the ,journal of biological chemistry vol 259, no. 23 ... · the ,journal of biological chemistry fc...
TRANSCRIPT

THE ,JOURNAL OF BIOLOGICAL CHEMISTRY fc 1984 hy The American Society of Biological Chemists, Inc.
Vol 259, No. 23, Issue of December 10, PP. 14880-14887,19&9 Printed in U. S. A.
Evidence for Two RNA Polymerase Activities in Euglena gracilis Chloroplasts*
(Received for publication, May 25, 1984)
Bruce M. Greenberg, Jonathon 0. Narita, Camille DeLuca-Flaherty, Wilhelm GruissemS, Keith A. Rushlowf, and Richard B. Hallickn From the Department of Chemistry, University of Colorado, Boulder, Colorado 80309
Two types of RNA polymerase activity were isolated from Euglena gracilis chloroplasts and compared. One polymerase is tightly bound to chloroplast DNA; this complex is called the transcriptionally active chromo- some (TAC) (Rushlow, K. E., Orozco, E. M., Jr., Lipper, C., and Hallick, R. B. (1980) J. Biol. Chern. 255,3786- 3792). The other activity is found in a soluble extract of Euglena chloroplasts. The soluble extract is depend- ent upon an exogenous DNA template for activity. The two activities can be isolated in two distinct subchlo- roplast fractions from a single chloroplast preparation. The soluble extract is selective for transcription of transfer RNA genes, whereas the TAC is selective for ribosomal RNA genes. TAC and the soluble extract respond differently to KC1 and M 8 + . The soluble ex- tract is sensitive to heparin, and TAC is resistant. The two activities have different temperature optima. Based on this evidence, we conclude that Euglena chlo- roplasts have at least two distinct RNA polymerase activities.
~________ ____ ~~ _ _ _ _ _ ~ ~
Chloroplasts contain autonomous chromosomes that are in general approximately 150 kilobase pairs in size (for reviews see Refs. 1-3). Chloroplast genomes from several organisms have been well characterized. rRNA, mRNA, and tRNA genes have all been located on chloroplast DNA restriction maps. Many of these genes have been sequenced. Several in uitro chloroplast transcription systems have also been described (4-18). Since detailed information about chloroplast gene structure and organization is available and since chloroplast RNA polymerase activities are readily detected, one can now begin to study the molecular aspects of chloroplast gene expression in oitro.
Euglena gracilis is an ideal system for the study of chloro- plast transcription. The chloroplast genome organization is well understood. rRNA, tRNA, and mRNA gene loci have been extensively characterized (3,19-25,40). In addition, two different in uitro transcription systems from Euglena chloro- plasts have been reported (3, 10-12, 33). One is an RNA polymerase activity that is tightly bound to chloroplast DNA
* This work was supported by Grants GM28463 and 21351 to R. B. H. from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Current address, Department of Botany, University of California, Berkeley, CA 94720.
3 Current address, Syngene Products and Research, Inc., P. 0. Box 2211, Ft. Collins, CO 80522.
fl To whom correspondence should be addressed.
~~
(10, 11). This preparation, termed the TAC,’ specifically transcribes rRNA genes. The other activity is found in a soluble extract of Euglena chloroplasts. We recently reported the soluble extract to be active in template-dependent tRNA biosynthesis (12) as well as several post-transcriptional tRNA maturation reactions. Transcription systems very similar to both TAC and the soluble extract have also been isolated from spinach chloroplasts (5-9, 13, 14).
In this report we compare the Euglena chloroplast TAC and the soluble extract RNA polymerase activities directly. The properties compared include subchloroplast location, gene selectivity, temperature activity profiles, effect of diva- lent cations and salt concentration, and drug resistance. The data are consistent with the proposal that there are at least two different RNA polymerase activities in Euglena chloro- plasts. One activity is involved in rRNA precursor biosyn- thesis and the other in tRNA gene expression.
EXPERIMENTAL PROCEDURES
Isolation of Euglena Chloroplasts-E. gracilis Klebs, Pringsheim strain Z cells were grown autotrophically to a density of 1.0 to 1.5 X lo6 cells/ml at 25 “C with continuous illumination as previously described (11). Cells were harvested by continuous flow centrifugation in a Szent-Gyorgi and Blum rotor. Chloroplasts were isolated by flotation on sucrose in the presence of EDTA (32). The EDTA serves to strip the chloroplast outer membrane of cytoplasmic proteins and nucleic acids. Our previous report (12) on isolation of a soluble transcription extract from Euglena was based on differential centrif- ugation in Percoll gradients for chloroplast purification. We have since found that chloroplasts isolated by sucrose flotation give more consistent results.
Isolation of TAC-TAC was isolated as described previously (33), except that the ammonium sulfate concentration in the Triton X- 100-containing buffer during gel filtration was 0.4 M.
