the ecology and evolution of stoichiometric phenotypes · known about the ecological and...
TRANSCRIPT

TrendsThere is growing evidence for intraspe-cific variation in stoichiometric traits ofanimals, for a genetic basis of this var-iation, and for rapid evolution of thesetraits. In light of such observations,ecological stoichiometry has the poten-tial to address evolutionary processesand identify mechanisms by whichenvironmental heterogeneity drivesthe origins of biological diversity.
We suggest that addressing the evolu-tion of stoichiometric reaction-norms
TREE 2192 No. of Pages 10
OpinionThe Ecology and Evolution ofStoichiometric PhenotypesMiguel C. Leal [11_TD$DIFF],,1,* Ole Seehausen,1,2 and Blake Matthews3
Ecological stoichiometry has generated new insights into how the balance ofelements affects ecological interactions and ecosystem processes, but little isknown about the ecological and evolutionary dynamics of stoichiometrictraits. Understanding the origins and drivers of stoichiometric trait variationbetween and within species will improve our understanding about the eco-logical responses of communities to environmental change and the ecosys-tem effects of organisms. In addition, studying the plasticity, heritability, andgenetic basis of stoichiometric traits might improve predictions about howorganisms adapt to changing environmental conditions, and help to identifyinteractions and [12_TD$DIFF]feedbacks between phenotypic evolution and ecosystemprocesses.
could help in understanding when,and to what extent, variability in elemen-tal phenotype in natural populations isdue to plasticity of stoichiometric traitsor genetic variation. We also proposestrengthening the link between stoichio-metric trait variability and fitness bybuilding on nutritional geometry theoryand function-valued methods. In addi-tion, and because stoichiometric traitsmay not only respond to environmentalvariation but also influence ecosystemdynamics, studying the elemental phe-notypes can contribute to our under-standing of the nature of eco-evolutionary [12_TD$DIFF]feedbacks.
1Department of Fish Ecology andEvolution, Swiss Federal Institute ofAquatic Science and Technology(Eawag), Center for Ecology,Evolution, and Biogeochemistry, 6047Kastanienbaum, Switzerland2Division of Aquatic Ecology andEvolution, Institute of Ecology andEvolution, University of Bern, 3012Bern, Switzerland3Department of Aquatic Ecology,Eawag, Center for Ecology, Evolution,and Biogeochemistry, 6047Kastanienbaum, Switzerland
*Correspondence:[email protected] (M.C. Leal,).
Ecological Stoichiometry: A Functional Trait PerspectiveThere is a growing interest in understanding intraspecific functional trait variability so as to makebetter predictions about population dynamics, species interactions, and ecosystem functioning[1,2]. Although stoichiometric traits (see Glossary) are useful for connecting organisms toecosystem processes [3–5], little is known about how intraspecific variation in these traitsmediates ecological and evolutionary dynamics. Stoichiometric traits are an integral part ofecological stoichiometry (ES) theory, which focuses on the balance of multiple elements inecological interactions and ecosystem processes [6]. Stoichiometric traits can vary amongspecies [5–7], populations, and individuals of the same species [8–10], can influence howorganisms affect energy flow and nutrient cycling in ecosystems (i.e., as functional effect traits)[6,11], and can mediate evolutionary responses to environmental variation and change (i.e., asfunctional response traits) [4,5]. Recent studies have also begun to document the occurrence[8,12,13] and ecological consequences [3,14] of rapid evolution of stoichiometric traits. Overall,such stoichiometric traits are a useful focal point for studying both interactions and [12_TD$DIFF]feedbacksbetween phenotypic evolution and ecosystem processes [4_TD$DIFF].
ES theory posits that performance (e.g., growth rate) is partly determined by the match betweenthe stoichiometry of resources and organism elemental phenotype (EP), which comprisesmultiple stoichiometric traits. However, what is the relationship between stoichiometric traitsand fitness variation, and, more generally, how do organisms adapt to elemental imbalances intheir diet or their resource uptake? Increasing nutrient supply can impact on the evolution of life-history traits by changing allocation trade-offs, particularly those associated with nutrientimbalances that negatively influence performance [15,16]. For example, we might expect eitherdirectional selection on stoichiometric traits to reduce elemental imbalances between a popu-lation and its resources [15,17], or fluctuating selection to maintain stoichiometric trait variationwithin populations when there is variation in resource quality through time. It is also possible thatagents of selection on stoichiometric traits vary in their importance over the lifetime of organisms[15]. Juveniles and adults often vary in their nutritional needs because the nutritional demandsfor rapid growth (e.g., high dietary phosphorus, P) early in life might differ from those for
Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy http://dx.doi.org/10.1016/j.tree.2016.11.006 1© 2016 Elsevier Ltd. All rights reserved.

TREE 2192 No. of Pages 10
GlossaryGrowth rate hypothesis (GRH):predicts that rapid growth often hascomparatively higher P demand tosupport increased allocation to P-richribosomal RNA to meet the demandsof protein synthesis.Heritability: variation of a trait that isexplained by genetic differences. Itcan be calculated as the ratio ofgenetic to total variance (includingenvironmental).Homeostasis: physiologicalmechanisms that maintain the internalenvironment of an organism in arelatively constant state. In ecologicalstoichiometry, homeostatic processesreduce variation in the elementalcomposition of an organism despitechanges in resource stoichiometry.Nutrition: interaction of nutrientsavailable in food with physiologicalfunctions of an organism(metabolism, growth, reproduction,health, and disease) that affectresource ingestion, assimilation, andexcretion.Phenotypic plasticity: a singlegenotype expresses multiplephenotypes in response toenvironmental cues, with eachphenotype matching a differentselective optimum. The range ofphenotypes produced by a particulargenotype exposed to differentenvironmental conditions can beassessed using reaction norms.Stoichiometric traits: individualmeasures of elemental composition,assimilation, allocation, or excretion
reproduction later in life (e.g., dietary lipids). Overall, the fitness consequences of maintaining (ornot maintaining) stoichiometric homeostasis remain poorly understood.
