the global diversity of protozoa and other small species...the global diversity of protozoa and...
TRANSCRIPT

International Journal for Parasitology 28 (199%) 2948
The global diversity of protozoa and other small species B. J. Finlay*
Institute of Freshwater Ecology, Ambleside. Cumbria LA22 OLP, U.K.
Received 30 June 1997; accepted 25 July 1997
Abstract
It is widely believed that the number of species of micro-organisms in the world is extremely large. Here, we offer the contrasting view-that the number may be quite modest. Most of the work reviewed re&rs to the ciliated protozoa. As with all microbial groups, we must define our concept of “species”, and for ciliates, the “morpbospecies” concept appears to be at least as robust as any other. Critical examination of published descriptions of &l&es provides a “best estimate” of 3744 for the global number of free-living morphospecies. Of these, ‘793 are associated with marine sediments, and 1370 with freshwater sediments. In an independent analysis based on extrapolation (assuming the ubiquity of species) from ecological datasets, we estimate the numbers of qecies in marine and freshwater sediments a$ 597 and 732, respectively (i.e. within a factor of two of the figures obtained from taxonomic analysis). This a~entconvergence of independent estimates will strengthen if, as is likely, the number of nominal species is further reduced by taxonomic revision. These relatively low numbers of species are consistent with (a) the vast amount of ~1~~ information indicating typically cosmopolitan distributions for ciliates and other microbes, and (b) recent experimentiti evidence that most free-living ciliates are rare or cryptic-seldom detectable, but present, and “waiting” for suit&le conditions to arrive. In summary, most ciliates (and other micro-organisms) are probably ubiquitous, endemics are rare, global species richness is relatively low, and, at least in the case of the ciliates, most species have already been discovered. C 1998 Australian Society for Parasitology. Published by Elsevier Science Ltd.
Key words: &diversity; protozoa; ciliate; micro-organisms; species richness; cosmopolitan; morphospecies; ubiquity; aggregate
It is not a simpk task to provide accurate esti- mates of giohal species richness in any of the larger taxonomic groups. Even for the well-studied insects, then? is considerable variation in published estimates [I]; and with respect to the micro-organ- isms, the task is generally considered to he rather difficult [2-l. The fundamental problem with micro-organisms is that in many cases there is only
*E-mail: [email protected].
a fragile consensus on what constitutes a species, and until that consensus is strengthened, we cannot have a sound strategy for estimating the global number of species.
Let us begin by looking at a specific example within the protozoa. The rich morphofogical var- iety of ciliated protozoa (Fig. 1) has been inves- tigated for more than 200years, and ciliate morphology has provided the criteria for defining species 18-111. However, this practice has not always been without its prohh~~. The ccmtinuous development of new metho& @A as E!M) for pro- bing deeper .into the morphology of cells has
SOO20-7519/98 %19.00+0.00 0 1998 Australian Society for Parasitology. Published by Elsevier Science Ltd. Printed in Great Britain PII: SOO20-7519(97)00167-7


B. J. Fin& 1 International Journal for Parasitology1 28 i 1998) 2948 31
unravelled layers of complexity and variation that could not have been imagined 200years ago; but perhaps the considerable exploratory potential of these techniques contributes more to illustrating the fractal geometry of protozoa than to the clearer definition of species differences. Moreover, some investigators have better tools than others, and bet- ter tools allow more weight to be placed on specific criteria. Some workers have broad experience of protozoan diversity in the natural environment; others have huge experience of variation in lab- oratory cultures of protozoa. All those who study ciliates and other protozoa, like all naturalists, tend to form a personal view of protozoan diversity. And, like at1 other naturalists, protozoologists tend
to fall into one camp or the other--the “lumpers” or the “splitters”.
It is, therefore, not surprising that if we try to gain some quantitative idea of species diversity in the ciliates, we find a rather murky picture. We find a remarkably large number of synonyms for nominal species, and genera that endure chaotic cycles of expansion and contraction, with occasional “revision” into extinction. Improved clarity might be gained by adopting a more objec- tive approach., and by focusing on specific groups within the protozoa. Therefore, as a first step, we have looked at those ciliates that are capable of an independent existence in the natural environment--- the free-living ciliated protozoa.
Fig. 1. A selection from the variety of morphospecies of free-living ciliates living in fresh waters. Those placed towards the top left are typically found in the open water of lakes; those close to the centre at the base are all anaerobic. The remainder are generally found in sediments and detritus, and attached to submerged surfaces (e.g., aquatic animals and plants). All are drawn to scale (scale bar = 1 mm). (Adapted from Finlay et al. [12].) 1. Cinetochilum margaritaceum; 2. Halteria sp.; 3. Cyclidium citrullus; 4. Urotricha sp. 5. Thigmogaster sp.; 6. Mesodinium sp.; I. Philasterides sp.; 8. Chilodonelia uncinata; 9. Litonotur uninucleatus; IO. Cyclidium plouneaurr; 11. Aspidisca cicada: 12. Lacrymaria sp.: 13. Halteria grandinelia; 14. Trimyema sp.; 15. Zosterodasys sp.; 16. Glaucoma scintillans; i 7. Ctedectoma wilberti; 18. Calyptotricba lanuginosa; 19. Saprodinium sp.; 20. Cristigera setosa; 21. Cyctidium sp.; 22. Pseudomnicrothurax dubius; 23. Sathrophilus muscorum; 24. Paranophrys sp.; 25. Askenasia sp.; 26. Metacystis tessehta; 27. Hastatella radians; 28. Bahmanema biceps; 29. Uronema marinum; 30. Cyclidium glaucoma; 3 1. Colpoda cucullus; 32. Dexiotricha media; 33. Colpidium campylum; 34. Ptacus iuciae: 35136. Tetrahymena pyrijormis-complex; 37. Placus sp.; 38. Cinetochilum margaritaceum; 39. Blepharisma hyalinum; 40. Loxohs SD.; 41. Paramecium aurelia-complex; 42. Lacrymaria olor; 43. Tetrahymena vorax; 44. Entosiphon sp. (euglenid); 45. Spurhidium suhatum; 46. Pleuronema coronatum; 41. Chilodonella sp.; 48. Urocentrum turbo; 49. Ileonema dispar; 50. Nassula tumida; 51. Sugittariapoltganalis; 52. Trithigmostoma cucullulus; 53. Frontonia leucas; 54. Lacrymaria elegans; 55. Paramecium caudatum; 56. Lacrymaria op.; 57. Eqlotes sp.; 58. Drepanomonas revoluta; 59. Tachysoma sp.; 60. Tachysoma pellionella; 61. Styiomychia putrina; 62. Urosoma eienkowski: 63. Holosticha sp.; 64. Keronopsis monilata; 65. Ancystropodium maupasi; 66. Urosoma cienkonski; 61. Gastrostyla steinii; SK. Styloqxhia mytilus; 69. Uroleptuspiscis; 70. Lacrymaria olor; 11. Metopus striatus; 12. Bothrostoma undulans; 73. Tropidoatractuc acuminates; 74. Metopus sp.; 75. Brachonella spiralis; 16. Caenomorpha sp.; 77. Caenomorpha uniseriahs; 18. Saprodinium dentatum; 19. tsocychdium globosum; 80. Plagiopyla nasuta: 81. Metopus es; 82. Saprodinium sp.; 83. Cvclidium porcatum; 84. Discomorphellu pectinata; 85. Mylestoma uncinatum; 86. Saprodinium diffile; 81. Trimyema compressurn; 88. Vaginicola crystallina; 89. Ophrydium versatile; 90. Trichophrya epistylidis; 91, Vorticella natans; 92. Acineta sp.; 93. Loxophyllum heft*s; 94. Chikodonetla sp.; 95. Sphaerophrya magna; 96. Epistylis sp.; 97. Paruroleptus caudatus; 98. Stentor sp.; 99. Stentor polymorphus: 100. Dendrosoma radians; 101. Stylocohz striuta; 102. Aspidisca cosiata 103. Stentor roeseli; 104. Epistylis flavicans; 105. Nassula picta; 106. Paramecium bursaria; 107. Front&a sp.; 108. Deltopylum rhabdoides; 109. Ophrydium eichornii; 110. Loxodes striaius; 11 I. Dileptus sp.; 112. Trachelius ovum; 113. Arcuospathidium vermiforme; 114. Epistylis plicatilis; 115. Kahlilembus attenuatus; 116. Endosphaera terehrans; 117. Vorticella mayert; 118. Litonotus Cygnus; 119. Amphileptus sp.; 120. Litonotus Cygnus; 121. Litonotus fasciola; 122. Loxophyllum hems; 123. Litonotuv fi-tsriofa: 124. Loxophyllum sp.; 125. Hastatella aesculacantha; 126. Hastatella radians; 127. Phascolodon vorticella; 128. Scrombidmm aclox: 129. Bursaria truncatella; 130. Urozona bti’tsehlii; 13 1. Disematostoma gyrans; 132. Urotricha fircata; 133. Monodinium balbiani; 1.34. Actinobolina sp.; 135. Urotricha furcata; 136. Tintinnopsis sp.; 137. Bursaridium pseudobursaria; 138. Prorodon palurlris; 139. His- tiobalantium ma&s; 140. Balanion planctonicum; 141. Hypotrichidium conicum; 142. Dtsematostoma (Leurophrys) tetraedrica: 143. Tintinnopsis laeustris; 144. Lembadion sp.; 145. Enchelyomorpha vermicularis; 146. Tintinmidium fluviatile; 147. Platynamatum so&/e; 148. Prorodon ovum; 149. Pleuronema sp.; 150. Urotricha sp.; 151. Halteria sp.; 152. Strombidium sp.; 153. Strombidium sulcatum; 154. Coleps sp.; 155. Coleps hirtus; 156. Prorodon discolor; 157. Codonella cratera; 158. Strombidium viride; 159. Pro&on sp.: 160. Promdon sp; 161. Bursel~opsisgargamellae; 162. Frontonia acuminata; 163. Strobilidium adhrrens; 164. Spirosromum sp.; 165. Spiro.~tomum minus: I66 Homalozoon vermiculare; 167. Vorticeffa sp.