Preparation of the Euglena Chloroplast-soluble Extract-This pro- cedure is based on a previously published method (12), with some minor modifications. A flow chart of this procedure is shown in Fig. 2. All steps were done at 0-4 “C, unless otherwise stated. Chloroplasts purified by flotation on sucrose (32) were resuspended in 3 volumes of 10 mM Tris-HC1 (pH 7.9), 1 mM EDTA, 5 mM dithiothreitol, frozen on dry ice, and thawed. Then 3 volumes of 50 mM Tris-HC1 (pH 7.91, 10 mM MgC12, 2 mM dithiothreitol, 25% (w/v) sucrose, 50% (v/v) glycerol were added followed by 1 volume of saturated (4.1 M) am- monium sulfate to a final concentration of 0.5 M. This suspension was stirred slowly for 20 min. The suspension was next centrifuged for 3 h at 50,000 rpm in a Spinco Ti 70 rotor, and the supernatant was recovered. (The TAC can be isolated from the resulting pellet.) The proteins were then precipitated with 0.31 g of ammonium sulfate/ ml of supernatant (final concentration of ammonium sulfate was 2.9 M). The precipitated proteins were collected by centrifugation at 25,000 rpm in a Spinco SW-27 rotor for 30 min. The protein pellet was resuspended in 100 ~1 of 20 mM HEPES.Cl (pH 7.9), 100 mM KCI, 12.5 mM MgC12, 0.1 mM EDTA, 2 mM dithiothreitol, 17% (v/v)
The abbreviations used are: TAC, transcriptionally active chro- mosome; HEPES, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid.
14880

RNA Polymerase Activities of Euglena gracilis Chloroplasts 14881
glycerol/ml of chloroplast. The solution was dialyzed against the same solution overnight with several changes of buffer. Insoluble material was removed by centrifugation, and the extract was stored a t -70 "C. The extract is stable at this temperature for at least 6 months without a detectable loss of activity.
Euglena Chloroptost DNA-Recombinant plasmids used as DNA templates for in uitro transcription are listed in Table I. Restriction maps for the Euglem chloroplast DNA templates are shown in Fig. 1.
In Vitro Transcription and RNA Analysis-RNA was transcribed from TAC as described by Rushlow et al. (11, 33). RNA synthesis was measured by incorporation of nucleotides into trichloroacetic acid-insoluble material (10) or by polyacrylamide gel electrophoresis of RNA transcripts. Gels contained 10% acrylamide (acrylamide/ bisacrvlamide, 191). 7 M urea, 90 mM Tris base, 90 mM boric acid. 0.5 mM EDTA (pH 8.3).
For transcription reactions involving the Euglena chloroplast sol- uble extract, a- reaction cocktail was made containing 500 p~ ATP, 500 FM CTP, 50 I . ~ M GTP, 50 PM UTP, 10 FCi each of [w3'P]GTP and [a-32P]UTP, 5 mM Tris.HC1 (pH 7.9), 3.25 mM MgClz, 2 mM dithiothreitol, 2% glycerol, and 100 pg/ml of DNA. The extract was diluted 1:l with 20 mM HEPES, pH 7.9, 20 mM KC1, 12.5 mM MgCl,, 0.1 mM EDTA, 2 mM dithiothreitol, 17% glycerol. 10 pl of 1:l diluted soluble extract were added to 10 pl of the reaction cocktail, and the reation was incubated for 60 rnin at 25 "C unless stated otherwise. Reactions were stopped by incubation with proteinase K/sodium dodecyl sulfate (12) for 15 min at 37 "C. The reaction mixtures were then extrated with phenol/chloroform/isoamyl alcohol (25:24:1, v/v/ v) and applied to 10% polyacrylamide, 7 M urea gels. Following electrophoresis, RNAs were detected by autoradiography. Incorpora- tion of nucleotides into tRNAs was quantitated by liquid scintillation counting of tRNA-sized products which were excised from the gels.
an elution buffer containing 0.5 M NaCl, 10 mM Tris.HC1 (pH 7.6), RNA needed for further analysis was extracted from gel slices with
5 mM EDTA, and 1% (v/v) phenol. In uitro RNA transcripts were hybridized to membrane filter blots of chloroplast DNA restriction fragments as previously described (11, 36). RNase TI fingerprints were performed as previously described (13, 14).
Rifampicin, heparin, and a-amanitin were used as previously de- scribed (12). The drugs were added to the reaction cocktail prior to addition of the soluble extract.
RESULTS
Different RNA Polymerase Activities in Two Distinct Sub- chloroplast Fractions-The two Euglena chloroplast RNA po-
lymerase activities described in this paper are isolated by procedures outlined in Fig. 2. The chloroplast-soluble extract is made by lysis of the chloroplasts in a low-salt buffer followed by extraction and precipitation of the stromal pro- teins with ammonium sulfate. This produces an RNA polym- erase activity in a soluble chloroplast protein preparation. This RNA polymerase loses activity if detergents are present during chloroplast lysis. TAC is isolated by detergent lysis of the chloroplasts followed by chromatographic fractionation via gel filtration (Fig. 2 and Ref. 33). The RNA polymerase of the TAC remains tightly bound to chloroplast DNA in the presence of 0.5 M ammonium sulfate. The TAC RNA polym- erase activity is insensitive to 1-2% Triton X-100 in the lysis buffer. Conditions have not been found to solubilize the Euglena TAC RNA polymerase activity from the endogenous chloroplast DNA template while maintaining its full activity. I t is also possible to isolate TAC from the 0.5 M ammonium sulfate pellet that is normally discarded from the soluble extract preparation (Fig. 2). This TAC activity cannot be distinguished from TAC isolated from intact chloroplasts either in enzymatic properties or in yield. Thus, it is possible to isolate both activities from the same chloroplasts.
Different R N A Transcription Products Are Produced by the TAC and the Soluble Extract-The RNA polymerase and processing activities in the soluble extract produce tRNAs when DNAs containing Euglena chloroplast tRNA genes are added as templates (12-14). The products of transcription in the soluble extract with added plasmid (pPG14) or total Euglena chloroplast DNA are shown in Fig. 3 (Panel Z, lunes B and C). If no DNA is added to the extract some background transcription is observed due to a slight contamination by endogenous chloroplast DNA with the tightly bound RNA polymerase (i.e. TAC activity) (Fig. 3, Panel I , lane A) . This background comprises 10% of the total transcriptional activ- ity when pPG14 is added as template (data not shown). Transcripts are also produced by the soluble extract RNA polymerase activity using the endogenous DNA as template.
