the effect of stage of growth on the rate of drying of cut grass at 20°c
TRANSCRIPT

Crass and Forage Science {1979) Volume 34,139-144.
The effect of stage of growth on therate of drying of cut grass at 20°C
LEWIS JONESGrassland Research Institute, Hurley, U.K.
Abstract
The drying of Italian ryegrass (RvP) tillers undercontrolled conditions at three stages of growth isdescribed. The rate of water loss was dependenton tiller morphology as well as on water content.Leaf laminae dried faster than stems and alsoaided the loss of water from stems. Drying timewas shortest for vegetative tillers despite theirhigh water content, and longest for reproductivetillers shortly before ear emergence. The practicalimplications in relation to haymaking are dis-cussed.
Introduction
Despite increases in silage production, haymak-ing is still the commonest method of conservinggrass in the UK. For safe storage as hay it isnecessary to remove more than 90% of the watercontained in the crop when mown. During fielddrying, losses of dry matter (DM) occur throughthe activity both of respiratory enzymes presentin the plant tissue and of micro-organisms presenton the plant surface. Losses increase as the dryingprocess is prolonged as do those arising fromleaching by rain and fragmentation duringmechanical handling; on average in the UK. one-third of the DM present at mowing is lost,equivalent to about 3-5 Ml annually. More rapidremoval of water following mowing therefore in-creases the efficiency of the haymaking process.
Whilst evaporation potential is determined by
Correspondence: Mr L. Jones. Grassland Research Institute,Hurley. Maidenhead. Berks, SL6 5LR. U.K.
OI42-5242/79/0600-OI39S02.00© 1979 Blackwell Scientific Publications
weather conditions, the actual rate of water lossfrom a hay crop depends both on the environ-ment immediately surrounding the cut tillers inthe swath and on limitations imposed by the dry-ing tissue itself. In most haymaking studies theeffects on drying of these two factors have beenconfounded.
A study of limitations imposed by the dryingtissue can be made by exposing grass tillers to aconstant environment. Such studies, which so farhave been restricted to isolated leaves and stems(Lesham et al., 1972; Harris et al. 1974) or tochopped material (Menzies and O'Callaghan,1971; Rees, 1974), have shown that the leaf driesmuch faster than the stem. Further, Shepherd(1964) and Tutberg (1975) working on legumeshave demonstrated the importance of the leaf inaiding the loss of water from the stem.
In grasses, crop water content (expressed in thispaper as kg water per kg DM) may vary withgrowth stage by a factor of three; the proportionof stem (including leaf sheath) ranges from 20% invegetative tillers to 80% in reproductive tillers atanthesis (Figure I). Further, the structure anddegree of exposure of stems also differs betweengrowth stages, and differences in the dryingcharacteristics of tillers at varying stages of de-velopment have been suggested (Morris. 1972).An investigation was therefore undertaken tostudy such drying characteristics when tillers atdifferent growth stages and their constituentparts were dried in a constant environment.