32 B. J. Finlay 1 International Journalfor Parasitology 28 (1998) 2948
2. The discovery of new ciliate species-historical trends
Figure 2 illustrates the history of the discovery of new free-living ciliate species, from the year 1758 until the end of 1993. These species belong to 774 genera. One genus (Spathidium) contains at least 100 species, and no fewer than 372 genera are rep- resented by a single species. The rate of description of new species has varied greatly over time. There are correlations with historical events such as world wars, and peaks of taxonomic activity can be allo- cated to specific individuals or groups of indi- viduals, e.g., Ehrenberg and Dujardin around 1830-1940; Faur6Fremiet and Penard around 1920, and massive peaks in the 1920s and 1930s due, especially, to the efforts of Kahl. Dragesco et al. are largely responsible for the peaks in the 1950s and 196Os, and Foissner et al. for those in the 1980s. The underlying rising trend in the number of species described in the period since 1945 is about 1% per annum.
In our analysis of this literature, we encountered two related problems. The most obvious was dupli- cation-the same organism being described under different names by different (and sometimes the same!) taxonomist. The second problem was the apparent lack of objective criteria for defining spec- ies-how different do two organisms have to be to have the status of separate species? Each of these problems, of synonyms and species concepts, is now considered.
3. Synonyms and revisions
The rate of change in the number of nominal species depends on opposing forces-the naming of new species, and taxonomic revisions. These forces are obviously unequal in strength, because the num- ber of nominal species continues to rise. Of course, genuinely new species are found, but we must also consider the problem of ignorance of what has pre- viously been published, with the consequence that the same organism is described with different names. These names may subsequently be judged to be synonyms (see Fig. 3).
Taxonomic revision will reduce the rate of
200 (4 I I
1750 1800 1850 1WJO 1050
moo (b> 1
L 2OOQ s
4
g CJ 1000
0 I 1750 18OQ 1850 1900 1950 2000
Year Fig. 2. (a) The number of new species of free-living ciliates described each year. (b) The cumulative number of new species of free-living ciliates described in the international literature from 1758 until the end of 1993. (Adapted from Finlay et al. [12].)
l
Fig. 3. Taxonomic history of the ciliate genus Loxo&s. Arrows are used to connect equivalent species, “forms” and “types”. We have considered only those species that can be unambiguously integrated into the taxonomic history of the genus. Broken lines indicate that the connected species may not be identical. Boxes indicate a statement by the original author that Loxodes exists only as three conspecific “forms” or “types”.

B. J. Finlay / International Journal for Parasitology 28 (1998) 29-48
vii
“TiF
____--- ---- _-_-------------------- ---------I L ---- .i‘e-g~swwTm’
c ------ -_ _ _-- -&.;.;-,-- _-___--- i--+; ---- !qr+ -_--____--
--_ __ - ‘: 4: ----!
+&& qdL.
.“sq q- Ln!k
fk
1 *IUS
r------ ---- --------__------__
: 7-c L-Qsy- A i
LA-
*-