TABLE I Plasmid DNAs used for in uitro transcription
Plasmid Vector Chloroolast DNA Genes Reference
pPG14
pEZC3OO
pPG76
pEZC2 pEZC800 pEZC514 pPG689 pPG50
pEZCl
pEZC734
pMB9
pBR325
pBR322
pBR322 m13mp8 pBR325 pMB9 pMB9
pBR322
pBR325
EcoRI-G"
EcoRI-Q
BamHI-Sal19
Hind111 21 A Sau3A-Hind111 fragment' EcoRI I EcoRI J' EcoRI P
HindIII 23
EcoRI N and a PuuI-EcoRI fragmentg
trnV-UAC, tmN-GUU, trnR-ACG, trnL-UAGb
trnT-UGU, trnG-GCC, trnM- CAU, trnS-GCU, trnZ-UUG
trnY-GUA, trnH-GUG, trnM- CAU, trnW-CCA, trnE-UUC, trnG-UCC
trnP-GAA, trnC-GCA, trn?-CAT trnP-GAA, trnC-GCA trnL-UAA, psbAd tRNA genes' 16 S rDNA, trnI-GAU, tmA-UCG,
23 S rDNA (3'-end), 5 S rDNA, 16
tufA,h tmL-UGA, trnR-ACG,
23 S rDNA (5"end)
S rDNA (5"end)'
trnN-GUU (3'-end)
20
22
21
40 40 40 19, 40 24
25
35
All chloroplast DNAs endonuclease fragments.
are from the E. gracilis chloroplast genome. They are designated as restriction
tRNA genes-are designated by tm followed by the single letter code indicating the amino acid. Gene
This Sau3A-Hind111 fragment is a 600-base pair subclone from the HindIII 21 insert of pEZC2. psbA is the locus coding for the 32-kilodalton protein from photosystem 11. EcoRI I also contains an unassigned
nomenclature is described in Ref. 34.
reading frame. e These tRNA genes have not been sequenced. 'This Euglena chloroplast DNA fragment also contains the spacer region between rRNA operon repeats.
This PuuI-EcoRI fragment is a 1100-base pair fragment from EcoRI G. tufA is the locus coding for elongation factor Tu.

14882 RNA Polymerase Activities of Euglena gracilis Chloroplasts
Pffi 14 (EcoRI-HindP Fragment of ECO G 20kbp)
q@
pEZC300 (EcoQ,2.0kbp) .
PPG 76 (Barn-Sal 9,3.2 kbp 1 iF
(t.“ + 9 BamHT EcoRE
PEZC2 (Hind21,2.8kbp)
$? I J *
trnx trnF trnC _.
I rn -b ECORI trnL psbA
pPG689 (EcoJ’, 4.0kbp) +tRNA Gene Loci-1 &
fl9
Intact
Detergent Lysis *-\ Lysis
Low Salt
DNA With Bound RNA-Polymerase ..p-nand
(TAC)
Centrifugation
Precipitate
Concentrated TAC
Soluble Extract
FIG. 2. Flow chart for preparation of the Euglena chloro- plast TAC and soluble extract. The details for these preparations are described under “Experimental Procedures” (see also Refs. 12 and 33). All steps are performed at 0-4 “C. Note, the membrane pellet from the soluble extract preparation can be used as starting material for TAC preparation.
PPGSO (EcoP, 2.6kbp) 232 rDNA trpA tC“I 16s rDNA g*
4
c\\\\\~\\\\\\\\\\\\\\\\i <u“ “
EcoRI
Hindm
pEZC734 (4.0kbp) “mRNA” trnR trnN
I EcoRI Hindm trnL
EcoN From EcoG
”. PVUI
/ L f i V V
(2.9kbp) ( l lkbp)
FIG. 1. Physical maps of plasmid DNAs used for in vitro transcription. The Euglena chloroplast DNA inserts from the plas- mid DNAs are illustrated. Details for these plasmid DNAs are given in Table I. The boxes on the maps indicate the site of the genes. The names of the genes are given above the gene loci (see Ref. 34 for nomenclature). The direction of transcription is indicated by the arrows. All of Eco G (8.2 kilobase pairs (kbp)) is present in pPG14, but only the EcoRI-Hind111 (2.0 kilobase pairs) fragment is shown on the map. The sizes of the inserts are in kilobase pairs. These maps are not all drawn to the same scale.
Southern hybridization was used to investigate whether transcription in the soluble extract of DNA containing Eu- glena chloroplast tRNA genes as exogenous templates led to the production of tRNAs. 32P-labeled RNA from the tRNA region of the gel was eluted and hybridized to restriction endonuclease digests of Euglena chloroplast DNA (Fig. 3,
Panel ZZ). When no DNA was added to the extract the transcribed RNA hybridized to restriction fragments that correspond to the rRNA genes (Fig. 3, Panel ZZ, lune A). These are the Euglena chloroplast EcoRI restriction fragments B, F, L, and P (23, 24). This hybridization pattern is identical to the pattern one sees from RNA produced by TAC (11). Therefore, we conclude that most of the background tran- scription in the soluble extract is contributed by TAC. The RNA produced by transcription of pPG14 DNA in the soluble extract hybridizes to restriction endonuclease fragments of Euglena chloroplast DNA that contain tRNA operons (see Fig. 3, Panel 11, lane B, and Ref. 19). The strongest hybridi- zation signal is to the EcoRI G fragment. This is the Euglena chloroplast DNA fragment that is contained in pPG14 (see Fig. 1 and Table I) . The hybridization to EcoRI J’ and H and the enhanced hybridization to EcoRI P are due to tRNA genes with very high homology to the genes in EcoRI G (39, 40). When total chloroplast DNA is added to the soluble extract, the RNA product hybridizes to all known tRNA gene loci with about equal intensity (Fig. 3, Panel II, lane C, and Refs. 19 and 40). Hybridization to the rRNA operons is essentially unchanged by the addition of pPG14 or chloroplast DNA.
