the evolution of transport systems in plants introduction xylem phloem summation; integration...
TRANSCRIPT

The evolution of transport systems in plants
Introduction Xylem Phloem Summation; integration
http://www.xs4all.nl/~steurh/eng/rhynia.html

evolution or revolution ?
In the beginning………….
hydrocarbonsair
water
light
stress
ozoneoxygencarbon
nitrogenhydrogen
land
migration
? ? ? ?

The symplast and apoplast
Symplast - Defined as that region of the living plant, that is bounded (enclosed by) a plasmamembrane. The cells forming this group are called a DOMAIN, and DOMAINS are connected via PLASMODESMATA.
Apoplast. By definition, all regions of living plant cells NOT bounded by a plasmamembrane. This must include the CELL WALL as well as intercellular spaces. These two constitute FREE SPACE in the plant. Apoplast thus involved in the (free) movement of substances and the principal conduit in this case, must be the xylem or xylem-associated cells that LACK a plasmamembrane.
next

xylem and phloem - the parallels
Phloem and xylem have distinct physiological functions, and their distinctly-different functions give them a unique parallelism.
The xylem conducts water, usually acropetally, from root to shoot and the leaf, (in a sense, from source to sink) whilst the phloem in the crudest sense, transports carbohydrate and other substances from a site of manufacture (source) to a site of utilization (sink).
In both cases, tubes are involved – sieve tubes in the phloem, and xylem vessels and narrower trachieds in the xylem. Both systems are associated with parenchymatous elements, and both are relatively delicate structures.
It is now more than 160 years since, for example, the publication of Hartig’s descriptions of the bark of trees. There is much new information concerning structure and function, but also, we have become more aware of the fossil record, allowing us to trace the evolution of these remarkable conduits through some 500 million years of evolution.
//3

phloem - early work
1. In 1837, Theodor Hartig published his work on the organization of the trunks of forest trees, and commented on the then “completely uninvestigated elementary organs” of what was called the ‘sap skin’ (Safthaut) of trees.
2. Significantly, Hartig described three types of cell – Siebfasern, Siebröhren and keulenförmige Saftröhen – in English, the sieve elements and laticifers common in Euphorbia for example.3. The Siebröhren and Siebfasern correspond entirely to what we describe today as sieve tubes and in the latter case, to sieve cells, associated cambial cells, phloem parenchyma as well as sclerenchymatous elements.
4. In 1858, Nägeli coined the term ‘phloem’ top describe the food conducting systems in higher plants.
// 1-4

Moving on.Developing the xylem transport system
Initially, transport of water in plants that had just emerged onto land, would have been a physiological necessity –
Why?

Ancient origins
Indeterminate hydroid or stromatoporoids in Starostinella nordica
Upper Permian ca 280 MYSince then many examples in which hydroids appear.

Stelar arrangements and tissue organisation
All comparisons serve to underscore the fundamental relationship in early vascular plants between the evolution of increased complexity in stelar architectures and the evolution of complex lateral branches and leaves.
water transporting function is the crucial factor in the evolution of the various steles.
Vascular plants seem to be the ideal study group for integrating developmental process directly into analysis of homology and estimation of phylogenetic relationship.
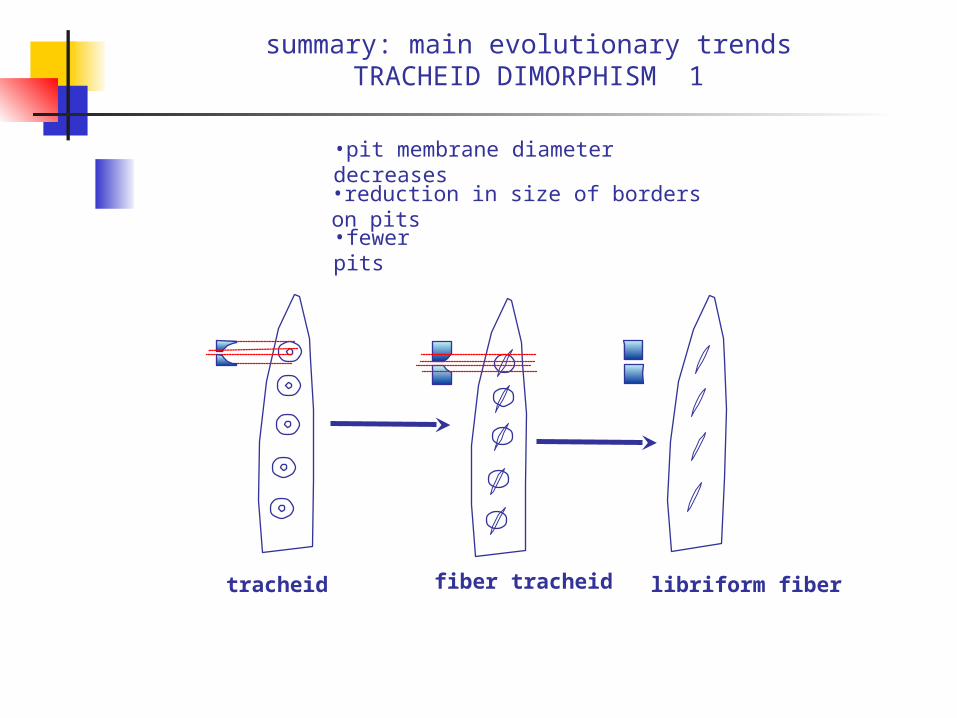
summary: main evolutionary trendsTRACHEID DIMORPHISM 1
•pit membrane diameter decreases•reduction in size of borders on pits
•fewer pits
tracheid fiber tracheid libriform fiber

summary: main evolutionary trends TRACHEID DIMORPHISM 2
tracheid dimorphism:vasicentric tracheids +septate fiber tracheids
fiber tracheid dimorphism:fiber tracheids + vasicentric tracheids
libriform fiber
fiber dimorphism: parenchyma-like fibers (or parenchyma) + libriform fibers
fiber tracheidtracheid
SPECIAL EVOLUTIONARY TRENDS

Narrow is best ?
Tracheids in their various forms are thus the principal water conducting cells in gymnosperms, as well as in vesselless dicotyledons.
They present many advantages in terms of transport.
Think for example, of the boreal forests – what is the species composition?
Mostly gymnosperm, some vesselless angiosperms
Why?
click