34 B. .I. Finlay / International Journalfor Parasitology 28 (1998) 2948
increase in the number of new species. Consider again the extreme example of Spathidium. The genus has not been revised in the last 60years. It currently contains more than 100 species, and many are very similar to one another. How can it be justified to squeeze more “new” ciliates into such a genus before rationalising the existing complement of nominal species? And if we look at the 730 “new” ciliate species described in the period 1978-1993, we find that many have been allocated to “old” genera; in fact most are placed in genera that have not benefited from revision since 1935, and some have never been revised. More than half of the recent increase in the number of new species is due to species allocated to genera that are probably in need of revision. Taxonomic revisions are a potent force in rationalising the numbers of nominal spec- ies. In most recent, authoritative revisions, the num- ber of nominal species has been reduced to about half, or less, of the original number [ 121.
4. Species concepts
One factor that contributes greatly to the rising number of new ciliate species is the lack of a solid species concept. At present, the boundaries between species are often unclear, and the probability of a new species being “discovered” and described often depends on the techniques and other resources that are available at the time. It is abundantly clear that we need a practicable concept of species that can be applied throughout the whole range of the free- living ciliates.
5. Biological species
Within many nominal species, discrete popu- lations probably exist, between which gene flow is restricted or non-existent. These populations, known as syngens, sibling species or biological spec- ies (the terms are largely interchangeable), are often difficult, even impossible to separate on mor- phological grounds. Paramecium aurelia is one of the best studied of these ciliates, and it has been shown to comprise at least 14 sibling species [13]. But let us take a step back, and look for a moment
at the ciliates as a whole. We find that they are remarkably diverse in the repertoire of their sexual habits, and some manage with no sex at all. We find outbreeders with multiple mating types, inbreeders performing intraclonal conjugation, autogamous ciliates practising self-fertilisation, and forms that are apparently incapable of sex, and so permanently asexual. Paramecium bursaria can form conjugation triplets, with the third partner carrying out autog- amy [14], and up to half of the Tetrahymenas iso- lated from nature were found not to have a micronucleus [ 151, so they were asexual. Are sibling species relevant to our quest for a practicable spec- ies concept for ciliates? The answer, I believe, is “no”; but let us look at some of the evidence.
Paramecium caudatum consists of at least 16 sibling species, but mating is known to occur among four of them [16]. A further complication is that mating type is not always fixed.
The Tetrahymena pyriformis complex currently consists of 16 sibling species [ 171. Four are amic- ronucleate, and thus incapable of mating. Two others can mate with each other, with the pro- duction of viable progeny.
Valbonesi et al. [18] collected 120 strains of Euplotes crassus, of which 38 were observed to mate. These were placed in groups: strains in one group could mate with those in a second group, but not with strains in a third group. Strains in the third group would, however, mate with those in the second group; so genetic exchange could occur, directly or indirectly, among all 38 strains. Such complex mating behaviour cannot be rationalised in terms of any practicable biological species concept.
Although they are undoubtedly biologically interesting, we might question the contribution that biological species make to the business of charac- terising ciliate diversity. A biological species con- cept cannot be applied to sexless and inbreeding ciliates; nor to the complex “mating continuum” found in ciliates such as Euplotes. Mating between sibling species can occur, and only a small fraction of the total number of sibling species, and of the inventiveness of their sexual exploits, is ever likely to be known.
The diversity of cultured tetrahymenine ciliates has also been subjected to sequence analysis of

B. J. Finlay / International Journal for Parasitology 28 ( 1998) 29-48 15
rRNA. Nanney et al. [19] and Jerome and Lynn [20] found that they fell into distinct groups, or “ribosets”. One riboset emerged with eight mor- phologically identical species that were “riboso- mally” identical, and yet all were different, biological species. So, what do these sibling species tell us about ciliate diversity? Two morphologically identical species can be reproductively isolated from each other, but “ribosomally” identical. A further pair of morphologically identical species can be reproductively isolated, yet “ribosomally” quite different from each other. In Tetrahymena, at least, genetic isolation and genetic divergence appear to be poorly correlated, although morphology remains fairly constant. The most plausible expla- nation is that stabilising selection has preserved the typical tetrahymenid morphological phenotype because it probably represents some adaptive peak [21]. In other words, evolution has preserved the morphology that allows representatives of the genus Tetrahymena to continue to carry out the same core ecological functions (e.g., bacterivory in organically-rich particle aggregates) irrespective of the variation that has arisen in their reproductive behaviour and in their ribosomal DNA.
In any event, the number of sibling species described so far is not quantitatively important. Approximately 2000 morphological species have been described in the past 50 years. For sibling spec- ies in the same period, the figure is just over 100.
6. Tbe cibte mqWspecies-towards an operatioaal deSinitio0
In ecological studies in particular, “species diver- sity” of ciliates usually means diversity of form and function [22]. For this reason, the “morphospecies” can be used as a pragmatic definition of “species”: a collection of forms that fit into a defined range of morphological variation; and, most important, forms that apparently occupy the same ecological niche. A ciliate morphospecies might have the fol- lowing characteristics:
1. Morphospecies and the ecological niche. Form and function are closely correlated in ciliates, e.g., “worm-like” interstitial karyorelictids; per-
2.
3.
4.
5.
7.
itrich stalks; the adoral membranellar “umbrella” in planktonic oligotrichs. If two eili- ates are morphologically identical or very nearly so, then it is likely that they occupy identical or very similar niches. In particular, they will probably feed in the same way, on the same types of food particles, in the same type of habitat. Morphological limits. A morphospecies can be defined in terms of limits for various mor- phological characteristics (e.g., cell length, num- ber of kineties, argyrome type). Some characteristics (e.g., the number of k&reties and the number of ciliary units they contain) may be extremely variable and scale with cell size, clonal age, and other factors 1231; but others will be conservative (e.g., argyrome type, type of oral apparatus). The morphological limits will be derived using data from a large number of ceils, from a range of physiological states (see Fig. 4). Global distribution. A morphospecies is usually cosmopolitan in its distribution, and it will thrive wherever it finds a suitable habitat. Variation in some features (e.g., the continuous mor- phological variation in tintinnid loricae [24]) may be correlated with geography. Siblings selfers and sexless species. The mor- phospecies may consist of sibling species, amic- ronucleate-asexual cells, obligatory selfers, and cells that do not or cannot mate. Crme flow may occur between different sibling species; either directly, or through intermediates. Correlation with molecular characteristics. It may be possible to divide a morphospecies into groups, using defined molecular techniques. If these groups cannot also be ascribed specific morphological characteristics, there may be little justification for referring to them as discrete species.
In many cases, the relevant features of both form and function may have less to do with the ciliates themselves than with functional associations involving ciliates and other micro-org;tnisms. Many free-living ciliates, perhaps the majority, harbour

36 B. J. Finlay / International Joumal,for Parasitology 28 (1998) 2948
130 - . .
“0 10 I I 1
50 150 250 350 450 550 650
Cell length (urn Fig. 4. Non-overlapping distributions of morphological characteristics (length of oral aperture and length of cell) in two species of the genus Loxodes (L. magnus and the smaller L. striates). (Adapted from Finlay and Berninger [80].)
symbiotic micro-organisms [25, 261 or organelles (e.g., chloroplasts) sequestered from micro-organ- isms [27, 281. Most anaerobic ciliates have endo- symbiotic methanogens that act as hydrogen sinks [29]; in Paramecium, symbiotic bacteria confer killer traits upon the host [30]. Half of the biomass of the marine interstitial ciliate Kentrophoros consists of ectosymbiotic sulphide-oxidising bacteria which the ciliate farms as a food source [3 I]. An extraordinary diversity of freshwater ciliates harbour unicellular algae that photosynthesise inside the host ciliate [25, 321. These associations are usually permanent or, as in the case of sequestered chloroplasts, con- tinuously re-formed. In all cases, two or more gen- omes permanently co-exist but-more important- the relevant phenotype is not that of the ciliate itself, but that of the consortium. Take the example of Euplotes daidaleos, a ciliate that contains sym- biotic green algae (Chlorellae). Like many other freshwater ciliates, E. daidaleos has a preferred spa- tial niche (Figs 5 and 6)-at, or just below, the oxic- anoxic boundary in the stratified water column of
a pond or lake-a place where it is largely free from predation by aerobic metazoans. Different ciliate species have different methods of coping with life in a low-oxygen environment [29, 33-351. Euplotes daidaleos manages because it makes use of its algal symbionts. It lives at a depth in the water column where there is just enough light for the consortium to carry out net photosynthesis: the symbionts pro- vide all the oxygen required for aerobic respiration by the ciliate, so the consortium becomes an aerobic “island” surrounded by anoxic water [36]. In addition, by living where it does, the consortium is exposed to an elevated concentration of free carbon dioxide which diffuses from the underlying sedi- ment. This ensures that photosynthesis by the sym- bionts is not carbon limited, so the ciliate can receive part of its requirement for organic carbon from the sugars produced and secreted by the sym- bionts. In this case, the ecological niche is filled not by a ciliate, but by a symbiotic consortium that incorporates a ciliate, and it is the phenotype of the consortium on which evolution will operate.