All tRNA Operons, but not rRNA Operons, Are Transcribed in the Soluble Extract-Plasmid DNAs containing various regions of the Euglena chloroplast genome were assayed as templates for transcription in the soluble extract (Fig. 1). pPG14, pEZC300, pPG76, pEZC800, pEZC2, pEZC514, and pPG689 all contain tRNA operons outside of the rRNA oper- ons (19-22,40). pPG50 and pEZCl contain parts of the rRNA operons (24,25). pEZC734 contains tufA and two tRNA genes from the EcoRI fragment G (35).
With all seven plasmid DNA templates containing Euglena chloroplast tRNA genes that are outside the rRNA operons the soluble extract produces tRNA-sized products (Fig. 4,

RNA Polymerase Activities of Euglena gracilis Chloroplasts 14883
I H
85. 75
A / \ \
A B C ABC silly
A 8 C 1 2 1 2 1 2 1 2
FIG. 3. Southern hybridization analysis of the products of in vitro transcription. Panel I, polyacryl- amide gel electrophoresis of in vitro transcription products. Transcription was done in the soluble extract under standard conditions. RNA products were detected by autoradiography. The right part of panel I is an enlargement of the left part. The transcript sizes are in nucleotides as estimated from known standards. Lane A, no added DNA; Lane E, pPG14; Lane C, chloroplast DNA, Panel 11, hybridization of the RNA products from in vitro transcription to restriction endonuclease fragments of Euglena chloroplast DNA. Chloroplast DNA was digested and electropho- resed in agarose. The DNA was then transferred to nitrocellulose filters. RNA used as probes was eluted from the gel shown in panel I . DNA fragment letters refer to the EcoRI fragments of chloroplast DNA (see Ref. 3). Lane 1, Euglena chloroplast DNA digested with EcoRI; Lane 2, Euglena chloroplast DNA digested with HindIII. Lane A, RNA probe from Lane A in panel Lane B, RNA probe from Lane E in panel I; Lane C, RNA probe from Lane C in panel I .
lunes 2-8). When no DNA is added, no tRNAs are produced (Fig. 4, lune I ) . The tRNA-sized products derived from tran- scription reactions involving pPG14, pEZC300, pEZC514, pPG76, and pEZC2 were further resolved by two-dimensional electrophoresis. Each of the resulting 19 individual tRNAs was characterized by RNase TI fingerprint analysis. In addi- tion, 15 individual tRNAs were further characterized by sec- ondary RNase A analysis of the RNase TI oligonucleotides. In each case, the tRNAs were found to be correctly transcribed and processed in the soluble extract. A fingerprint of tRNAL*" transcribed from EcoRI G (pPG14) is shown as an example (Fig. 5). This fingerprint is identical to a fingerprint of the same tRNA in a previous publication (see Fig. 4C in Ref. 13), in which tRNAku was shown to be correctly transcribed and processed at the 5'- and 3'-ends by the spinach chloroplast- soluble extract.
When the trnI-GAU and trnA-UCG genes' from the 16 S rRNA-23 S rRNA spacer region (pPG50) were used as a template in the soluble extract, no tRNAs were produced (Fig. 4, lane 9). When the 5'-leader region of the 16 S rRNA gene was added (pPG5O or pEZC1) no transcription above back- ground was observed (Fig. 4, lanes 9 and 10). This includes tRNA-sized transcripts and high-molecular-weight RNAs, ~ ~~~~
Gene nomenclature is described in Ref. 34. ~ ~~ ~~~ ~
such as are seen for TAC reactions (Fig. 4, lune 12). Note also that the 5 S rRNA gene is apparently not transcribed in the soluble extract (Fig. 4, lune IO). This would be a transcript of approximately 120 nucleotides. Therefore, the soluble extract will recognize tRNA genes outside of the three Euglena chlo- roplast rRNA operons but will not respond to any of the genes from the rRNA operons. It has already been shown that transcripts from the rRNA operons are selectively produced by the TAC (11). Therefore, the TAC RNA polymerase activ- ity and the soluble extract RNA polymerase activity appar- ently respond to different gene loci, i.e. rRNA genes and tRNA genes, respectively, to yield the complementary mixture of rRNAs and tRNAs required for chloroplast protein synthe- sis.
An interesting result was obtained when pEZC734 DNA was used as template in the soluble extract. This plasmid contains intact trnL-UAG, trnR-ACG, and a partial trnN- GUU (Fig. 1). However, only the 85-nucleotide tRNAk" tran- script is obtained (Fig. 4, compare lunes 2 and II). The fact that tRNAArK is apparently not transcribed is consistent with the prediction that the trnR locus is part of a trnV-trnN-trnR polycistronic transcription unit. Sequences 5' upstream from trnV, not present in pEZC734, are likely required for trnR expression (see "Discussion").