Vessel bearing dicotyledon
Transition from vessel-less to vessel-bearing wood. Qualitatively, a marked drop in the length of the tracheid to primitive vessel occurs.
Vessel end retains remnant primary wall strands
Vessel elements and tracheids much shorter than those in vessel-less wood
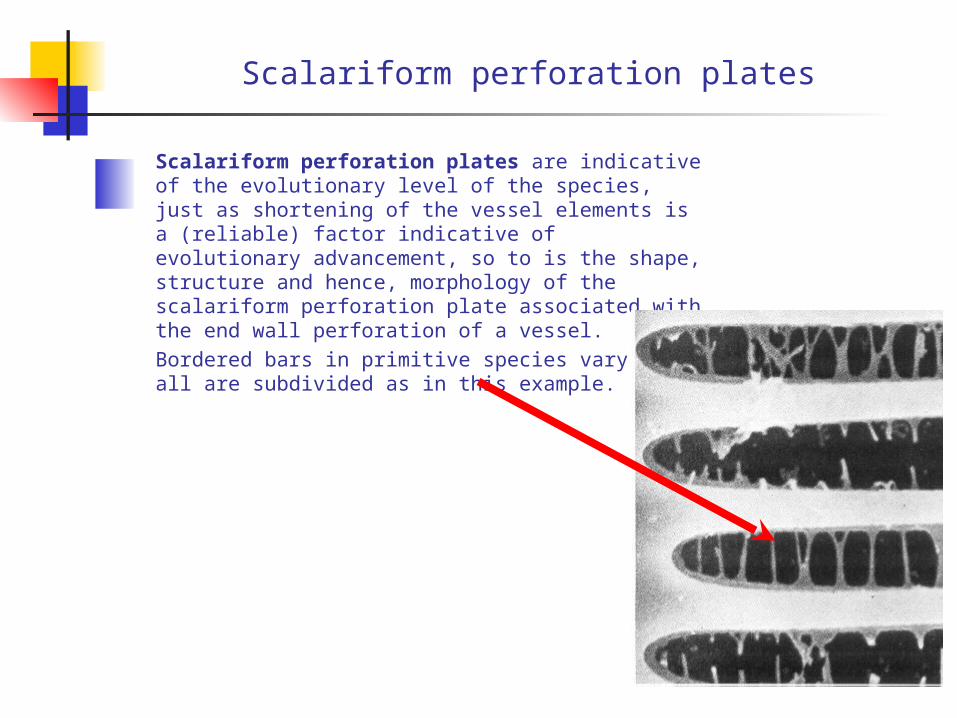
Scalariform perforation plates
Scalariform perforation plates are indicative of the evolutionary level of the species, just as shortening of the vessel elements is a (reliable) factor indicative of evolutionary advancement, so to is the shape, structure and hence, morphology of the scalariform perforation plate associated with the end wall perforation of a vessel.Bordered bars in primitive species vary but all are subdivided as in this example.
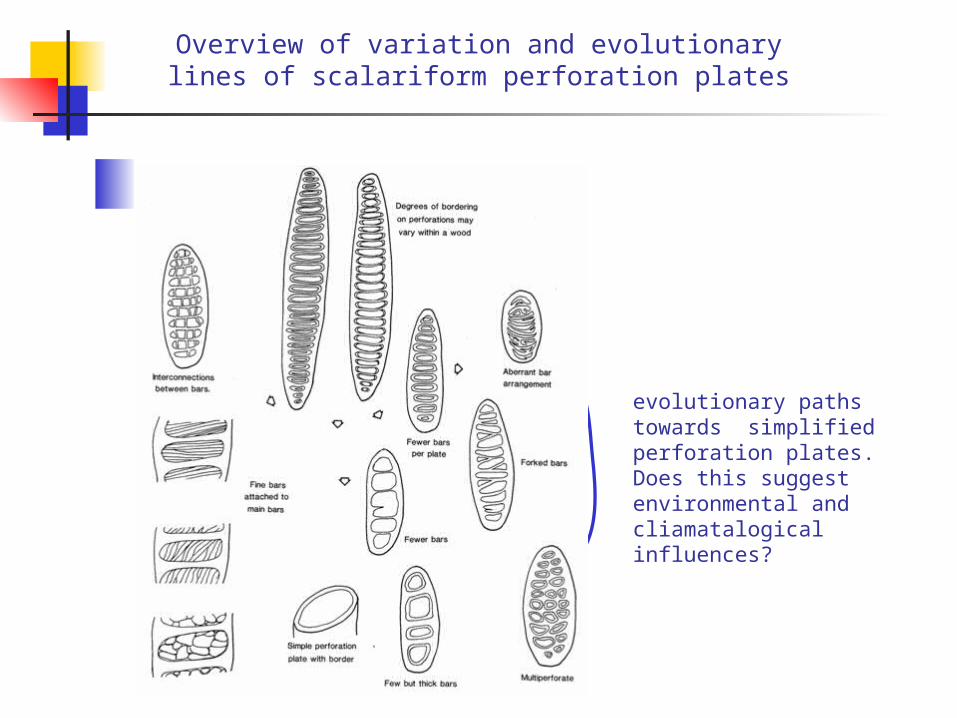
Overview of variation and evolutionary lines of scalariform perforation plates
evolutionary paths towards simplified perforation plates. Does this suggest environmental and cliamatalogical influences?
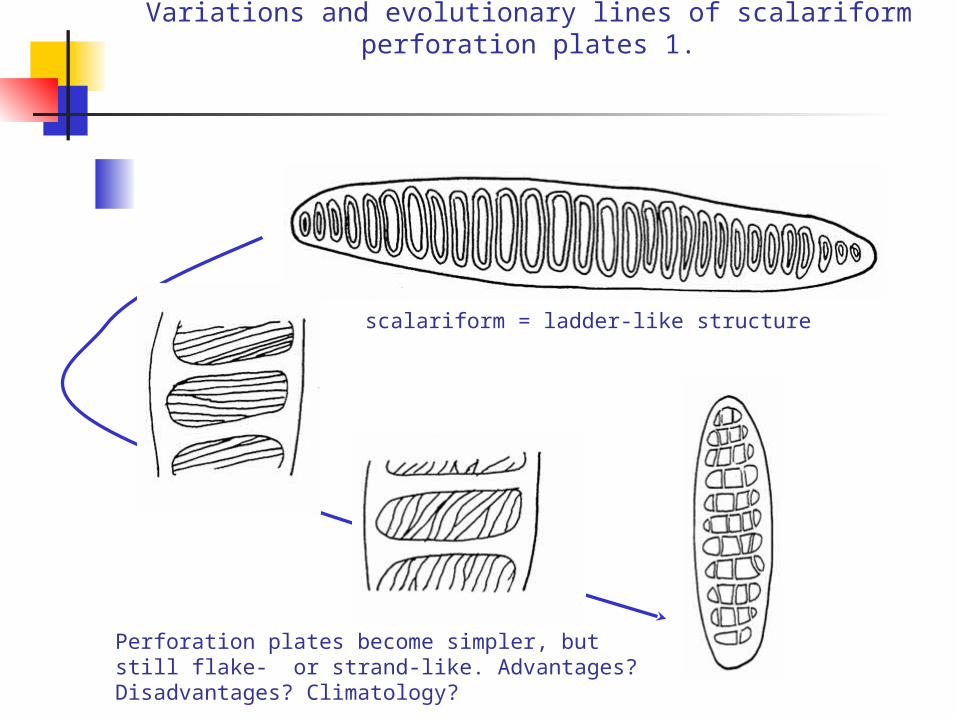
Variations and evolutionary lines of scalariform perforation plates 1.
Perforation plates become simpler, but still flake- or strand-like. Advantages? Disadvantages? Climatology?
scalariform = ladder-like structure
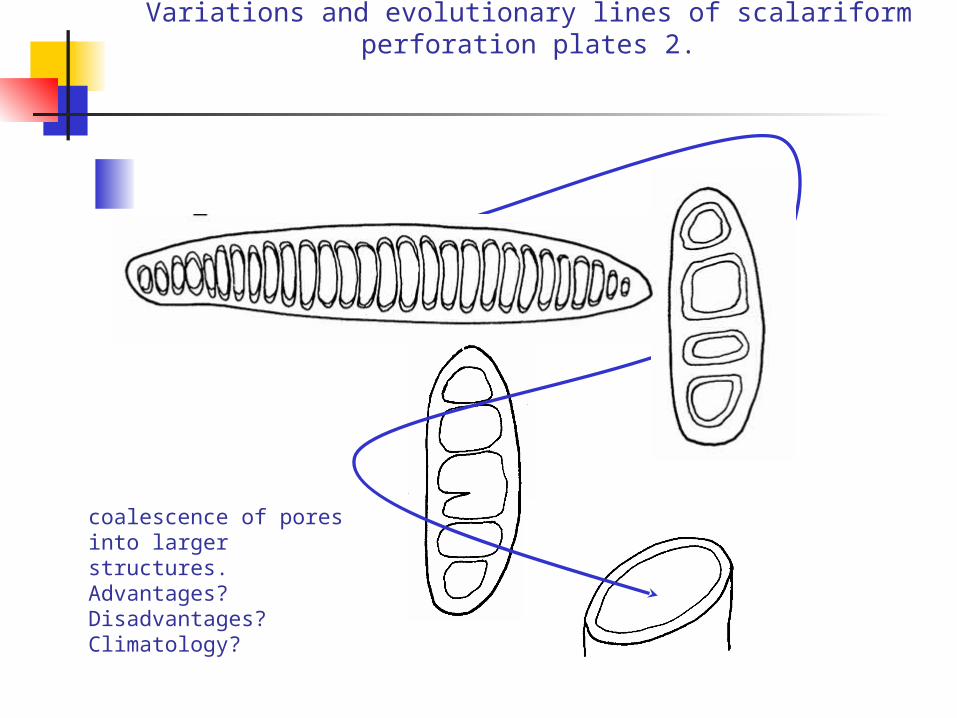
Variations and evolutionary lines of scalariform perforation plates 2.
coalescence of pores into larger structures.Advantages? Disadvantages? Climatology?

