the integration of recombination and physical maps in a large-genome monocot using haploid genome...
Post on 20-Dec-2015
219 views
TRANSCRIPT

The Integration of Recombination and Physical Maps in a Large-Genome Monocot Using Haploid Genome Analysis in a Trihybrid Allium Population
Khrustaleva et al., 2005

Constructing integrated genetic and physical maps
Four approaches:
I. Construction of physical map by contig assembly of large insert DNA clones (BACs and YACs).
II. Combination of in situ hybridization of BACs or YACs on plant chromosomes with recombination maps
III. Use deletion or translocation lines to create physical landmarks on genomes and to relate these to recombination maps
IV. Visualization of recombination points in interspecific hybrids via genomic in situ hybridization (GISH)

Research GoalsI. Construct integrated physical and recombination maps of a
large-genome monocot (Allium)
II. Contribute to the study of chromosome organization in these species

Why this is important:
Enables a thorough analysis of recombination frequencies along Allium chromosomes
Allows for a comparison of genetic and physical distances
Makes it possible to study the physical distribution of two types of AFLP markers produced by restriction enzyme combinations sensitive (PstI/MseI) and nonsensitive (EcoRI/MseI) to methylation

Procedure
Utilized GISH and AFLP technology to construct integrated maps
Used Allium trihybrid population:
Allium cepa X (Allium roylei x Allium fistulosum)
Compared AFLP profiles of individual genotypes with corresponding recombinant chromosomes
Constructed integrated physical and recombination maps of chromosome 5 and 8 for A. roylei and A. fistulosum simultaneously
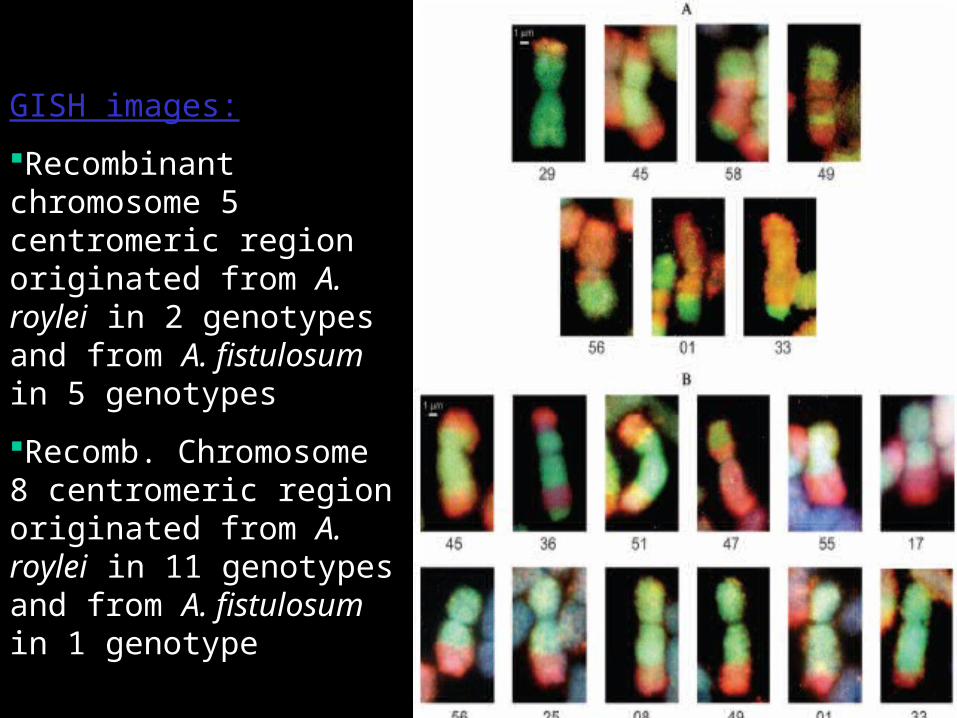
GISH images:
Recombinant chromosome 5 centromeric region originated from A. roylei in 2 genotypes and from A. fistulosum in 5 genotypes
Recomb. Chromosome 8 centromeric region originated from A. roylei in 11 genotypes and from A. fistulosum in 1 genotype
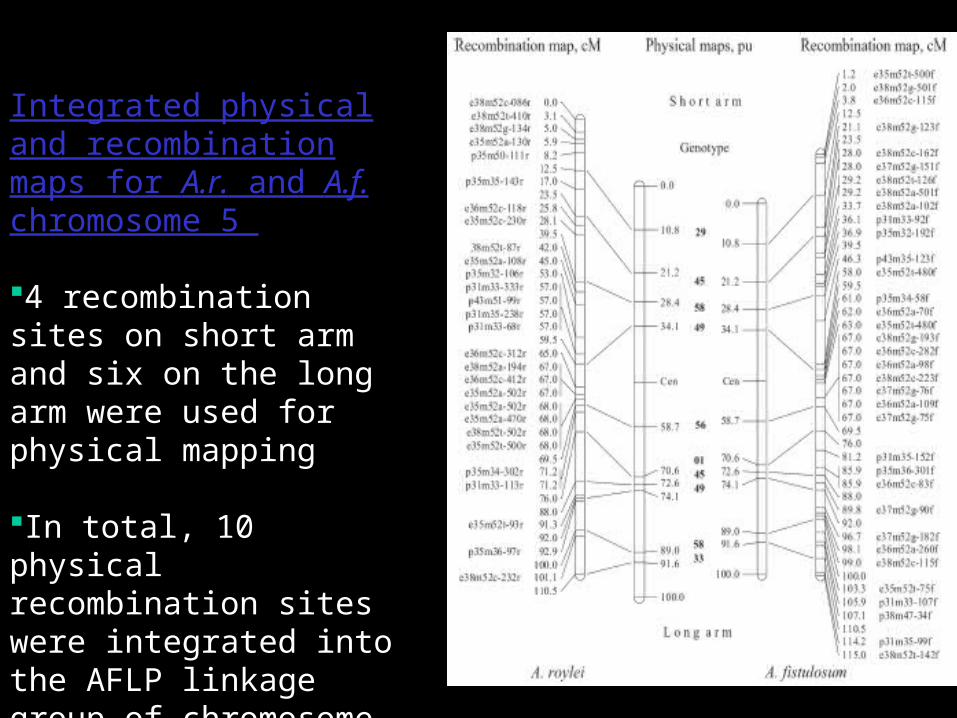
Integrated physical and recombination maps for A.r. and A.f. chromosome 5
4 recombination sites on short arm and six on the long arm were used for physical mapping
In total, 10 physical recombination sites were integrated into the AFLP linkage group of chromosome 5