Materials and methods
Italian ryegrass (Lolium muUiflorum), cv. RvP,grown in the field was cut at a height of 5 cm witha reciprocating knife mower at three stages ofgrowth designated R|, R2 and V (Table 1). Oneach occasion, five samples were taken of (a)whole tillers, (b) laminae and (c) stems, the two
139
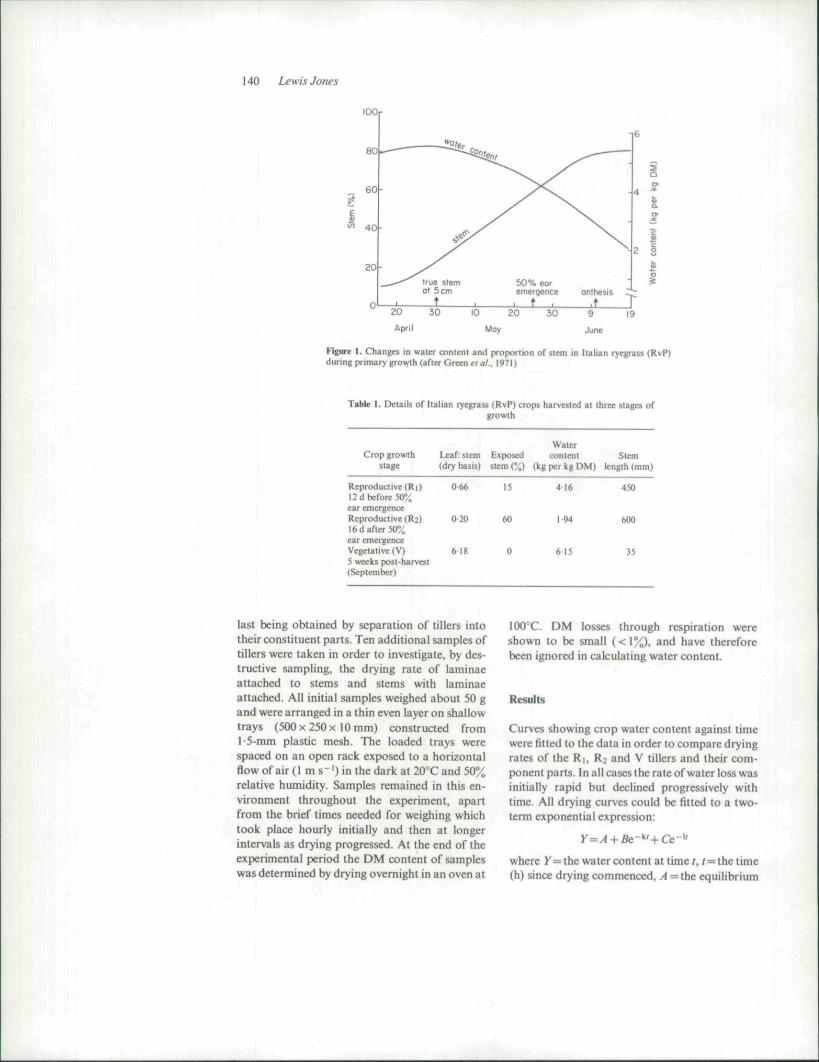
140 Lewis Jones
lOOr
19
Figure 1. Changs in water conieni and proportion of stem in Italianduring primary growth (after Green era/.. 1971)
(RvP)
Table 1. Details of Italian ryegrass (RvP) crops harvested at three stages ofgrowth
WaterCrop growth l.eaf: stem Exposed content Stem
stage (dry basis) stem (%) (kg per kg DM) length (mm)
Reproductive(Ri)12 d before 50%ear emergenceReproductive (Rj)16 d after 50%ear emergenceVegetalive(V)5 weeks f>osl-harvest(September)
0-66
0 20
518
15
60
4 16
I 94
6-15
450
600
35
last being obtained by separation of tillers intotheir constituent parts. Ten additional samples oftillers were taken in order to investigate, by des-tructive sampling, the drying rate of laminaeattached to stems and stems with laminaeattached. All initial samples weighed about 50 gand were arranged in a thin even layer on shallowtrays (500 x 250 x 10 mm) constructed from1 5-mm plastic mesh. The loaded trays werespaced on an open rack exposed to a horizontalflow of air (I m s"') in thedark at 20°C and 50%relative humidity. Samples remained in this en-vironment throughout the experiment, apartfrom the brief times needed for weighing whichtook place hourly initially and then at longerintervals as drying progressed. At the end of theexperimental period the DM content of sampleswas determined by drying overnight in an oven at
lOCC. DM losses through respiration wereshown to be small (<1%), and have thereforebeen ignored in calculating water content.
