paper explores a model agent to illustrate how the perspective and tools of dynam-
ical systems theory can be applied to the analysis of situated, embodied agents capable of minimallycognitive behavior. Specifically, we study a model agent whose “nervous system” was evolved using a
genetic algorithm to catch circular objects and to avoid diamond-shaped ones. After characterizing
the performance, behavioral strategy and psychophysics of the best-evolved agent, its dynamics areanalyzed in some detail at three different levels: (1) the entire coupled brain/body/environment sys-
tem; (2) the interaction between agent and environment that generates the observed coupled dynam-
ics; (3) the underlying neuronal properties responsible for the agent dynamics. This analysis offersboth explanatory insight and testable predictions. The paper concludes with discussions of the overall
picture that emerges from this analysis, the challenges this picture poses to traditional notions of rep-
resentation, and the utility of a research methodology involving the analysis of simpler idealized mod-els of complete brain/body/environment systems.
Keywords dynamics · minimally-cognitive behavior · categorical perception
1 Introduction
“May you live in interesting times.” This ancient Chi-nese curse has never been more appropriate than it isof cognitive science today. Once upon a time, life wassimple. Cognition was computation, and everyoneknew what that meant: formal manipulation of quasi-linguistic symbolic representations by syntactic rules.Then, in the early 1980s, connectionism began tomuddy the theoretical waters by significantly expand-ing our notion of what a representation could be andthe form a computation could take.
Now notions of situatedness, embodiment, anddynamics are threatening to once again revolutionize
some of our most basic assumptions about what cog-nition is and how it should be understood. For a situ-ated, embodied agent, taking action appropriate toboth its immediate circumstances and its long-termgoals is the primary concern, and cognition becomesonly one resource among many in service of thisobjective. An agent’s physical body, the structure ofits environment, and its social context can play asimportant a role in the generation of its behavior as itsbrain. Indeed, in a very real sense, cognition can nolonger be seen as limited to an agent’s head, but canbe distributed across a group of agents and artifacts.
Dynamical approaches emphasize the temporaldimension of cognition, and the way in which an
Copyright © 2003 International Society for Adaptive Behavior(2003), Vol 11(4): 209–243.[1059-7123(200312) 11:4; 209-243; 041345]
Correspondence to: Randall D. Beer, Department of Electrical Engineering and Computer Science, Case Western Reserve University, Cleveland, Ohio 44106-7071, USA.E-mail: [email protected]; Tel: +1-216-368-2801; Fax: +1-216-368-2816.

210 Adaptive Behavior 11(4)
agent’s behavior arises from the ongoing interactionbetween its brain, its body and its environment. Onthis view, the focus shifts from accurately represent-ing an environment to continuously engaging thatenvironment with a body so as to stabilize coordinatedpatterns of behavior that are adaptive for the agent.Rather than assigning representational content to neu-ronal states, the mathematical tools of dynamical sys-tems theory are used to characterize the structure ofthe space of possible behavioral trajectories and theinternal and external forces that shape the particulartrajectory that unfolds. Indeed, a dynamical approachto situated action raises important questions about thevery necessity of notions of representation and com-putation in cognitive theorizing.
Not surprisingly, such claims have met with agreat deal of skepticism, and there is considerablecontroversy regarding the fundamental significance ofsituatedness, embodiment, and dynamics to cognitivescience. Are these notions just minor corrections tothe classical picture of cognition, or do they radicallyalter the entire enterprise of cognitive science? Unfor-tunately, as in the early days of connectionism, the ini-tial debates have often been based more on competingintuitions than facts. Concrete dynamical models of awide range of cognitive phenomena are under construc-tion (including language, cognitive development, deci-sion-making, perception, and action (for reviews, seePort & van Gelder (1995) and Beer (2000)). But in themeantime, one powerful way to improve our intuitions,clarify the key issues and sharpen the debate is throughthe careful study of simpler idealized models of mini-mally cognitive behavior, the simplest behavior thatraises issues of genuine cognitive interest (Beer, 1996).Detailed dynamical analyses of situated and embodiedmodel agents can help us to expand our theoreticalimagination and to better ground the ongoing debates inconcrete examples (Beer, 1997, 1998). What kinds ofcognitive phenomena can dynamical notions accountfor? What kinds of mathematical tools can be brought tobear, and what sorts of insights do they provide? What isthe nature of a dynamical explanation and how does itdiffer from more traditional modes of explanation incognitive science? What are the advantages and disad-vantages of such an explanation? We will explorethese and other questions in the context of a detaileddynamical analysis of a particular model agent.
What sort of minimally cognitive behavior shouldwe examine? Stably partitioning the world into
objects with distinctive properties is one of the mostfundamental activities that an agent must perform.Indeed, it has been argued that categorical perception,as this activity is often called, may provide a neces-sary substrate for higher cognition (Harnad, 1987),making it particularly appropriate for detailed study asa minimally cognitive behavior. In categorical percep-tion, the continuous signals received by sense organsare sorted into discrete categories, whose membersresemble one another more than they resemble mem-bers of other categories. The paradigmatic example ofcategorical perception is the perception of phonemeboundaries in continuous speech (Liberman, Cooper,Shankweiler, & Studdert-Kennedy, 1967), but otherexamples include color categorization in humans(Boynton, 1975) and species-specific sound categoriza-tion in monkeys, birds, frogs, toads and insects (Ehret,1987; Wyttenbach, May, & Hoy, 1996). A variety ofneural network models of categorical perception havebeen developed and analyzed (Harnad, Hanson, &Lubin, 1995; Tijsseling & Harnad, 1997; Damper &Harnad, 2000; Cangelosi, Greco, & Harnad, 2000).
In this paper, I present a dynamical analysis of apreviously developed model agent that “visually” dis-criminates between objects (Beer, 1996). This agent iscontrolled by a dynamical “nervous system” that wasevolved using a genetic algorithm to catch circularobjects and avoid diamond-shaped objects. The inten-tion here is not to propose a serious model of categor-ical perception, but rather to use this model agent toexplore the implications of dynamical explanation forcognitive agents. For this reason, we will take theanalysis of this agent very seriously, systematicallyexamining in considerable depth each aspect of itsbehavior and the mechanisms underlying that behav-ior. While some may find the level of technical detailtedious, it is a basic premise of this work that a properappreciation of the nature of dynamical explanationrequires a thorough grounding in concrete examples.We must also address nontrivial issues involved inextending the techniques utilized for dynamical analy-sis of sensorimotor behavior to minimally cognitivebehavior. Furthermore, the ability to analyze a singleminimally cognitive agent is a crucial prerequisite forabstracting general principles from the analysis ofmany such agents. Think of this exercise, then, as aform of mental calisthenics, an intellectual warm-upfor the dynamical analyses of a wider range of agentsand behaviors.

Beer Categorical Perception 211
The paper is organized as follows. The next sec-tion presents an overview of the conceptual frame-work within which the work described in this paper iscarried out, one that attempts to do justice to theembodied, situated and dynamical nature of cognitiveagents. This is followed by a review of our object dis-crimination experiments. I then summarize the overallperformance and the behavioral strategy used by thebest agent that was evolved. Next, I describe theresults of psychophysical experiments that demon-strate categorical perception in the evolved agent,determine the object features that matter to the dis-crimination, and characterize the manner in which thedecision to catch or avoid is made. I then analyze thisevolved agent, first characterizing the dynamics of theentire agent-environment system, then decomposingthis in terms of the interactions between the agent andits environment, and finally examining the neuralimplementation of the agent dynamics. The paperconcludes with discussions of the overall picture thatemerges from this analysis, the relationship of thispicture to traditional notions of representation, and theutility of a research methodology based on analyzingsimpler idealized models of complete brain/body/environment systems.
2 An Embodied, Situated, Dynamical Perspective on Cognition
What does it mean to take an embodied, situated ordynamical perspective on cognition, let alone all threesimultaneously? There are probably as many answersto this question as there are advocates or critics ofthese various positions. Thus, I will make no attempthere to provide a critical survey of the ongoingdebates or to develop and defend a general situated ordynamical position. Fortunately, this ground is beingwell-covered elsewhere (Clark, 1997). Rather, themore modest aim of this section is to merely provide abrief sketch of the general conceptual frameworkwithin which the particular analysis described in thispaper will be carried out (Beer, 1995a, 1997).
As I intend them, embodiment and situatednessare closely related concepts. Both reject the notionthat purely intellectual ratiocination is the definingfeature of the cognitive. Rather, embodiment empha-sizes the role of an agent’s own body in its cognition,while situatedness emphasizes the role of an agent’s
immediate physical and social environment (Gibson,1979; Suchman, 1987; Beer, 1990; Brooks, 1991a;Varela, Thompson, & Rosch, 1991; Ballard, 1991;Hutchins, 1995; Hendriks-Jansen, 1996; Clancey, 1997;Agre, 1997; Glenberg, 1997; Lakoff & Johnson, 1998;Arkin, 1998; Pfeifer & Scheier, 1999). On this view,situated action is the fundamental concern, and cogni-tion is an elaboration of this more basic capacity, andone resource among many that can be brought to bearas an agent encounters its world. After all, nervoussystems originally evolved to coordinate behavior, notto balance checkbooks. An embodied nervous systemcan utilize the natural biomechanics of its body, thegeometry of its sensory surfaces, and its ability toactively control these surfaces to considerably sim-plify many problems. Likewise, a situated agent canutilize and manipulate the physical and functionalorganization of the space around it, as well as thesocial organization in which it exists, to offload prob-lems to its environment. On this view, cognitivebehavior arises from the interaction between a brain, abody and an environment (Figure 1), and cannot prop-erly be studied as a product of the mind/brain alone(Beer, 1995a). Given that nervous systems and bodiesco-evolve with one another and with their environ-ment, the need for such a coupled perspective ishardly surprising. What is surprising is the extent towhich such an embodied, situated perspective can
Figure 1 A dynamical perspective on a situated, em-bodied agent. The nervous system, body and environmentof an agent are each conceptualized as dynamical sys-tems that are in constant interaction. From this viewpoint,an agent’s behavior arises from this interaction betweenthese subsystems and cannot properly be attributed to anyone component in isolation from the others.

212 Adaptive Behavior 11(4)
alter our very conception of cognition, as well as thedomains in which we seek to understand its underly-ing mechanisms.
By a dynamical perspective, I mean simply one inwhich the concepts and mathematical tools of dynam-ical systems theory are applied to the analysis of thetemporal behavior of a cognitive system (Ashby,1952; Skarda & Freeman, 1987; Thelen & Smith,1994; Kelso, 1995; Elman, 1995; Beer, 1995 ; Port &van Gelder, 1995; van Gelder, 1998; Beer, 2000; The-len, Schöner, Scheier, & Smith, 2001). A dynamicalsystem is a mathematical object that unambiguouslydescribes how the state of some system evolves overtime. More formally, a dynamical system is a triple
consisting of an ordered time set T, a statespace S, and an evolution operator thattransforms an initial state at time toanother state at time . The time set T maybe continuous or discrete. The state space S may benumerical or symbolic, continuous or discrete, or ahybrid of the two, and it may be finite- or infinite-dimensional depending on the number of variablesrequired to fully describe the state of the system. Theevolution operator may be given explicitly ordefined implicitly, it may be deterministic or stochas-tic and it may have inputs (non-autonomous) or not(autonomous). Sets of differential or difference equa-tions, cellular automata, finite state machines and Tur-ing machines are all examples of dynamical systems.Dynamical systems theory offers a wide variety oftools for analyzing the temporal behavior of such sys-tems, including the identification of invariant sets(e.g., equilbrium points, limit cycles, etc.), a charac-terization of their local and global structure (e.g., sta-bility), and their dependence on parameters (e.g.,bifurcations) (Abraham & Shaw, 1992; Strogatz,1994).
Thus, to apply the tools of dynamical systems the-ory to a cognitive process is to seek a set of state vari-ables that uniquely characterize the behavior of theprocess and to uncover the laws by which these statesevolve in time. Armed with such a dynamical model,the dynamicist then attempts to understand the organ-izing features of its dynamics and the response ofthose dynamics to inputs or changes in system param-eters. Here the focus is on how the system’s behaviorunfolds over time and the various “forces” that shapethis trajectory. The importance of the states that thesystem passes through lies not so much in any content
that they may be assigned, but rather in their sensitiv-ity to subsequent inputs and the future behavior thatthey make possible.
While embodied, situated, and dynamical per-spectives can be and have been independently devel-oped and defended, these viewpoints work muchbetter as a unit. Taking a dynamical perspective on sit-uated, embodied action, an agent’s nervous system,its body and its environment are each described asdynamical systems, which are coupled (Figure 1).Because all of the individual components aredescribed in the same mathematical language, it ismuch easier to approach questions involving theirinteraction. To say that an agent’s behavior arisesfrom the interaction between its brain, body, and envi-ronment, for example, simply means that it is a trajec-tory of the complete dynamical system formed bytheir coupling, and the explanatory focus shifts to theproperties of this trajectory and the mechanisms bywhich it is generated and stabilized. This commonlanguage can be applied to cognitive processes, theneurophysiological processes that support it, non-cog-nitive human behavior, and the adaptive behavior ofsimpler animals, as well as the emergence of cogni-tion in development and evolution. Thus, in dynami-cal language, a unified theoretical framework forcognitive science broadly construed begins to seempossible.
3 An Object Discrimination Task
In a previous series of experiments, model agentswere evolved that could “visually” discriminatebetween circles and diamonds, catching the formerwhile avoiding the latter. These experiments weredesigned to produce evolved examples of categoricalperception, the best of which we will study in Sections4–8. Complete details of these experiments can befound in an earlier paper (Beer, 1996).
The agent has a circular body with a diameter of30, and can move horizontally as objects fall fromabove (Figure 2A). Horizontal velocity is proportionalto the sum of opposing forces produced by twomotors. The agent’s “eye” consists of seven rays dis-tributed over a visual angle of π/6. An intersectionbetween a ray and an object causes an input to beinjected into the corresponding sensory neuron, withthe magnitude of the injected input inversely propor-
T S φt, ,⟨ ⟩φt : S S→
x0 S∈ t0 T∈xt S∈ t T∈
φt

Beer Categorical Perception 213
Figure 2 Basic setup for the categorical perception experiments. (A) The agent can move horizontally while objectsfall from above. It uses an array of seven distance sensors to discriminate between circular (left) and diamond-shaped(right) objects, catching the former while avoiding the latter. (B) Examples of the class of hybrid objects, RH (φ; α) as, αis varied from 0 (circle) to 1 (diamond). (C) The network architecture consists of seven sensory neurons fully connectedto five fully interconnected interneurons, which are in turn fully connected to two motor neurons. All connections andneurons are bilaterally symmetric.