Lateral wall pitting, in tracheids and vessels
scalariform transitionalopposite alternate
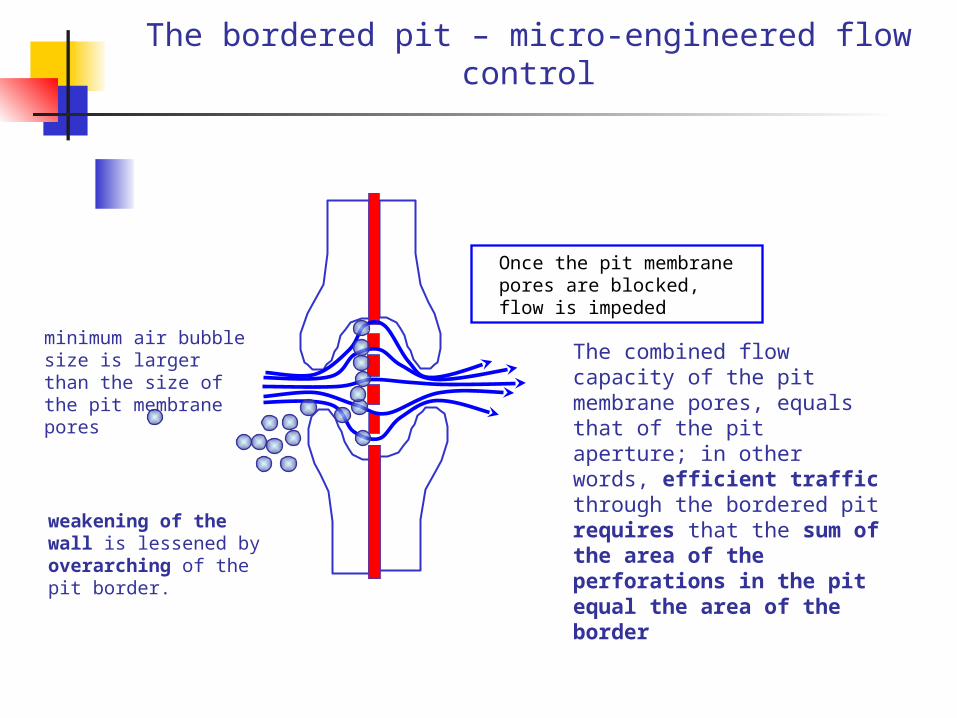
The bordered pit – micro-engineered flow control
The combined flow capacity of the pit membrane pores, equals that of the pit aperture; in other words, efficient traffic through the bordered pit requires that the sum of the area of the perforations in the pit equal the area of the border
weakening of the wall is lessened by overarching of the pit border.
minimum air bubble size is larger than the size of the pit membrane pores
Once the pit membrane pores are blocked, flow is impeded
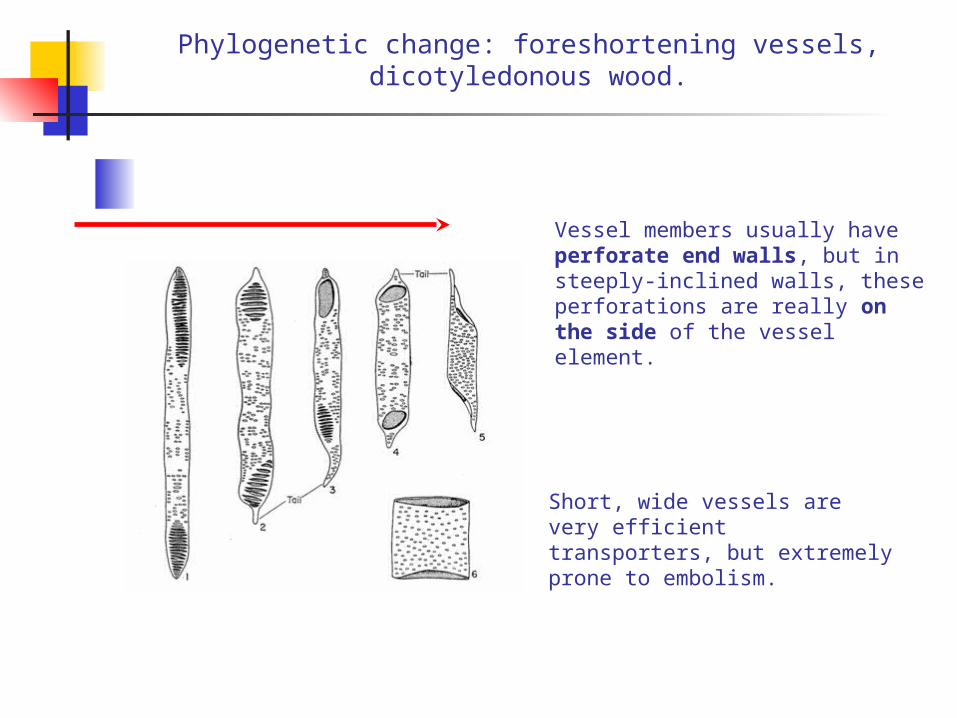
Phylogenetic change: foreshortening vessels, dicotyledonous wood.
Vessel members usually have perforate end walls, but in steeply-inclined walls, these perforations are really on the side of the vessel element.
Short, wide vessels are very efficient transporters, but extremely prone to embolism.
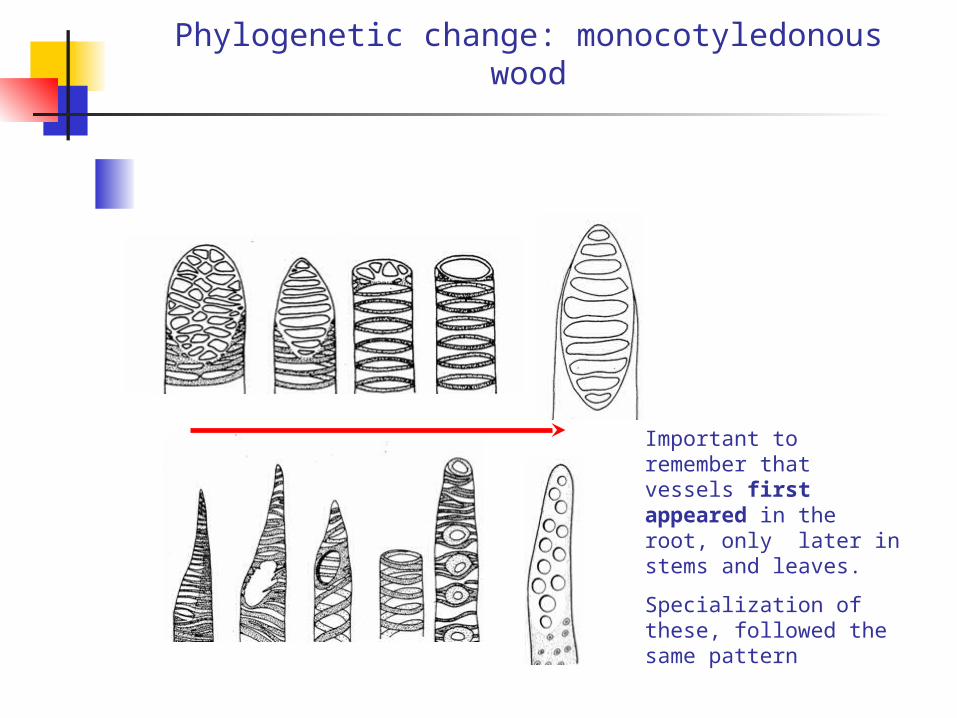
Phylogenetic change: monocotyledonous wood
Important to remember that vessels first appeared in the root, only later in stems and leaves.