Results
Curves showing crop water content against timewere fitted to the data in order to compare dryingrates of the R], R2 and V tillers and their com-ponent parts. In all cases the rate of water loss wasinitially rapid but declined progressively withtime. All drying curves could be fitted to a two-term exponential expression:
where K= the water content at time/, r=the time(h) since drying commenced, 4̂ =the equilibrium

Stage of growth and drying rate of grass 141
Figure 2. Changes in water content (o, means of five replicates) at 20°C, 50%relative humidity and 1 m s ^ ' airflow, fitted drying curve and componentexponential curves for Italian ryegrass {RvP) herbage
water content, 5= the water content lost at rate k,C=lhe water content lost at rate 1, k and 1 =theexponential decay rate constants for B and Crespectively (kg water per kg water present per h),e = base of natural logarithms (2.718) and when/ = 0, y=^+B+C=initial water content.
As the equilibrium water content (A) isapproached, the rate of water loss becomes veryslow. Only in laminae was the value of A estab-lished before the experiments were terminated;during all curve-fitting the asymptote was con-strained to 012—the value obtained by Shepherd(1957) for laminae and stems, and confirmed forlaminae in the present experiments.
Figure 2 shows the two-term exponential curvefitted to typical data and also the componentsingle exponential curves. Drying is considered intwo phases: (i) a rapid phase of water loss predo-minantly described by the exponential Bc~^', fol-lowed by (ii) a protracted phase predominantlydescribed by the exponential Ce"". The para-
meters of the model are presented in Table 2 forall the drying curves.
Drying of detached laminae and stems
Laminae dried at a much faster rate than stems(Figure 3). Water loss (per unit of water present)from laminae was three times greater than thatfor stems over the rapid phase of drying and seventimes over the protracted phase. Further, therapid drying phase in laminae accounted for agreater proportion of their total water loss than instems (Table 2). When laminae had dried to astage suitable for storage as hay (0 25 kg water perkg DM), the water content of R I, R:and V stemswas 1-50, 1 00 and 2 00 kg per kg DM respect-ively.
Stems of V tillers lost water faster than those ofR tillers throughout the drying period, whilstwater loss was more rapid in R2 than R] stems. At

142 Lewis Jones
Table 2. Values for parameters of two-term exponentialequations describing water loss from whole tillers, andlaminae and stem fractionsof Italian ryegrass (RvP) cutat three stages of growth and drying as separate (S) orattached (A) units. (Thin layer drying a! 2O''C and 50°/̂relative humidity. Equilibriuni wa[crconteniO't2kgper
kg DM).
Reproductive ( R i )TillerLamina S
AStem S
A
Reproductive (R2)TillerLamina S
AStem S
A
Vegetative (V)TillerLamina S
AStem S
A
4284-043-893-91394
2-051901'891951 90
5-77
B
-1292
ni-84
137
-2968-510406
)-I55-48 4-72620 t-256-72 4-066 72 t35
C
1-041-001 061'951-45
0-640 100 260-790-72
0 500641-833-542-25
k
0 200450-280-090-18
0 230430-320-15020
0-370'400-680-250-33
1
00130-135006600090 011
0019006900920-0150-014
00280 1120 16400200-016
all three growth stages, the rate at which the stemslost water during the rapid phase was about tentimes greater than during the protracted phase(Table 2).
Drying of attached laminae and stems
The stem dried faster when part of the whole tillerthan when separated from its laminae—an indi-cation that laminae aided the loss of water held inthe stem; in the R tillers, this aid retarded thedrying of laminae (Figure 3). The faster drying ofthe stem continued until the water contents of Vand Ri laminae had fallen to about 0-50 and of R2laminae to 0 25 kg per kg DM. The rapid phase ofwater loss for the attached stem was the periodwhen this loss was aided by the laminae andaccounted for almost two-thirds of the total waterloss but only for one-tenth of the drying time. Theprotracted phase was thus the f>eriod of loss of theremaining water directly from the stem surface.