214 Adaptive Behavior 11(4)
tional to the distance to the object along that ray.When the rays are at their maximum length, no inputis injected, while the maximum input is injected forrays of zero length.
Objects fall straight down from the top of theenvironment with a fixed vertical velocity of –3 andan initial horizontal offset from the agent in the range±50. Circular objects had a diameter of 30 and dia-monds had sides of length 30. Both for uniformity ofnotation and for later analysis, it is convenient todefine a parameterized class of hybrid objects thatcontinuously interpolate between a circle and a dia-mond, even though the agent was exposed only to cir-cles and diamonds during evolution. This is mosteasily done in polar coordinates. If the circle isdefined by RC (φ) = 15 and the diamond is defined by
then the hybrid objects are given by RH (φ; α) = (1 – α) ×RC (φ) + αRD (φ). Sample objects produced as α var-ing from 0 to 1 are shown in Figure 2B.
The agent’s behavior is controlled by a continu-ous-time recurrent neural network (CTRNN) with thefollowing state equation (Beer, 1995c)
where s is the state of each neuron, τ is its time con-stant, wji is the strength of the connection from the jth
to the ith neuron, g is a gain, θ is a bias term, σ(x) = 1/(1 + e–x) is the standard logistic activation function,and I represents an external input (e.g., from a sensor).It will sometimes be convenient to refer to the outputof a neuron oi = σ(si + θi) rather than its state. The net-work architecture was bilaterally symmetric in theconnection weights, biases, and time constants. Thearchitecture consisted of seven ray sensory neurons
projecting to five fully interconnected interneurons,which in turn projected to two motor neurons control-ling horizontal motion (Figure 2C). All sensory neu-rons shared the same gain, bias, and time constant,while all other neurons had a gain of 1.
Thus, the equations for the complete model takethe form
(1)
where x is the horizontal position of the object relativeto the agent’s midline, y is the vertical position of theobject relative to the agent, and Ii (x, y; α) is the sen-sory input from the ith ray due to a hybrid object withthe given α at location (x, y) in agent-centered coordi-nates.
The connection weights, biases, time constants,and gain in the earlier equations were evolved. Theevolutionary algorithm used was a real-valued popula-tion-based hill climber (Bäck, 1996). The perform-ance measure to be maximized was Σ24
j = 1 pi /24, wherepi = 1 – | di | for circles (α = 0) and pi = | di | for dia-monds (α = 1), and di is the horizontal distancebetween the centers of the object and the agent whentheir vertical separation goes to zero on the ith trial. Atotal of 24 evaluation trials were uniformly distributedover the range of horizontal offsets.
Fourteen circle/diamond discrimination experi-ments were run with minor variations in the protocoldescribed earlier and with different random seeds.Seven of these achieved an average performance of> 90% on the discrimination task. Additional experi-ments that involved discriminating a circle from a linewere also run. The results of all of these experimentshave been previously reported (Beer, 1996). In thepresent paper, we focus on the detailed analysis of the
RD φ( )
15 2secφ1 tanφ+
------------------------- if 0 φ π/2≤ ≤
15 2secπ φ–1 tanπ φ–+
---------------------------------- if π/2 φ π≤ ≤
15– 2secφ1 tanφ+
---------------------------- if π φ 3π/2≤ ≤
15– 2secπ φ–1 tanπ φ–+
------------------------------------- if 3π/2 φ 2π≤ ≤
=
τis·i si– wjiσ gj sj θj+( )( ) Ii+
j 1=
N
∑+=
i 1 … N, ,=
y· 3–=
τs·i si– Ii x y α;,( ) i+ 1 … 7, ,= =
τis·i si– wjiσ g sj θ+( )( )
j 1=
7
∑+=
+ wjiσ sj θj+( ) i
j 8=
12
∑ 8 … 12, ,=
τis·i si– wjiσ sj θ+( ) i
j 8=
12
∑+ 13 14,= =
x· 5 σ s13 θ13+( ) σ s14 θ14+( )–( )=

Beer Categorical Perception 215
best circle/diamond discrimination agent that wasevolved over all of these experiments.
4 Performance and Behavior
The best evolved agent achieved a mean performanceof 99.83% on the 24 evaluation trials after 80 genera-tions of evolution, with a mean performance of97.08% on 10,000 random trials whose initial hori-zontal position was drawn from a uniform distributionover the range ± 50. The dependence of performanceon the initial horizontal position of an object is illus-trated in Figure 3. Note that there are very narrowspikes of poor performance near the centerline forboth circles and diamonds. In general, due to the bilat-eral symmetry of the task, objects whose initial posi-tions are very near the centerline are difficult tohandle. For example, a diamond that begins exactlycentered over the agent must be caught, because thereis nothing to break the bilateral symmetry of the body,environment and nervous system. There are also twobroader valleys of poor performance for circles. Thesefall between the locations of the 24 evaluation trialsused during evolution.
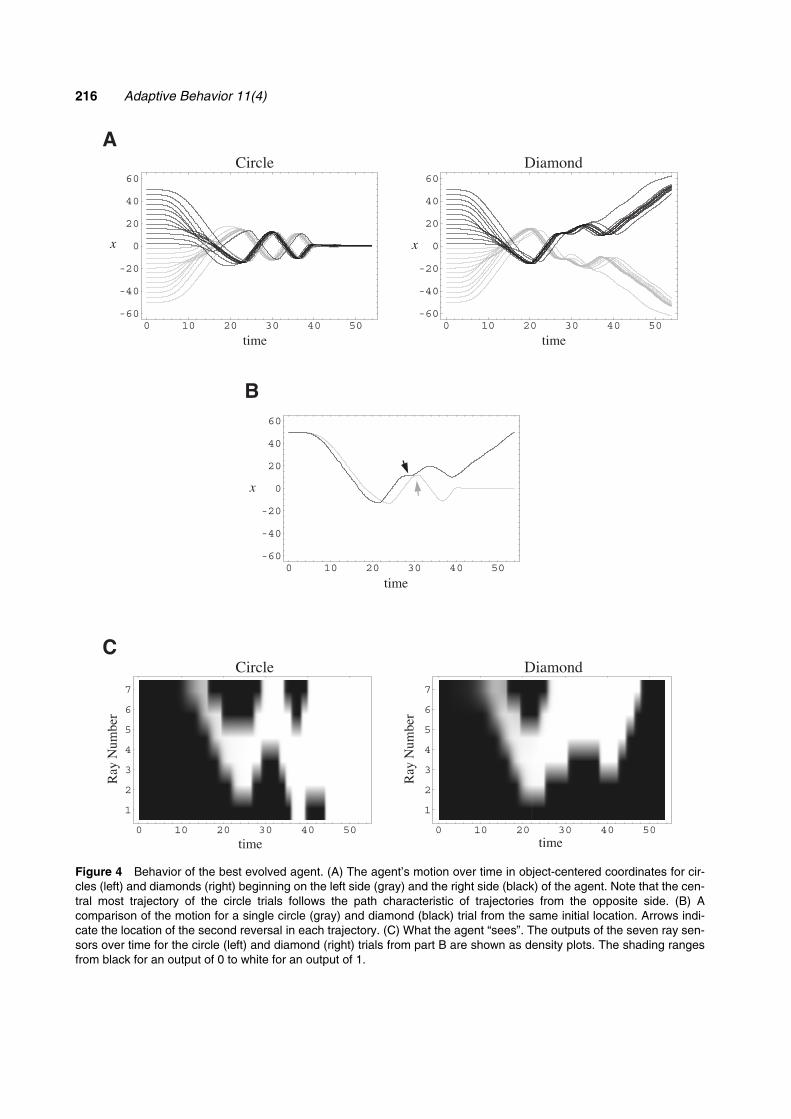
The basic strategy that this agent uses to distin-guish between circles and diamonds utilizes activescanning of the object as it falls (Figure 4A). Initially,the agent foveates any object by moving so as tocenter the object within its field of view. Next, theagent scans back and forth over the object. For a cir-cle, the amplitude of the scan decreases as the objectnears until it is centered, while the agent makes a largeavoidance movement for a diamond. Note that,although the agent responds more quickly to a dia-mond than a circle, the first half of the motion is qual-itatively very similar for both circles and diamonds(Figure 4B). However, the circle and diamondmotions differ qualitatively at the second reversal(Figure 4B, arrow), with the agent moving backtoward a circle, but pausing and then moving furtheraway from a diamond. This suggests that the secondreversal may be a key event in the discrimination, andsomething that we would like to better understand.
Note how the motion trajectories group into twobilaterally symmetric bundles depending on whichside of the agent the object appears initially. There isone exception to this. For circles, the centermost trialon each side crosses over to join the bundle character-
istic of initial positions on the opposite side. This pairof central trajectories corresponds to the inner two“fingers” of high performance that are separated fromthe more peripheral trials by the low-performance val-leys mentioned above (Figure 3A). Thus, it appearsthat the central two trials are treated somewhat differ-ently from the remaining trials, and the low-perform-ance valleys may represent a transition regionbetween these two strategies.
The agent’s active responses to a falling objectcan be characterized by examining its reaction toobjects held at fixed distances (Figure 5). The agentfoveates any object placed slightly off center at largevertical distances (y = 200, 175). At intermediate dis-tances (y = 150, 125, 100), the agent actively scansobjects, with diamonds scanned at a higher frequencythan circles. At closer distances (y = 75), a circle is
Figure 3 Generalization performance over initial hori-zontal position of the best-evolved agent. (A) Circle catchperformance as a function of initial object location. (B)Diamond avoidance performance as a function of initialobject location.
216 Adaptive Behavior 11(4)
Figure 4 Behavior of the best evolved agent. (A) The agent’s motion over time in object-centered coordinates for cir-cles (left) and diamonds (right) beginning on the left side (gray) and the right side (black) of the agent. Note that the cen-tral most trajectory of the circle trials follows the path characteristic of trajectories from the opposite side. (B) Acomparison of the motion for a single circle (gray) and diamond (black) trial from the same initial location. Arrows indi-cate the location of the second reversal in each trajectory. (C) What the agent “sees”. The outputs of the seven ray sen-sors over time for the circle (left) and diamond (right) trials from part B are shown as density plots. The shading rangesfrom black for an output of 0 to white for an output of 1.

Beer Categorical Perception 217
Figure 5 Active perception in the best evolved agent. The agent’s motion over time in response to a circle (gray) ordiamond (black) fixed at a range of vertical distances is plotted.

218 Adaptive Behavior 11(4)
centered and a diamond is avoided. Finally, at evencloser distances (y = 50), a diamond is centered and acircle is avoided. Of course, this is not a situation thatthe agent would normally find itself in, because bythis point a circle would normally have already beencentered and a diamond would normally have alreadybeen avoided.
The agent’s strategy of actively scanning fallingobjects is an interesting one. Given the coarse spatialresolution of the agent’s seven rays, it is likely thatthis scanning accentuates the small differencesbetween a circle and a diamond. To see just how smallthese differences are, it is useful to visualize circlesand diamonds from the agent’s perspective (Figure4C). While the difference between a circle and a dia-mond is obvious by the end of the interaction (becausea caught circle fills the field of view while an avoideddiamond leaves the field of view empty), it would bequite a challenge to perform this discrimination usingonly static snapshots of the visual array near the begin-ning of the behavior. Active control of body positionand orientation has become an important theme invision research, where it appears to considerably sim-plify otherwise intractable problems (Ballard, 1991;Churchland, Ramachandran, & Sejnsowski, 1994).
5 Psychophysics
5.1 Labeling and Discrimination
The two major defining characteristics of categoricalperception are labeling and discrimination (Studdert-Kennedy, Liberman, Harris, & Cooper, 1970; Ehret,1987). Labeling requires that stimuli from a continuumare partitioned into distinct classes, whereas discrimi-nation requires that stimuli are better distinguishedbetween classes than within a class. For this agent, clas-sification is manifested in its approach or avoidancebehavior. In order to demonstrate labeling, the meancatch performance was plotted as the α parameter of ahybrid object smoothly varied from 0 (circle) to 1(diamond). The resulting curve is sigmoidal in shape,with the category boundary occurring at approxi-mately α = 0.3 (Figure 6A). Below this value, objectsare classified as circles and caught. Above this value,objects are classified as diamonds and avoided (i.e.,their catch performance is 0). In order to demonstratediscrimination, the mean difference in catch perform-
Figure 6 Demonstration of labeling and discriminationin the best evolved agent. (A) Average catch perform-ance as a function of α shows a sigmoidal shape, with atransition from catching to avoidance behavior at about α= 0.3. Note that there is also an anomalous peak of rela-tively high catch performance at about α = 0.85. Eachpoint represents mean value and standard deviation of1000 trials, using initial horizontal offsets drawn from auniform distribution over ± 50. (B) Average difference incatch performance for α values that differ by 0.1 as afunction of α is bell shaped, with a peak discrimination atabout α = 0.3. Note that there is a double anomalouspeak of high discrimination centered at about α = 0.85.Each point represents mean value and standard devia-tion of 1000 trials. (C) A density plot illustrating the struc-ture of the data underlying the plot in part A. The meancatch performance is plotted as a function of the initialhorizontal position and the α value of an object, withblack representing a catch performance of 0 and whiterepresenting a catch performance of 100%. The plot inpart A can be recovered by averaging over x, while theplots in Figure 3 represent α = 0 and α = 1 slices throughthis plot.

