the n-hexane and chloroform fractions of piper betle l. trigger different arms of immune responses...
TRANSCRIPT

International Immunopharmacology 9 (2009) 716–728
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r.com/ locate / in t imp
The n-hexane and chloroform fractions of Piper betle L. trigger different arms ofimmune responses in BALB/c mice and exhibit antifilarial activity against humanlymphatic filarid Brugia malayi
Meghna Singh a, Shilpy Shakya a, Vishal Kumar Soni a, Anil Dangi a,Nikhil Kumar b, Shailja-Misra Bhattacharya a,⁎a Division of Parasitology, Central Drug Research Institute, P. Box 173, Lucknow-226001, Indiab Betel Vine Biotechnology Laboratory, National Botanical Research Institute, Rana Pratap Marg, Lucknow-226001, India
⁎ Corresponding author. Tel.:+91522 261241118 (Off),2623938/2623405.
E-mail addresses: [email protected], shailj(S.-M. Bhattacharya).
1567-5769/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.intimp.2009.02.012
a b s t r a c t
a r t i c l e i n f oArticle history:Received 12 June 2008Received in revised form 1 February 2009Accepted 18 February 2009
Keywords:Piper betleImmunomodulationAntibodyTh1/Th2 cytokinesFlow cytometryAntifilarial activityBrugia malayi
Modulation of immune functions by using herbal plants and their products has become fundamental regime oftherapeutic approach. Piper betle Linn. (Piperaceae) is awidely distributed plant in the tropical and subtropicalregions of the world and has been attributed as traditional herbal remedy for many diseases. We have recentlyreported the antifilarial and antileishmanial efficacy in the leaf extract of Bangla Mahoba landrace of P. betlewhich is a female plant. The present report describes the in vivo immunomodulatory efficacy of the crudemethanolic extract and its n-hexane, chloroform, n-butanol fractions of the female plant at various dose levelsranging between 0.3 and 500 mg/kg in BALB/c. Attempts were also made to observe antifilarial activity of theactive extracts and correlate it with the antigen specific immune responses in another rodentMastomys couchainfected with human lymphatic filarial parasite Brugia malayi. The crude methanol extract and n-hexanefraction were found to potentiate significant (pb0.001) enhancement of both humoral (plaque forming cells,hemagglutination titre) as well as cell-mediated (lymphoproliferation, macrophage activation, delayed typehypersensitivity) immune responses in mice. The flow cytometric analysis of splenocytes of treated miceindicated enhanced population of T-cells (CD4+, CD8+) and B-cells (CD19+). The n-hexane fraction (3mg/kg)was found to induce biased type 2 cytokine response as revealed by increased IL-4+ and decreased IFN-γ+ T-cell population while the chloroform fraction (10 mg/kg) produced a predominant type 1 cytokines. Crudemethanolic extract (100mg/kg) demonstrated amixed type 1 and type 2 cytokine responses thus suggesting aremarkable immunomodulatory property in this plant. The induction of differential T-helper cell immuneresponse appears ideal to overcome immunosuppression as observed in case of lymphatic, filarial Brugiamalayi infection which may also be extended to other infections as well.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Traditional systems of medicine provide an extremely vast body ofsource material for the development of new drugs and naturalproducts have become increasingly important as sources of pharma-cotherapeutics [1,2]. A global concern about infectious diseasesgenerates a need to identify novel means for their effective treatment.Immunomodulation using medicinal plants is seriously being con-sidered as an alternative approach to conventional chemotherapy fortreatment of these diseases. The concept of ‘Rasayana’ in traditionalIndian health system Ayurveda is also based on related principles ofbio enhancer [3], enhancing the body's defencemechanism. Betel vine(Piper betle L., family Piperaceae) is an important and ancient crop of
PABX 4221/4224; fax:+91522
ll rights reserved.