The TAC and the Soluble Extract RNA Polymerase Activi-
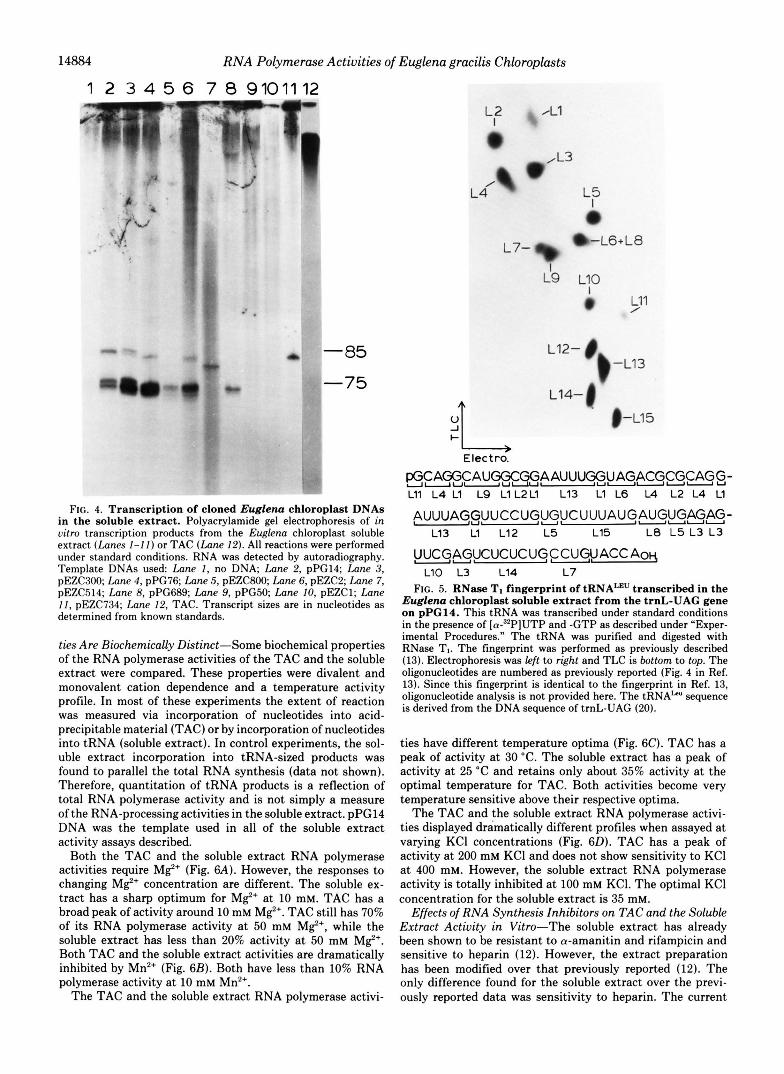
14884 R N A Polymerase Activities of Euglena gracilis Chloroplasts
1 2 3 4 5 6 7 8 9101112 L2 /L1
I
-85
-75
FIG. 4. Transcription of cloned Euglena chloroplast DNAs in the soluble extract. Polyacrylamide gel electrophoresis of in oitro transcription products from the Euglena chloroplast soluble extract (Lanes 1-1 1) or TAC (Lane 12). All reactions were performed under standard conditions. RNA was detected by autoradiography. Template DNAs used: Lane 1, no DNA; Lane 2, pPC14; Lane 3, pEZC3OO; Lane 4, pPG76; Lane 5, pEZC800; Lane 6, pEZCZ; Lane 7, pEZC514; Lane 8, pPG689; Lane 9, pPG50; Lane 10, pEZC1; Lane 1 1 , pEZC734; Lane 12, TAC. Transcript sizes are in nucleotides as determined from known standards.
ties Are Biochemically Distinct-Some biochemical properties of the RNA polymerase activities of the TAC and the soluble extract were compared. These properties were divalent and monovalent cation dependence and a temperature activity profile. In most of these experiments the extent of reaction was measured via incorporation of nucleotides into acid- precipitable material (TAC) or by incorporation of nucleotides into tRNA (soluble extract). In control experiments, the sol- uble extract incorporation into tRNA-sized products was found to parallel the total RNA synthesis (data not shown). Therefore, quantitation of tRNA products is a reflection of total RNA polymerase activity and is not simply a measure of the RNA-processing activities in the soluble extract. pPG14 DNA was the template used in all of the soluble extract activity assays described.
Both the TAC and the soluble extract RNA polymerase activities require Mg2+ (Fig. 6A). However, the responses to changing M e concentration are different. The soluble ex- tract has a sharp optimum for Mg2+ at 10 mM. TAC has a broad peak of activity around 10 mM M e . TAC still has 70% of its RNA polymerase activity at 50 mM M$+, while the soluble extract has less than 20% activity at 50 mM M e . Both TAC and the soluble extract activities are dramatically inhibited by Mn2+ (Fig. 6B). Both have less than 10% RNA polymerase activity at 10 mM Mn2+.
The TAC and the soluble extract RNA polymerase activi-
L I- Electro.