Beer Categorical Perception 219
ance for hybrid objects separated by a ∆α of 0.1 wasplotted as a function of α. The resulting curve is bell-shaped, with a peak discrimination occurring near thecategory boundary at α = 0.3 (Figure 6B).
Note that both the labeling and discriminationplots are marred by an anomalous peak of high catchperformance around α = 0.85 (near-diamond shapes).Recall that the agent was only evolved to discriminatecircles (α = 0) and diamonds (α = 1); it was neverexposed to objects with intermediate values of α dur-ing evolution. Thus, the sigmoidal labeling curve andthe bell-shaped discrimination curve were interpolatedby the evolved network, and it is hardly surprising thatmistakes might appear at intermediate values to whichthe agent was never exposed during evolution. As willbe demonstrated later, this anomalous peak providesan important clue to the organization of the agent-environment dynamics that has evolved.
Finally, note that there is a fair amount of varia-bility in both the labeling and discrimination plots.We already know that there are two valleys of avoid-ance behavior near the centerline for circles (Figure3A) and that there are extremely narrow spikes ofcatch behavior near the centerline for diamonds (Fig-ure 3B). Such misclassifications clearly contribute tovariability in the mean performance statistics. In orderto obtain a more detailed picture of the structureunderlying this variability for general α, we can plotcatch performance as a function of the initial horizon-tal offset of an object as α is varied from 0 to 1 (Figure6C). In this plot, light bands correspond to catchingbehavior and dark bands correspond to avoidancebehavior. Both the category boundary around α = 0.3and the anomalous peak at about α = 0.85 have a verysimilar structure, with central and peripheral horizon-tal offsets switching behavior first as α is increased,followed by horizontal offsets at about ± 20, andfinally intermediate offsets. A faint peak near α = 0.6also exhibits a very similar structure (this small peakcan also be seen in Figure 6A).
5.2 What Features Matter to the Discrimination?
What object features does the evolved agent use tomake its categorization? Since the size of the circle issuch that it can be inscribed within the diamond, thetwo objects differ only in the four corners shown inFigure 7A, only three of which are visible to the agent
as the object falls. A coarse determination of the fea-tures important to discrimination can be made byexamining the agent’s response to the eight possiblecombinations of these three features. As the Figureshows, the presence of a single corner on either side ofthe object is sufficient to trigger avoidance, while thepresence or absence of the corner facing the agentappears to make very little difference to the agent’sresponse. This suggests that object width is the majordetermining factor in discrimination.
In order to test this hypothesis, we can examinethe agent’s response to a variety of different objectswhose width varies from 30 (the width of the circle) to
(the width of the diamond) (Figure 7B).In all cases, the resulting labeling curves are qualita-tively similar to Figure 6A (shown as a solid blackcurve in Figure 7B for reference), with the categoryboundary occurring at a width of 33.1 (SD 0.6) and theanomalous peak occurring at a mean width of 40.3 (SD
1.0). Note that, as would be expected, the labelingcurve of hybrid objects is more similar to that for cir-cles (dark gray curve) for α values closer to 0 and it ismore similar to that for diamonds (dotted curve) for αvalues closer to 1. The location of the category bound-ary varies in a consistent way with type of object, sug-gesting that the agent’s behavior is also somewhatsensitive to the amount that an object protrudestoward the agent.
5.3 When is the Decision Made?
At what point does the agent make its decision tocatch or avoid an object? We can probe this questionby switching the identity of the object at differentpoints in time and asking when the agent has commit-ted itself to a response. Figure 8A shows how the finalcatch performance at a single horizontal offset variesas a circle is switched to a diamond at later and latertimes (left) and how the final avoidance performanceat a single horizontal offset varies as a diamond isswitched to a circle at later and later times (right). Forexample, a high value in the circle plots (left) meansthat, even though the circle is switched to a diamondat that point, the agent still responds as if it were a cir-cle and was thus already committed to catching it.However, the multiple sharp changes in performancethat occur in both plots as the time of the switch is var-ied demonstrate that the agent’s behavior remainsquite sensitive to object identity until very late in the
30 2 42.43≈

220 Adaptive Behavior 11(4)
interaction. The plots for other horizontal offsetsexhibit the same general pattern (Figure 8B), andsome features of this pattern correlate closely with theagent’s behavior (Figure 8C). For example, the brief
peak in catch performance for circles at about t = 33and the sharp rise in avoidance performance for dia-monds at about t = 29 coincide with the secondreversal.
Figure 7 A demonstration that object width is the primary feature used by the best evolved agent to discriminate cir-cles from diamonds. (A) The agent’s performance on objects possessing all possible combinations of the three discretefeatures (shown in black at top) that distinguish a circle from a diamond. Note that the agent only catches objects withno left or right corners; the presence of either one or both left/right corners leads to avoidance. (B) Variation in catch per-formance with object width for a variety of different objects, including a horizontal line (dashed gray curve), wedgeswhose height is the same as the circle and the diamond (dashed black curves), diamonds (dotted black curve), the hy-brid objects, RH (φ; α) (solid black curve), ellipses whose height is the same as the circle (solid light gray curve), and cir-cles and ellipses whose height is the same as a diamond (solid dark gray curve). Note that, as their width is varied, allobjects produce a qualitatively similar response in the agent.

Beer Categorical Perception 221
Taken at face value, these results suggest that the“decision” is repeatedly made and unmade as theagent and the object interact. For example, the agentseems to be completely uncommitted during the initialfoveation of a circle, but is then largely committed to
catching during the first scan (although there are timeswhen this commitment wavers). The agent is thenuncommitted during subsequent scans, but committedto catching at each reversal. By the final reversal neart = 38, the agent is largely committed to catching the
Figure 8 The effect of switching object identity at different times on the final decision to catch or avoid an object in thebest evolved agent. (A) Catch performance as a function of the time of switch from a circle to a diamond (left) and avoid-ance performance as a function of the time of switch from a diamond to a circle (right) for a single initial horizontal offset.(B) Catch (left) and avoidance (right) performance as a function of time of switch for twelve different initial horizontal po-sitions. In both cases, note that the agent’s sensitivity to switches in object identity appears to vary considerablythroughout the interaction. (C) The normal behavioral trajectories for the initial horizontal offsets used in (B). Note thecorrespondence between features of the behavioral trajectories and features of the performance plots in (B).

222 Adaptive Behavior 11(4)
Plate 1 The dynamics of the best evolved agent. (A) Steady-state horizontal velocity fields for circles (left) and dia-monds (right). Each point represents the long-term horizontal velocity that the agent would eventually adopt if an objectwere fixed at that location in its field of view. These fields correspond to an x· projection of the coordinates of the equilib-rium points of the evolved continuous-time recurrent neural network as a function of x and y. Regions in which x· is di-rected toward the midline (centering regions) are colored blue, whereas those in which x· is directed away from themidline (avoidance regions) are colored red. The magnitude of x· is represented by the intensity of a color, with x· = 0 rep-resented as black. Green regions are those in which multiple stable equilibrium points exist. The straight white lines in-dicate boundaries across which the object intersects different numbers of rays. These ray tangencies divide the visualfield into disjoint regions of smooth dynamics. (B) The fields from part A with the movement trajectories from Figure 4Asuperimposed. The trajectories are color-coded with instantaneous horizontal velocity using the same scheme as in partA. If the interaction were frozen at any point along a trajectory, the color of that point would eventually approach thecolor of the corresponding point of the steady-state field. However, since both the agent and the object are moving, theinstantaneous velocity lags behind the steady-state velocity. (C) Some additional example trajectories. The inner trajec-tories are misclassifications taken from the regions of low performance in Figure 3. The outer trajectories demonstratethat all sufficiently peripheral objects are avoided.

Beer Categorical Perception 223
object. The pattern is less clear-cut for diamonds.However, by the time of the pause near t = 27, theagent is fairly committed to avoiding the object(although there are times during the final scan whenthis commitment wavers). Thus, it is not until theexpression of the decision is nearly complete that astable decision can really be said to have been made.These results suggest that the generation of a catch oravoidance response is a complex, temporally extendedprocess, and it makes little sense to talk about the agenthaving made a decision before its behavioral expressionof that decision. A similar view of decisions as tempo-rally extended processes shaped by both sensory sig-nals and neuronal state has begun to emerge from thedecision-making literature (Townsend & Busemeyer,1995; Platt, 2002).
6 The Dynamics of the Coupled Agent–Environment System
Let us now turn to an analysis of the dynamics under-lying the behavioral and psychophysical phenomenadescribed in the previous two sections. The first stepin such an analysis is to characterize the dynamicalstructure of the entire coupled system formed by theevolved nervous system, the agent’s body and the fall-ing object (Figure 1). There are a number of questionsthat we would like to answer about the dynamics ofthis agent–environment system. How is its dynamicsstructured so that almost all circle trajectories result incentering, while almost all diamond trajectories leadto avoidance? How does this structure change as αincreases so that the agent’s response changes fromcatch to avoid, resulting in a category boundarybetween circles and diamonds at about α = 0.3? Whatcauses the anomalous peak at about α = 0.85?
From the equations describing the coupled agent–environment system (equation 1), we can make sev-eral general observations about its dynamics. First, thesystem is 16-dimensional, because 16 separate differ-ential equations are required to uniquely describe itsmotion. Second, this system has no closed limit setssuch as equilibrium points or limit cycles because they· = –3 equation means that y is always decreasing.Third, discontinuities are introduced into the dynam-ics through the ray sensory neurons (neurons onethrough seven) because Ii (x, y; α) is a discontinuousfunction of x and y. There are locations where an
infinitesimal change in position can cause an objectthat was not intersecting a ray to suddenly intersect it,or vice versa. These ray tangencies divide the statespace into discrete cells of smooth dynamics whichdepend on the size and shape of the object (see Plate 1),making the agent a hybrid dynamical system (Johnson,1994). One final general observation that we can makeis that the coupled agent-environment system is actu-ally a one-parameter family of systems, because chang-ing the object from a circle to a diamond amounts tochanging the α parameter from 0 to 1.
In order to say anything more specific about thiscoupled dynamics, we must examine its detailedstructure. While it is not possible to directly visualizethe trajectories of such a high-dimensional system,carefully chosen projections can provide importantinsights into this structure. By examining various pro-jections of the dynamics, it was discovered that com-binations of x, y and an interneuron are most useful.The three-dimensional projection (x, y, o9) leads toparticularly clear visualizations of the dynamics.Interestingly, this projection includes an environmen-tal state variable (the vertical position of the object), abody state variable (the horizontal position of thebody relative to the object), and a neuronal state varia-ble (the output of interneuron 9).
In the (x, y, o9) projection, the trajectories beginas a line of initial states and move to decreasing y val-ues as the object falls (Figure 9). When the leadingedge of the object reaches the leading edge of theagent, a fitness value is assigned based on their hori-zontal separation. By adjusting network parameters,the evolutionary algorithm has been asked to structurethe dynamics of the entire coupled system so that theline of initial states is concentrated near x = 0 by theend of the trial for circles, but is dispersed to largepositive or negative values of x for diamonds.
Figure 9 shows the structure of the 24 evaluationtrials as α is varied from 0 to 1, with the trajectoriesshaded according to final catch performance so that itis easier to distinguish those that are caught fromthose that are avoided. Note that the motion trajecto-ries shown in Figure 4a are simply the projection ofthe trajectories shown in Figure 9 onto the (x, y) plane.As expected, there are no closed limit sets and the tra-jectories merely flow from left to right. However, theydevelop quite a bit of structure as the interactionunfolds. In particular, note how the line of initialstates is separated into two distinct bundles of trajec-

224 Adaptive Behavior 11(4)
Figure 9 The dynamics of the coupled agent–environment system. The (x, y, o9) projections of twenty-four trajectoriesof the system are shown. The plots are arranged in a counter-clockwise direction as α varies from 0 (circle) to 1 (dia-mond). The trajectories are shaded according to catch performance, with lighter shades representing high catch per-formance and darker shades representing low catch performance. Note that the trajectories are organized into twobundles that wind around one another. For small values of α, these bundles collide on the axis of bilateral symmetry andthe objects are caught. As α increases, the bundles progressively unwind. As can be seen in the α = 0. 8 and α = 0.9plots, the anomalous peak in Figure 6 corresponds to the two bundles colliding at their third crossing.