B. J. Finlay / Iniemational Journal for Parasitology 28 (1998) 2948 37
Fig. 5. The ciliate EupZotes daidaleos (length 90 pm), packed with photosynthetic endosymbionts (zoochlorellae). The ciliate was retrieved from anoxic (illuminated) water in a pond. Note the accompanying 16-41 cluster of the anaerobic, photosynthetic bacterium Thiopedia, present at the same depth in the water column. (Adapted from Finlay et al. [36].)
There are published reports that ciliates can lose their symbionts when they are cultivated in the lab- oratory (e.g., the methanogen endosymbionts of the anaerobic ciliate Trimyema compressurn [37]). One possible reason is that the characteristics that con- tribute to fitness in nature may be rather different to those that are relevant in laboratory cultures, and the morphospecies that is selected for in the laboratory may in a sense be an artefact, differing in some important respects from its symbiont-car- rying antecedents in the natural environment.
In nature, ciliates may invariably reproduce themselves in asexual clones. Mutations in these
clones may produce morphological variants, but these may be so minor that they do not reduce significantly the overall fitness of the organism. Propagation of these mutations in asexual clanes will produce iuzdepen&nt lineages with slightly divergent morphotypes. There are several conse- quences: the boundaries of the morphospecies may become rather fuzzy; a morpbaspecies may, in the course of time, produce enough variants to fill out a continuum of forms; and “conservative” mor- phological characters may show systematic vari- ation (e.g., in Euplotes minuta the number of longitudinal ciliary rows changes with clonal age [23]). One way to handle this problem may be to borrow and adapt a device used in plant taxonomy [38]. (NB Higher plants, too, support a very large number of typically asexual species-e.g., dan- delions). The solution is the “aggregate”.
An aggregate is nothing more than a device of convenience, used to group together two or more nominal species that are morphologically similar and difficult to separate. In the case of ciliates, an aggregate would have three key characteristics: (a) it would have no formal taxonomic m;eaning, {b) its name would be permanent, and unaffected by translocations of the nominal species it contains, and (c) it would have no particular phylogenetic significance. Under what circumstances would nominal species be grouped together in an aggre- gate?
1 When they cannot consistently be separated from each other, or when they lie in a continuum of overlapping forms (e.g., cifiates of the cir- rotype 10, double argyrome [Euplotes charon] morpbtypej.
2. When sibling species cannot be discriminated morphologically (e.g., T. pyriformis agg. and Paramecium aurelia agg.).
3. When nominal species can be separated only with molecular methods OF EM of intra-cellular structures (e.g., mitochondria), or on the basis of differences in their geographic distribution.
Adopting this procedure, one can envisage that part of a ‘%pecies” list from an ecological project in the future would look something like the following:

38 B. J. Finlay / International Journalfor Parasitology 28 (1998) 29-48
Eupiotes charon agg. 9. Estimating the global number of morpbospecies
contains at least 13 overlapping nominal species [39] Halteria viridis agg.
contains at least five (probably indistinguishable) nominal species of Halteria and Pelagohalteria with zoochlorellae
Loxodes magaas Stokes, 1887 (morphospecies) Loxodes striatas (Engelmann, 1862) Penard, 1917 (morphospecies) Metopus palaeformzs Kahl, 1927
a morphospecies with several synonyms [40] and a poly- morphic life-cycle
Paramecium aurelia agg. contains all sibling species (primaureiia quadecaurelia)
From our examination of the published litera- ture, the current best estimate for the number of nominal free-living ciliate species is 3744. We would expect this number to remain fairly stable in the future, or to be reduced, possibly to around 3000, in response to new taxonomic revisions of crowded genera. But how good an estimate is this of the global number of ciliate species? In order to answer this question, we have focused on habitats that are relativelv well-studied-the marine interstitial and the freshwater benthos.
0
I Ciliates without
I Ciliates with
symbiotic algae symbic .c algae
Photo irradiance @mol mqs”) l
10 20 30 40 1 I ’ I 9 I -
0, (mol m’“) 0.2 0.4
Free COS1(mol ni”) 0
Fig. 6. Vertical distribution of free-living ciliates, with and without photosynthetic endosymbionts, in the water column of a freshwater pond. Ciliates with endosymbionts produced a distinct abundance peak at 2.6m where there was no detectable dissolved oxygen in the water. (Adapted from Finlay et al. [36].)
2.5

B. J. Finlay / International Journalfor Parasitology 28 (1!?98) 29-48 39
We assume that microbes in general, including ciliates, are ubiquitous. In some places they may be extremely rare or present only as the occasional cyst or other cryptic form, but there is a reasonable chance that a species can be found, at least period- ically, anywhere in the biosphere. This would be due mainly to passive dispersal which in terms of absolute numbers is responsible for dispersal of phenomenally large numbers of protozoa. The agents involved would include hurricanes and tor- nadoes, convective transfer into the atmosphere (with subsequent deposition in rainfall), transport by flying insects and in the damp fur and feathers of migrating animals, in ground water, in the ballast of marine tankers, etc. There is a large literature on this (see [41-44]). If we take the assumption of ubiquity and use it to extrapolate from ecological datasets which relate to relatively small areas, will we obtain global estimates that accord with those obtained by taxonomic analysis? We have tried to answer this question, using large datasets for free- living ciliates in the marine interstitial and in the freshwater benthos. The marine sites were in Scan- dinavia, predominantly Denmark. The freshwater sites were in the United Kingdom and in Nigeria.
The summary data for the numbers of ciliates and ciliate species recorded in these studies appear in Table 1. If these data are then plotted as ranked species (logarithmic) abundances, we find that the curves have linear terminal components (Fig. 7); that is to say, the abundances of the successively rarer species decrease logarithmically. Further- more, the slopes of these trends seem fairly insen- sitive to the size of the datasets. The slope for the Esthwaite Water data is roughly the same as that
Table 1 Summary information for ecological datasets
for the larger dataset that includes all the freshwater benthic sites. This feature is consistent with the idea of the ubiquity of species: if the larger datasets had included a number of species with exclusively local distributions, their respective slopes would be less steep than those for the smaller datasets from smal- ler areas.
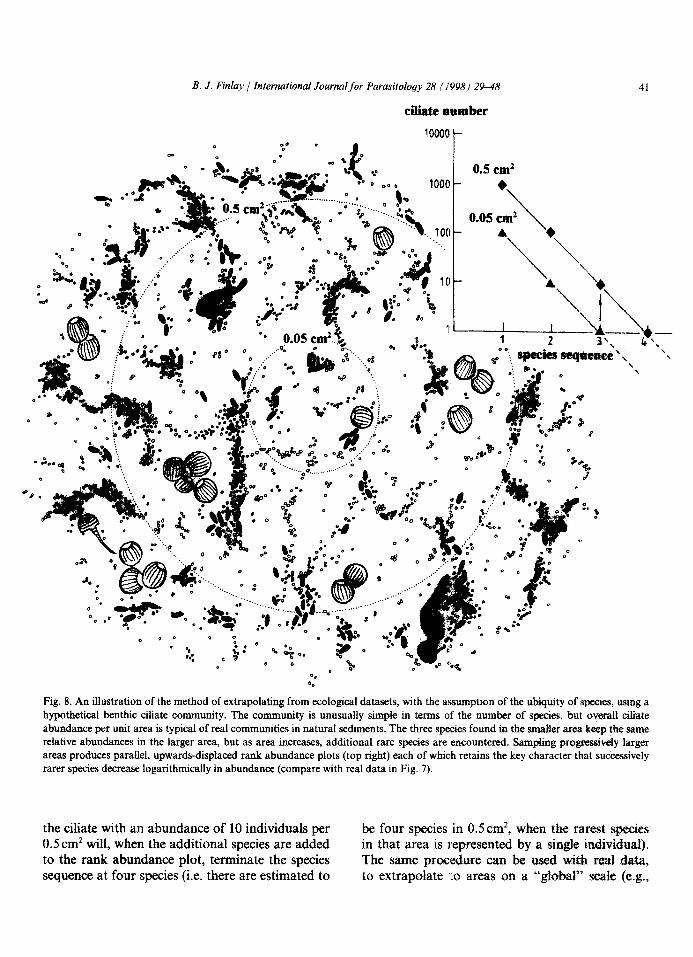
Now, if our datasets had been much larger, as would have been the case if we had examined much larger areas of sediment, we can safely assume that some very rare species would have been revealed. We would also have found a proportionate increase in numbers of the more common species originally recorded. It is reasonable therefore to extend each terminal linear trend below an abundance of one ciliate to indicate the additional species that would be recovered by examining a larger area. This pro- cess is best described with an hypothetical example, as shown in Fig. 8. This shows an unusually simple community of ciliates, but it serves to illustrate the procedure. The inner circle represents 0.05cm2 of sediment. It contains only three species, represented by 100, 10 and 1 individuals, respectively. Now, suppose that we could record every ciliate in an area of sediment 10 times as large (i.e. 0.5 cm’), surrounding and including the inner circle. Our rare ciliate represented by only a single individual in the smaller area would, if it retained the same degree of rarity within the enlarged area, now be rep- resented by an estimated 10 individuals. Moreover, all other ciliate species will keep the same relative abundances the:y had in the smaller area, so the slope of the new, upwards displaced, rank-abun- dance plot will have the same slope as the original. This new theore!.ical plot (for 0.5 cm’) indicates that
-___- Marine interstitial
Freshwater benthos
Helsingrar Beach” All Marine Sites”
Esthwaite Water” All freshwater sitesb
Number of ciliates recorded and identified
48 186 79 342
20 486 35 837
Number of ciliate species recorded _____....._ -...-.. ~_._-.-.. --
85 151
104 125
“Fenchel[8 11. “Finlay [82. 831; Finlay et al. [36, 841.