Specialization of these, followed the same pattern
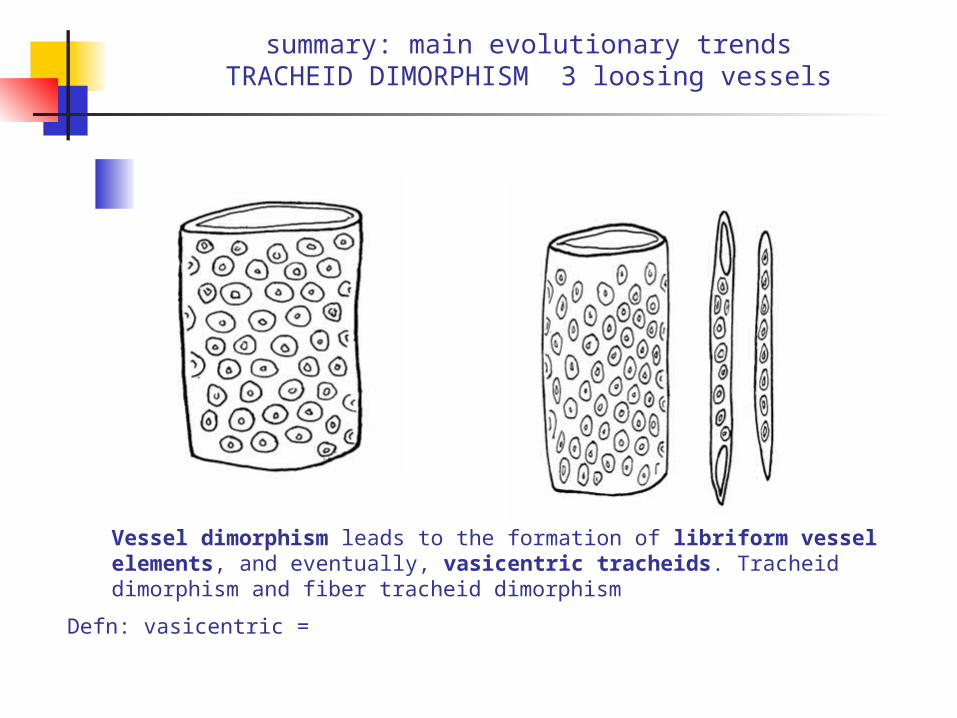
summary: main evolutionary trendsTRACHEID DIMORPHISM 3 loosing vessels
Vessel dimorphism leads to the formation of libriform vessel elements, and eventually, vasicentric tracheids. Tracheid dimorphism and fiber tracheid dimorphism
Defn: vasicentric =
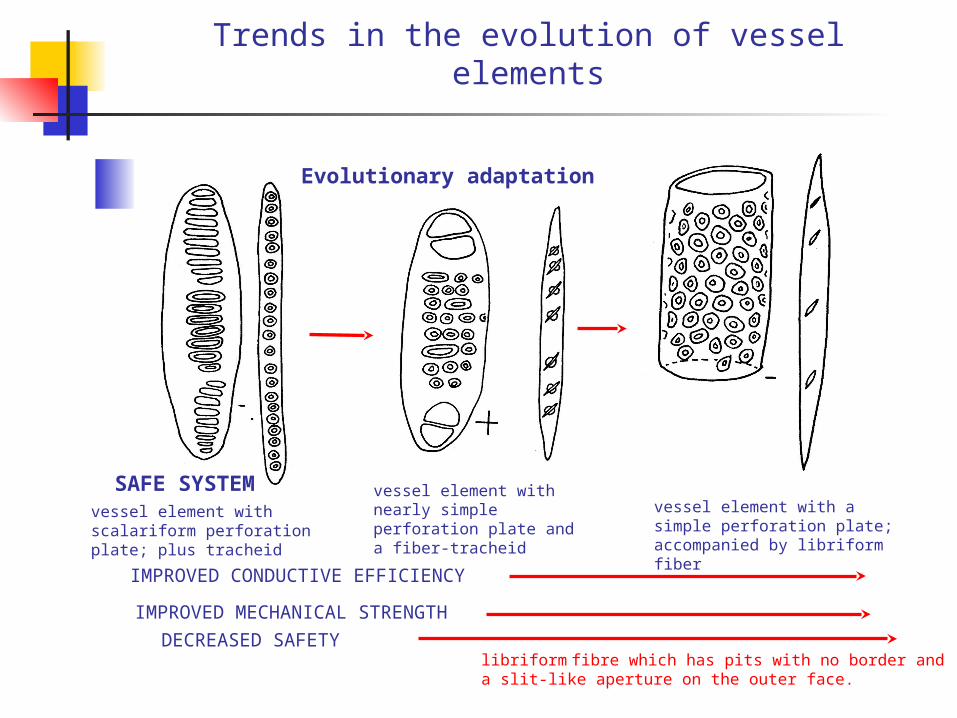
Trends in the evolution of vessel elements
vessel element with scalariform perforation plate; plus tracheid
vessel element with nearly simple perforation plate and a fiber-tracheid
IMPROVED CONDUCTIVE EFFICIENCY
IMPROVED MECHANICAL STRENGTH
SAFE SYSTEM
Evolutionary adaptation
vessel element with a simple perforation plate; accompanied by libriform fiber
DECREASED SAFETYlibriform fibre which has pits with no border and a slit-like aperture on the outer face.
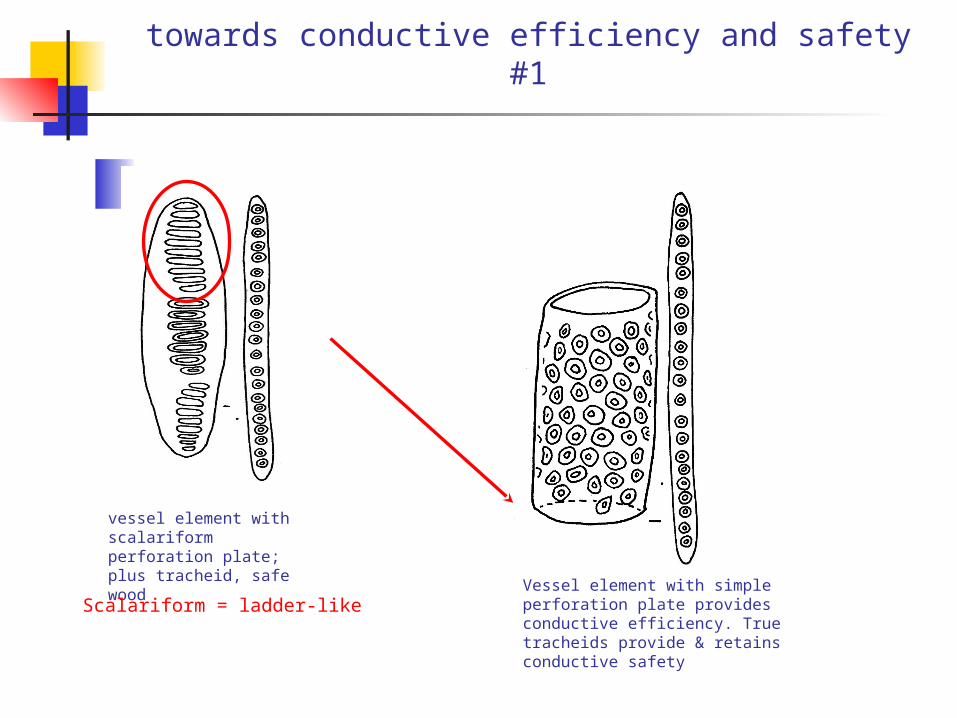
towards conductive efficiency and safety #1
vessel element with scalariform perforation plate; plus tracheid, safe wood
Vessel element with simple perforation plate provides conductive efficiency. True tracheids provide & retains conductive safety
Scalariform = ladder-like