Integrated physical and recombination maps for A.r. and A.f. chromosome 8
4 recombination sites on proximal half of short arm and 10 on the long arm were used for physical mapping (8 located in interstitial part, 2 in distal part
In total, 14 physical recombination sites were integrated into the AFLP linkage map of chromosome 8
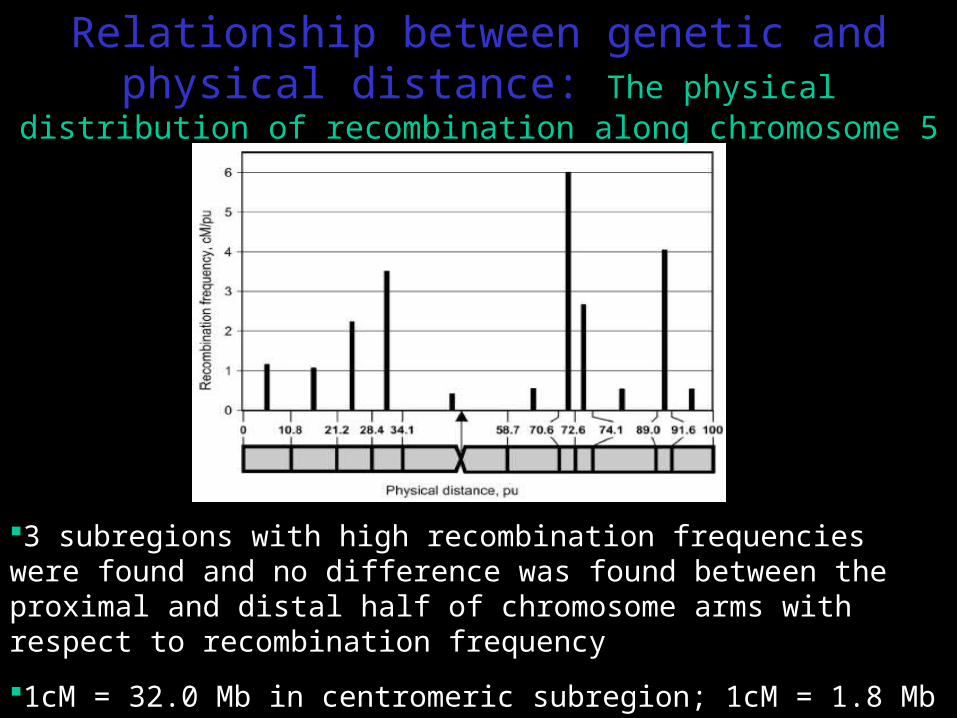
Relationship between genetic and physical distance: The physical distribution of recombination along
chromosome 5
3 subregions with high recombination frequencies were found and no difference was found between the proximal and distal half of chromosome arms with respect to recombination frequency
1cM = 32.0 Mb in centromeric subregion; 1cM = 1.8 Mb in subregion with highest recombination frequency (70.6-72.6 pu)
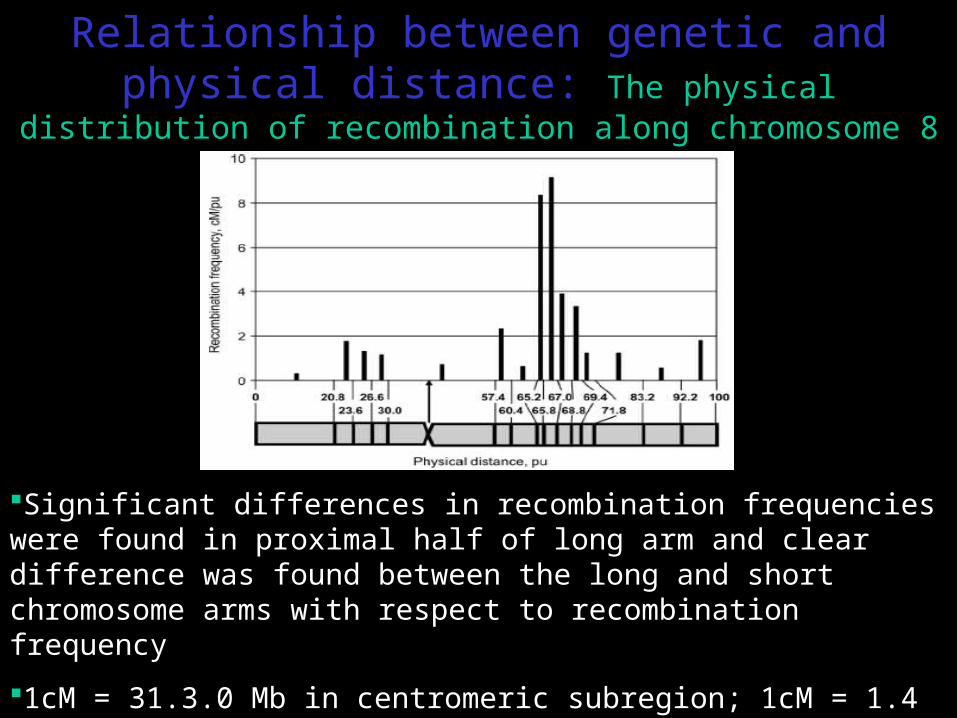
Relationship between genetic and physical distance: The physical distribution of recombination along
chromosome 8
Significant differences in recombination frequencies were found in proximal half of long arm and clear difference was found between the long and short chromosome arms with respect to recombination frequency
1cM = 31.3.0 Mb in centromeric subregion; 1cM = 1.4 Mb in subregion with highest recombination frequency (65.8-67.0 pu)