40 B. J. Finlay 1 International Journalfor Parasitology 28 (1998) 29-48
No. of ciliates
looooo +
1000
100
10
0 20 40 60 80 100 120 140
Species sequtaee Fig. 7. Rank abundance plots for ciliate species recorded from the sediment of a freshwater lake (Esthwaite Water, in England) over a period of 2 years, and from a variety of lakes and rivers (“all freshwater sites”) in the U.K. and in Nigeria. See Table 1.
B. J. Finlay / International Journalfor Parasitology 28 11’998) 2948
ciliaee sumber
10000 t
.'..._ t . . . . ..( f*
w. .....
I‘\ .t
0.05 cm2 . . ..,JOO \
‘,. Al t
41
_. -- I 1*-,---&-
\ 4 *\ , e‘, \ \ \ \
Fig. 8. An illustration of the method of extrapolating from ecological datasets, with the assumption of the ubiquity of species, using a hypothetical benthic ciliate community. The community is unusually simple in terms of the number of species, but overall ciliate abundance per unit area is typical of real communities in natural sediments. The three species found in the smaller area keep the same relative abundances in the larger area, but as area increases, additional rare species are encountered. Sampling progressively larger areas produces parallel, upwards-displaced rank abundance plots (top right) each of which retains the key character that successively rarer species decrease logarithmically in abundance (compare with real data in Fig. 7).
the ciliate with an abundance of 10 individuals per be four species in 0.5 cm’, when the rarest species 0.5 cm2 will, when the additional species are added in that area is represented by a singe individual). to the rank abundance plot, terminate the species The same procedure can be used with real data, sequence at four species (i.e. there are estimated to to extrapolate to areas on a “gfobar’ s&e (e.g.,

42 B. J. Finlay ] International Journalfor Parasitology 28 (1998) 29-48
2 x lo6 km’-the area of inland fresh waters in the world). If we do this, the projected total is 597 species for the marine interstitial, and 732 species of freshwater benthic ciliates.
How realistic are these extrapolations from eco- logical data obtained for relatively small areas? We find that they are within a factor of two of the numbers of species derived from an analysis of spec- ies descriptions in the international published litera- ture (Fig. 9). Furthermore, there is good reason for believing that the correspondence between the two types of estimate may be even better than this, because, as discussed above, the estimate from taxo- nomic analysis is probably still too high. So, our converging estimates of global species richness may, in the course of time, flow even closer to each other than they do at present.
These estimates are also in line with some published, authoritative guesses. After examining marine interstitial ciliates in different parts of the world, Dragesco [45] suggested that “the total num- ber of mesopsammic ciliates must certainly exceed 600”. Some, however, may question our estimates as being rather low (e.g., [46]), for we are saying that there are only 1300-2200 niches for free-living ciliates living in marine and freshwater sediments. Unfortunately it is difficult to carry this argument
much further because it is even more difficult to characterise the niches of ciliates than it is to describe ciliate morphospecies. Moreover, a poten- tial ciliate niche with an adequate supply of suitable bacteria or micro-algae as food may also be a poten- tial niche for some heterotrophic flagellates, gastrotrichs, rotifers, nematodes and other micro- fauna. Ciliates may not have unhindered access to such a wide variety of niches as is commonly imagined.
10. Cosmopolitanism and endemics
We have assumed throughout that the great majority of ciliate species probably do have cosmo- politan distributions and that as a consequence, their global species number is limited. What evi- dence is there for this?
As with most protists [44,47-561, there is no good evidence that ciliates have a biogeography. On the contrary, it seems that the same species are found wherever their “preferred” habitat is found (e.g., [571). Second, local species richness seems to be a significant proportion of global species richness. The number of ciliate species that can be recovered
THE GLOBAL NUMBER OF FREE-LIVING CILIATE SFECIES
Summary information from independent datasets
Taxonomic analysis total piiJ
Lt t marine 1592 n 121521 non-marine
tit d\
marine 17931 m 17821 113701 ftwhwa&r interstitial 15971 other
marine non-marme Other. m beathos
K f Extrapolation from ecological datasets
Fig. 9. The convergence of independent estimates (from taxonomic analysis, and by extrapolation from ecological datasets), for the global number of free-living ciliates living in the marine interstitial and in the freshwater benthos.