We discuss here how stoichiometric traits can be an important intersection between ecologicalinteractions, evolutionary dynamics, and ecosystem processes. To illustrate this, we review theprimary drivers of intraspecific variation in the EP and the evidence that stoichiometric traits canevolve, potentially via multiple agents of natural selection.We advocate a better understanding ofhow stoichiometric trait variability [e.g., %P, %nitrogen (N), and the N:P ratio] relates to fitnessvariation, and suggest that studying the evolution of stoichiometric reaction norms might help todisentangle the role of phenotypic plasticity and genetic variation in determining the EP. Wehighlight how approaches from nutritional geometry (e.g., [18,19]) and function-valued methods(e.g., [20,21]) [5_TD$DIFF] can help [13_TD$DIFF]strengthening the link between fitness variation and elemental imbalancesbetween consumers and their resources. We end by discussing how stoichiometric traits canimprove our understanding of eco-evolutionary dynamics and [12_TD$DIFF]feedbacks between ecology andevolution.
EP Variability: Causes and ConsequencesThe EP is made up of multiple stoichiometric traits associated with the composition, acquisition,assimilation, allocation, and excretion of key elements of an individual organism [22] (Box 1).Autotrophs are more stoichiometrically variable than heterotrophs owing to their rapid responseto nutrient availability and ability to store nutrients [6]. For heterotrophs, there is growing evidenceof variation in homeostasis among species, among individuals within populations [4,9], andwithin individuals throughout their ontogeny [10,17].
Variation in EP between and within populations is often attributed either to spatial heterogeneityin resource quantity and quality [9,23], or to non-spatial sources of intraspecific variation in EP,including body size, diet, morphology, and sex [7,10,24]. In addition, variation in EP amongindividuals has been attributed to variation in the cellular concentration (e.g., by weight) of nucleicacids, such as RNA and DNA, particularly when there is variation in ploidy or in elementalallocation associated with growth [12,25–27]. Such genomic variation at the cellular level canaffect organismal stoichiometry, and thereby shape the nutritional quality of organisms (either asprey or detritus), and modify energy fluxes through the food web [11].
that are usually defined by theirelemental content or ratios betweenelements.
Box 1. Agents and Targets of Selection
Phenotypic traits are defined as quantifiable measurements of an organism and can evolve when there is covariancebetween phenotype and fitness. The stoichiometric traits that define the elemental phenotype (EP) of an organism canboth evolve in response to environmental selection and modify the environmental conditions that might determineselection pressures.
The EP is made up of several traits, such as organismal elemental composition (e.g., % of dry mass or the ratio ofelements), which determines how acquired nutrients are used and recycled. These stoichiometric traits might affect thesurvival, growth, and reproduction of individuals, and consequently be targets of selection. Potential selection agentssuch as nutrient availability (i.e., food quantity and/or quality) and predation pressure are likely to affect fitness throughchanges in element allocation from routine metabolic processes to demanding physiological traits occurring duringgrowth and reproduction, which can also reroute energy allocation away from homeostatic processes. Food qualityaffects the growth rate of consumers [1_TD$DIFF] and therefore can be considered [2_TD$DIFF] a selection agent. The selection targets would bethe physiological mechanisms driving nutritional demand that are affected by food quality. Another target of selectionmight be genome size because it is composed of nucleotides and amino acids that require key elements, such as N andP, which are reallocated from DNA to RNA to sustain rapid growth [26,58]. Consequently, the N and P content of foodhave been suggested to act as selection agents in nutrient-limited environments because they can mediate the relativesuccess of individuals differing in genome size, particularly between diploid and polyploid organisms [11,25]. Parasitismmight also be a selection agent driving the evolution of EPs through nutrient-driven within-host infection dynamics [59].
2 Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy
TREE 2192 No. of Pages 10
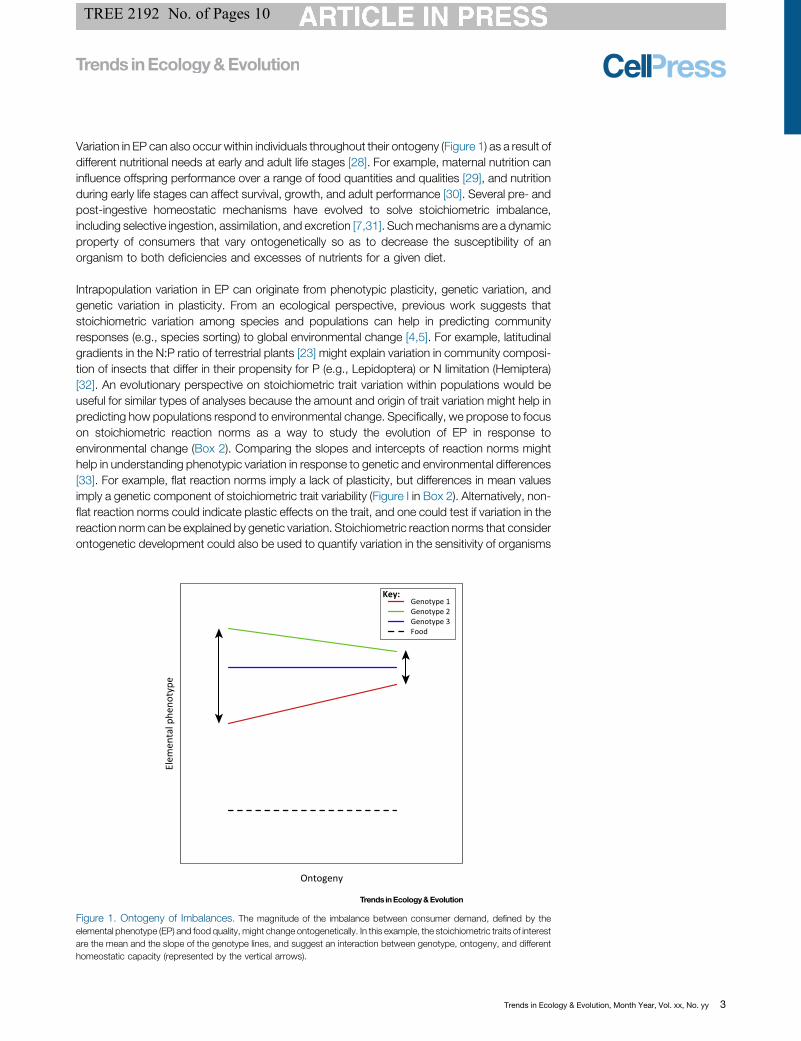
Variation in EP can also occur within individuals throughout their ontogeny [14_TD$DIFF] (Figure 1) as a result ofdifferent nutritional needs at early and adult life stages [28]. For example, maternal nutrition caninfluence offspring performance over a range of food quantities and qualities [29], and nutritionduring early life stages can affect survival, growth, and adult performance [30]. Several pre- andpost-ingestive homeostatic mechanisms have evolved to solve stoichiometric imbalance,including selective ingestion, assimilation, and excretion [7,31]. Suchmechanisms are a dynamicproperty of consumers that vary ontogenetically so as to decrease the susceptibility of anorganism to both deficiencies and excesses of nutrients for a given diet.