Vegetative Iv)
20 40 60
3. Drying curves at three growth slagcs for laminae and stems drying as separate and attached units and for whole tillers (thinlayer drying al 20°C, 50/o relative humidity and I m s " ' airflow)
attached stemtseparaiestemtattached laminatseparate laminaj
-• whole lillerjt curves based o n 6 x 2 weighings, t curves based on 1 5 x 5 weighings

Stage of growth and drying rale of grass 143
Drying of the whole tiller
The proportion of leaf to stem differed betweentillers al the three growth stages. The dryingcurves reflect these differences; thus the curves forV and R2 tillers were similar to those for attachedlaminae and stems respectively, with that of Ribeing intermediate (Figure 3).
For the whole tiller the rapid drying phase isthe loss of almost all the laminae water plus thatlost from stem when aided by laminae. The pro-portion of water lost in the rapid phase thusdepends on the ratio of lamina to stem (Table 1);it amounted to 90%, 75% and 65% for V, R, andR2 tillers, respectively.
The amount of water remaining and thus lostdirectly from the stem, was twice as much in R| asin V and R2. Further, the rate ofloss of this waterwas slowest in R tillers, particularly R[.
Thus, a water content suitable for storage (0-25kg per kg DM) was attained first by the V tillers,despite their greater initial water content. In con-trast, R2, containing only one-third as muchwater as the V tillers, required twice their dryingtime. Ri tillers, although of intermediate watercontent originally, needed a drying time at leastthree times as long as that of the V tillers (Table3).
Table 3. Time for tillers of Italian rye-grass (RvP) cut at three growth stages todry to a water content suitable for bulkstorage as hay, 0-25 kg p»cr kg DM (h)(Thin layer drying at 20°C. and 50%
relative humidity)
Growth stage Time (h)
Reproductive (Ri) 127-7 ±2 2Reproductive (R2) 79-8 ±2-8Vegetative regrowth (V) 37-2 ± 6-3
Discussion
The finding that a two-term exponential expres-sion gave a good fit to the drying characteristics ofwhole tillers and their component parts was simi-lar to the findings of Menzies and O'Callaghan(1971) and Rees (1974) working on the dryingcurves for chopped grass. It is not easy to accountfor the rapid and protracted drying rates on abiological basis, yet they presumably reflect, atleast in part, the relative ease with which water
can be removed from intercellular spaces andconducting vessels compared with its removalfrom cell sap and cytoplasm.
Water is lost from the plant as water vapour,from evaporating sites close to its surface. A dry-ing curve therefore reflects the resistance encoun-tered by liquid water en route to the evaporatingsite, plus the resistance to the diffusion of watervapour through the plant surface. Further, theease with which vapour can move away from theplant surface (so maintaining a gradient of watervapour concentration between the surface andthe surrounding atmosphere) will also influencethe rate of drying. Plant structure therefore playsan important part in determining the rate of waterloss, probably accounting for much of the differ-ential drying of laminae and stems.
Laminae have a much larger surface area perunit of water contained than do stems. In addi-tion, the bulk of the liquid water is closer to thesurface in laminae. The surfaces of laminae alsooffer a lower diffusive resistance to water vapour,as a result of tbe numerous stomatal pores pre-sent. During curve fitting it was found for leafymaterial that a three-term exponential expressionimproved the fit over the initial period of drying, asimilar finding to that of Menzies and O'Callag-han (1971). This extra term probably describesthe very rapid loss of water through open sto-mata.
Stomata close within an hour of the com-mencement of drying (Clark el al., 1977); subse-quently, water vapour must diffuse through thewaxy cuticle which covers all uncut laminae andstem surfaces. The continuing faster drying oflaminae after stomatal closure compared withstems suggests a lower cuticular resistance forlaminae. Such a finding is similar lo that ofLeshem el al. (1972) working on cocksfoot grownindoors; they showed also that removal of theepicuticular wax doubled the rate of water lossfrom laminae but had no effect on the drying ofstem intemodes. In field-grown material, the dif-ference in cuticular resistance of laminae andstems will thus be accentuated by the removal ofepicuticular wax through the mutual abrasion ofplant parts blown about in the wind, as shown forclover by Hall and Jones (1961).