L7- * a-L6+L8
L9 L10 I
L12- 0 )-L13
L14- 1 ' #-L15
PGCAGGCAUGGCGGAAUUUGGUAGfiCqFGCAGG- Ll1 L4 L1 L9 L1 L2L1 L13 L1 L6 L4 L2 L4 Ll 4-u-uuu-u-
AUUUAGSUUCCUGUG.JCUUUAUGAUGUGAGAG- L13 L1 L12 L5 L15 L8 L5 L3 L3
L10 L3 L14 L7 FIG. 5. RNase TI fingerprint of tRNALEU transcribed in the
Euglena chloroplast soluble extract from the trnL-UAG gene on pPG14. This tRNA was transcribed under standard conditions in the presence of [c~-~'P]UTP and -GTP as described under "Exper- imental Procedures." The tRNA was purified and digested with RNase TI. The fingerprint was performed as previously described (13). Electrophoresis was left to right and TLC is bottom to top. The oligonucleotides are numbered as previously reported (Fig. 4 in Ref. 13). Since this fingerprint is identical to the fingerprint in Ref. 13, oligonucleotide analysis is not provided here. The tRNAL" sequence is derived from the DNA sequence of trnL-UAG (20).
ties have different temperature optima (Fig. 6C). TAC has a peak of activity at 30 "C. The soluble extract has a peak of activity at 25 "C and retains only about 35% activity at the optimal temperature for TAC. Both activities become very temperature sensitive above their respective optima.
The TAC and the soluble extract RNA polymerase activi- ties displayed dramatically different profiles when assayed at varying KC1 concentrations (Fig. 6D). TAC has a peak of activity a t 200 mM KC1 and does not show sensitivity to KC1 a t 400 mM. However, the soluble extract RNA polymerase activity is totally inhibited a t 100 mM KCI. The optimal KC1 concentration for the soluble extract is 35 mM.
Effects of RNA Synthesis Inhibitors on TAC and the Soluble Extract Actiuity in Vitro-The soluble extract has already been shown to be resistant to a-amanitin and rifampicin and sensitive to heparin (12). However, the extract preparation has been modified over that previously reported (12). The only difference found for the soluble extract over the previ- ously reported data was sensitivity to heparin. The current

RNA Polymerase Activities of Euglena gracilis Chloroplasts
Temperature PC 1
C
7
50
14885
B
[KCI] (mM) FIG. 6. Physical properties of the Euglena chloroplast soluble extract and the TAC RNA polymerase
activities. The soluble extract activity was measured by incorporation of [32P]UMP into the tRNA product. TAC RNA polymerase activity was measured by incorporation of tritiated nucleotides into acid-insoluble material. For all soluble extract assays, pPG14 DNA was used as template. Panel A, effect of [MgC12] on transcription. Panel B, effect of [MnS04] on transcription. Panel C, temperature activity profile. Panel D, effect of [KC11 on transcription. All reactions were done under standard conditions except the parameter of interest was varied. A, TAC; 0, soluble extract (SOL EX). Optimal incorporation was always in the range of 2 to 10 pmol.
extract is sensitive to 20 pg/ml heparin (data not shown). The extract made by Gruissem et al. (12) was sensitive to 2 pg/ml heparin. TAC is insensitive to rifampicin, a-amanitin, and heparin (10).
DISCUSSION
We have presented evidence for two types of RNA polym- erase activity in E. gracilis chloroplasts. TAC contains an RNA polymerase that is tightly bound to chloroplast DNA (10, 33) and specifically produces rRNA transcripts (11). A soluble extract of E. gracilis chloroplasts has been found to contain a different type of polymerase activity (12) that requires exogenous DNA templates and transcribes tRNA gene loci.
These two activities are isolated by different procedures. However, TAC can also be isolated in normal yields (based on chloroplast volume) from the chloroplast thylakoid mem- brane pellet that is generated during the soluble extract preparation. The soluble extract RNA polymerase activity remains in the stromal supernatant. Therefore, both activities can be recovered in different subchloroplast fractions from the same chloroplasts.
The two chloroplast RNA polymerase activities differ in their gene selectivity. In eukaryotic nuclei there are three classes of RNA polymerases (28, 29). RNA polymerase I specifically transcribes rRNAs, RNA polymerase I1 specifi- cally transcribes mRNAs, and RNA polymerase I11 specifi- cally transcribes tRNAs, 5 S rRNAs, and other small RNAs. A similar selectivity is observed for the two Euglena chloro-
plast RNA polymerase activities. TAC is specific for tran- scription of the rRNA operons, including trnA and trnI in these operons, even though the entire Euglena chloroplast genome is present (11). The soluble extract RNA polymerase activity transcribes all known Euglena chloroplast tRNA gene loci, except for trnA and trnI which are in the rRNA operons. Also, the soluble extract activity apparently does not recognize the rRNA genes.
We have not yet determined if chloroplast mRNAs can be transcribed in the soluble extract. If mRNAs are being tran- scribed in the soluble extract, it will be interesting to see if there is a third chloroplast RNA polymerase activity that is specific for mRNA genes or if one of the two activities described in this paper is responsible for mRNA transcription.