Intrapopulation variation in EP can originate from phenotypic plasticity, genetic variation, [15_TD$DIFF]andgenetic variation in plasticity. From an ecological perspective, previous work suggests thatstoichiometric variation among species and populations can help in predicting communityresponses (e.g., species sorting) to global environmental change [4,5]. For example, latitudinalgradients in the N:P ratio of terrestrial plants [23] might explain variation in community composi-tion of insects that differ in their propensity for P (e.g., Lepidoptera) or N limitation (Hemiptera)[32]. An evolutionary perspective on stoichiometric trait variation within populations would beuseful for similar types of analyses because the amount and origin of trait variation might help inpredicting how populations respond to environmental change. Specifically, we propose to focuson stoichiometric reaction norms as a way to study the evolution of EP in response toenvironmental change (Box 2). Comparing the slopes and [16_TD$DIFF]intercepts of reaction norms mighthelp in understanding phenotypic variation in [17_TD$DIFF]response to genetic and environmental differences[33]. For example, flat reaction norms imply a lack of plasticity, but differences in mean valuesimply a genetic component of stoichiometric trait variability (Figure I in Box 2). Alternatively, non-flat reaction norms could indicate plastic effects on the trait, and one could test if variation in thereaction norm can be explained by genetic variation. Stoichiometric reaction norms that considerontogenetic development could also be used to quantify variation in the sensitivity of organisms
Ontogeny
Elem
enta
l phe
noty
pe
Genotype 1Genotype 2Genotype 3Food
Key:
Figure 1. Ontogeny of Imbalances. The magnitude of the imbalance between consumer demand, defined by theelemental phenotype (EP) and food quality, might change ontogenetically. In this example, the stoichiometric traits of interestare the mean and the slope of the genotype lines, and suggest an interaction between genotype, ontogeny, and differenthomeostatic capacity (represented by the vertical arrows).
Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy 3
TREE 2192 No. of Pages 10
to elemental imbalance throughout ontogeny. In practice it is difficult to have continuous accessto the complete ontogenetic EP through time, but breeding and rearing different genotypes incommon-garden experiments could be a powerful way to identify how much phenotypicvariation throughout ontogeny is explained by genetic differences.
Link Stoichiometric Traits to FitnessOne fundamental challenge for understanding the evolution of the EP is to establish a better linkbetween EP and fitness. To start, it is important to recognize that ES studies typically use carbon(C) as a proxy for biomass to track energy flows between organisms and their environment.However, in most cases [18_TD$DIFF] components of the body that are considered nutrients are more likely tobe determinants of fitness variation than solely the elements and their ratios (e.g., C:P, N:P). In
Box 2. Stoichiometric Reaction Norms
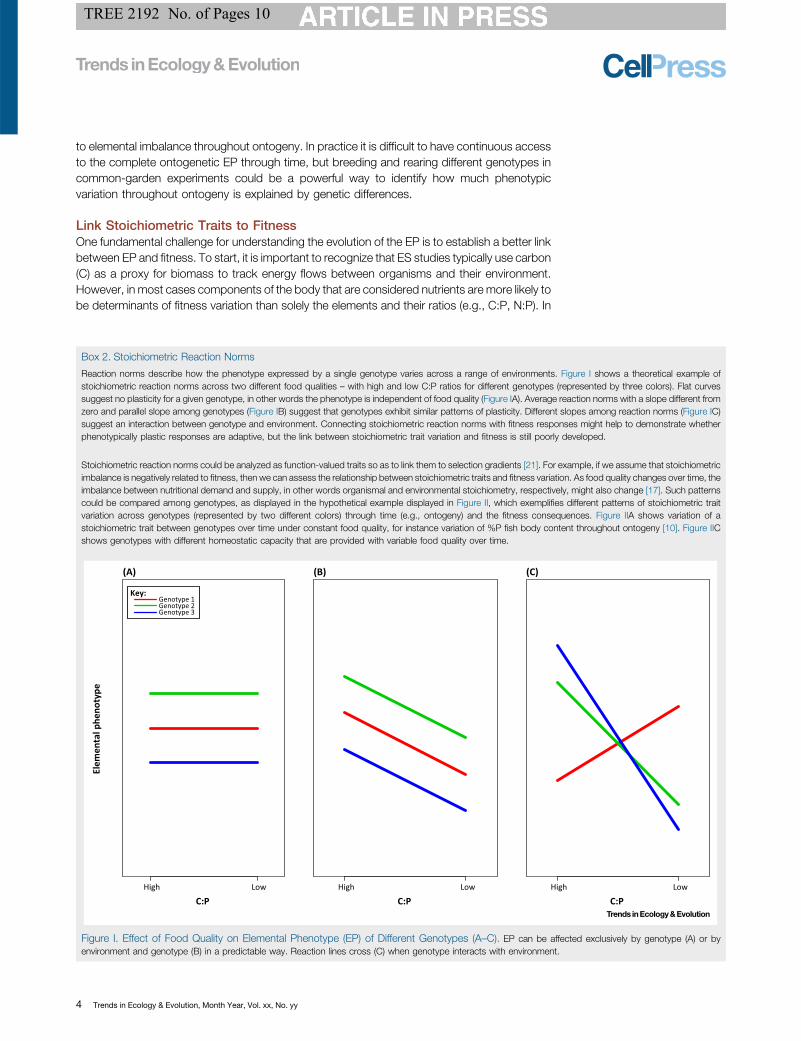
Reaction norms describe how the phenotype expressed by a single genotype varies across a range of environments. Figure I shows a theoretical example ofstoichiometric reaction norms across two different food qualities – with high and low C:P ratios for different genotypes (represented by three colors). Flat curvessuggest no plasticity for a given genotype, in other words the phenotype is independent of food quality (Figure IA). Average reaction norms with a slope different fromzero and parallel slope among genotypes (Figure IB) suggest that genotypes exhibit similar patterns of plasticity. Different slopes among reaction norms (Figure IC)suggest an interaction between genotype and environment. Connecting stoichiometric reaction norms with fitness responses might help to demonstrate whetherphenotypically plastic responses are adaptive, but the link between stoichiometric trait variation and fitness is still poorly developed.