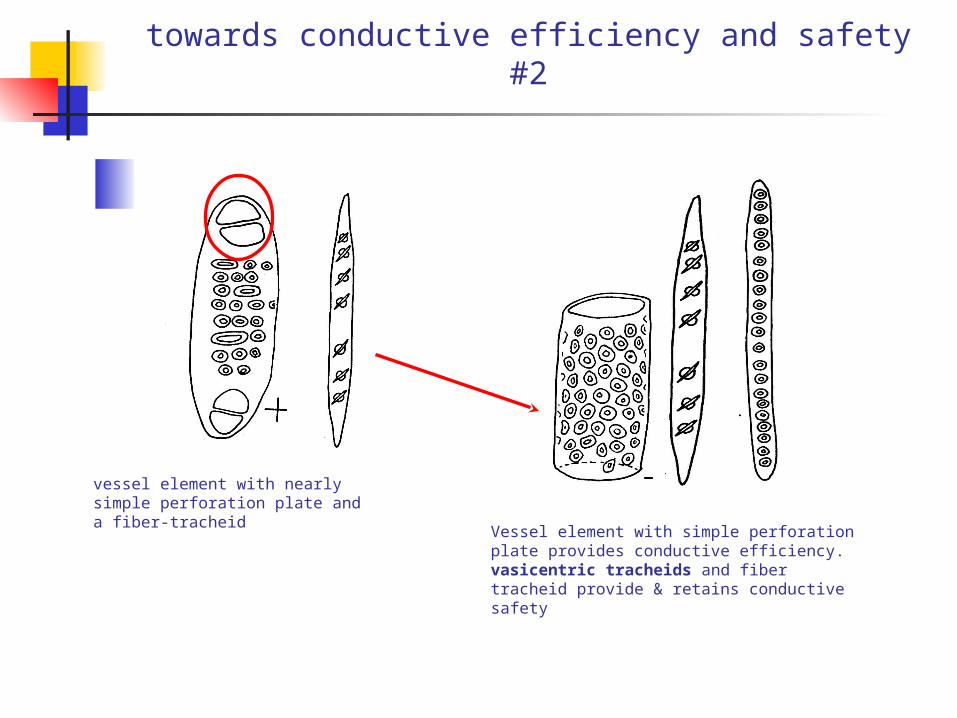
towards conductive efficiency and safety #2
vessel element with nearly simple perforation plate and a fiber-tracheid
Vessel element with simple perforation plate provides conductive efficiency. vasicentric tracheids and fiber tracheid provide & retains conductive safety
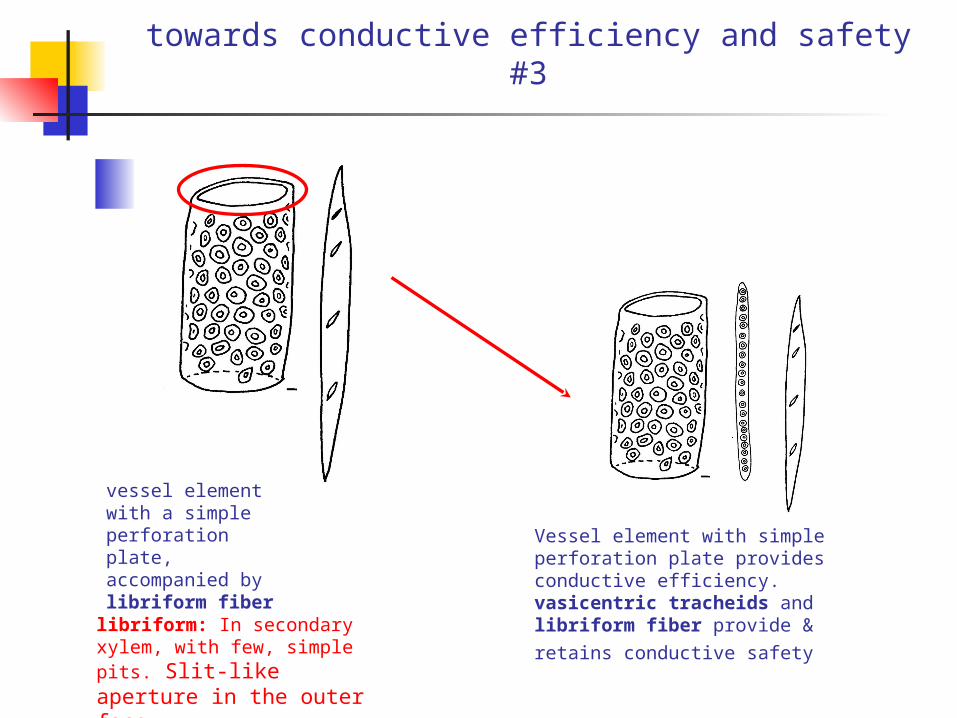
towards conductive efficiency and safety #3
vessel element with a simple perforation plate, accompanied by libriform fiber
Vessel element with simple perforation plate provides conductive efficiency. vasicentric tracheids and libriform fiber provide & retains conductive
safety libriform: In secondary xylem, with few, simple pits. Slit-like aperture in the outer face