B. J. Finlay / International Journal for Parasirologv 28 ( 1998) 29-48 43
from 100 cm’ of freshwater or marine sediment rep- resents lO-20% of the global species number [58, 591. Third, the diversity of free-living species that have been described in the international literature is relatively small and unlikely to increase sig- nificantly in the future [12]. Fourth, ciliates and other protists have high absolute abundance and, in many cases, effective passive dispersal (e.g., [4, 43, 44, 60-633). They are continually being dis- tributed “everywhere”, and newly-formed habitats (e.g., freshwater ponds) are rapidly colonised (e.g., [64, 651). Protists are, in fact, ubiquitous.
But perhaps this assumption is suspect-perhaps we are unable to detect subtle but important differ- ences separating species, so we identify ciliates from different places as the same species only because we are unable to tell them apart. A “cosmopolitan morphospecies” could consist of many similar spec- ies, each with its own geographical distribution. There is, however, one piece of evidence indicating that this is not usually the case. In those ciliate genera in which reproductively isolated populations (sibling species) are known, we might expect this reproductive isolation to be correlated with geo- graphic isolation, but the evidence is to the contrary. Most sibling species in the P. aureliu com- plex have cosmopolitan distributions [66], and many in the T. pyrifarmis complex have been found on two or more continents [67, 681. Strains of Stylonychia Iemnae isolated in North America con- jugated with those isolated in Europe, producing viable ex-conjugants; and the two groups of strains were genetically very similar [69]. It appears as if those examples holding the most promise of reveal- ing a species biogeography within a common mor- photype, fail to do just that.
A further argument against cosmopolitanism is fuelled by the so-called “endemics”. These tend to be found in unusual or poorly-studied habitats, such as solution lakes [70], wetlands in tropical Africa [9] or sea-ice in the Antarctic [71]. But is it not likely that “endemics” are found in these places because of the habitats that the places provide? Perhaps, if the habitat was found elsewhere, we would find the same species there also. A good example is provided by the “endemics” of Antarctic sea-ice that were subsequently found in the Arctic [71, 721. And there are many other examples. Wil- bert and Kahan [73] described a very large and
unusual ciliate (Condylostomu reicki) from Solar Lake in Sinai. Iit was subsequently found in tropical Africa [9], but before it could acquire the epithet of a pantropical distribution, Petz et al. ]71] described a Condyloston;ra in Antarctic sea-ice which, they believed, most closely resembled C. reichi. Endemics tend to acquire a broader geographical distribution in. response to additional sampling effort. Thus Bryometopus hawaiiensis, an “endemic” of the Hawaiian archipelago [74], was soon found in wet moss by a river in central Spain [75]. There are many other exmples from non- ciliate protists. Among the more spectacular of recent discoveries is the ana.e~obir with long rod-,shaped bacteria, discovered in the deep anoxic basin of Mariager Fjord in Denmark and named Postgaardi ?riar@pem& (Fig. 10) [76]. The basin is separated from the sea by a 20 km-long channel of oxygenated brackish water. 14n identical flagellate has now been found in the anaerobic, sulphidic zone of a meromictic lake in the Vestfold Hills of Antarctica [77].
11. Ubiquity
Ciliate species are ubiquitous, and if they find an appropriate habitat on different continents they will be termed cosmopolitan. This is so, even for species living in habitats that are relatively rare. Those living and growing on unusual “islands’” that are separated by large distances (e.g., the sea-ice of the Arctic and the Antarctic) may never have been detected in intermediate regions, but the available evidence does indicate the reality of global dispersal of these “island” species, even if the magt&ude of this dispersal is small compared with that of the many common ciliate morphospecies co-occurring in common habitats.
But if ciliates are ubiquitous, we should, with sufficient patience, be able to find them in any suit- able sample of the natural environmeat. This is what we have now begun to do. We simply took a small sample of sediment from a freshwater lake and, in a separate experiment, a small sampie of marine sand, and manipulated them to produce a variety of niches for the wide range of cift‘ate species that we suspected might be present. The manipu-

44 B. J. Finlay 1 International Journal for Parasitology 28 (1998) 2948
Fig. 10. The anaerobic flagellate Postgaardi mariagerensis with ectosymbiotic bacteria, recently found in the deep, anoxic basin of a Danish fjord (Fenchel et al. [76] [including illustration]), and in the sulphidic layer of a freshwater pond in the Antarctic (Simpson et al. [77]).
lations involved treatment of sediment with various temperatures, redox gradients, food sources, light regimes, etc. and various combinations of these. These simple procedures did create niches for popu- lation growth by a previously hidden diversity of ciliate species. Although only 20 species could be detected in a small volume of sediment freshly col-
lected from a small pond, subsequent manipu- lations encouraged the growth of a total of 135 species over a 3-month period (Fig. 11). It is likely that with more patience, and more imaginative manipulations of sediment, further ciliate niches would have been created and an even greater variety of “ubiquitous” ciliates would have revealed them- selves. The total for the pond is now (as at June 1997) 203 species, and the number continues to increase as new micro-habitats within the pond are examined in detail. This total, from this one small pond, represents about 20% of the total for fresh- water benthic sites worldwide, so local species rich- ness is a significant proportion of the global species number-as we would expect for a group of pre- dominantly ubiquitous organisms.
Perhaps an even more impressive example is afforded by the representatives of the chrysomonad genus Paruphysomonas living in the same l-hectare pond. At present, 50 described species are known from around the world-all of them discriminated on the basis of the scale morphology (incidentally, only a single species was known until these flagel- lates were examined in greater detail using EM- see Andersen [78]). Recent examination of a few millilitres from the sediment-water interface in the pond has so far uncovered 27 of these 50 species (KJ Clarke, personal communication).
Finally, we can illustrate just how different is global biodiversity at the microbial level. Although a large proportion of all microbial species can be found in a small area, the rate of addition of new species falls off rapidly with increasing area sampled. The best known general equation for the species-area relation in macroscopic animals and plants is: S= CA”, where S is number of species, A is area, and C and z are constants that vary from one group of organisms to another [79]. In most studies of the macrofauna and flora of islands and continents, z takes a value in the range 0.12-0.35. The value is usually lower (0.12-O. 17) for organisms such as birds that are easily dispersed, and it is higher for animals such as land snails living on islands. The average slope of the species-area relation for ciliates (z=O.O43; Fig. 12) falls well below either of these ranges. This is consistent with the high rates of dispersal assumed for ciliates, other protozoa and probably all free-living micro-organ- isms, providing yet another illustration of the theme

B. J. Finlay / International Journal for Parasitology 28 (I 1998) 29-48 45

46 B. J. Finlay / International Journal for Parasilology 28 (1998) 29-48
insects = 0.31 Ciliates = 0.04
6
5
T 1
-10 -5 0 5 10
Log Area (km2) Fig. 12. Species-area relationships for free-living ciliates obtained by extrapolation from two large independent ecological datasets (see Table 1): upper curved line, freshwater benthos; lower curved line, marine interstitial. The two superimposed triangles mark the global estimates from taxonomic analysis for freshwater benthic (open triangle) and marine interstitial (filled triangle) ciliates. The latter are in fair agreement with the low slope extrapolated from the ecological datasets, with the assumption of species ubiquity. This is in marked contrast to the much higher slope of the regression drawn for the insects (using data provided by Gaston [l]) which, like most macrofauna and flora, do have a biogeography. (Adapted from Finlay et al. [85].)
that has run through this presentation: most ciliates (and other micro-organisms) are probably ubiqui- tous, endemics are rare, global species richness is relatively low and, in the case of ciliates, most spec- ies have already been discovered.
AcknowledgementsThis paper is based on work carried out in the last few years with financial support from the Natural Environment Research Council (U.K.). The British Council (new IMAGES) provided financial support for the presentation of this lecture in Australia.
References
[l] Gaston KJ. Regional numbers of insect and plant species. Functional Ecol. 1992; 6: 243-247.
[2] May RM. How many species? Phil Trans R Sot Lond 1990; 330: 293-304.
[3] Vickertnan K. The diversity and ecological significance of protozoa. Biodivers Conserv 1992; 1: 334-341.
[4] Fenchel T. There are more small than large species? Oikos 1993; 68: 375-378.
[S] O’Donnell AG, Goodfellow M, Hawksworth DL. Theor- etical and practical aspects of the quantification of biodi-