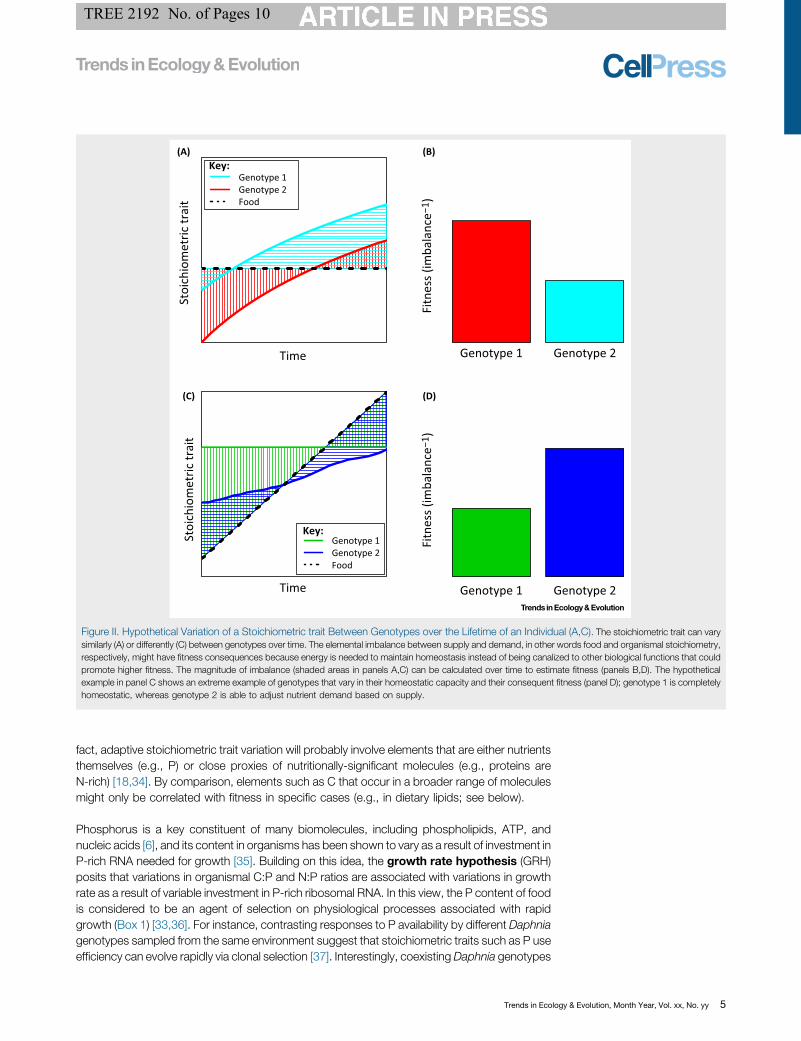
Stoichiometric reaction norms could be analyzed as function-valued traits so as to link them to selection gradients [21]. For example, if we assume that stoichiometricimbalance is negatively related to fitness, then we can assess the relationship between stoichiometric traits and fitness variation. As food quality changes over time, theimbalance between nutritional demand and supply, in other words organismal and environmental stoichiometry, respectively, might also change [17]. Such patternscould be compared among genotypes, as displayed in the hypothetical example displayed in Figure II, which exemplifies different patterns of stoichiometric traitvariation across genotypes (represented by two different colors) through time (e.g., ontogeny) and the fitness consequences. Figure IIA shows variation of astoichiometric trait between genotypes over time under constant food quality, for instance variation of %P fish body content throughout ontogeny [10]. Figure II [8_TD$DIFF]Cshows genotypes with different homeostatic capacity that are provided with variable food quality over time.
C:P
Elem
enta
l phe
noty
pe
LowHigh
(A)
Genotype 1Key:
Genotype 2Genotype 3
C:PLowHigh
(B)
C:PLowHigh
(C)
Figure I. Effect of Food Quality on Elemental Phenotype (EP) of Different Genotypes (A–C). EP can be affected exclusively by genotype (A) or byenvironment and genotype (B) in a predictable way. Reaction lines cross (C) when genotype interacts with environment.