The fact that tRNAA1" and tRNA"' from the rRNA operons are not produced in the extract (pPG50) lends support to the notion that the soluble extract requires a specific chloroplast promoter sequence to initiate transcription. If transcription by this activity was random, one would expect tRNAA1" and tRNA"' to be produced from nonselective transcripts due to the accurate processing reactions in the extract. Indeed, when TAC RNA is purified and reincubated with the soluble ex- tract, tRNA-sized transcripts are obtained (data not shown). Thus the rRNA spacer tRNAs are capable of being processed and, therefore, must not be transcribed by the soluble extract. For eukaryotic nuclear in vitro tRNA transcription systems, sequences within the tRNA genes are utilized as promoters (37, 38). If this were the case in the Euglena chloroplast- soluble extract, one would again expect trnI and trnA to be expressed, since internal promoter sequences are conserved

14886 RNA Polymerase Activities of Euglena gracilis Chloroplasts
in most tRNAs, and these tRNAs have high sequence homol- ogy to the transcribed species (40). For example, trnA and trnG of pPG14 are 78% homologous, and the first 26 nucleo- tides are exactly identical (20). Additional support for the conclusion that sequences 5‘ external to tRNA genes are required as promoters is based on the transcription data for pEZC734. This plasmid is missing the 5’ half of a trimeric tRNA gene locus (Fig. l), but it still contains an intact tRNAA”” gene. tRNAA”” is not produced in the soluble extract from pEZC734, while as an internal positive control, an isolated tRNAL”” on the opposite DNA strand is correctly expressed (Fig. 4, lune 11). Therefore, it appears the soluble extract RNA polymerase activity has a requirement for a 5’ external promoter sequence, although other factors may also be important. This is consistent with our previous report based on Bat31 deletion analysis that the analogous RNA polymerase activity in spinach chloroplasts requires a 5’ external promoter (13). We are currently conducting experi- ments to further test this hypothesis and better define the Euglena chloroplast tRNA promoter.
Several biochemical properties of the TAC and the soluble extract RNA polymerase activities were compared. The only properties the two activities have in common are manganese sensitivity and rifampicin and a-amanitin resistance. By con- trast, the TAC and the soluble extract RNA polymerase activities differ in their responses to temperature, M$+ con- centration, KC1 concentration, and heparin. The soluble ex- tract and the TAC activities have similar MS’ concentration optima. They do differ significantly in response to higher M$+ concentrations. The effect of KC1 on the two activities is quite striking. The TAC activity is insensitive to high KC1 concentrations, whereas the soluble extract shows a very sharp response at low concentrations. The two activities also have different sensitivities to heparin. The TAC activity is resistant to heparin, while the soluble extract is quite sensitive to heparin. These results should be interpreted with some caution, since the TAC is a highly purified preparation, while the soluble extract is an impure stromal protein preparation. Therefore, some of the enzymatic properties reported could be influenced by other components in the extract. Neverthe- less some of the enzymatic properties are so distinct ( i x . KC1 response) that these polymerase activities can be readily distinguished in vitro.
The data presented in this paper can be interpreted in one of two ways. First, these two RNA polymerase activities might use the same core RNA polymerase, i.e. they could have common polypeptide subunits. If this is the case, then the two activities could simply differ in the factors used to transcribe different genes. This is reminiscent of the way Escherichia coli RNA polymerase is altered when E. coli is infected with phage T4 (41). That is, a protein factor encoded on T4 DNA changes the specificity of E. coli RNA polymerase from E. coli promoters to phage promoters. However, the use of different factors by the two chloroplast RNA polymerase activities to find different promoters would not explain the differences observed in DNA binding and biochemical properties. Alter- natively, chloroplasts could have two distinct RNA polymer- ases. This would account for the differing gene selectivity, DNA binding properties, and biochemical properties. There- fore, we find the latter interpretation to be more attractive. However, until both Euglena chloroplast RNA polymerases are purified, it will be impossible to determine whether these two activities have different core structures.
I t would appear from our data that there are at least two different Euglena chloroplast RNA polymerase activities. Two similar activities have been described in spinach chloroplasts
and other chloroplasts (4-9, 13-18). Therefore, these two types of activities might be common to all chloroplasts. This means that chloroplast transcription systems are probably not strictly analogous to prokaryotes, which only contain one class of RNA polymerase (26, 27). Chloroplast transcription systems are probably not analogous to yeast mitochondrial systems either, since only one type of RNA polymerase has been isolated from yeast mitochondria to date (30, 31). How- ever, chloroplast transcription systems may be analogous to eukaryotic nuclear transcription systems (26, 27). It could be that chloroplasts employ three types of RNA polymerase activities: one for rRNA transcription, one for tRNA tran- scription, and one for mRNA transcription. Alternatively, the two activities described in the present report might be suffi- cient to transcribe all chloroplast genes.
Acknowledgments-We wish to acknowledge the assistance of Drs. Thomas R. Cech, Peter Gegenheimer, and Marvin R. Paule in criti- cally reading the manuscript.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15. 16.
17.
18.
19.
20.
21.
22.
23.
24.
REFERENCES Whitfeld, P. R., and Bottomley, W. (1983) Annu. Reu. Plant
Bohnert, H. J., Crouse, E. J., and Schmitt, J. M. (1982) in Encyclopedia of Plant Physiology (Parthier, B., and Boulter, D., eds) Vol. 14B, pp. 475-530, Springer-Verlag, New York
Hallick, R. B., Greenberg, B. M., Gruissem, W., Hollingsworth, M. J., Karabin, G. D., Narita, J. O., Nickoloff, J. A., Passavant, C. W., and Stiegler, G. L. (1983) in Structure and Function of Plant Genomes (N. A. T. 0. Advanced Studies Institute) (Ci- ferri, O., and Dure, L., 111, eds) pp. 155-166, Plenum Press, New York
Wollgiehn, R., and Parthier, B. (1982) in Encyclopedia of Plant Physiology (Parthier, B., and Boulter, D., eds) Vol. 14B, pp. 125-170, Springer-Verlag, New York
Briat. J. F.. Laulhere, J. P., and Mache, R. (1979) Eur. J . Biochem.
PhYSiOl. 34 , 279-310
98; 285-292
509
. .