B. J. Finlay / International Journalfor Parasitology 28 (1998) 29-48 47
versity among microorganisms. Phil Trans R Sot Lond B 1994; 345: 65-73.
[6] Boucher G, Lambsbead JD. Ecological biodiversity of mar- ine nematodes in samples from temperate, tropical, and deep-sea regions. Conserv Biol 1995; 9: 1594-1604.
[7] UNEP. In: Heywood VH, executive editor. Global biodi- versity assessment. Cambridge: Cambridge University Press, 1995.
[8] Corliss JO. The ciliated protozoa. Oxford: Pergamon Press, 1979.
[9] Dragesco J, Dragesco-Kern&is A. CiliCs libres de l’Afrique intertropicale. Collection faune tropicale No. 26. Paris: ORSTOM, 1986.
[IO] Lynn, D. Systematics of ciliates. In: Hausmann K, Brad- bury PC, editors. Ciliates: cells as organisms. Stuttgart: Gustav Fischer, 1996;51-72.
[l l] Lynn DH, Corliss JO. Ciliophora. In: microscopic anatomy of invertebrates, vol 1, Protozoa. Wiley-Liss, 1991; 333- 467.
WI
u31
1141 iI51
[I61
Cl71
1181
1191
PO1
WI
PI
[231
[241
PI
Finlay BJ, Corliss JO, Esteban GF, Fenchel T. Biodiversity at the microbial level: the number of free-living ciliates in the biospbere. Q Rev Biol 1996; 71: 221-237. Sonnebom TM. The Paramecium aurelia complex of 14 sibling species. Tram Am Microsc Sot 1975; 94: 155-178. Greil KG. Protozoology. Berlin: Springer-Verlag, 1973. Preparata RM, Meyer EB, Preparata FP, Simon EM, Vossbrinck CR, Nanney DL. Ciliate evolution: the riboso- ma1 phylogenies of the tetrahymenine ciliates. J Mol Evol 1989; 28: 427-441. Khadem N, Gibson I. Enzyme variation in Paramecium c&turn. J Protozool 1985; 32: 622-626. Dini F, Nyberg D. Sex in ciliates. Adv Microb Ecol 1993; 13: 85-153. Valbonesi A, Ortenzi C, Luporini P. The species problem in a ciliate with a high multiple mating type system, Euplotes crcIssus. J Protozool 1992; 39: 45-54. Nanney DL, Meyer EB, Simon EM, Preparata R-M. Com- parison of ribosomal and isozymic phylogenies of tetrahy- menine ciliates. J Protozool 1989; 36: l-8. Jerome CA, Lynn DH. Identifying and distinguishing sib- ling speties in the Tetrahymena pyriformis complex (Cili- ophora, Oligohymenophorea) using PCR/RFLP analysis of nucleolar ribosomal DNA. J Eukaryot Microbial 1996; 43: 492-497. Nanney DL. Genes and phenes in Tetrahymena. Bioscience 1982; 32: 783-788. Finlay BJ, Fenchel T. Ecology: role of ciliates in the natural environment. In: Hausmann K, Bradbury PC, editors. Cili- ates: cells as organisms. Stuttgart: Gustav Fischer, 1996;417440. Frankel J. An analysis of the spatial distribution of ciliary units in a ciliate, Euplotes minuta. J Embryo1 Exp Morph01 1975; 33: 553-580. Boltovskoy D, Alder VA, Spinelli F. Summer Weddell Sea microplankton: assemblage structure, distribution and abundance, with special emphasis on the tintinnina. Polar Biol 1989; 9: 447456. Finlay BJ. Physiological ecology of free-living protozoa. Adv Microb Ecol 1990; 11: l-35.
[26] Gdrtz H-D. Symbiosis in ciliates. In: Hausmann K. Brad-
bury PC, editors. C&&es: cells as organisms. Stuttgart: Gustav Fischer, 1996;441-462.
[27] Stoecker DK., Michaells AE, Davis LH. Large proportion of marine planktonic cilia&s found to contain functional chloroplasts. Nature 1987,326: 790-792.
[28] Jonsson PR. Photosynthetic assimilation of inorg&c car- bon in marine oligotrich ciliates (Ciliopbora, Oligo- trichina). Mar Microb Food Webe 1987; 2: 5568.
[29] Fenchel T, Finlay BJ. In: May RM, Harvey P, editors. Ecology and evolution in anoxic worlds. Oxford series in ecology and evolution. Oxford: Oxford University Press, 1995;276.
[30] Quackenbush RL. Endosymbionts of killer paramecia. In: Gtirtz H-D, editor. Paramecium. Berlin: Springer-Verlag,
[311
1321
[331
[341
[351
1361
[371
[381
[391
[401
[411
[421
[431
WI
[451
1988;406-418. Fenchel T, Finlay BJ. Kentrophoros: a mautbless ciliate with a symbiotic kitchen gar&. Qpbelia 1989; 30: 75-93. Finlay BJ, Beminger U-G, Clarke KS, Cow&& AJ, Hindle RM, Rogerson A. On the abundance and diattibution of protozoa and their food in a productive freshwater pond. Eur J Protistol 1988; 23: 205-217. Finlay BJ. Oxygen availability and seasonal migrations of ciliated protozoa in a freshwater lake. J Gen Microbial 1981; 123: 173-178. Finlay BJ. Nitrate respiration by protozoa (Loxodes spp.) in the hypolimnetic nitrite maximum of a freshwater pond. Freshw Biol 1985; 15: 333-346. Finlay BJ, Fenchel T. Photosensitivity in the ciliated pro- tozoon Loxo&~ pigment granules, absorption and action spectra, blue tight perception and ecological significance. J Protozool1986; 33: 534-542. Finlay BJ, Maberly SC, Esteban G. Spectacular abundance of ciliates in anoxic pond water: contributim of symbiont photosynthesis to host respiratory requirements. FEMS Microbial Ecol 1996; 20: 229-235. Wagener S, Bardele CF, Pfennig N. Functional integration of Methanobacterium formieicum into the anaerobic ciliate Trimyema compressurn. Arch Microbial 1990: 153: 496- 501. Heywood VH The *‘species aggregate” in theory and prac- tice. In: Symposium on biosystematics. Utrecht: Inter- national Bureau for Plant Taxonomy and Nomenclature, 1963. Jones TC, Gates MA. A morphometric analysis of the Euplotes charon morphotype (Ciliophora: Euplotida). J Eukaryot Microbial 1994; 41: 441-450. Esteban G, Fenchet T, Finlay BJ. Diversity of free-living morphospecies in the ciliate genus Metopus. Arch Pro- tistenk 1995; 146: 137-164. Gislkn T. Aerial plankton and its conditions of life. Biol Rev 1948; 23: 109-126. Tailing JF. The element of chance in pond populations. Naturalist 1951;Oct-Dec:157-170. Maguire B. The passive dispersal of small aquatic organ- isms and their colonization of isolated bodies of water. Ecol Monogr 1963; 33: 161-185. Kristiansen J. Dispersal of freshwater algae--a review. Hydrobiology 1996; 336: 151-157. Dragesco J. On the biology of sand-dwelling ciliates. Sci Prog 1962; 50: 353-363.
[46] Levandowsky M, Corliss JO. Arguments with Hutner. or