4 Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy
TREE 2192 No. of Pages 10
Time
Stoi
chio
met
ric tr
ait
(A)
Genotype 1Key:
Key:
Genotype 2Food
Fitn
ess (
imba
lanc
e−1 )
(B)
Time
Stoi
chio
met
ric tr
ait
Genotype 1Genotype 2Food
(C)
Genotype 2Genotype 1
Genotype 2Genotype 1
Fitn
ess (
imba
lanc
e−1 )
(D)
Figure II. Hypothetical Variation of a Stoichiometric trait Between Genotypes over the Lifetime of an Individual (A,C). The stoichiometric trait can varysimilarly (A) or differently (C) between genotypes over time. The elemental imbalance between supply and demand, in other words food and organismal stoichiometry,respectively, might have fitness consequences because energy is needed to maintain homeostasis instead of being canalized to other biological functions that couldpromote higher fitness. The magnitude of imbalance (shaded areas in panels A,C) can be calculated over time to estimate fitness (panels B,D). The hypotheticalexample in panel C shows an extreme example of genotypes that vary in their homeostatic capacity and their consequent fitness (panel D); genotype 1 is completelyhomeostatic, whereas genotype 2 is able to adjust nutrient demand based on supply.
fact, adaptive stoichiometric trait variation will probably involve elements that are either nutrientsthemselves (e.g., P) or close proxies of nutritionally-significant molecules (e.g., proteins areN-rich) [18,34]. By comparison, elements such as [19_TD$DIFF]C that occur in a broader range of moleculesmight only be correlated with fitness in specific cases (e.g., in dietary lipids; see below).
Phosphorus is a key constituent of many biomolecules, including phospholipids, ATP, andnucleic acids [6], and its content in organisms has been shown to vary as a result of investment inP-rich RNA needed for growth [35]. Building on this idea, the growth rate hypothesis (GRH)posits that variations in organismal C:P and N:P ratios are associated with variations in growthrate as a result of variable investment in P-rich ribosomal RNA. In this view, the P content of foodis considered to be an agent of selection on physiological processes associated with rapidgrowth (Box 1) [33,36]. For instance, contrasting responses to P availability by different Daphniagenotypes sampled from the same environment suggest that stoichiometric traits such as P useefficiency can evolve rapidly via clonal selection [37]. Interestingly, coexisting Daphnia genotypes
Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy 5

TREE 2192 No. of Pages 10
can differ strongly in their reproductive output over a broad range of dietary C:P ratios (e.g., 100to 800) [38], suggesting that nutrient availability could be an important dimension of life-historyevolution [15]. Although there is strong support for the GRH for invertebrates [35,39], less isknown about the relationship between EP and fitness for organisms that store P in tissues withslow turnover, for example vertebrates that store P in bone [6].
[20_TD$DIFF]Nitrogen is one of the main constituents in biological structural molecules, and tissue Ncontent is often used as a proxy for proteins [6]. Several abiotic and biotic factors have beenshown to affect N-based stoichiometric traits for a variety of organisms, ranging from primaryproducers to aquatic and terrestrial consumers [8]. N limitation can have important fitnessconsequences, particularly for herbivores because [21_TD$DIFF]the N content [22_TD$DIFF] of herbivores like that of allheterotrophs is notably higher than that of autotrophs [32,40]. This stoichiometric imbalancein herbivores has likely led to different adaptations to N-poor food, such as producingbiomass using low-N materials (e.g., chitin instead of protein), or modifying elementalallocation to different body parts that differ in N content [41]. N limitation might also affectperformance across life-history stages [42] or lead to the evolution of novel trophic inter-actions, such as omnivory or cannibalism, as alternative ways to meet nutrient demand andincrease reproductive output [43,44].
Carbon occurs in multiple nutrients, such as lipids and carbohydrates, and therefore might co-vary with C-rich macro-compounds, such as dietary fatty acids [45]. For instance, variation in theratio of essential fatty acids to C content of seston can strongly affect growth rate and eggproduction in Daphnia [46]. This is often observed in lakes where the dominance of cyanobac-teria in phytoplankton communities, which lack important sterols and long-chain polyunsatu-rated fatty acids, directly affects Daphnia consumers and has driven specific adaptations [47].However, establishing a direct link between organismal C-based stoichiometric traits and fitnessis likely more difficult for other lipid molecules [23_TD$DIFF]because different lipids can vary in their biologicalfunction but have similar C content.
A useful way to link stoichiometric traits with fitness variation is to build on approaches fromnutritional geometry (Box 3). This approach uses a multidimensional nutrient space where eachdimension represents a different nutrient [18,19,48–51]. A standard approach in nutritionalgeometry is to rear animals on a range of food qualities (e.g., different protein and carbohydratecontents are represented in each axis) which delimit an area in nutritional space. The location ofthe intake target within the available nutrient space represents the optimal combination ofnutrients that fulfill the requirements of the animal. With this approach, consumer performanceover a range of different nutritional scenarios can be measured as development rates orreproductive output. By doing so, one can assess the relationship between specific nutrients,and their elemental composition, and variation in fitness components [48,49]. Such approachescould also be extended by assessing how different genotypes vary in their performance within amultidimensional space determined by stoichiometric trait axes. This can be done for differentlife-history stages bymeasuring different fitness components, such as growth for early life stagesand reproductive output for adults (Figure I in Box 3).
Establishing a link between stoichiometric traits and variation in fitness components throughoutontogeny is usually performed separately for different life stages because proxies of fitness areoften quantified differently for juveniles and adults. This could be improved by using homeostasisas a continuous [24_TD$DIFF] proxy of one fitness [25_TD$DIFF]component through the life of an organism (Figure [26_TD$DIFF]II in Box2). This approach would assume that stoichiometric imbalance between consumers andresources is negatively correlated with performance, and account for ontogenetic changesin homeostatic mechanisms [27_TD$DIFF] (Figure 1) as well as variability in resource quality and quantity. Suchcomplex functions could be analyzed using a function-valued trait approach [20,21].