Beer Categorical Perception 225
Plate 2 The circle steady-state response fields of the bilateral pairs of interneurons 8/12 and 9/11. (A). A density plot ofthe feedforward response fields of each interneuron in response to a circle placed at each point in the agent’s field ofview. Here only the feedforward connections from the sensory neurons to the interneurons are considered. White re-gions correspond to an output of 1 while black regions correspond to an output of 0. (B) A density plot of the interneuronresponse fields when the recurrent interactions are also taken into consideration. Note that intensity of the feedforwardresponse fields is modified in places and that regions of multistability appear (green). (C) The steady-state horizontal ve-locity fields of each interneuron, color-coded in the same way as in Plate 1. These fields show the steady-state horizon-tal velocity produced by each interneuron in response to a circle placed at each point in the field of view. (D) The netsteady-state horizontal velocity field formed by summing the four fields in Part C. Note that this summed response fieldclosely matches the steady-state horizontal velocity field shown in Plate 1A (left).

226 Adaptive Behavior 11(4)
tories that wind around each other several times. Eachtime the agent scans to the right, the output ofinterneuron 9 develops a peak (three arrows at top ofα = 0 plot) that allows the bundle to cross over theother bundle’s corresponding scan to the left. Notethat the trajectory bundle resulting from objects thatinitially appeared to the right of the agent exhibits twopeaks, while the bundle resulting from objects that ini-tially appeared to the left exhibits only one. Of course,due to bilateral symmetry, plots of the (x, y, o11) pro-jection would be mirror images of these, with thepeaks occurring during scans to the left and the orderand number of peaks for objects initially on the left ofthe agent reversed as compared with objects initiallyon the right.
How do these plots help us to answer the ques-tions raised at the beginning of this section? Let usbegin by examining the circle trials (α = 0). Aftercrossing over one another three times, the two circletrajectory bundles appear to “collide” and merge at thetrajectory of bilateral symmetry rather than crossing afourth time (arrow at bottom). Note that one must becareful in drawing such a conclusion from a singlelower-dimensional projection. However, examinationof other projections verifies that the two trajectorybundles do indeed interpenetrate at this point (sincethe agent dynamics is non-autonomous, distinctbehavioral trajectories can pass through the same neu-ronal state). From the point of this collision on, bothbundles follow the trajectory of bilateral symmetry(the trajectory that the dynamics follows in responseto an object dropped at the midline), resulting in accu-rate catches. As α increases, the trajectories appear tounwind. While they retain their basic shape, they shiftto larger values of y so that the peaks occur earlier intime. As the bundles shift earlier in time with increas-ing α, more and more trajectories miss the final colli-sion and instead diverge from the trajectory ofbilateral symmetry until, by α = 0.5, the two bundlesdo not intersect at all. As α continues to increase, thetwo trajectory bundles continue to shift earlier in timeuntil they begin to collide at the third crossing at α =0.8, resulting in catches that produce the anomalouspeak. As α increases even further, the bundles shift tothe point where, by α = 1, they cross only twice beforediverging.
These observations immediately suggest a predic-tion. From our psychophysical analysis, we know thatthe most important effect of increasing α is to increase
the width of the object. Assuming that the two trajec-tory bundles continue to unwind with increasingobject size, we would expect that two additional colli-sions should occur as the bundles intersect at the sec-ond and then the first crossing. This reasoningpredicts that two additional anomalous peaks in catchperformance should be observed for objects widerthan the α = 1 diamond. As shown in Figure 10A, thisis indeed the case. The catch performance is plottedfor diamonds ranging in width from 30 to 100. From30 to 42.4, the plot is a compressed version of the dot-ted black line in Figure 7B, with the anomalous peakappearing as a narrow spike. From a width of about47.7 to about 54.5, there is a second short anomalouspeak. Finally, beginning around a width of 59, there isa final slow increase in catch performance. Figure10B shows how a collision of the two trajectory bun-dles at the second crossing is responsible for the sec-ond short peak (w = 51.2) and a collision at the firstcrossing is responsible for final broad peak (w = 66),as predicted. Note that, between these two anomalouspeaks, the bundles have unwound past the secondcrossing (w = 54.8). Beyond the start of the last anom-alous peak, the trajectory bundles have unwound com-pletely (w = 89.6).
7 The Analysis of Agent–Environment Interaction
The characterization of the overall dynamical struc-ture of the coupled agent–environment system in theprevious section certainly provided some explanatoryand even predictive insight into this agent’s behavior.However, it could be argued that this characterizationis primarily descriptive in nature, detailing what thestructure is without explaining how it arises. The sec-ond step in a dynamical analysis is to examine howthis structure emerges from the interaction betweenthe agent and its environment. What produces theactive scans? What causes the two trajectory bundlesto develop and to have the structure that they do? Whydo circle trajectories move back toward the midlineat the second reversal, while diamond trajectoriesmerely pause and then continue moving away? Whathappens when the two bundles collide at the trajectoryof bilateral symmetry? What causes the two bundlesto unwind with increasing α? Why is object width thedetermining factor in discrimination? What underlies

Beer Categorical Perception 227
Figure 10 Confirming a prediction of additional peaks as object width increases. (A) A plot of catch performance for adiamond whose width varies from the width of a circle to more than twice the width of a normal diamond. Note that addi-tional anomalous peaks occur. The dashed vertical lines correspond to the predicted widths at which anomalous peaksshould begin based on the reasoning described in the text. (B) Trajectory plots for diamond widths within the third peak(w = 51.2), between the third and fourth peak (w = 54.8), and at two different points within the fourth peak (w = 66 and w= 89.6). The format is the same as in Figure 9. Note that the two bundles continue to unwind with increasing object widthand that the two new anomalous peaks correspond to collisions occurring at the second or first crossing.

228 Adaptive Behavior 11(4)
the misclassifications that occasionally occur? Why isthe moment of decision to catch or avoid so difficultto pin down?
7.1 Steady-State Effects
Once again, we have many different choices as to howto decompose the dynamics of the agent–environmentsystem and how to visualize it. Clearly, whateverdecomposition we choose should emphasize the fac-tors underlying movement, since the agent’s decisionis expressed in its motion over time and it is thisbehavior that we want to explain. Thus, we willdecompose the agent–environment dynamics into: (1)the effect that the relative positions of the object andthe agent have on the agent’s motion; (2) the effectthat the agent’s motion has on the relative positions ofthe object and the agent.
Imagine a particular object fixed at some (x, y)location in the agent’s field of view. This specifies aconstant set of sensory inputs to the evolved neuralcircuit as described by the Ii (x, y; α) terms in equation1. For constant inputs, the neural circuit is an autono-mous dynamical system. It turns out that the only limitsets exhibited by this circuit are equilibrium points,whose positions and stabilities we can calculate.Although each stable equilibrium point is 14-dimen-sional, the only quantity that directly affects theagent’s motion is the horizontal velocity x·, which isdetermined by o13 and o14 as described in equation 1.If we wait long enough after fixing the object in theagent’s field of view, the state of the neural circuitwill be found very near one of its stable equilibriumpoints. Thus, for each point in the agent’s field ofview, we can plot a representation of the steady-statevelocity that would be produced by an object locatedat that point (Plate 1A). These points are color-codedaccording to whether the steady-state x· is directedtoward (blue) or away (red) from the centerline. Theintensity of the color represents the magnitude of x·,with an x· of zero colored black. Green points indicateobject locations where the neural circuit has more thanone stable equilibrium point. In this case, the long-term horizontal velocity is not uniquely determined byobject position alone, but depends on the system’sstate. Note the straight, sharp borders between regionsof different color that radiate at several angles fromthe bottom of the plots. These correspond to the raytangencies described earlier (white lines in Plate 1A),
which separate the regions of smooth dynamics wherethe object intersects different subsets of rays.
Superimposed on these steady-state velocityfields are the trajectories of motion of the objectthrough the agent’s field of view for the 24 trials usedduring evolution (Plate 1B). These trajectories arecolor-coded according to their instantaneous horizon-tal velocity using the same color scheme describedearlier. If the interaction were frozen at any pointalong a trajectory, then the instantaneous velocitywould approach the steady-state value over time as theinternal state of the neural circuit approached the cor-responding stable equilibrium point. However, sinceboth the agent and the object are moving, the instanta-neous velocity lags behind the steady-state value. Aswe shall see, this subtle interplay between sensoryinput and internal state is crucial to accurate discrimi-nation.
How can these plots be used to understand theagent’s behavior? Consider the circle trials shown inPlate 1B (left). The most striking feature of this plot ishow the blue centering regions repeatedly turn the tra-jectory bundles back toward the midline until theybecome trapped in the central black regions of no hor-izontal movement. Initially, all trajectories begin indark-blue regions corresponding to equilibrium pointsthat attract the agent’s velocity toward the center,slowly at first, and then with increasing velocity as theregions brighten. The trajectories overshoot the mid-line due to the lag mentioned above between theactual internal state of the agent and the changingequilibrium points that are attracting it (it takes sometime for the neurons to respond to a sudden change insensory input). This lag is also the reason that the briefpassage of the outermost trajectories through a redregion near the top of the plot has little effect on them.Note that the trajectories turn red as soon as they crossthe midline. This is because, although a motion of theobject toward the right is a centering movement whenthe object is on the left side of the midline, it becomesan avoidance movement when the object is on theright side of the midline and vice versa. As these nowred trajectories pass through blue centering regions onthe opposite side of the midline, they eventually turnblue and move back toward the center. This overshootand turning back toward the midline occurs twicemore. Thus, the active scans are produced by the inter-play between the steady-state horizontal velocity fieldand a kind of “neuronal inertia” that causes the agent

Beer Categorical Perception 229
to repeatedly overshoot the midline. If this inertia isnot allowed to build up (e.g., by decreasing the timeconstants of the interneurons), then the agent’s dis-crimination performance drops sharply.
The last time the trajectory bundles approach themidline, they pass briefly through a red avoidanceregion before moving into the central black region andbecoming trapped. The black region corresponds toobject positions for which there exists a single stableequilibrium point whose x· projection is 0. If the agenthad sufficient neuronal inertia at this point, it wouldpass right through this region. In fact, there is a slightovershoot of the midline. However, this overshoot isnot sufficient to carry the agent out of the blackregion. As the object continues to fall, the trajectoriesbriefly intersect the corners of the central red regions,causing a small transient movement that actuallyserves to further center them (we will examine suchtransient effects shortly). Thus, what happens whenthe two trajectory bundles collide is that they run outof neuronal inertia in the upper black diamond-shapedregion of the steady-state horizontal velocity field,leaving neither sensory inputs nor internal state to ini-tiate further horizontal motion.
Now consider the diamond trials shown in Plate1B (right). Although the details are somewhat differ-ent, the upper half of the steady-state horizontal veloc-ity field and the initial motion of the trajectories issimilar to that for circles. The most striking differencefrom the circle trials is that the central red regions ofthe steady-state horizontal velocity field in the lowerhalf of the plot (and the black regions separatingthem) are much larger for diamonds than for circles,and penetrate much higher into the agent’s field ofview. These red avoidance regions have the effect ofpushing the trajectory bundles away from the midlineafter the first scan, so that they never pass into thecentral black regions. Whereas the circle trajectoriespass through only blue regions at the second reversal,turning them back toward the midline, the diamondtrajectories pass through the top of the expanded cen-tral red region, causing only a brief pause rather than areversal. The diamond trajectories then enter a bluecentering region that pushes them back toward themidline, but the large central red region again pushesthem away, resulting in a final avoidance. Because thesize and location of the central regions are determinedby the width of the object (compare the left and rightsides of Plate 1A), it is easy to see how wider objects
unwind the trajectory bundles (Figure 9) as theexpanding red avoidance regions push the trajectoriesaway from the midline earlier and earlier in the inter-action.
Using this decomposition of the agent-environ-ment dynamics, we can account for the shape of thecatch performance plot (Figure 6A) as follows. As thewidth w of an object increases, the regions of smoothdynamics (delimited by the white lines in Plate 1A)enlarge and shift upwards (compare the left and rightsides of the Figure), causing the central black regionto intersect each of the four midline crossings in turn.Catch peaks begin whenever the trajectory bundlespass through a sufficiently wide portion of the blackregion that they become trapped. At w = 30, the origi-nal peak occurs when the mean vertical offset of thefourth midline crossing falls approximately 13.6below the top of the black region. For a diamond, theray tangencies occur at y = mi (x ± w/2), where mi isthe slope of the ith ray. By solving for the ray tangencyintersections that define the upper central blackregion, we find that its top occurs at y = m6w / 2. If wesolve for the object widths at which the other threemidline crossings occur 13.6 units below the top ofthe central black region, we obtain quantitative pre-dictions for where the three anomalous peaks shouldbegin. As the dashed lines in Figure 10A show, thesepredictions are fairly accurate. Catch peaks end whenthe red avoidance region surrounding the lower por-tion of the central black diamond become sufficientlylarge that the trajectories are pushed away before theyever enter the black region. This happens only threetimes because, for sufficiently wide objects, the cen-tral lower black triangle-shaped region becomes solarge that no trajectories can avoid becoming trappedwithin it.
This decomposition of the agent–environmentinteraction can also explain a number of other featuresof the agent’s behavior. Misclassifications occur whenthe agent’s internal state gets sufficiently out of phasewith the steady-state velocity field. For example, inthe valleys of low circle performance (Figure 3A), thescan is delayed to the point where these trajectoriespass through enough of the red avoidance region tocarry them out of the black region that normally trapscircle trajectories (Plate 1C, left). Similarly, in thenarrow spikes of low diamond performance (Figure3B), the initial scan is delayed long enough that thetrajectories become trapped in the central black region