India and has been ascribed many medicinal properties [4,5]. Theplant is indigenous throughout the Indian Malaya region and used inMadagascar, Bourbon and the West Indies. Leaves of betel vine havebeen used with condiments such as areca nut, kattha, cloves,cardamom, fennel and candied rose for chewing purposes and nearlyone billion people consume it daily in one form or the other globally[6]. The betel leaves act as breath-freshener and are used in the Indiansystem of medicine and health and include medicinal properties suchas digestive, antacid, decongestant, carminative, stimulant, anti-pyretic, anti-inflammatory, anti-allergic, anti-septic, hepatoprotective,radio-protective, anti-fungal, anti-platelet, anti-fertility, nematocidaland anti-oxidant [7–16]. The chief constituent of the leaves is a volatileoil which contains phenols, betel-phenol; chavibetol and chavicol;cadinene and hydroxychavicol, which have been ascribed to possessanti-oxidant and anti-carcinogenic activities [8,17]. This crop isclaimed to have hundreds of landraces broadly grouped into threetypes, Bangla, Kapoori and others [18,19]. The plant is known to haveexplicit dioecy and the male and female plants can be easily identified

717M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
[13]. We have earlier reported the antiparasitic activity of the P. betleleaf extracts in vitro [20]; however, there is no definite report on itsimmunomodulatory and in vivo antiparasitic activity so far.
In the present study, the crude methanol extract from the leaves ofBangla landrace and its fractions viz. n-hexane, chloroform and n-butanol, were evaluated for their immunomodulatory activity inBALB/c mice employing both humoral and cellular immune assaysand an attempt was made to explore the molecular mechanism ofimmunomodulation. Further, in vivo antifilarial activity of the crudemethanol extract and its active fractions was assessed using humanlymphatic filariid, B. malayi experimentally maintained in the rodenthostMastomys coucha and the antifilarial results were correlated withthe immunomodulation potential of the plant.
2. Materials and methods
2.1. Plan of work
The crude methanol extract (MeOH) was administered to BALB/cmice orally at 500, 100, 30 mg/kg×14 days while fractions were fed at100, 30, 10 and 3 mg/kg orally×14 days. Of these, few animal groupswere immunized with SRBC (Sheep red blood corpuscles) on day 9post onset of treatment. At the end of treatment immunized animalswere euthanized humanely to determine specific antibody (HA titreand PFC assay) and cellular immune response (DTH) and unim-munized mice were used for assaying cellular proliferation (LTT) andmacrophage function (NO production). Once the immunomodulatoryactivity was established by these 4 parameters, active extract/fractions were further explored to dissect the sub-clones of T cells(Th1/Th2) and cytokine production using flow cytometric analysis.Apart from immunomodulatory activity, the active extract andfractions were also exploited for their antiparasitic (antifilarial)activity against human lymphatic filarial parasite Brugia malayi inrodent model (Mastomys coucha) and the immune response of thetreated filarial host was also studied. The results of immunomodula-tory efficacy of extracts were corroborated with the antifilarialactivity.
Scheme 1. Fractionation
2.1.1. MaterialsMedium RPMI-1640 (Sigma, USA) containing Phenol Red was used
in all the studies except colorimetric Nitric Oxide assay wheremedium devoid of Phenol Red was used. The medium was fortifiedwith 1% antibiotic-antimycotic cocktail (Sigma, USA) and 10% Foetalbovine serum (GIBCO, USA) for in vitro cell culture. Concanavalin A(Con A), lipopolysaccharide (LPS), HEPES and 2-mercaptoethanolwere purchased from Sigma Chemical Co. (St. Louis, MO). Allantibodies were purchased from Becton and Dickinson (BD) (SanDiego, CA, USA). All other chemicals used were of analytical gradeavailable locally.
2.1.2. Preparation of plant extractP. betle landraces were grown in the botanical garden of National
Botanical Research Institute, Lucknow under fully protected cultiva-tion and fully-grown mature leaves were harvested, washed, weighedand loaded in Clevenger apparatus for preparation of decoction [20].The crude methanolic extract was prepared by using 1.5 kg of shadedried leaf powder and extracted with 5 l of methanol until there wasno green colour in the extract. The yield of methanol extract was 73 gof which 70 g was dissolved in minimal amount of water andpartitioned with n-hexane. The yield of n-hexane fraction was about23 g. The water layer was further partitioned with chloroform whichyielded fraction weighing 9 g. Further partitioning was done with n-butanol which yielded 5 g extract and the residual 31 g was taken asaqueous fraction. The percent yield of different fractions was 32, 13,7.5 and 44 respectively for n-hexane, chloroform, n-butanol andaqueous. Methanolic extract (MeOH) and its fractions were dried invacuum and stored in vacuuo under cold condition. The crude MeOHextract and n-hexane, n-butanol and chloroform were selected forassaying the immunomodulatory activity as some of these extractsdemonstrated antiparasitic activity in vitro in our earlier study [20](Scheme 1).