6 Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy

TREE 2192 No. of Pages 10
Stoichiometric Eco-Evolutionary DynamicsMultiple agents of selection can drive the evolution of stoichiometric traits [8], but it is stillunknown when this phenotypic evolution might affect biotic and abiotic conditions, and therebyinfluence prevailing environmental sources of natural selection [3]. Variability in resource demandby heterotrophs can affect nutrient recycling in natural ecosystems, as has been observed forfish with high P content that display low excretion rates [52,53]. In such cases, stoichiometrictrait variation in the community [54], or at the population level [55], can change the balance offreely-available nutrients in the environment. Such changes to nutrient availability might subse-quently affect the ecological and evolutionary dynamics of both autotroph and heterotrophpopulations that vary in their nutrient use physiology and elemental composition [11,26,36].
The dual role of stoichiometric traits as both functional effect and response traits makes themespecially likely to be involved in [12_TD$DIFF]feedbacks between phenotypic evolution and ecosystemprocesses. Models integrating ES with eco-evolutionary dynamics have begun to illustrate howthe evolution of EPs can influence different levels of biological organization, from individuals topopulations, communities, and ecosystems [3]. Although there are clear ES examples that theEP of animals can modify the environment [8], and that the evolution of EPs can be driven byenvironmental change [37], a better understanding of how stoichiometric traits evolve inresponse to natural selection will be useful to identify potential feedback between ecological,evolutionary, and ecosystem processes. One completely open question, for instance, concerns
Box 3. Application of a Geometric Approach to ES
In general, ES still lacks an approach to link stoichiometric traits to fitness and selectionmechanisms that fit a broad rangeof organisms. Mechanisms such as selective feeding, differential assimilation of ingested nutrients, selective excretion ofnutrients in excess, and nutrient storage have likely evolved in response to nutrient availability [60]. However, the evolutionof stoichiometric-related traits and the mechanisms to obtain, retain, and concentrate resources throughout life remainunclear. ES has made progress toward establishing the link between elements and growth rate as a key life-history traitthat is associated with fitness (e.g., GRH). However, ES could benefit from approaches such as nutritional geometry [18],where elemental ratios can be used inmultidimensional geometric models to assess fitness across elemental phenotypes [9_TD$DIFF](Figure I), and from the analysis of function-valued traits (Box 2) to study adaptive plasticity. Variation in food qualityrequirements through life (e.g., early stages require more P for growth, and late stages requiremore C for energy storage),together with variability in available resources [10_TD$DIFF] (Figure II), might change the strength and directionality of selection [3_TD$DIFF].
Early stage
%Phosphorus%Nitrogen
Gro
wth
Adult
%Phosphorus%Nitrogen
Repr
oduc
�ve
outp
ut
Figure I. Hypothetical 3DModel ShowingOrganismal Fitness Response to FoodQuality Represented by Nand P Content (Following a Nutritional Geometry Approach [18]). Note that in this example stoichiometric traits(%N and %P) are used instead of macromolecules (e.g., carbohydrates, proteins). Different optimal scenarios may beobserved depending on nutritional strategies and experience throughout life history. In the hypothetical scenario shownhere, early stages notably benefit (i.e., grow faster) from increased food quality, whereas adults do not benefit.
Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy 7

TREE 2192 No. of Pages 10
Early stage
Gro
wth
Adult
Repr
oduc
�ve
outp
ut
Food quality
Figure II. Two-dimensional Cut of the Plots Shown in Figure I Noting Different Performances of Early Stagesand Adults in Response to Variation in FoodQuality. This hypothetical scenario suggests that food quality is crucialfor early stages to grow and allocate high-quality food into somatic tissues. Adult stages might not be equally sensitive tofood quality because increased reproductive output is likely associated with non-nutrient-related attributes such asmorphological or behavioral traits.
the genetic architecture of EP variation, and therefore howmuch EP variation can actually evolvewithin a single panmictic population. This leads to the question of how speciation and speciesrichness within ecosystems affects EP variation, a potential theme for future speciation andbiodiversity research.
Two recent studies have addressed the rapid evolution of stoichiometric traits that are involvedin eco-evolutionary feedbacks. In a theoretical study, Yamamichi et al. [8] used a modelingapproach to show that the rapid evolution of EP affects the nutrients available in the environ-ment, and that this affected evolutionary processes. Consumers that rapidly evolved to newnutrient conditions caused a feedback that ultimately affected both adaptive evolution andpopulation stability. In an elegant and unique experimental study, Declerck et al. [14] showedthat phenotypic variation in growth rates and nutrient demand of a herbivorous rotifer affectednutrient recycling, particularly when P-limited food was provided. The subsequent changes inresource availability favored the phenotypes with a selection history of P limitation, therebydriving evolutionary processes and consequent feedback on standing stocks of primaryproduction. This ultimately caused changes in population structure, grazing pressure, andthe biomass ratio of consumers and resources. Both [7_TD$DIFF] studies show the potential involvement ofstoichiometric traits in eco-evolutionary dynamics and feedbacks.Moving forward, we can gaina better mechanistic understanding of the reciprocal interactions between EP variation andecosystem dynamics by experimentally testing (i) how much individual organisms within asingle genetic population can vary in their stoichiometry, (ii) how organisms with differentstoichiometries affect ecosystem dynamics, (iii) how nutrient limitation and organismal elemen-tal imbalance relate to fitness variation, and (iv) how organism-mediated modifications to theenvironment, particularly those governed by stoichiometric traits, affect fitness variation insubsequent generations [56].
8 Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy

TREE 2192 No. of Pages 10
Outstanding QuestionsHow variable are stoichiometric traitsacross ontogeny, and what are thefitness consequences of homeostaticimbalance resulting from variation infood quality?
What are the primary agents of selec-tion that drive stoichiometric traitevolution?
How does selection driven by nutrientlimitation act at the genomic, proteo-mic, and metabolomic levels, and whatare the consequences for individuals,populations, and ecosystems?
How and when do ecosystem changesmediated by stoichiometric traits oforganisms lead to eco-evolutionaryfeedbacks?