B. J. Finlay / International Jourrzal.for Parasiloiog.v 28 (1998) 29-48 48
1471
1481
1491
WI
WI
1521
[531
[541
[551
t561
[571
[581
[591
why there are so few kinds of protozoa. J Protozool 1977; of the Tetrahymenapyriformis complex. Trans Am Microsc 24: 481483. Sot 1976; 95: 664682. Sandon H. The composition and distribution of the proto- zoan fauna of the soil. Edinburgh: Oliver & Boyd, 1927. Lackey JB. A study of some ecologic factors affecting the distribution of protozoa. Ecol Monogr 1938; 8: 503-527. Bary BM. Studies on the freshwater ciliates of New Zealand. Part II. An annotated list of species from the neighbourhood of Wellington. Proc R Sot NZ 1950; 78: 31 l-323.
WI
F91
Stout JD. Reaction of ciliates to environmental factors. Ecology 1956; 37: 178-191. Ogden CC, Hedley RH. An atlas of freshwater testate amoebae. Oxford: British Museum/Oxford University Press, 1980. Larsen J, Patterson DJ. Some flagellates (Protista) from tropical marine sediments. J Nat Hist 1990; 24: 801-937. Bowers NJ, Pratt JR. Estimation of genetic variation among soil ciliates of Colpoda infura (Stokes) (Protozoa: Ciliophora) using the polymerase chain reaction and restric- tion fragment length polymorphism analysis. Arch Pro- tistenkd 1995; 145: 29-36. Ekebom J, Patterson DJ, Vlars N. Heterotrophic flagellates from coral reef sediments (Great Barrier Reef, Australia). Arch Protistenkd 1996; 146: 251-272. Tyler PA. Endemism in freshwater algae. Hydrobiologia 1996; 336: 127-135. Patterson DJ, Simpson AGB. Heterotrophic flagellates from coastal marine and hypersaline sediments in Western Australia. Eur J Protistol 1996; 32: 423448. Smith HG. The distribution and ecology of terrestrial protozoa of sub-Antarctic and maritime Antarctic islands. Sci Rep Br Antarct Surv, No 95. Cambridge: BAS/NERC, 1978.
[701
[711
Corliss JO, Daggett P-M. “Paramecium aurelia” and “Tetrahymena pyriformis”; current status of the taxonomy and nomenclature of these popularly known and widely used ciliates. Protistologica 1983; 19: 307-322. Ammermann D, Schlegel M, Hellmer K-H. North Amer- ican and Eurasian strains of Stylonychia lemnae (Cili- ophora, Hypotrichida) have a high genetic identity, but differ in the nuclear apparatus and in their mating behavior. Eur J Protistol 1989; 25: 67-74. Esteban G, Finlay BJ, Embley TM. New species double the diversity of anaerobic ciliates in a Spanish lake. FEMS Microbial Lett 1993; 109: 93-100. Petz W, Song W, Wilbert N. Taxonomy and ecology of the ciliate fauna (Protozoa, Ciliophora) in the endopagial and pelagial of the Weddell Sea, Antarctica. Stapfia 1995; 40: l-223.
1721 Agatha S, Spindler M, Wilbert N. Ciliated protozoa (Cili- ophora) from Arctic sea ice. Acta Protozool 1993; 32: 261- 268.
1731
[741
[751
[761
[771
Finlay BJ, Esteban GF, Fenchel T. Global diversity and body size. Nature 1996; 383: 132-133. Fenchel T, Esteban GF, Finlay, BJ. Local versus global
1781
1791
WI
Wilbert N, Kahan D. Ciliates of Solar Lake on the Red Sea shore. Arch Protistenkd 1981; 124: 70-95. Foissner W. Bryometopus hawaiiensis sp n., a new colpodid ciliate from a terrestrial biotope of the Hawaiian Archi- pelago. Ann Naturhist Mus Wien 1994; 96B: 19-27. Olmo JL, Tellez C. An European population of Bry- ometopus hawaiiensis Foissner, 1994 (Protozoa: Cili- ophora). Acta Protozool 1996; 35: 317-320. Fenchel T, Bernard C, Esteban G, Finlay BJ, Hansen PJ, Iversen N. Microbial diversity and activity in a Danish fjord with anoxic deep water. Ophelia 1995; 43: 45-100. Simpson AGB, van den Hoff J, Bernard C, Burton HR, Patterson DJ. The ultrastructure and systematic position of the euglenozoon Posrgaardi mariagerensis Fenchel et al. Arch Protistenkd 1997; 147: 2 13-225. Andersen RA. Diversity of eukaryotic algae. Biodivers Conserv 1992; 1: 267-292. MacArthur RH, Wilson, EO. The theory of island biog- eography. Princeton, NJ: Princeton University Press, 1967. Finlay BJ, Beminger U-G. Coexistence of congeneric cili- ates (Karyorelictida: Loxodes) in relation to food resources in two freshwater lakes. J Anim Ecol 1984; 53: 929-943. Fenchel T. The ecology of marine microbenthos, IV. Struc- ture and function of the benthic ecosystem, its chemical and physical factors and the microfauna communities with special reference to the ciliated protozoa. Ophelia 1969; 6: 1-182.
diversity of microorganisms: cryptic diversity of ciliated protozoa. Oikos 1997; 80: 220-225.
[60] Parsons WM, Schlichting HE, Stewart KW. In-flight trans- port of algae and protozoa by selected Odonata. Trans Am Microsc Sot 1966; 85: 52&527.
[61] Maguire B, Belk D. Paramecium transport by land snails. J Protozool 1967; 14: 445447.
[62] Schlichting HE, Sides SL. The passive transport of aquatic microorganisms by selected Hemiptera. J Ecol 1969; 57: 759-764.
[63] Finlay BJ, Tellez C, Esteban G. Diversity of free-living ciliates in the sandy sediment of a Spanish stream in winter. J Gen Microbial 1993; 139: 2855-2863.
[64] Scourfield DJ. The nannoplankton of bomb-crater pools in Epping Forest. Essex Nat 1944; 27(9): 23 1-241.
[65] Holmberg 0, Pejler B. On the terrestrial microfauna of Surtsey during the summer of 1970. Surtsey Res Prog Rep 1972; 6: 69-72.
[66] Nyberg, D. The species concept and breeding systems. in: GBrtz H-D, editor. Paramecium. Berlin: Springer-Verlag, 1988;41-58.
[67] Nanney DL, McCoy JW. Characterization of the species
t811
P21
1831
P41
P51
Finlay BJ. Temporal and vertical distribution of cilio- phoran communities in the benthos of a small eutrophic loch with particular reference to the redox profile. Freshw Biol 1980; 10: 15-34. Finlay BJ. Effects of seasonal anoxia on the community of benthic ciliated protozoa in a productive lake. Arch Protistenkd 1982; 125: 215-222. Finlay B, Bannister P, Stewart J. Temporal variation in benthic ciliates and the application of association analysis. Freshw Biol 1979; 9: 45-53. Finlay BJ, Esteban GF, Fenchel T. Protozoan diversity: converging estimates of the global number of free-living ciliate species. Protist 1998 (in press).