The work of Menzies and O'Callaghan (1971)and Rees (1974) on chopped grass suggests thatvariation in stem length would only have a smalleffect on water loss; differences between stems atthe three growth stages thus also mainly reflect

144 Lewis Jones
structural difference. Faster drying of V com-pared with R stems probably results from theabsence of true stem in the former. The work ofHarris et al. (1974) suggests that the faster dryingof R; compared with R| stems is due to a greaterdegree of true stem exposure in R2.
The higher the water content of the tiller thegreater was the efTecl of the attached lamina inaiding stem drying rate, as Shepherd (1964) foundwith white clover, so that tillers with a high initialwater content benefited most.
The drying requirement of a tiller has beenshown to be primarily dependent on the propor-tion of lamina to stem but it is also affected by theinitial water content and the degree of stem expo-sure. Thus, the liller just before ear emergence,with its high water content and low proportionboth of lamina and of exposed stem, is the mostdifficult to dry.
Practical implications
The data suggest that crop conditioning treat-ments to accelerate drying should mainly aim toreduce the differential drying of laminae andstenis.
The long protracted drying phase shown by alltillers and particularly by reproductive tillers atthe nutritionally desirable stage of growth (R,),highlights the advantage to be gained fromdeveloping techniques which allow for the earlytermination of field drying. Assuming a similarpattern to occur under swath conditions, thecurves suggest that maximum advantage, both intenns of reduced weather risk and minimal shat-tering of dry laminae, would be gained from ter-mination of field drying when tiller water contenthas fallen to about 0-7 kg per kg DM (40% wetbasis).
The relative ease of drying of vegetativeregrowth suggests that greater attention should
be paid to the possibilities of producing hay fromsuch crops.
Acknowledgments
The author thanks the Agricultural ResearchCouncil and the Grassland Research Institute forproviding facilities to conduct the work, Ms J. C.Prickett for valuable technical assistance and DrJ. M. Cobby for curve fitting and statistical analy-sis. Thanks are also due to Professor A. Lazenbyand Dr E. L. Leafe for encouragement and forcriticism of the manuscript.
References
CLARK B.J..PR10ULJ.-L. and CouMRcH. (1977) The physiolo-gical response lo culling in Italian ryegrass. Journal of iheBriiiifi Grassland Society, 32, 1-15,
GREEN JO. . C ( « R A L L A.J. and TERRY R . A . (1971) Grass speciesand varieties. Technical Report, Grassland Research Institute.Hurley, No. 8.
HALL D M , and JONES R . L . ( t%l ) Physiological significance ofsurface wan on leaves. Nature (London), 191.95-96.
HARRISC.E-, THArNE R. and SARISALOH.I .M. (1974) Effcclive-ness of some mechanical. Ihennal and chemical laboraloryireaimenis on the drying rates of leaves and stem internodesof grass. Journal of Agricultural Science, Cambridge, 83,353-358.
LESHEM V,. THAINE R. . H A R R B C . E . and CANAWAV R, J . (1972)Water loss from cut grass with spKial reference to hay-making. Annals of Applied Biology. 72. 89-104,
MENZtK D.J. and O'CALLAGHAN J . R , (1971) The effect oftemperature on the drying rale of grass. Journal of Agricul-tural Engineering Research, 16, 213-222.
MORRIS R M , (1972) The rate of water loss from grass samplesduring hay-iype conservation. Journal of the British Grass-land Society, 71,99-lOi.
REES D . V . H . (1974) Investigations on the drying of herhage attemperatures up to 50°C. Journal of the British GrasslandSociety, 79.47-55.
SHEPHERD W, (1957) Moisture relations of hay species. Austra-lian Journal of Agricultural Research, 9, 436-445
SHEPHERD W . (1964) Paths and mechanisms of moisture move-ments in detached leaves of white clover. 1. Losses of petiolemoisture direct from petioles and via laminae. Annals ofBotany N.S., 28.207-220.
TULBERG J . N . (1975) Studies of lucerne hay (tying. PhD thesis.University of Queensland. Brisbane.
(Received 14 July 1978)