Concluding Remarks and Future DirectionsThere is growing evidence that the EP can be highly variable across species, populations, andindividuals, and can evolve in response to natural selection, but the effect of EP variation onecosystems and its reciprocal effect on phenotypic evolution are still poorly understood[28_TD$DIFF] (seeOutstanding Questions). A necessary future step for ES to address this gap will be to strengthenthe link between stoichiometric traits and heritable fitness using approaches other than thealready established link between P and growth rate [35]. The recent integration between ES andnutritional geometry [48,49] can be extended to study how stoichiometric trait variation is relatedto fitness. The link between homeostasis and fitness can also be integrated with other frame-works encompassing homeostatic regulation in animals. For instance, Dynamic Energy Budgettheory (DEB), which provides simple mechanistic rules describing the uptake and use of energyand nutrients [57], could be better integrated with ES [48] and across different organizationallevels. Such an approach might provide a better mechanistic basis for understanding andpredicting population and ecosystem responses to stoichiometric constraints of individuals, anduncover new mechanisms underlying eco-evolutionary [12_TD$DIFF]feedbacks. Further progress integratingES with other frameworks that address evolutionary dynamics will improve our understanding ofhow environmental heterogeneity might drive the origins of biological diversity, and howbiological diversity can be affected by the accelerating environmental change that ecosystemsare experiencing.
AcknowledgmentsWe would like to thank three anonymous reviewers for helpful feedback that greatly improved the manuscript.
References
1. Bolnick, D.I. et al. (2011) Why intraspecific trait variation matters incommunity ecology. Trends Ecol. Evol. 26, 183–192
2. Turcotte, M.M. and Levine, J.M. (2016) Phenotypic plasticity andspecies coexistence. Trends Ecol. Evol. 31, 803–813
3. Matthews, B. et al. (2011) Toward an integration of evolutionarybiology and ecosystem science. Ecol. Lett. 14, 690–701
4. Yu, Q. et al. (2015) Stoichiometric homeostasis predicts plantspecies dominance, temporal stability, and responses to globalchange. Ecology 96, 2328–2335
5. Carnicer, J. et al. (2015) Global biodiversity, stoichiometry andecosystem function responses to human-induced C–N–P imbal-ances. J. Plant. Physiol. 172, 82–91
6. Sterner, R.W. and Elser, J.J. (2002) Ecological Stoichiometry: TheBiology of Elements from Molecules to the Biosphere, PrincetonUniversity Press
7. Burress, E.D. et al. (2013) Species-specific ontogenetic diet shiftsamong neotropical Crenicichla: using stable isotopes and tissuestoichiometry. J. Fish Biol. 82, 1904–1915
8. Yamamichi, M. et al. (2015) Rapid evolution of a consumer stoi-chiometric trait destabilizes consumer–producer dynamics. Oikos124, 960–969
9. El-Sabaawi, R.W. et al. (2012) Widespread intraspecific organis-mal stoichiometry among populations of the Trinidadian guppy.Funct. Ecol. 26, 666–676
10. Pilati, A. and Vanni, M.J. (2007) Ontogeny, diet shifts, and nutrientstoichiometry in fish. Oikos 116, 1663–1674
11. Leitch, A.R. et al. (2014) Impact of genomic diversity in riverecosystems. Trends Plant Sci. 19, 361–366
12. Gorokhova, E. et al. (2002) Functional and ecological significanceof rDNA intergenic spacer variation in a clonal organism underdivergent selection for production rate. Proc. R. Soc. B 269,2373–2379
13. Weider, L.J. et al. (2005) Genotype � environment interactions,stoichiometric food quality effects, and clonal coexistence inDaphnia pulex. Oecologia 143, 537–547
14. Declerck, S.A. et al. (2015) Rapid adaptation of herbivore con-sumers to nutrient limitation: eco-evolutionary feedbacks topopulation demography and resource control. Ecol. Lett. 15,553–562
15. Snell-Rood, E. et al. (2015) Life-history evolution in the anthro-pocene: effects of increasing nutrients on traits and trade-offs.Evol. Appl. 8, 635–649
16. Boersma, M. and Elser, J.J. (2006) Too much of a good thing: onstoichiometrically balanced diets andmaximal growth. Ecology 87,1325–1330
17. Bullejos, F.J. et al. (2014) Shifts in food quality for herbivorousconsumer growth: multiple golden means in the life history. Ecol-ogy 95, 1272–1284
18. Raubenheimer, D. et al. (2009) Nutrition, ecology and nutritionalecology: toward an integrated framework. Funct. Ecol. 23, 4–16
19. Simpson, S.J. and Raubenheimer, D. (1999) Assuaging nutritionalcomplexity: a geometrical approach. Proc. Nutr. Soc. 58, 779–789
20. Hadjipantelis, P.Z. et al. (2013) Function-valued traits in evolution.J. R. Soc. Interface 10, 20121032
21. Stinchcombe, J.R. et al. (2012) Genetics and evolution of function-valued traits: understanding environmentally responsive pheno-types. Trends Ecol. Evol. 27, 637–647
22. Jeyasingh, P.D. et al. (2014) Testing the ecological consequencesof evolutionary change using elements. Ecol. Evol. 4, 528–538
23. Reich, P.B. and Oleksyn, J. (2004) Global patterns of plant leaf Nand P in relation to temperature and latitude. Proc. Natl. Acad. Sci.U.S.A. 101, 11001–11006
24. El-Sabaawi, R.W. et al. (2012) Environmental and organismalpredictors of intraspecific variation in the stoichiometry of a neo-tropical freshwater fish. PLoS One 7, e32713
25. Neiman, M. et al. (2013) Can resource costs of polyploidy providean advantage to sex? Heredity 110, 152–159