230 Adaptive Behavior 11(4)
and they are caught (Plate 1C, right). The differentpatterns of active scanning at different vertical dis-tances (Figure 5) can be explained by the differencesin the steady-state horizontal velocity fields for circlesand diamonds at different vertical distances along themidline. The complexity of the decision plots (Figure 8)is due to the fact that switching object identity corre-sponds to replacing the steady-state horizontal veloc-ity field for one object with that for the other.Depending on the position of the object and the inter-nal state of the agent at the time of the switch, the sub-sequent interaction of the agent with the new velocityfield can sometimes cause the object to be caught andsometimes to be avoided. Finally, the outer red avoid-ance regions observed in both circle and diamondvelocity fields predict that the agent will avoid all suf-ficiently peripheral objects. As shown by the periph-eral trajectories in Plate 1C, this is indeed the case.
7.2 Transient Effects
The steady-state horizontal velocity fields providesome qualitative insight into how the interactionbetween this agent and its environment generates theobserved categorization behavior, and even providesome quantitative predictive power. However, they donot fully explain the fine details of the behavior. Forexample, note that some trajectories pass into the cen-tral black region in all cases shown in Plate 1B andC. Why is it that some become trapped there (Plate1B, left and Plate 1C, right) while others do not (Plate1B, right and Plate 1C, left)? In order to answer suchquestions, we must look in more detail at the transientstructure of the agent dynamics when the objectpasses into the central black region of the steady-statevelocity fields, taking into account the neuronal iner-tia provided by the lag of the CTRNN state.
Figure 11 illustrates the transient effects underlyingthe agent’s behavior for two trajectories that becometrapped in the black region (C1 and D2) and two trajec-tories that do not (C2 and D1). The actual trajectory ofthe second largest principal component of the CTRNNstate is plotted against x· (black curves) from the time anobject enters the black region until the time it exits orthe trial ends for one example of each of the four cases.At each point along a trajectory (black dots), the agentis following a particular instantaneous trajectory (graycurves) toward the equilibrium point currently attract-ing it (gray points). Note that although the vertical loca-
Figure 11 The role of transient effects in the dynamicsof the best evolved agent for two circle (C) and two dia-mond (D) trajectories. (A) The passage of each of thefour trajectories through the central black diamond-shaped region in the steady-state horizontal velocity fieldfor circles (left) and diamonds (right) is shown. C1 is anormal circle catch from Plate 1B (left). C2 is a mistakencircle avoid from Plate 1C (left). D1 is a normal diamondavoid from Plate 1B (right). D2 is a mistaken diamondcatch from Plate 1C (right). (B) The second principalcomponent of continuous-time recurrent neural network(CTRNN) output is plotted against horizontal velocity asthe agent enters the central black diamond for the four cas-es shown in part A. PCA 2 can be considered a depictionof the internal state or “neuronal inertia” of the CTRNN.PCA 2 was chosen over PCA 1 because the former led toless overlap of trajectories in this plot. The labeled pointsshow the state of the agent at the time the object entersthe black region and the sequence of black dots showhow this state evolves over time until the agent leaves theblack region. The gray curves show how the state wouldevolve if the interaction were frozen at selected pointsalong the trajectory. Note that the two circle trajectories(C1 and C2, light gray) are ultimately headed toward an x· =0 equilibrium point (light gray point), and the two diamondtrajectories (D1 and D2, dark gray) are ultimately headedtoward a slightly different x· = 0 equilibrium point (darkgray point). However, due to differences in internal state,only C1 and D2 actually reach the equilibrium points andthe agent stops horizontal motion, whereas C2 and D1never reach the equilibrium points before the object pass-es out of the black region.

Beer Categorical Perception 231
tion of the equilibrium point is slightly different forcircles (light gray) and diamonds (dark gray), bothoccur at x· = 0, which is why this region is colored blackin the steady-state velocity fields. If the object remainedin this region long enough, the black trajectories wouldeventually reach the equilibrium points at x· = 0 and theagent would stop moving in all four cases. However,because the CTRNN dynamics is parameterized by the(x, y) location of the object in the agent’s field of viewand both the object and agent are moving, the CTRNNdynamics is constantly changing, causing the state tofollow a slightly different instantaneous trajectory ateach time step (gray curves).
With this background, we are ready to explain whysome trajectories pass through the central black regionof the steady-state velocity fields while others becometrapped there. For all four trials, the state begins at x· <0, so the object is moving to the left in the agent’s fieldof view. For a normal circle trial (C1), the state brieflycrosses into the x· < 0 region, leading to a small right-ward movement of the object, and then reaches theequilibrium point. This ceases horizontal motion of theagent and results in a catch. For the case in which a cir-cle is mistakenly avoided (C2), a qualitatively similartrajectory is followed. However, due to the differentCTRNN state and the different sequence of relativepositions of the object and the agent, larger positivevalues of x· are reached and a much larger rightwardmovement is generated, causing the object to reversedirection and move out of the black region before thestate reaches the equilibrium point. For a normal dia-mond trial (D1), the time spent in the black region is sobrief that the state barely has a chance to move towardthe equilibrium point. Finally, for the case in which adiamond is mistakenly caught (D2), the CTRNN stateand sequence of object positions are such that themotion toward the equilibrium point is even faster andmore direct than for a normal circle trial. Thus, the par-ticular trajectory that the agent follows depends on theinterplay between the internal neural state and thechanging (x, y) location of the object in the agent’s fieldof view.
8 Neural Implementation of Agent Dynamics
The final step in a dynamical analysis of this agent isto examine how the agent dynamics described in the
previous section is actually instantiated in the evolvedneural circuit. How do the specific neuronal and syn-aptic parameters in this network produce the observedsteady-state horizontal velocity fields? What neuralproperties give rise to the central black region soessential to catching circles? What neuronal interac-tions underlie the transient effects involved in over-shooting the midline during scans and in trappingtrajectory bundles in the central black region for cir-cles? We will use a graphical illustration of theevolved circuit architecture (Figure 12) to presentanswers to all of these questions.
One observation that follows immediately fromFigure 12 is that interneuron 10 has a high threshold,inhibits itself, and is either strongly inhibited or onlyvery weakly excited by the sensory neurons. This sug-gests that this neuron will remain off under most con-ditions and thus may not play any significant role inthe circuit’s operation. Lesion studies largely confirmthis. When interneuron 10 is removed, diamondavoidance performance only drops from 99.38 to99.18% (N = 1000). However, circle catch perform-ance drops from 95.08 to 88.77% due to a small butcrucial effect that interneuron 10 has at one point inthe behavior. Nevertheless, this difference is smallenough that we will not consider interneuron 10 fur-ther in our analysis.
8.1 Steady-State Effects
Our first task is to account for the structure of the steady-state horizontal velocity fields (Plate 1). This is mosteasily done in three steps. First, we will examine theeffect of the feedforward sensory connections in the cir-cuit on the interneuron response fields (Plate 2A). Thenwe will consider the contribution of the recurrent inter-actions between the interneurons (Plate 2B). Finally, wewill examine the consequences of the interneuronresponses on the agent’s motion (Plate 2C) and comparethe combined effect of movement responses to theobserved steady-state horizontal velocity fields (Plate 1).Our analysis will focus on the circle fields. The mecha-nisms underlying the diamond fields are very similar andwe have already explained the differences in size andshape between the circle fields and the diamond fields interms of how the locations of the lines of ray tangencydiffer for the two classes of objects.
The feedforward response fields for the bilateralinterneuron pairs 8/12 and 9/11 are shown in Plate 2A.

232 Adaptive Behavior 11(4)
These fields represent the output of each neuron inresponse to a circle at each point in the visual field.Note that interneurons 8/12 primarily respond toobjects on the same side, whereas interneurons 9/11primarily respond to objects on the opposite side.
These features are relatively easy to explain. Both thesensory neurons and the interneurons are intrinsicallyoff. The left side of interneuron 8’s response field isblack due to the inhibition it receives from sensoryneurons 1 and 2, while the right half of interneuron 9’s
Figure 12 A graphical depiction of the evolved continuous-time recurrent neural network (CTRNN) of the best agent.The feedforward architecture is shown in (A), while the recurrent interactions between the interneurons are shown in(B). Neurons are shaded according to their bias, with higher threshold neurons (which require more excitation to acti-vate) shaded darker. Excitatory connections are shaded gray and inhibitory connections are shaded black, with thewidth of the line proportional to the strength of the connection. The symbol above the neuron number in part B reflectsthe sign of the self-connection and the size of the symbol represents its strength. Thus, interneuron 10 has a highthreshold and strongly inhibits itself. It is strongly inhibited by sensory neurons 1, 2, 6 and 7, very weakly excited by sen-sory neurons 3–5, strongly excited by interneurons 8 and 12, and very weakly excited by interneurons 9 and 11. In turn,interneuron 10 provides modest inhibition to the other interneurons and modest excitation to motor neurons 13 and 14.

Beer Categorical Perception 233
response field is black due to the strong inhibitionfrom sensory neurons 5, 6 and 7. The remainder of thefields are produced by the pattern of excitation andinhibition from adjacent rays in a distributed fashion.Of course, the response fields and underlying synapticmechanisms for interneurons 12 and 11 are simply themirror images of those of interneurons 8 and 9 respec-tively. Most importantly, note the central blackregions in all four response fields. These are due to thefact that inhibition dominates excitation when the cen-tral five or all seven sensory neurons are active. Thisis ultimately the mechanism responsible for the cen-tral black region in the horizontal steady-state velocityfields that traps circle trajectories.
The recurrent interactions between the interneu-rons modify the feedforward response fields as shownin Plate 2B. The most striking difference is thatregions of multistability appear (green) in which thenetwork can adopt different steady-state patterns ofactivation in response to the same input depending onthe state it is in when the input occurs. The centralregion of multistability is primarily due to the mutualinhibition between interneurons 8 and 12, while theperipheral regions of multistability are produced bythe mutual inhibition between interneurons 8 and 11and between interneurons 12 and 9. In addition, theresponses of interneurons 8 and 12 are both strongerthan the feedforward responses alone due to self-exci-tation, while the responses of interneurons 9 and 11are weaker due to inhibition from inteneurons 12 and8 respectively.
Finally, each of the steady-state interneuronresponse fields produce a steady-state horizontalvelocity field via their projections to the motor neu-rons (Plate 2C). These fields are color-coded in thesame way as those in Plate 1. Note that all fourinterneurons produce motion away from the side theyare on. In all cases, this is accomplished by inhibitingthe motor neuron on the same side while exciting themotor neuron on the opposite side. Simply summingthese individual horizontal velocity fields produces anestimate of their combined effect on the agent’smotion (Plate 2D). Of course, this estimate is onlyapproximate due to the nonlinear nature of the neuralactivation function (because of the saturation in σ(x),σ(x + y) ≠ σ(x) + σ(y) in general). However, thissummed horizontal velocity field corresponds quiteclosely in both structure and intensity to the observedvelocity field (compare Plate 2D with Plate 1A, left).
Thus, the step-by-step reconstruction of the agent’ssteady-state horizontal velocity field illustrated inPlate 2 can largely account for this important featureof its dynamics.
8.2 Transient Effects
As in the previous section, the steady-state responsesof the interneurons to an object and the effects ofthose responses on motion do not give the full story.Transient effects also play an essential role. In order toillustrate the importance of these effects, I will exam-ine the neural activity underlying the termination ofactive scanning and the final catching of circles (thiswas also explored in Figure 11). Because they cannotrespond instantly to a change in input, all neuronscontribute to some extent to the neuronal inertia thatproduces scanning, as shown by the fact that modify-ing the time constants of any of them lead to deficitsin behavior. However, because all sensory signalsmust pass through the interneurons to affect behavior,we will focus on the transient responses of theinterneurons during circle trials here.
Figure 13A shows the outputs of interneurons 8,9, 11 and 12 along circle catch trajectories superim-posed over the interneurons’ feedforward responsefields (from Plate 2A). Note that, for the most part,interneurons are only active along specific segmentsof the trajectory bundles and generally only oneinterneuron is active at a time (although there is sig-nificant overlap in places). This is presumably due tothe almost universal mutual inhibition between thefour interneurons. For example, for objects that beginto the right of the agent, interneuronal activationoccurs in the following sequence: 11, 12, 9, 8, 11, 9,12. We will focus on interneuron 9, which is activeduring left-to-right scans (Note that these brightregions of activity correspond to the peaks in the α = 0plot in Figure 9).
How are scans terminated to produce a catch? Inthe last section, we saw that whether or not a trajec-tory is trapped by the x· = 0 equilibrium point corre-sponding to the central black region depends on thesystem’s state when the black region is entered (Fig-ure 11). However, what neuronal mechanism under-lies this behavior? Since the motor neurons requiredifferential activation to produce movement andinterneuron 9 is the only interneuron active duringleft-to-right scans, it stands to reason that it and its

234 Adaptive Behavior 11(4)
bilateral partner interneuron 11 are the principal pro-ducers of these scans. As can be seen in Figure 12,activity in interneuron 9 is primarily due to excitationfrom sensory neurons 1, 2 and 4, which produces thebright regions of excitation on the left side of its visualfield. Normally, activity in interneuron 9 is terminatedby inhibition from sensory neurons 5, 6 and 7 as the
trajectory bundles pass into the right half of the visualfield. However, notice that the activity during the finalleft-to-right scan is terminated early (Figure 13A,interneuron 9, white arrow). What causes this earlytermination? Unlike in previous left-to-right scans,interneuron 12 becomes active as the bundlesapproach the midline on the final scan. Since interneu-
Figure 13 Transient behavior of the interneurons during circle catch trials. (A) Motion trajectories are superimposed onthe feedforward interneuron response fields from Plate 2A. The trajectories are shaded according to the output of eachinterneuron, with black corresponding to an output of 0 and white corresponding to an output of 1. Note that, in mostcases, only one interneuron is active at a time along each trajectory bundle. An important exception to this is that in-terneurons 9/12 and 8/11 are simultaneously active on the final scan before catching. (B). Interneuron 9 activity in re-sponse to circle trials beginning on the right of the agent when the connection from interneuron 12 to interneuron 9 istransiently lesioned as soon as the trajectories enter the black region. Note that the activity in interneuron 9 now lastslonger (compare white arrow here to white arrow in the interneuron 9 plot in part A), causing the trajectories to overshootthe central black region, and these circles are now avoided.