2.1.3. Animals and their treatment with P. betle extract and its fractionsAge and sex matched (18–20 g) inbred mice strain BALB/c were
used for the treatment with different extracts and fractions. The
scheme of P. betle.

718 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
animals were housed under standard conditions of temperature (23±1 °C), relative humidity (55±10%), 12 h/12 h light/dark cycles atNational Laboratory Animal Centre (NLAC), CDRI, Lucknow, India andfed with standard pellet diet and water ad libitum. All the animalhandling and experimental protocols employed in the present studywere duly approved by the Institutional Animal Ethics Committee(IAEC). The crude methanolic extract and its fractions were preparedas suspension in distilled water along with 1% gum acacia as a vehicle.Animals were grouped (each having five) and fed with thesepreparations orally using canula daily for 14 consecutive days.MeOH preparation was administered at 500, 100 and 30 mg/kg dosewhile fractions were fed at lower doses of 100, 30, 10, 3, 1 and 0.3 mg/kg. Control mice received the vehicle (1% gum acacia in distilledwater) under identical conditions using the canula. Each experimentwas repeated with the same number of mice.
2.2. Assessment of humoral immune response
2.2.1. Haemagglutinating titre (HA Titre)Sheep red blood cells (SRBC) were aseptically collected from a
normal healthy sheep in the Alsever's solution and stored at 4 °C.Groups of treated and untreated (n=5) mice were intraperitoneallyimmunised with 1×107 (SRBC, 0.1 ml) on day 9 since the start oftreatment and autopsied on day 15 for sera collection (HA titer) andPFC counts. Serum antibody levels were determined by the Haemag-glutination technique [21,22], in U-bottom plate (NUNC) and theendpoint agglutination of SRBC was considered as positive titre.
2.2.2. Plaque forming cell (PFC) assayThe above mice were autopsied and spleens were aseptically
removed to make single cell splenocytes suspension for PFC assay[23], in agarose petridishes. The plaques were counted and the valuesexpressed as counts per 1×106 cells.
2.3. Assessment of cellular immune response
2.3.1. Delayed type hypersensitivity reaction (DTH) by foot padmeasurementMicewere primedwith 2×108 SRBC suspensions in right-hind foot
pad, PBS in the contra lateral paw and the thickness of footpad wasmeasured at 24, 48 and 72 h post SRBC challenge [24], using aPlethsymometer.
2.3.2. Lymphoproliferative responseSplenic T and B-lymphocyte proliferative response of treated and
untreated mice was determined in triplicate wells of a 96 well flatbottom plates (NUNC) after stimulation with an optimal concentra-tion (2.5 μg/ml) of Concavaline-A (Con-A) (Sigma, USA) andLipopolysaccharide (LPS) (Sigma, USA) at 37 °C for 48 h in 5% CO2
in air. Cells were harvested after 16 h of pulsing with [3H]-thymidine(0.5 μCi/well) and radioactive incorporation was measured on β-counter (Backman coulter) to evaluate stimulation indices [25].