pumpkin sunflower

Tracheary element evolution –(& the vesselless dicotyledon)
Narrower tracheids have circular pits on end and lateral wallsWider tracheids may bear scalariform pits on overlap areas (end walls), as well as on their lateral walls.
Tracheid end wall. These are really micropores in the pit membrane (scalariform perforations develop from this). Are there advantages? disadvantages? Much argument still.
Example? Winteraceae resist freeze-thaw conditions, 0 – 6% loss of hydraulic conductance only, vs >20% for vessel-containing dicots. Clearly an advantage. Reasons for the possible loss of vessels and the ecological events underlying this process remains mysterious!

pine, TS pine, LS
50 50 µµmm 50 µm

Whose got xylem?
Division Psilophyta: Psilopsids, characterised by presence of microphylls. No differentiation of shoot and root.
Division Lycophyta: Have microphylls. Division Sphenophyta: The horsetails Single genus (Equisetum) –
jointed stems, conspicuous nodes. Leaves scale-like. Division Pterophyta: Ferns. All possess a megaphyll. Division Coniferophyta: Conifers. Gymnosperms with active cambial
growth (secondary tissue) simple leaves. Division Cycadophyta: Cycads. Gymnosperms with sluggish cambial
activity. Division Ginkophyta: Considerable cambial growth, fan-shaped
leaves. Open dichotomous venation. Division Gnetophyta: Gymnosperms with many angiosperm-like
features. Only Gymnosperms in which vessels occur. Division Anthophyta: Flowering plants. Megaphylls, secondary
growth. Contain dicotyledons (cambium) and monocotyledons (no cambium).
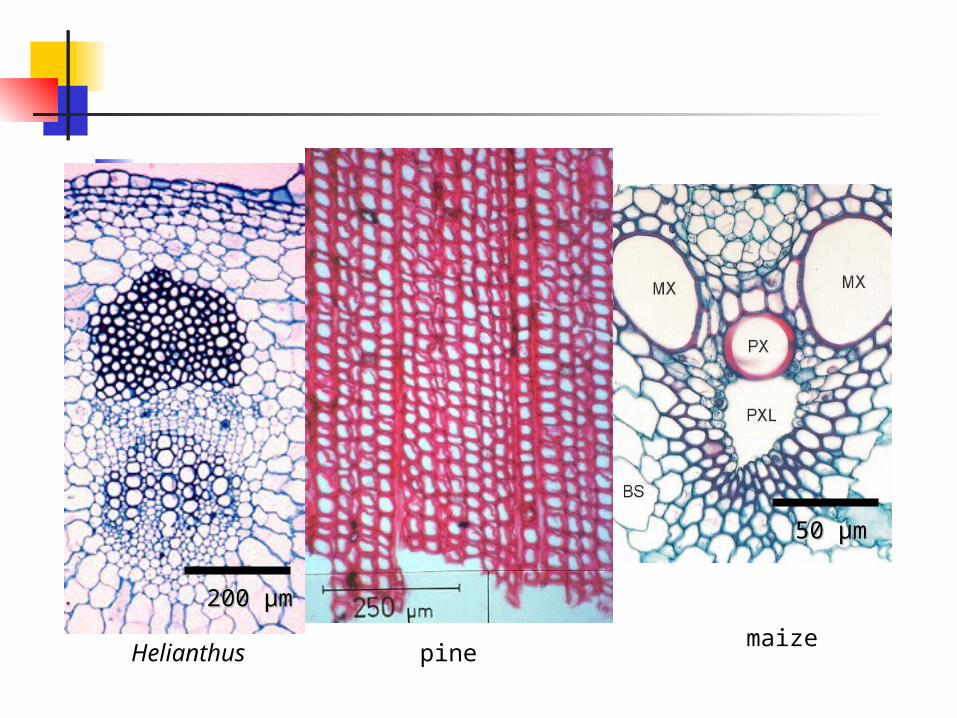
Helianthus pinemaize
200 200 µµmm
50 50 µµmm

Summary
First land plants had hydroids – simple in structure – may have been lignified
Later evolution of more complex wall structureEvolution of tracheoidsTracheids become the principal water conducting elementsVessels – essentially complex, with compound perforations
(scalariform common)Mesomorphs/subtropical to tropical wood – evolution of simple
perforation plates, some crassulae Crassulae: Transversely oriented thickenings in tracheid walls of
gymnosperms accompanying the pit pairs. Also called Bars of Sanio).
Shortening and widening of vessels in warmer climates.