Physical distribution of AFLP markers for chromosome 5
Uneven distribution of the 64 AFLP markers from linkage group assigned to chromosome 5
Short arm: high density = 1.9 markers/pu in proximal subregion
Long arm: high density = 1.5 markers/pu in 2 subregions and 1.3 markers/pu in 1 subregion

Physical distribution of AFLP markers for chromosome 8
Uneven distribution of the 56 AFLP markers from 15 physical subregions on chromosome 8; 78.6% of markers on long arm
Proximal long arm: high density in 4 regions = 1.7 markers/pu in 1st, 3.3 markers/pu in 2nd, 2.5 markers/pu in 3rd, 1.7 markers/pu
Marker density in short arm low in all subregions (0.2-1.1 markers/pu)

Conclusions: comparisons of Allium with other large-genome monocots
Similarities:
Recombination hotspots restricted to few chromosomal regions
Physical density of markers corresponds closely to distribution of recombination
High degree of suppression of recombination in centromeric regions
Differences:
Recombination predominantly occurs in proximal half of chromosome arm
~58% of PstI/MseI markers occur in proximity of centromeric region

Possible explanations of differences
High level of recombination in proximal half of Allium chromosomes suggests a high density of genes in this area
Gene density higher in distal regions for cereals
Recombination events occur in gene-rich areas
Or, variation of recombination rates could be due to sequence heterologies in distal parts of A. roylei and A. fistulosum chromosomes
Sequence heterologies can significantly reduce recombination rate
Low degree of sequence identity between the two parental homeologs

Possible explanations of differences
~58% of PstI/MseI markers proximally located on Allium chromosomes
PstI methylation-sensitive enzyme
Expressed genes are typically hypomethylated
PstI/MseI markers predominantly located in gene-rich areas
Therefore, nonmethylated genic areas are found in more proximal regions of chromosome
Contrasts with RFLP mapping results in cereals

Unlocking variability: inherent variation and developmental traits of garlic plants originated
from sexual reproduction
Shemesh et al., 2008

Traits of garlic (Allium sativum)
Agricultural garlic completely sterile, propagated only from cloves
limits variation
improvement of economically important traits restricted
High variability in garlic from Central Asia, main center of origin
Growth habit and flowering significantly affected by
storage conditions
environment during the current and previous growing seasons

Broadening the variability of garlic
Collected over 300 garlic landraces and wild populations across Central Asia (local genotypes possess economically useful traits lost during 10,000 years of domestication)
Over 30 accessions produced flowers and set seeds
Detailed characterization of seedlings’ development and assessment of inherent variation
Availability of seed allows the study of the complete life cycle, free of plant history (storage, growth temperatures of previous years affect vegetatively propagated plant development)

NBS profiling reveals polymorphism in seedling population
Extracted DNA from 28 seedlings and mother plant
10 fragments from mother were polymorphic in progeny
All but 2 had 1-8 foreign DNA fragments originating from cross-pollination
2 have same fragment pattern as mother, presumed to be selfings

Ontogeny of seedlings:
Seed germination and juvenile state
Seed shape and color and seedling morphology typical to Allium
Germination lasted few weeks
Single or cluster bulbs formed at end of first growing season
Seedlings’ population varied in bulbing ability, bulbing time, and bulb traits (size, color, shape and clove number)

Ontogeny of seedlings:
Reproductive state
Transition from vegetative to reproductive phase occurred after 6-22 foliage leaves and leaf primordia are produced
Garlic genepool highly variable
No morphological and developmental differences evident between 1st and 2nd year of floral development
1st year seedlings can be used for study of blooming, avoiding field and storage history effects

Ontogeny of seedlings:
Florogenesis and topset development
Florogenesis same as vegetatively propagated garlic
Flower primordia initiation followed by vegetative meristem initiation in inflorescence, forms thin leaves or small dormant bulbs (topsets)
Varied in number and size of topsets/leaves under equal environmental conditions

Effect of storage temperature on ontogenesis of seedlings: Vegetative traits
No effect on sprouting time, leaf growth, or time from planting to first occurrence of secondary growth
Bulbing, secondary sprouting of axillary buds strongly affected
Despite genetic variability, lower temperature produced bulbs after ~15 leaves, higher temperatures caused rotting prior to clove formation except one plant developed bulb after ~22 leaves

Effect of storage temperature on ontogenesis of seedlings: Reproductive
traits
Transition to reproductive state greatly affected
Cold treatment required for flowering

Phenotypic variability under equal growth conditions
In third year of development, populations segregated into 3 main phenotypes: branching (20%), multifloral (60%), regular (20%)

ConclusionsConfirmed the essential role of cross-pollination
Cross-pollinated plants highly heterozygous which is preferred genotypic state for seedling survival and plant vigor
Seed populations showed large variation in all morphological and physiological traits, similar variability as vegetatively propagated garlic
Variability by restored sexual reproduction may serve as rich source for genetic studies and breeding work, development of new and improved cultivars propagated from seed