2.4. Estimation of nitric oxide (NO) and Reactive oxygen species (ROS)using peritoneal macrophages
The peritoneal macrophages cells were collected from treated andcontrol mice, plated (3×105cells/well) into flat bottom 96-wellculture plate (NUNC) and macrophages were allowed to adhere toplastic bottom for 2 h at 37 °C in 5% CO2 in air [26]. A set of triplicatewells was stimulated overnight with 0.1 μg/well LPS and culturesupernatant was collected for estimating the NO production by freshlyprepared Griess Reagent by reading absorbance at 550 nm usingNaNO2 as standard [27]. A standard curve was plotted using sodiumnitrate (Sigma, USA) to quantitate the test results and the concentra-tion of nitric oxide is expressed in nM NO/1×106 cells. Intracellularreactive oxygen species (ROS) in peritoneal macrophage cells was
determined through a fluorometric assay using 2′, 7′-dichlorofluor-escin diacetate (DCF-DA) (Sigma, USA) in flow cytometry as describedearlier [28]. Briefly, freshly harvested macrophage cells of both treatedand untreated groups were adjusted to a concentration of 5×106
cells/ml in phosphate buffer saline (PBS), washed with PBS×3 andtransferred to FACS tubes (1×106 cells/tube). For probe loading, cellswere incubated with the DCF-DA at a final concentration of 1 μM, for15 min at 37 °C and thenwashed twice in PBS. ROS levels in individualliving cells were determined by individually measuring theirfluorescence intensity on FACS Calibur (Becton Dickinson, USA).Data were analyzed by CellQuest Software (Becton Dickinson, USA)and mean ROS values were evaluated for cell populations.
2.5. Flow cytometric (FACS) measurement of CD3+/CD4+ and CD3+/CD8+
T-lymphocyte sub-population
Immunophenotyping to assess lymphocyte subset population byflow cytometry was performed on a Becton Dickinson (BD) FACSCalibur using fluorochrome conjugated monoclonal antibodies (Ser-otec) directed against co-receptors to CD3+ (FITC) and CD4+/CD8+/CD19+ (phycoerythrin or PE) antigens following the manufacturer'sprotocol. Splenocytes suspension was prepared from spleens oftreated and untreated control mice on day 15 of the commencementof treatment. 1×106 cells were initially blocked with Mouse SeroblockFcR at room temperature (RT) for 10 min, washed and labeled initiallywith rat anti-mouse FITC-CD3+ for 10min at RT for further incubationwith rat anti-mouse PE-CD4+ or CD8+ for another 10 min. The thirdtube served as CD3+ control. Cell pellet was suspended in sheath fluidand analyzed by FACS using CellQuest analysis software (Becton-Dickinson, San Diego, CA) after gating the forward- and side-scattersettings to exclude debris. For each determination 20,000 cells wereanalyzed and the results are reported as percentage of each cellpopulation.
2.6. Intracellular cytokines by flow cytometry
Themeasurement of intracellular cytokines in the spleenwas doneas per manufacturer's instructions using antibodies and reagents fromBD. Briefly, splenocytes (4×106/ml) were incubated with Brefeldin A(10 μg/ml) (Serotech) in dark for 6 h in CO2 incubator at 37 °C andreincubated with mouse Seroblock FcR at RT for another 10 min. Cellswere washed in PBS and incubated with FITC-rat anti mouse CD3+
antibody. Leucoperm A and Leucoperm B (Serotech) was added at RTfor 15 min and cells were dispensed in four tubes each containing1×106 cells/100 μl. PE-rat antimouse monoclonal antibodies tocytokines IL-4 and IFNγ were added to separate tubes, cells werewashed and finally suspended in 250 μl of 0.5% Para formaldehyde forFACS readings.
2.7. Evaluation of antiparasitic activity of P. betle extracts against lymphaticfilarial parasite B. malayi in vivo in the rodent host Mastomys coucha
2.7.1. Experimental animal and treatmentSub periodic strain of B. malayi was transmitted to rodent host
Mastomys coucha by subcutaneous inoculation of 100 infectivelarvae (L3) of B. malayi recovered from mosquito vector Aedesaegypti after day 9±1 of infective blood meal on microfilaraemicmastomys [29,30]. Mastomys carrying 5–6 month old B. malayiinfection and displaying progressive rise in microfilaraemia wereselected and divided into ten groups (n=5). The crude MeOHextract and its fractions were fed by oral route daily for fiveconsecutive days. Crude methanol extract was administered at logdoses of 300, 100 and 30 mg/kg while the fractions wereadministered at 100 and 30 mg/kg. Infected control group receivedthe vehicle only. Five uninfected and untreated normal mastomysserved as another control.