Xylem transportXylem transport

The phloem
Sieve plate: That part of the wall of sieve tube elements (members) bearing one or more highly differentiated sieve areas.
Definitions
Sieve element: The cell of the phloem that is specialized and involved in the long-distance transport of food substances; sieve tubes are further classified into sieve cells and sieve tube elements (or members).
Sieve cell: A long slender element, with relatively unspecialized sieve areas, with tapering end walls that lack sieve plates; found in the phloem of gymnosperms.
Sieve tube element (member): One of the component cells of the sieve tube; found primarily in the flowering plants and typically associated with companion cells.
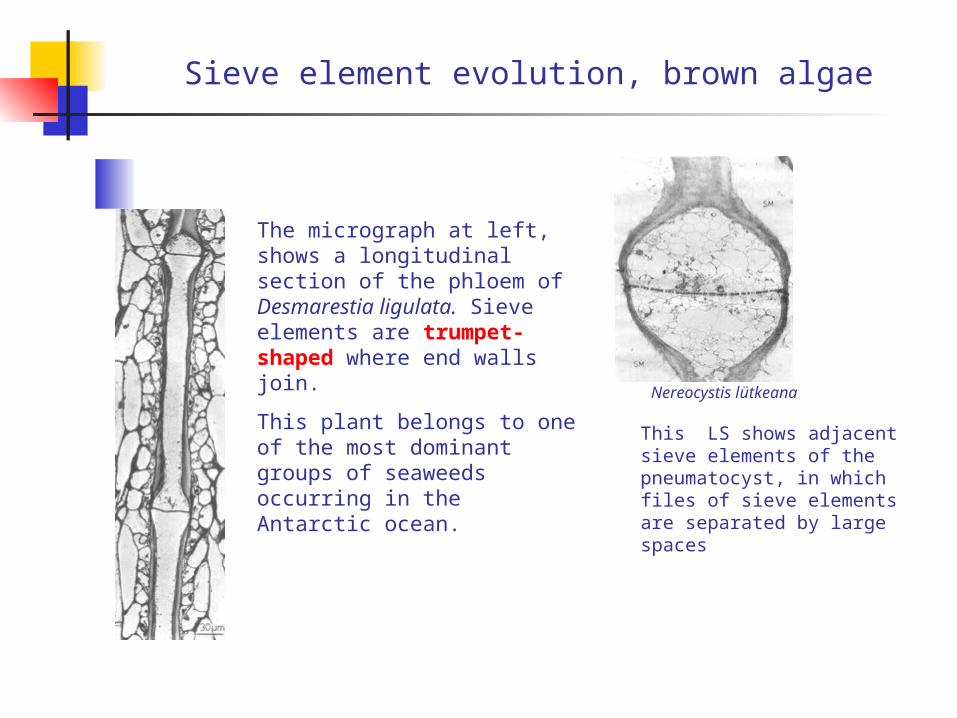
Sieve element evolution, brown algae
The micrograph at left, shows a longitudinal section of the phloem of Desmarestia ligulata. Sieve elements are trumpet-shaped where end walls join.
This plant belongs to one of the most dominant groups of seaweeds occurring in the Antarctic ocean.
Nereocystis lütkeana
This LS shows adjacent sieve elements of the pneumatocyst, in which files of sieve elements are separated by large spaces

organized transport in seaweeds sieve cells or sieve tubes
1. The perforated end wall is one of the most characteristic features of the phloem in the marine algae. The size of the perforations are species dependant.Pore sizes range from as small as plasmodesma (100 nm or so) or as wide as the sieve pores in vascular plants.
1/3
2. Phloem systems tend to be well-developed for long-distance transport of organic and inorganic nutrient. 2/3
3. In Laminariales, cross-connections exist, interconnecting longitudinal strands either via cross-connecting sieve elements, or via lateral sieve areas. In Macrocystis these lateral sieve area pores are smaller in diameter than those occurring in the cross walls, thus one could define the sieve elements as true sieve tubes and the perforated end walls are thus true sieve plates.
3/3

the phloem lower orders, Gnetophytes
Within the cycads, the phloem consists of sieve elements and narrow parenchyma cells.
The Cycadophytinia represent a group of plants, extant since the Jurassic (208my), with strong links to the angiospermae.
Longitudinal sections through part of the phloem in Cycas revoluta, showing differentiated (SE) and differentiating sieve elements (DSE)
Note: The term ‘sieve element’ is used here, to describe more primitive sieve cells and more advanced sieve tube members.

primitive dicotyledonous plants
The photomicrographs to the left, show details of the phloem tissue in the phloem of Magnolia kobus in which the cell wall of the sieve elements, can be see to be thickened, and of wavy outline. This wall is termed the nacreous wall, which is less cellulosic and pectin-poor compared with the outer wall layer. These inner walls have been shown to be polylamellate in many species.

higher plant sieve plate pores
As seen in this example from Beta vulgaris, the sieve plate pores in mature sieve elements (bottom micrograph) are large, and if prepared properly for electron microscopy, are devoid of callose. They are approximately 0.5 µm in diameter.
0.5 µm
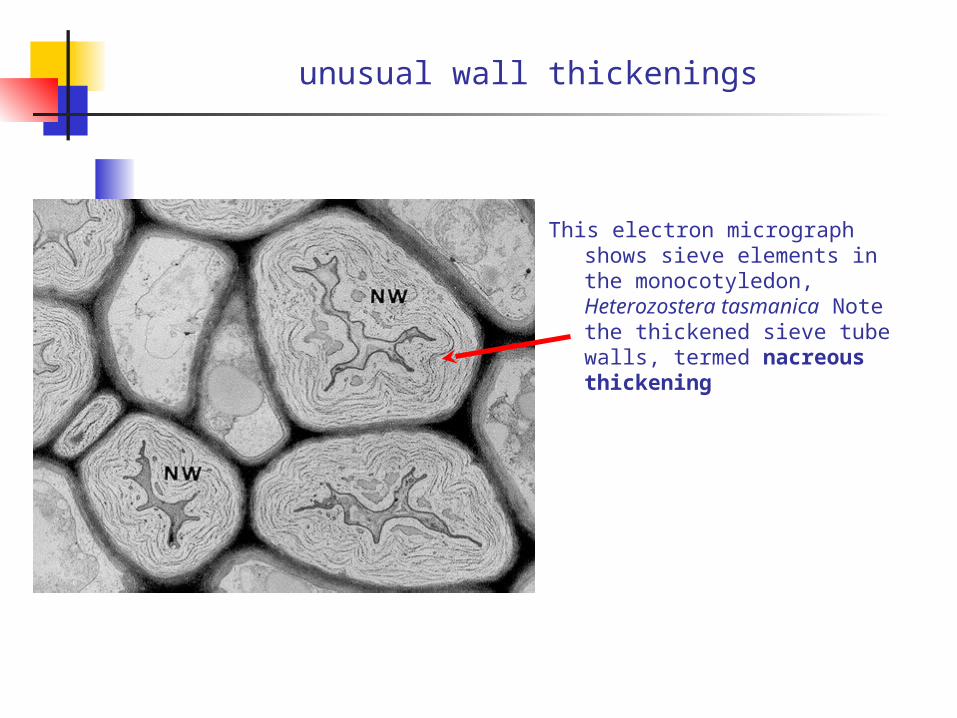
unusual wall thickenings
This electron micrograph shows sieve elements in the monocotyledon, Heterozostera tasmanica Note the thickened sieve tube walls, termed nacreous thickening

fossil records, carboniferous (362my) gymnosperms
Left: Cordaixylon, ts outer phloem. Parenchyma cells appear smaller than the sieve cells (S).
Right: Callistophyton, showing alternation of sieve cells with parenchyma cells. R = Ray tissue
EL Taylor Phloem evolution: Ch 14, Behnke and Sjoland Sieve Elements