Beer Categorical Perception 235
ron 12 strongly inhibits interneuron 9, its activityremoves the source of left-to-right movement.Interneuron 12’s activity also serves to terminate theleft-to-right motion itself, because interneuron 12excites and inhibits the opposite motor neurons frominterneuron 9.
If this explanation is correct, it would predict thattransiently lesioning the connection from interneuron12 to 9 just as the trajectories pass into the blackregion of inhibition during the final scan should allowthe trajectory bundles to pass through this regioninstead of becoming trapped there. As shown in Fig-ure 13B (white arrow), this is indeed the case. Notethat the length of time that interneuron 9 is active dur-ing the final scan is now comparable to that of earlierscans, causing the agent to move past the object andeventually produce an avoidance movement verymuch like that seen for diamonds (see Plate 1B, right),except that it is executed one scan later than usual.
9 Discussion
9.1 Summary
This paper has utilized a simpler idealized modelagent to illustrate in some detail how the perspectiveand tools of dynamical systems theory can be appliedto the analysis of embodied, situated agents capable ofminimally cognitive behavior. A 14-neuron continu-ous-time recurrent neural network was evolved toallow an agent to visually discriminate between circu-lar and diamond-shaped objects in its environment bycatching the former and avoiding the latter. The bestevolved agent used a strategy in which it foveated andactively scanned any object before eventually catch-ing or avoiding it. It was also shown that this agentexhibited labeling and discrimination behavior char-acteristic of categorical perception, that object widthwas the primary determining factor in discrimination,and that the decision to catch or avoid was a tempo-rally extended process rather than a discrete event.
The majority of the paper then focused on analyz-ing the dynamics of this evolved agent at three differentlevels: (1) the entire coupled brain/body/environmentsystem; (2) the interaction between agent and environ-ment that generates the observed coupled dynamics; (3)the underlying neuronal properties responsible for theagent dynamics. We found that the coupled dynamics
was structured into a pair of bundles that twisted aroundone another, and that these two bundles unwound withincreasing object width. Objects were caught wheneverthe two bundles collided at the axis of bilateral symme-try. These collisions underlay the category boundarybetween circles and diamonds, as well as producing ananomalous peak of catch performance. This structurealso predicted two additional anomalous peaks thatwere subsequently observed. We were then able toexplain these features of the coupled dynamics in termsof the interaction between the steady-state horizontalvelocity fields of the agent dynamics and the neuronalinertia caused by the lag of CTRNN state in response tochanging sensory inputs. Specifically, circle trajectoriesrepeatedly overshot the midline and were then turnedback toward it by blue centering regions until theybecame trapped in central black regions, whereas largerred avoidance regions ultimately pushed diamond tra-jectories away from the midline. Finally, we demon-strated how the horizontal steady-state fields wereinstantiated in the evolved CTRNN, as well as the neu-ronal interactions that underlay the trapping of circletrajectories in the central black region. This analysissuccessfully predicted that transiently lesioning a par-ticular connection at a crucial point in time would causecircles to be avoided rather than caught.
9.2 The Dynamics of Embodied, Situated Cognition
The primary technical goal of this paper has been toshow how dynamical analysis can be applied to a min-imally cognitive agent. One very important questionthat remains to be addressed is the extent to which thespecific result obtained here will generalize, both toother agents evolved on the same object discrimina-tion task and to other related tasks. Such a generaliza-tion has successfully been performed for evolvedneural circuits for walking (Chiel, Beer, & Gallagher,1999; Beer, Chiel, & Gallagher, 1999). A key step inthat generalization was a detailed study of a large set ofhigh-fitness walkers that identified the range of possi-ble solutions and their similarities and differences. Sucha generalization study remains to be performed for theobject discrimination task. Nevertheless, we can atleast begin to sketch the general picture that is sug-gested by our analysis.
First and foremost, this analysis has illustratedwhat it means to say that an agent’s behavior is a

236 Adaptive Behavior 11(4)
property only of the coupled brain/body/environmentsystem, and cannot be attributed to any one subsys-tem. In a very real sense, the evolved CTRNN doesnot “know” the difference between circles and dia-monds. It is only when embodied in its particular bodyand situated within the environment in which itevolved that this distinction arises over time throughthe interaction of these subsystems. For example, ifthe object ceases to fall, the agent produces stereo-typed responses depending on the vertical distance tothe object (Figure 5). The vertical motion of the objectas it falls and the horizontal motion of the objectthrough the agent’s field of view as the agent movescombine with neural activity to produce a temporallyextended approach or avoidance response that is morelike the gradual accumulation of small differencesthan it is a discrete decision. Even though onlyCTRNN parameters have been modified by the evolu-tionary algorithm, it is a property of the dynamics ofthe entire coupled system that has been selected for(specifically, the horizontal separation between theagent and object at the end of a trial).
A second general theme that has emerged fromthe analysis of this model agent is that, if we seek togo beyond mere characterization of the coupleddynamics to an explanation of its generation, we mustdecompose the coupled brain/body/environment sys-tem into its components and study how their interac-tions give rise to the observed behavior. The maindecomposition strategy that we have pursued here hasbeen to separate the agent from its environment andask how the structure of the environment affects theagent’s behavior and how the agent’s behavior affectsthe structure of its environment. Specifically, we char-acterized the steady-state structure of the agent’smovement field as a function of the object’s relativeposition in the agent’s field of view (Plate 1). Thiskind of steady-state decomposition can be made math-ematically rigorous when the time scale of the agentdynamics is much faster than the time scale of the sen-sory inputs from the environment. Even when thiscondition is not strictly satisfied, a great deal of quali-tative insight can often be extracted from such asteady-state decomposition.
When the timescales of the agent significantlyoverlap those of its environment, as they do here andin many biological systems, then one cannot ignorethe transient effects that arise from the lag betweenactual agent state and the equilibrium state. For exam-
ple, this lag produces the differences shown in Plate 1between the instantaneous horizontal velocity along atrajectory and the steady-state horizontal velocitygiven by the field. As the agent’s state evolves fromits current point, the agent’s resulting actions and theenvironment’s own dynamical evolution change thesensory input that the agent receives and modify thesubsequent trajectories that are available to it. In thisway, both the agent’s dynamics and that of its envi-ronment continually shape the unfolding behavioraltrajectory, as well as the future sensitivity of that tra-jectory to subsequent sensory input. As with any feed-back system, it can be very difficult to disentanglecause and effect. Effects “play through the system” tobecome causes. Indeed, the entire discriminationbehavior takes place on an extended transient of thecoupled system. This certainly does not mean that thecontribution of an agent’s dynamics to its behaviorcannot be understood. However, these results do sug-gest that we may need to modify our expectationsregarding the demands we can reasonably place upona componential explanation, stemming from the sim-ple fact that the agent and its environment are reallyjust two components of a single larger system.
The analysis described in this paper also hasimportant implications for our understanding of per-ception. Traditionally, perception has been conceivedas a process whereby an agent is informed about thestate of its environment. But, as we have seen above, adynamical agent follows a trajectory specified by itsown dynamical laws and its current state. By paramet-rically shaping those laws, sensory inputs can cer-tainly influence an agent’s behavior. They cannot,however, in general steer it along any desired path orplace it into a state uniquely corresponding to a givenstimulus. Because the behavioral consequences of agiven sensory input can differ greatly depending onthe internal state of the agent when it occurs, ongoingneural activity sets a context for perceptual processing(Arieli, Sterkin, Grinvald, & Aertsen, 1996). Forexample, although the steady-state behavior specifiedby the perception of an object approaching the midlineis the same in all four cases shown in Figure 11(namely, to stop moving), the actual behavior differsdue to differences in internal state.
Thus, it is perhaps most accurate to view percep-tion as a perturbation to an agent, with the resultingbehavior determined by the structure of the agent’sdynamics and its internal state (Maturana & Varela,

Beer Categorical Perception 237
1980). On this view, evolution structures an agent’sdynamics so that its behavior is appropriately sensi-tive or insensitive to the different classes of perturba-tion that it may encounter (indeed, the agent createsthese classes by its differential sensitivity). If theagent had been evolved to catch both circles and dia-monds rather than to discriminate between them, thenits dynamics would have had to suppress rather thanamplify the behavioral effects of the small sensorydifferences between them. Of course, given the tightcoupling between action and perception, a dynamicalagent need not sit by passively while the environmentshoves it around. Rather, it can modify its behavior soas to actively structure the perturbations it receives,making active perception a natural consequence ofthis view.
Dynamical systems theory provides a powerfulset of mathematical and computational tools for char-acterizing the dynamics of a brain/body/environmentsystem, only a few of which we have utilized here.However, there are clearly many challenges facing adynamical approach to cognition. Even in the rela-tively simple agent considered in this paper, it is stillquite impossible to fully visualize or characterize itscomplete 16-dimensional dynamics. Fortunately, sucha complete characterization is usually unnecessary.The key is to focus only on the essential degrees offreedom, since only a tiny fraction of the full statespace is actually visited during typical behavioralinteractions. For example, note that the actual catego-rization behavior of the agent analyzed in this paperoccurs in the neighborhood of the trajectory bundlesshown in Figure 9. Appropriate projections allowed usto gain considerable insight into the operation of thisevolved agent. Furthermore, many of the quantitativetools of dynamical systems theory do not require visu-alization for their application. Finally, it is likely thatnew tools will need to be developed, particularly inthe areas of systematic reduction of dimensionalityand the analysis of the non-autonomous dynamicalsubsystems that arise when a coupled brain/body/environment system is decomposed (Abarbanel &Rabinovich, 2001).
9.3 Stalking the Wily Representation
How does the concept of representation, the corner-stone of a computational theory of mind, fit into a sit-uated/embodied/dynamical explanatory scheme? It is
difficult to imagine a more controversial question inthe foundations of cognitive science than this one(Carello, Turvey, Kugler, & Shaw, 1984; Brooks,1991b; Bickhard & Terveen, 1995; Beer, 1995b;Clark & Toribio, 1995; Harvey, 1996; Ramsey, 1997;Bechtel, 1998; Clark & Grush; 1999; Wheeler &Clark, 1999; Markman & Dietrich, 2000; Keijzer,2001). Much of the debate hinges on exactly what oneis or is not willing to call a representation. Taken liter-ally, the word “representation” means re-presentation,to present again. Thus, a sketch may re-present theessentials of a scene to someone who did not see itdirectly. Traditionally, internal representations areinvoked as mediators between the outside world andinternal cognitive processes. The basic idea is thatthese internal representations stand in for (re-present)external things in a way that both carries a coherentsemantic interpretation and plays a direct causal rolein the cognitive machinery.
Because representation is such a ubiquitous andfundamental concept in cognitive science, an ongoingcritical assessment of representational notions is espe-cially important. Thus, at the risk of inflaming what isalready a sometimes acrimonious debate, it is worthunderstanding the challenges that dynamical analysesare beginning to raise for traditional representationalnotions. It is highly unlikely that any one of thesechallenges could ever definitively disprove the repre-sentation hypothesis. “Representation” is far too mal-leable a label for that. But minimally-cognitive agentsdo provide concrete examples within which we canexplore the utility of these various notions and, bystretching our intuitions about the kinds of mecha-nisms that can account for sophisticated non-reactivebehavior, they enlarge the intellectual playing field onwhich these debates are carried out. Remember that askeptical stance is supposed to be a good thing in sci-ence. Someone who runs around yelling that the sky isfalling is a trouble-maker. A skeptic just wants toknow: How do you know that the sky won’t falltoday? Do you know what’s keeping the sky up? Whatexactly do we mean by “the sky” anyway?
Consider the idea of an internal representation“standing in for” environmental entities that are notpresent in the agent’s immediate environment (e.g.,absent, hypothetical or fictitious objects, events,actions, or states of affairs), guiding behavior in theirstead (Haugeland, 1991; Clark, 1997). Historically,this aspect of internal representation was one of the