Briat, J. F., and Mache, R. (1980) Eur. J . Biochem. 111, 503-
Blanc, M., Briat, J. F., and Laulhere, J. P. (1981) Biochim.
Briat, J. F., Droix, M., Loiseaux, S., and Mache, R. (1982) Nucleic
Lerbs, S., Briat, J. F., and Mache, R. (1983) Plant Mol. Biol. 2,
Hallick, R. B., Lipper, C., Richards, 0. D., and Rutter, W. J.
Rushlow, K. E., Orozco, E. M., Jr., Lipper, C., and Hallick, R. B.
Gruissem, W., Narita, J. O., Greenberg, B. M., Prescott, D. M.,
Gruissem, W., Greenberg, B. M., Zurawski, G., Prescott, D. M.,
Greenberg, B. M., Gruissem, W., and Hallick, R. B. (1984) Plant
Tewari, K. K., and Goel, A. (1983) Biochemistry 2 2 , 2142-2148 Smith, H. J., and Bogorad, L. (1974) Proc. Natl. Acud. Sci. U. S. A.
Jollv. S. 0.. and Boeorad, L. (1980) Proc. Natl. Acud. Sci. U. S. A.
Biophys. Acta 655 , 374-382
Acids Res. 10,6865-6878
67-74
(1976) B i o ~ h ~ m i s t ~ 15, 3039-3045
(1980) J. Biol. Chem. 255 , 3786-3792
and Hallick, R. B. (1983) J. Cell. Biochem. 22,31-46
and Hallick, R. B. (1983) Cell 35,815-828
Mol. Bioi. 3 , 97-109
71,4839-4842 I .
77,822-826 Schwarz. Z.. Jollv. S. 0.. Steinmetz. A. A.. and Bogorad. L. (1981)
Proc. Natl. Acid. Sci.’U. S. A. 76, 3423-3427 - Orozco. E. M.. Jr.. and Hallick, R. B. (1982) J. Biol. Chem. 2 5 7 ,
325813264 I ,
Orozco, E. M., Jr., and Hallick, R. B. (1982) J. Bwl. Chem. 2 5 7 ,
Hollingsworth, M. J., and Hallick, R. B. (1982) J. Biol. Chem.
Karabin, G . D., and Hallick, R. B. (1983) J. Biol. Chem. 258 ,
3265-3275
257 , 12795-12799
5512-5518 Orozco, E. M., Jr., Gray, P. W., and Hallick, R. B. (1980) J. Bid.
Orozco, E. M., Jr., Rushlow, K. R., Dodd, J. R., and Hallick, R. Chem. 255, 10991-10996

RNA Polymerase Activities of Euglena gracilis Chloroplasts 14887
B. (1980) J. Biol. Chern. 2 5 5 , 10997-11003 Biomedical Press, Amsterdam 25. Karabin, G. D., Narita, J. O., Dodd, J. R., and Hallick, R. B. 33. Rushlow, K. E., and Hallick, R. B. (1982) in Methods in Chloro-
(1983) J. Biol. Chern. 2 5 9 , 14790-14796 p h t Molecular Biology (Edelman, M., Hallick, R. B., and Chua, 26. Chamberlin, M. J. (1976) in RNA Polymerase (Losick, R., and N.-H., eds) pp. 543-550, Elsevier/North-Holland Biomedical
Chamberlin, M., eds) pp. 17-67, Cold Spring Harbor Press, Press, Amsterdam Cold Spring Harbor, NY 34. Hallick, R. B., and Bottomley, W. (1983) Plant Mol. Biol. Reporter
ed., Vol. 15B, pp. 61-86, Academic Press, New York 35. Passavant, C. W., Stiegler, G. L., and Hallick, R. B. (1983) J.
berlin, M., eds) pp. 285-329, Cold Spring Harbor Press, Cold 36. Southern, E. M. (1975) J. Mol. Biol. 98 , 503-517 Spring Harbor, NY 37. DeFranco, D., Schmidt, O., and So11, D. (1980) Proc. Natl. Acad.
29. Lewis, M. K., and Burgess, R. R. (1982) in The Enzymes (Boyer, Sci. U. S. A. 77, 3365-3368 P. D., ed) 3rd ed., Vol. 15B, pp. 109-153, Academic Press, New 38. Galli, G., Hofstetter, H., and Birnstiel, M. L. (1981) Nature York (Lond.) 294,626-631
30. Levens, D., Lustig, A., and Rabinowitz, M. (1981) J. Biol. Chem. 39. Nickoloff, J. A., and Hallick, R. B. (1982) Nucleic Acids Res. 10,
31. Edwards, J. C., Levens, D., and Rabinowitz, M. (1982) Cell 31 , 40. Hallick, R. B., Hollingsworth, M. J., and Nickoloff, J. A. (1984)
32. Hallick, R. B., Richards, 0. C., and Gray, P. W. (1982) in Methods 41. Losick, R., and Pero, J. (1976) in RNA Polymerase (Losick, R., in Chloropht Molecular Biology (Edelman, M., Hallick, R. B., and Chamberlin, M., eds) pp. 227-246, Cold Spring Harbor and Chua, N.-H., eds) pp. 281-294, Elsevier/North-Holland Press, Cold Spring Harbor, NY
27. Chamberlin, M. J. (1982) in The Enzymes (Boyer, P. D., ed) 3rd 18,179-183
28. Roeder, R. G. (1976) in RNA Polymerase (Losick, R., and Cham- Biol. Chem. 258,693-695
256,1474-1481 8191-8210
337-346 Plant Mol. Bid . 3, 169-175