Evolutionary process, fossil records -- Lycopods, Sphenophyta and ferns
Taxon Details of Phloem
Lycopods Sieve elements, parenchyma; ca 15.7 µm xs; end wall horiz/obl;
SphenophytaSpenophyllum
Sieve elements; 12.5-21 µm xs; end wall horizontalPrimary and secondary sieve elements + parenchyma + interfasicular parenchyma.
Ferns
Ankyopteris; Botryopteris; Elapteris; Psaronius; Sauropteris
Adaxial & abaxial sieve elements; small & large sieve elements, (small = 10 27 µm; large = 48-120 µm). Sieve plates horiz – oblique; have sieve areas and sieve pores

Evolutionary process, fossil records - Progymnosperms
Taxon Details of Phloem
Progymnosperms‘aneurophyalean’
Sieve cells, 15-25 µm xs; very oblique end walls; sieve pores v. small, 0.5-0.6 µm
Callixylon (Devonian) Not understood, poor preservation
Calamopitys Sieve cells (primary and secondary);50-60 µm xs; oblique end walls
Callistophyton Sieve cells; secretory canals; 20-25 µm xs; very oblique end walls; v. long sieve cells (6mm).
CordaitesCordaixylon
Sieve cells 25 µm xs; cf 1.5 – 1.7 mm long; very tapered ends.
ConiferalesCupressinocortex
Sieve cells; 25-30 µm xs
Taxodioxylon Sieve cells; 20 µm xs 50 µm long

Fossil records - cryptogram & gymnosperm phloem similarities
Tissue origin:conducting cell:
tissue composition:
Vascular Cryptograms
sieve elements (primary)
sieve elements and parenchyma cells
Gymnosperms
sieve cells (primary and secondary)
sieve cells, parenchyma elements, fibers, sclereids, secretory cells
Sieve elements:
shape:
diameter:
length:
end wall:
sieve areas:
sieve pores:
callose:
rectangular to elongate
10-40 µm (up to 120 µm a)
<600 µm (>2.75 mm)
horizontal to very oblique
elliptical-rectangular & small
approx 1 µm dia.
present in some
elongate
20 -100 µm
1 – 9 mm
horizontal to very oblique
elliptical, larger app. 10x30µm
approx 1 µm dia.

Phloem phylogeny #1
Amongst fossil and extant species, the basic structure of the sieve element is fairly uniform in the cryptograms,.
Primary phloem is very poorly known from fossil Gymnosperms and the data presented in the previous two tables is based purely on the secondary phloem.
Whilst phloem has been noted in all groups of fossil Gymnosperms, it appears to have been researched in very few groups, including the cordaites, conifers, and Palaeozoic seed ferns.
In all instances phloem alternates with bands of fibers which may have interspersed axial parenchyma.
All in all the gymnosperms, the sieve cell is remarkably constant in its structure, with the first Middle Devonian progymnosperm record, being very similar to those in Carboniferous seed plants, and are comparable to extant sieve cells being 2.5 -> 9mm in length, with gradually tapering end walls.

Phloem phylogeny #2
Regardless of the group within the Gymnosperms, there is an intimate and regular relationship between axial parenchyma and conducting elements.
Fibers tend to be present and are variable in distribution, from the innermost (presumed functional) to the outermost regions closely spatially associated with the bark.
Sieve areas are discrete, uniform in size and shape, with distinct pores that can be counted (unlike cryptograms where pores require TEM or SEM for elucidation).
Callose has been observed, both as a collar surrounding the pores, or as definitive callose.

Phloem Phylogeny #3 – where did divergence occur?
Sieve elements in vascular cryptograms were considerably different structurally, (perhaps also functionally? From gymnosperms. By the early Carboniferous, these two cell types had diverged.PALEOZIC, (570)
Sieve cells and sieve tube members (elements) diverge. Seed fern Medullosa sieve cells alternate with parenchyma.
Carboniferous (362)
Slightly oblique end walls in cells occurring in the position which should be occupied by phloem in Rhynia and Trimerophyton.
Middle Devonian (~ 415)
MESOZOIC (245)
Cretaceous (145)
In cycadophytes such as Cycadeoidea sieve cells alternate with parenchyma

So, why two systems?
Even given that the xylem and phloem have distinct functions, with o being confined in an apoplasmic environment, and the other in a symplasmic environment, is this necessary?
One could argue that both functionalities could be contained, maintained and carried out in the same tube – or could one?
It could be argued that one does not ‘need’ a confinement process such as those typified by functional phloem, and functional xylem – or could one?
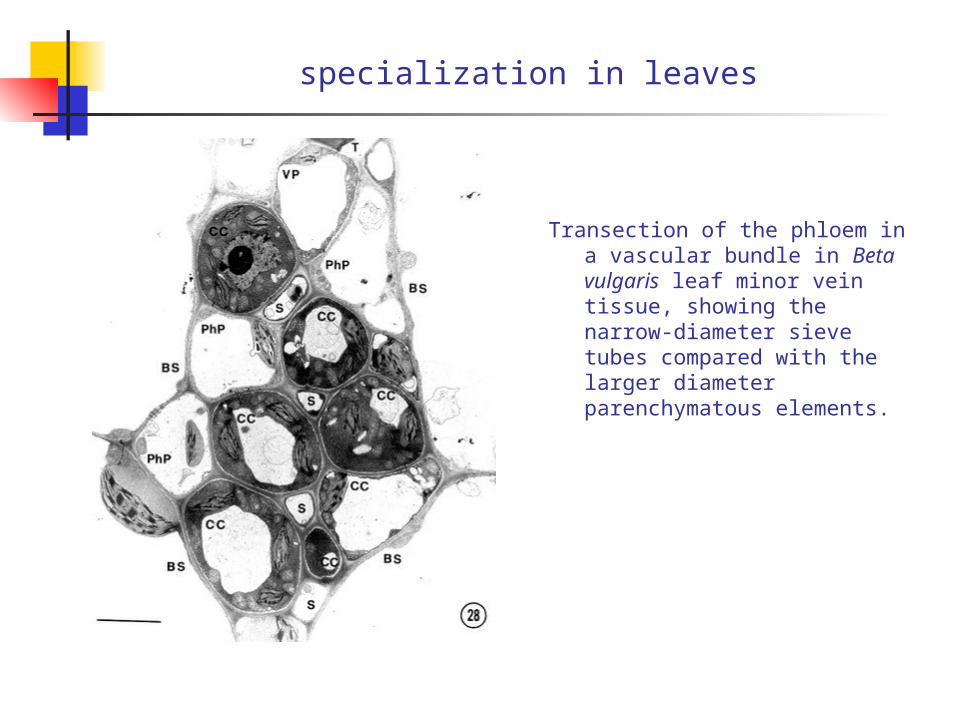
specialization in leaves
Transection of the phloem in a vascular bundle in Beta vulgaris leaf minor vein tissue, showing the narrow-diameter sieve tubes compared with the larger diameter parenchymatous elements.

reading
Behnke, H-D and RJ Sjölund (eds) 1990. Sieve elements. Comparative structure, induction and Development. Springer ISBN 3-540-50783-3.
Carlquist, S.1988. Comparative Wood Anatomy. Springer ISBN 3-540-18827-4. 583.04 Car.
Fahn, A 1967 Plant Anatomy. Pergamon Press. 581.3 Fah

That’s it……….