238 Adaptive Behavior 11(4)
primary theoretical concepts by which cognitiveapproaches sought to overcome the inability of thestimulus/response framework of their behavioristpredecessors to account for many features of humanbehavior (Gardner, 1985). Unlike reactive agents,the behavior of an agent possessing internal represen-tations can transcend the stimuli in its immediate envi-ronment. Even today, the limitations of reactivesystems are often used to criticize situated approachesmore generally.
Strictly speaking, however, internal state is theonly requirement for such non-reactive behavior(Beer, 1995b). If an agent has internal state, then itcan respond to the same environmental situation indifferent ways depending on that state, which in turncan depend upon its past experiences in that environ-ment. Such a dynamical agent can also initiate andcarry out sequences of actions even in the absence ofenvironmental triggers and feedback, and it canorganize its behavior in anticipation of future events.As an agent’s internal state becomes more and morecomplex, the relationship between perception andaction can become increasingly indirect. For thesereasons, merely by virtue of its internal state, anydynamical agent will produce non-reactive behaviorin general, as long as the timescale of its dynamics issufficiently slower than that of its environment. Yetfew would be willing to equate representation withinternal state, since internal state is a property of phys-ical systems in general. Finally, note that this observa-tion cuts both ways. Not only does internal state allowan agent to respond differently to the same sensorystimulus, but different sensory stimuli can place adynamical agent into the same internal state. Thismeans that internal states are not necessarily uniquelyassociated with external events; the state of the bodyand environment must also be taken into account tofully disambiguate the trajectory.
Another common claim of representationalismthat is challenged by dynamical analysis is the ideathat the units of cognitive machinery will be individu-ally semantically intelligible. The intuition here is thatthe objects and relations appearing in our conceptuali-zation of an agent’s environment and behavior mustalign in some sufficiently straightforward way withactual mechanisms internal to the agent. For example,since we conceptualize the object discrimination task interms of a “decision” to “approach” “circles” (whichare “smooth”) and “avoid” “diamonds” (which have
“sharp” “corners”), we expect to find representationsof such things within the agent, as well as processesthat manipulate and transform these representations.Indeed, a story is generally considered to be represen-tational insofar as it “depicts whole systems of identifi-able inner states (local or distributed) or processes(temporal sequences of such states) as having the func-tion of bearing specific types of information” (Clark,1997, p. 147).
There is no question that the patterns of activity ofthe interneurons play a key role in the operation of theagent analyzed here, but is this role best understood asa representational one? We have found no evidencefor circle detectors or corner detectors within thisagent’s evolved nervous system, and even though wehave shown that the agent’s behavior is sensitive toobject width, there is no evidence that any combina-tion of interneuronal activity plays the role of re-pre-senting that width. Neither does there appear to be anystraightforward neural correlate of the agent’s “deci-sion”. Rather, the interneurons are active at multipletimes during both circle and diamond trials, and inmultiple contexts (Figure 13). Whatever “meaning”this interneuronal activity has lies in the way it shapesthe agent’s ongoing behavior, and in the way it influ-ences the sensitivity of that behavior to subsequentsensory perturbations, not in coding particular fea-tures of the falling objects. For each possible internalstate and sensory input of the agent, its nervous sys-tem defines how its behavior should change. Indeed,such a vectorfield of change is one common definitionof what it means to be a dynamical system. This iswise dynamics to be sure, even dynamics for a pur-pose. But re-presentation?
Another common concern in many representa-tional theories of mind is the possibility of misrepre-sentation (see e.g., Cummins, 1989), in which aputative internal representation fails to properly trackits external referent in some circumstances. Since anydiscussion of mis-representation must presuppose anotion of re-presentation, there is little we can sayabout misrepresentation in the context of the evolvedagent. However, the related issue of classificationerror due to misbehavior is certainly relevant here,e.g., the mistakenly avoided circles and caught dia-monds in Plate 1C and the avoided circles in Figure13B. But these misclassifications are probably bestthought of as errors in dynamics that lead to a loss ofsynchronization between the agent and its environment.

Beer Categorical Perception 239
Like juggling, a successful categorization is dynami-cally assembled and maintained. While a coordinatedjuggling pattern can be destroyed by a perturbed ball ora mistimed toss, a coordinated classification behaviorcan be destroyed by a perturbed object or neuron. Butthe fact that perturbing a specific element leads to a par-ticular misbehavior can at best be taken as evidence ofa dynamical role for that element in the behavior. Itdoes not demonstrate a re-presentational role.
Since the object is always present in the agent’sfield of view, it might be argued that this categoricalperception task is not sufficiently “representation-hungry” to fully engage questions regarding the utilityof representational thinking (Clark & Toribio, 1995).There is some truth to this criticism, and it is certainlythe case that categorical perception may fall belowsome cognitive scientists’ threshold for cognitivelyinteresting behavior. However, even if cognitive inter-est is equated with the extent to which a behavior isinternally rather than externally driven, this task doesat least begin to engage the issues. As I have repeat-edly emphasized, the object alone does not drive theagent’s behavior; rather, the object’s effect on behav-ior is influenced by the agent’s evolving internal stateas the interaction proceeds. Thus, even in this simpletask, there is a certain amount of causal disconnectionbetween the object and the agent’s behavior. How-ever, it is important to further explore how the ideasdeveloped here carry over to even more offline tasks.Toward that end, ongoing work is aimed at extendingthe approach described in this paper to tasks involvingshort-term memory and selective attention, in whichthe agent’s behavior must depend in increasinglymore sophisticated ways on information that is notavailable in its immediate environment (Slocum,Downey, & Beer, 2000).
It is becoming widely recognized that traditionalnotions of representation may need to be modified inorder to do justice to the embodied, situated anddynamical facts emerging from recent work. Forexample, proposals are on the table to generalize thenotion of representation to include analog, context-dependent, action-oriented, distributed, and dynami-cal features. This raises the question of how appropri-ate the term “representation” will continue to be. Weare certainly free to redefine the term “representation”to mean anything we like, but some care must betaken in generalizing this concept. If “representation”comes to mean nothing more than correlated internal
state and “computation” comes to mean nothing morethan systematicity of behavior, then a computationaltheory of mind loses its empirical force (Beer, 1995b).
It may seem somewhat pointless to argue over aword. But the exemplar for computational theories ofcognition is still quasilinguistic symbols syntacti-cally manipulated, and by its connotations and itshistory the term “representation” carries with it pre-conceptions that may ultimately prove to be inappro-priate for understanding the mechanisms of cognitivebehavior. Thus, a little terminological hygiene seemsto be in order. If we can reliably identify semanti-cally intelligible internal configurations that can use-fully be described as re-presenting an agent’s internalgoals and external environment, then “representa-tion” it is. But if the mechanisms underlying situated,embodied cognitive behavior look nothing like re-presentations, and they act nothing like re-presenta-tions, then they are not representations, and continu-ing to call them representations is just going toconfuse everyone.
9.4 Frictionless Brains
The research methodology that has been pursued inthis paper is somewhat unusual, to say the least.Rather than studying the cognitive behavior of anexisting animal through experiment and/or modeling,I have invented an artificial agent that engages in acognitively interesting behavior and then analyzed themechanisms underlying this behavior. While suchapproaches are quite common in adaptive behaviorand artificial life research, they are still sufficientlynovel in cognitive science and neuroscience that it isworth briefly articulating the motivation behind thismethodology (Meyer, 1996; Beer, 1997; Parisi, 1997;Dean, 1998; Webb, 2000).
The integrated perspective illustrated in Figure 1raises special difficulties that any scientific approachto the neural basis of behavior must address. Studyingany one component of a brain/body/environment sys-tem is difficult enough, but studying all three compo-nents and their interactions in any animal is currentlybeyond our experimental capabilities. Not only mustone be able to control, measure and manipulate theproperties of a large fraction of an animal’s nervoussystem, but also the relevant aspects of its body andenvironment. Perhaps even more importantly, even ifthe required data were available, we currently lack the

240 Adaptive Behavior 11(4)
theoretical concepts and mathematical tools necessaryfor an integrated understanding of complete agent–environment systems. These experimental and theo-retical difficulties are only exacerbated by the com-plexity of cognitive processes.
The early theoretical development of a field typi-cally involves the careful study of simpler idealizedmodels that capture the essential conceptual featuresof the phenomena of interest. Such model systemshave a long history in physics. For example, it was notuntil Galileo’s consideration of such idealized situa-tions as frictionless planes that theoretical physics inthe modern sense of the word really began. The powerof such an idealization is that it simultaneously makesclear a deep principle of motion (acceleration, notvelocity, is proportional to force) and provides a well-defined way in which the complicating effects of fric-tion can be understood (as an external force acting onthe system). While testable quantitative predictions cansometimes be derived from simpler idealized models(e.g., when the friction coefficient of a physical surfaceis very low), their significance typically lies in the qual-itative understanding they provide rather than their abil-ity to account for specific empirical data.
Thus, in order to begin to flesh out the form that adynamical perspective on situated, embodied agentsmight take, we need to undertake a careful study ofsimpler idealized models of complete brain/body/environment systems, an endeavor that has beentermed computational neuroethology (Beer, 1990;Cliff, 1991). Instead of frictionless planes, we needfrictionless brains (and bodies and environments).Only in idealized model agents can we presently hopeto study the dynamics of a complete agent–environ-ment system and thus clarify the fundamental natureof such systems. And if we are interested in cognitivephenomena, then we must focus on model agents thatexhibit at least minimally cognitive behavior. Modelagents can help us to formulate the necessary theoreti-cal concepts, mathematical and computational tools,and experimental designs. They can serve not only asintuition pumps (Dennett, 1980), but also as theorypumps and mathematics pumps and even experimentpumps that can pave the way for more empiricallydriven analysis. They can also help to ground theongoing debates about the proper place of embodied,situated, and dynamical notions in cognitive scienceby providing concrete examples to build intuition andsharpen the discussion.
In the work described here, evolutionary algo-rithms (Mitchell, 1996) are used to evolve dynamicalnervous systems for situated model agents (Beer &Gallagher, 1992; Cliff , Harvey, & Husband, 1993;Nolfi & Floreano, 2000). There are, of course, manydifferent ways to construct simpler idealized modelagents that generate minimally cognitive behavior,and I certainly do not wish to claim that this is the onlyor best approach. However, there are certain advan-tages to this particular methodology that are worthmentioning. Continuous-time recurrent neural networksare arguably the simplest nonlinear, continuous-timeneural network model. They are relatively computa-tionally and analytically tractable, have a straightfor-ward neurobiological interpretation that captures someof the key features of nerve cells, and are universalapproximators of smooth dynamics (Funahashi &Nakamura, 1993; Kimura & Nakano, 1998). Thus, theyprovide an excellent substrate for exploring the dynam-ics of neural circuits. Evolutionary algorithms capturethe two key features of Darwinian evolution: (1) herita-ble variation; (2) differential reproduction under resourcelimitations (or natural selection). A significant advan-tage of an evolutionary approach is that, by selectingonly for the overall behavioral efficacy of the entireagent rather than a particular neuronal solution, we canminimize our a priori commitments to how a givenbehavior must be instantiated and thus better explorethe design space of possibilities that exists in a rela-tively unbiased way.
The study of simpler idealized model agents is notwithout its drawbacks. First, such models are neverlikely to make specific quantitative predictions thatare empirically testable in the way that more realisticmodels often can. However, the intent of idealizedmodels is not empirical prediction, but conceptualclarification. Detailed, realistic models are often asdifficult to understand as the original system. Clearlythe study of idealized models and empirically drivenanalysis of particular cognitive processes are comple-mentary approaches, and neither should proceed inisolation from the other. Second, there are many sub-jective decisions that must be made in order to formu-late an idealized model, such as choosing a task tostudy, choosing a body model, choosing an evolution-ary algorithm, etc. These decisions introduce theirown biases. But far from being a shortcoming, I viewthis as an opportunity. Each set of decisions consti-tutes hypotheses to be explored. Of course some of the