26. Hessen, D.O. et al. (2010) Genome streamlining and the elementalcosts of growth. Trends Ecol. Evol. 25, 75–80
27. Neiman, M. et al. (2009) Can phosphorus limitation contribute tothe maintenance of sex?. A test of a key assumption. J. Evol. Biol.22, 1359–1363
28. Perrin, N. and Sibly, R.M. (1993) Dynamic-models of energyallocation and investment. Annu. Rev. Ecol. Evol. Syst. 24,379–410
29. Frost, P.C. et al. (2010) Transgenerational effects of poor elemen-tal food quality on Daphnia magna. Oecologia 162, 865–872
Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy 9

TREE 2192 No. of Pages 10
30. Calado, R. and Leal, M.C. (2015) Trophic ecology of marineinvertebrates with bi-phasic life cycles – what are we still missing?Adv. Mar. Biol. 71, 1–70
31. Anderson, T.R. et al. (2005)Metabolic stoichiometry and the fate ofexcess carbon and nutrients in consumers. Am. Nat. 165, 1–15
32. Fagan,W.F. et al. (2002) Nitrogen in insects: implications for trophiccomplexity and species diversification. Am. Nat. 160, 784–802
33. Neiman, M. and Krist, A. (2016) Sensitivity to dietary phosphoruslimitation in native vs. invasive lineages of a New Zealand fresh-water snail. Ecol. Appl. 26, 2218–2224
34. Anderson et al. (2004) Stoichiometry: linking elements to biochem-icals. Ecology 85, 1193–1202
35. Elser, J.J. et al. (2003) Growth rate-stoichiometry couplings indiverse biota. Ecol. Lett. 6, 936–943
36. Lind, P.R. and Jeyasingh, P.D. (2015) Genotypic differences inphosphorus use physiology in producers (Chlamydomonas rein-hardtii) and consumers (Daphnia pulex) interact to alter primaryand secondary production. Evol. Ecol. 29, 551–563
37. Frisch, D. et al. (2014) A millennial-scale chronicle of evolutionaryresponses to cultural eutrophication in Daphnia. Ecol. Lett. 17,360–368
38. Chowdhury, P.R. et al. (2014) Functional genomics of intraspecificvariation in carbon and phosphorus kinetics in Daphnia. J. Exp.Zool. A Ecol. Genet. Physiol. 321, 387–398
39. Elser, J.J. et al. (2006) Ontogenetic coupling of growth rate withRNA and P contents in five species of Drosophila. Funct. Ecol. 20,846–856
40. Elser, J.J. et al. (2000) Nutritional constraints in terrestrial andfreshwater food webs. Nature 408, 578–580
41. Huberty, A. and Denno, R.F. (2006) Consequences of nitrogen andphosphorus limitation for the performance of two planthoppoerswith divergent life-history strategies. Oecologia 149, 444–455
42. Tigreros, N. (2013) Linking nutrition and sexual selection across lifestages in a model butterfly system. Funct. Ecol. 27, 145–154
43. Denno, R.F. and Fagan, W.F. (2003) Might nitrogen limitationpromote omnivory among carnivorous arthropods? Ecology 84,2522–2531
44. Simpson, S.J. et al. (2006) Cannibal crickets on a forced march forprotein and salt. Proc. Natl. Acad. Sci. U.S.A. 103, 4152–4156
45. Dalsgaard, J. et al. (2003) Fatty acid trophic markers in the pelagicmarine environment. Adv. Mar. Biol. 46, 225–340
46. Martin-Creuzburg, D. et al. (2008) Nutritional constraints at thecyanobacteria–Daphnia magna interface: The role of sterols. Lim-nol. Oceanogr. 53, 456–468
10 Trends in Ecology & Evolution, Month Year, Vol. xx, No. yy
47. Hairston, N.G., Jr et al. (2001) Natural selection for grazer resis-tance to toxic cyanobacteria: evolution of phenotypic plasticity?Evolution 55, 2203–2214
48. Sperfeld, E. et al. (2016) Bridging ecological stoichiometry andnutritional geometry with homeostasis concepts and integrativemodels of organism nutrition. Funct. Ecol. http://dx.doi.org/10.1111/1365-2435.12707
49. Sperfeld, E. et al. (2016) Woodstoich III: integrating tools of nutri-tional geometry and ecological stoichiometry to advance nutrientbudgeting and the prediction of consumer-driven nutrient recy-cling. Oikos. http://dx.doi.org/10.1111/oik.03529
50. Harrison, S.J. et al. (2014) Towards a synthesis of frameworks innutritional ecology: interacting effects of protein, carbohydrate andphosphorus on field cricket fitness. Proc. R. Soc. B 281,20140539
51. Raubenheimer, D. and Simpson, S.J. (2004) Organismal stoichi-ometry: quantifying non-independence among food components.Ecology 85, 1203–1216
52. Vanni, M.J. (2002) Nutrient cycling by animals in freshwater eco-systems. Annu. Rev. Ecol. Evol. Syst. 33, 341–370
53. El-Sabaawi, R. et al. (2016) Investment in boney defensive traitsalters organismal stoichiometry and excretion in fish. Oecologia181, 1209–1220
54. McIntyre, P.B. et al. (2008) Fish distributions and nutrient recyclingin streams: can fish create biogeochemical hotspots? Ecology 89,2335–2346
55. Rudman, S.M. et al. (2015) Adaptive genetic variation mediatesbottom-up and top-down control in an aquatic ecosystem. Proc.R. Soc. B 282, 20151234
56. Matthews, B. et al. (2016) Experimental evidence of an eco-evolutionary feedback during adaptive divergence. Curr. Biol.26, 1–7
57. Sousa, T. et al. (2010) Dynamic energy budget theory restorescoherence in biology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 365,3413–3428
58. Elser, J.J. et al. (2011) Stoichiogenomics: the evolutionary ecologyof macromolecular elemental composition. Trends Ecol. Evol. 26,38–44
59. Aalto, S.L. et al. (2015) A three-way perspective of stoichiometricchanges on host-parasite interactions. Trends Parasitol. 31,333–340
60. Warbrick-Smith, J. et al. (2006) Evolving resistance to obesity in aninsect. Proc. Natl. Acad. Sci. U.S.A. 103, 14045–14049