Beer Categorical Perception 241
choices will be wrong, but how will we know until wetry them and work through the consequences? Third,many of the details of an idealized model’s analysiswill be tied to the particular decisions made in its for-mulation, and thus be of questionable direct relevanceto any real cognitive system. But it is only by workingthrough the details of tractable concrete examples ofthe cognitive phenomena of interest that we can beginto glimpse whatever general theoretical principlesmay exist and whatever mathematical and computa-tional tools will be necessary to formulate them.
10 Conclusion
In response to a question about the relevance of think-ing of the brain as a complex dynamical system, thephilosopher of mind Patricia Churchland once replied“It’s obviously true, but so what? Then what is yourresearch program?” (Lewin, 1992, p. 164). Thepresent paper has described one possible way to beginto answer this question: (1) develop simpler idealizedmodels of minimally cognitive behavior in embodied,situated agents with as few a priori theoretical com-mitments as possible; (2) analyze the dynamics of theresulting brain/body/environment system with anopen mind about the mechanisms by which they mightoperate; (3) carefully consider the implications ofthese analyses for more realistic systems. The extentto which dynamical language will eventually becomea key element of the explanatory structure of cognitivescience remains to be seen. What does seem certain,however, is that dynamical analyses of cognitivebehavior, and the critical questions they raise, canonly broaden and clarify our understanding of themechanisms of cognition.
Acknowledgments
Portions of this analysis have been previously presented at theWorkshop on Dynamics, Computation and Cognition (held atthe Santa Fe Institute on 12–14 May 1996) and at the FourthInternational Conference on Simulation of Adaptive Behavior(held in Cape Cod, MA, USA on 9–13 September 1996). Iwould like to thank Dr Hillel Chiel and Doug Downey for theircomments on an earlier draft of this paper, as well as thereviewers. Part of this work was performed while I was on asabbatical leave at the Santa Fe Institute, and I gratefullyacknowledge their support throughout this time. This work was
also supported in part by grant RG0084/1997-B from theHuman Frontier Science Program and by grant EIA-0130773from the NSF.
References
Abraham, R. H., & Shaw, C. D. (1992). Dynamics: The geome-try of behavior. Addison–Wesley.
Abarbanel, H. D. I., & Rabinovich, M. I. (2001). Neurodynam-ics: Nonlinear dynamics and neurobiology. Current Opin-ion in Neurobiology, 11, 423–430.
Agre, P. E. (1997). Computation and human experience. Cam-bridge, UK: Cambridge University Press.
Arieli, A., Sterkin, A., Grinvald, A., & Aertsen, A. (1996).Dynamics of ongoing activity: Explanation of the largevariability in evoked cortical responses. Science, 273,1868–1871.
Arkin, R. C. (1998). Behavior-based robotics. Cambridge, MA:MIT Press.
Ashby, W. R. (1952). Design for a brain. Wiley.Bäck, T. (1996). Evolutionary algorithms in theory and prac-
tice. Oxford: Oxford University Press.Ballard, D. (1991). Animate vision. Artificial Intelligence, 48,
57–86.Bechtel, W. (1998). Representations and cognitive explana-
tions: Assessing the dynamicist’s challenge in cognitivescience. Cognitive Science, 22(3), 295–318.
Beer, R. D. (1990). Intelligence as adaptive behavior: an experi-ment in computational neuroethology. Academic Press.
Beer, R. D. (1995a). A dynamical systems perspective onagent–environment interaction. Artificial Intelligence, 72,173–215.
Beer, R. D. (1995b). Computational and dynamical languagesfor autonomous agents. In R. Port, & T. van Gelder (Eds.),Mind as motion, explorations in the dynamics of cognition(pp. 121–147). Cambridge, MA: MIT Press.
Beer, R. D. (1995c). On the dynamics of small continuous-timerecurrent neural networks. Adaptive Behavior, 3(4), 471–511.
Beer, R. D. (1996). Toward the evolution of dynamical neuralnetworks for minimally cognitive behavior. In P. Maes,M. Mataric, J. A. Meyer, J. Pollack, & S. Wilson (Eds.),From animals to animats 4: Proceedings of the FourthInternational Conference on Simulation of AdaptiveBehavior (pp. 421–429). Cambridge, MA: MIT Press.
Beer, R. D. (1997). The dynamics of adaptive behavior: Aresearch program. Robotics and Autonomous Systems, 20,257–289.
Beer, R. D. (1998). Framing the debate between computationaland dynamical approaches to cognitive science. Behavio-ral and Brain Sciences, 21(5), 630.
Beer, R. D. (2000). Dynamical approaches to cognitive science.Trends in Cognitive Sciences, 4(3), 91–99.

242 Adaptive Behavior 11(4)
Beer, R. D., & Gallagher, J. C. (1992). Evolving dynamicalneural networks for adaptive behavior. Adaptive Behavior,1, 91–122.
Beer, R. D., Chiel, H. J., & Gallagher, J. C. (1999). Evolutionand analysis of model CPGs for walking II. General prin-ciples and individual variability. J. Computational Neuro-science, 7(2), 119–147.
Bickhard, M. H., & Terveen, L. (1995). Foundational issues inartificial intelligence and cognitive science: Impasse andsolution. Elsevier.
Boyton, R. M. (1975). Color, hue and wavelength. In E. C. Car-terette, & M. P. Freidman (Eds.), Handbook of perception,volume 5 (pp. 301–347). Academic Press.
Brooks, R. A. (1991a). New approaches to robotics. Science,253, 1227–1232.
Brooks, R. A. (1991b). Intelligence without representation.Artificial Intelligence, 47, 139–159.
Carello, C., Turvey, M. T., Kugler, P. N., & Shaw, R. (1984).Inadequacies of the computer metaphor. In M. S. Gassan-iga (Ed.), Handbook of cognitive neuroscience (pp. 229–248). Plenum Press.
Cangelosi, A. , Greco, A., & Harnad, S. (2000). From robotic toilto symbolic theft: Grounding transfer from entry-level tohigher-level categories. Connection Science, 12(2), 143–162.
Chiel, H. J., Beer, R. D., & Gallagher, J. C. (1999). Evolutionand analysis of model CPGs for walking I. Dynamicalmodules. Journal of Computational Neuroscience, 7(2),99–118.
Churchland, P. S., Ramachandran, V. S., & Sejnsowski, T. J.(1994). A critique of pure vision. In C. Koch and J. Davis(Eds.), Large-scale neuronal theories of the brain (pp. 23–60). Cambridge, MA: MIT Press.
Clancey, W. J. (1997). Situated cognition: On human knowl-edge and computer representations. Cambridge, MA:Cambridge University Press.
Clark, A. (1997). Being there: Putting brain, body and worldtogether again. Cambridge, MA: MIT Press.
Clark, A., & Grush, R. (1999). Towards a cognitive robotics.Adaptive Behavior, 7(1), 5–16.
Clark, A., & Toribio, J. (1995). Doing without representing?Synthese, 101, 401–431.
Cliff, D. (1991). Computational neuroethology: A provisionalmanifesto. In J. A. Meyer, & S. W. Wilson (Eds.), Fromanimals to animats, Proceedings of the First InternationalConference on Simulation of Adaptive Behavior (pp. 29–39). Cambridge, MA: MIT Press.
Cliff, D., Harvey, I., & Husband P. (1993). Explorations inevolutionary robotics. Adaptive Behavior, 2, 73–110.
Cummins, R. (1989). Meaning and Mental Representation.Cambridge, MA: MIT Press.
Damper, R. I., & Harnad, S. R. (2000). Neural networks mod-els of categorical perception. Perception, and Psychophys-ics 62(4), 843–867.
Dean, J. (1998). Animats, & what they can tell us. Trends inCognitive Sciences, 2(2), 60–67.
Dennett, D. C. (1980). The milk of human intentionality.Behavioral and Brain Sciences, 3, 4128–430.
Ehret, G. (1987). Categorical perception of sound signals: Factsand hypotheses from animal studies. In S. Harnad (Ed.),Categorical perception: The groundwork of cognition (pp.301–331). Cambridge, UK: Cambridge University Press.
Elman, J. L. (1995). Language as a dynamical system. In R. F.Port, & T. van Gelder (Eds.), Mind as motion (pp. 195–225). Cambridge, MA: MIT Press.
Funahashi, K., & Nakamura, Y. (1993). Approximation ofdynamical systems by continuous time recurrent neuralnetworks. Neural Networks, 6, 801–806.
Gardner, H. (1985). The mind’s new science. Basic Books.Gibson, J. J. (1979). The ecological approach to visual percep-
tion. Lawrence Erlbaum.Glenberg, A. M. (1997). What is memory for? Behavioral and
Brain Sciences, 20, 1–55.Harnad, S. (Ed.) (1987). Categorical perception: The ground-
work of cognition. Cambridge, UK: Cambridge UniversityPress.
Harnad, S., Hanson, S. J., & Lubin, J. (1995). Learned categor-ical perception in neural nets: Implications for symbolgrounding. In V. Honovar, & L. Uhr (Eds.), Symbol proc-essors and connectionist network models in artificial intel-ligence and cognitive modeling: Steps toward principledintegration (pp. 191–206). Academic Press.
Harvey, I. (1996). Untimed and misrepresented. AISB Quar-terly, Winter 1996, No. 96, 20–27.
Haugel, J. (1991). Representational genera. In W. Ramsey, S.P. Stich, & D. E. Rumelhart (Eds.), Philosophy and con-nectionist theory (pp. 61–89). Lawrence Erlbaum.
Hendriks-Jansen, H. (1996). Catching ourselves in the act.Cambridge, MA: MIT Press.
Hutchins, E. (1995). Cognition in the wild. Cambridge, MA:MIT Press.
Johnson, S. D. (1994). Simple hybrid systems. InternationalJournal of Bifurcation and Chaos, 4(6), 1655–1665.
Keijzer, F. (2001). Representation and behavior. Cambridge,MA: MIT Press.
Kelso, J. A. S. (1995). Dynamic patterns: The self-organizationof brain and behavior. Cambridge, MA: MIT Press.
Kimura, M., & Nakano, R. (1998). Learning dynamical sys-tems by recurrent neural networks from orbits. NeuralNetworks, 11, 1589–1599.
Lakoff, G., & Johnson, M. (1998). Philosophy in the flesh.Basic Books.
Lewin, R. (1992). Complexity: Life at the edge of chaos. Mac-millan.
Liberman, A. M., Cooper, F. S., Shankweiler, D. P., & Stud-dert-Kennedy, M. (1967). Perception of the speech code.Psychological Review, 74, 431–461.

Beer Categorical Perception 243
Markman, A. B., & Dietrich, E. (2000). In defense of represen-tation. Cognitive Psychology, 40, 138–171.
Maturana, H. R., & Varela, F. J. (1980). Autopoiesis and cogni-tion. D. Reidel.
Meyer, J. A. (1996). Artificial life and the animat approach toartificial intelligence. In M. Boden (Ed.), Artificial intelli-gence (pp. 325–354). Academic Press.
Mitchell, M. (1996). An introduction to genetic algorithms.Cambridge, MA: MIT Press.
Nolfi, S., & Floreano, D. (2000). Evolutionary Robotics. Cam-bridge, MA: MIT Press.
Parisi, D. (1997). Artificial life and higher level cognition.Brain and Cognition, 34, 160–184.
Pfeifer, R., & Scheier, C. (1999). Understanding intelligence.Cambridge, MA: MIT Press.
Platt, M. L. (2002). Neural correlates of decisions. CurrentOpinion in Neurobiology, 12, 141–148.
Port, R. F., & van Gelder, T. (Eds.) (1995). Mind as motion:Explorations in the dynamics of cognition. Cambridge,MA: MIT Press.
Ramsey, W. (1997). Do connectionist representations earn theirexplanatory keep? Mind and Language, 12(1), 34–66.
Skarda, C., & Freeman, W. J. (1987). How brains make chaosin order to make sense of the world. Behavioral and BrainSciences, 10, 161–195.
Slocum, A. C., Downey, D. C., & Beer, R. D. (2000). Furtherexperiments in the evolution of minimally cognitivebehavior: From perceiving affordances to selective atten-tion. In J. Meyer, A. Berthoz, D. Floreano, H. Roitblat, &S. Wilson (Eds.), From animals to animats: Proceedingsof the Sixth International Conference on the Simulation ofAdaptive Behavior (pp. 430–439). Cambridge, MA: MITPress.
Strogatz, S. H. (1994). Nonlinear dynamics and chaos. Addi-son-Wesley.
Studdert-Kennedy, M., Liberman, A. M., Harris, K. S., &Cooper, F. S. (1970). Motor theory of speech perception:A reply to Lane’s critical review. Psychological Review,77, 234–249.
Suchman, L. A. (1987). Plans and situated actions. Cambridge,UK: Cambridge University Press.
Thelen, E., & Smith, L. B. (1994). A dynamic systemsapproach to the development of cognition and action.Cambridge, MA: MIT Press.
Thelen, E., Schöner, G., Scheier, C., & Smith, L. B. (2001).The dynamics of embodiment: A field theory of infantperseverative reaching. Behavioral and Brain Sciences,24, 1–86.
Tijsseling, A., & Harnad, S. (1997). Warping similarity spacein category learning by brackprop nets. In M. Ramscar, U.Hahn, E. Cambouropolos, & H. Pain (Eds.) Proceedingsof SimCat 1997: Interdisciplinary Workshop on Similarityand Categorization (pp. 263–269). Edinburgh, UK:Department of Artificial Intelligence, Edinburgh Univer-sity.
Townsend, J. T., & Busemeyer, J. (1995). Dynamic representa-tion of decision-making. In R. Port, & T. van Gelder(Eds.), Mind as motion: Explorations in the dynamics ofcognition (pp. 101–120). Cambridge, MA: MIT Press.
van Gelder, T. (1998). The dynamical hypothesis in cognitivescience. Behavioral and Brain Sciences, 21(5), 615–665.
Varela, F. J., Thompson, E., & Rosch, E. (1991). The embodiedmind. Cambridge, MA: MIT Press.
Webb, B. (2000). What does robotics offer animal behaviour?Animal Behaviour, 60, 545–558.
Wheeler, M., & Clark, A. (1999). Genic representation: Recon-ciling content and causal complexity. British Journal forthe Philosophy of Science, 50, 103–135.
Wyttenbach, R. A., May, M. L., & Hoy, R. R. (1996). Categor-ical perception of sound frequency by crickets. Science,273, 1542–1544.
About the Author
Randall D. Beer is a professor of Electrical Engineering and Computer Science at CaseWestern Reserve University, with a joint appointment in the Department of Biology. Hisresearch is focused on the modeling and analysis of brain/body/environment systems,and its implications for neuroscience, cognitive science and robotics.