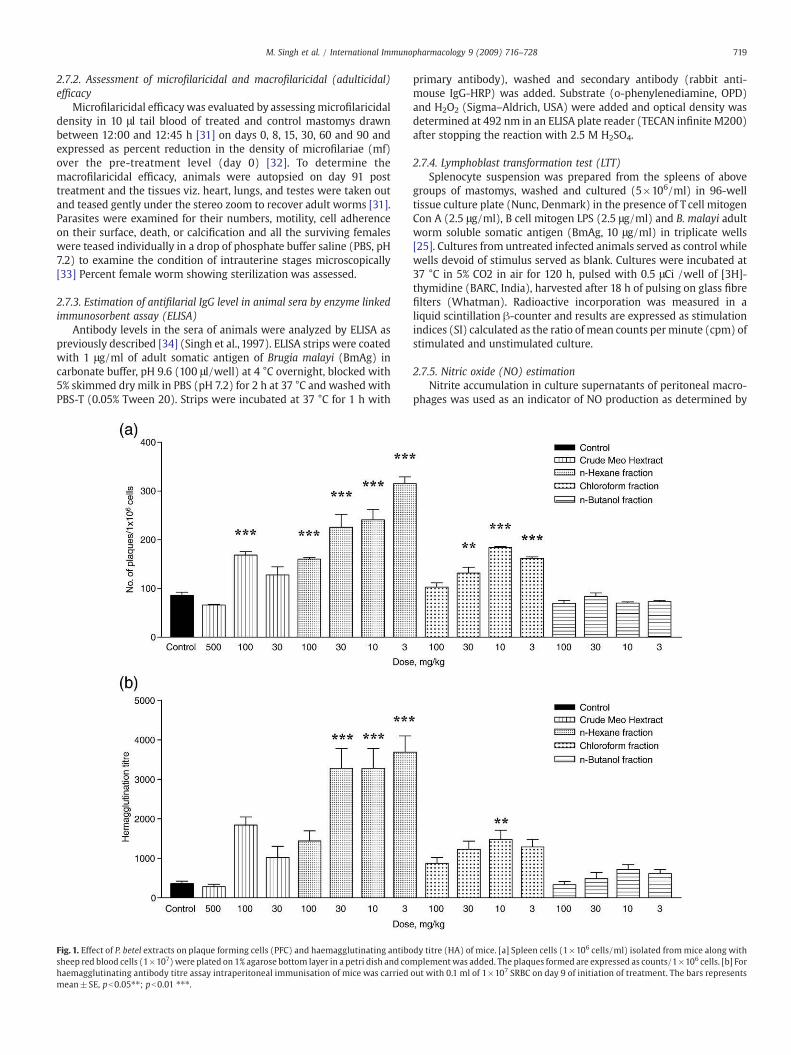
719M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
2.7.2. Assessment of microfilaricidal and macrofilaricidal (adulticidal)efficacy
Microfilaricidal efficacy was evaluated by assessingmicrofilaricidaldensity in 10 μl tail blood of treated and control mastomys drawnbetween 12:00 and 12:45 h [31] on days 0, 8, 15, 30, 60 and 90 andexpressed as percent reduction in the density of microfilariae (mf)over the pre-treatment level (day 0) [32]. To determine themacrofilaricidal efficacy, animals were autopsied on day 91 posttreatment and the tissues viz. heart, lungs, and testes were taken outand teased gently under the stereo zoom to recover adult worms [31].Parasites were examined for their numbers, motility, cell adherenceon their surface, death, or calcification and all the surviving femaleswere teased individually in a drop of phosphate buffer saline (PBS, pH7.2) to examine the condition of intrauterine stages microscopically[33] Percent female worm showing sterilization was assessed.
2.7.3. Estimation of antifilarial IgG level in animal sera by enzyme linkedimmunosorbent assay (ELISA)
Antibody levels in the sera of animals were analyzed by ELISA aspreviously described [34] (Singh et al., 1997). ELISA strips were coatedwith 1 μg/ml of adult somatic antigen of Brugia malayi (BmAg) incarbonate buffer, pH 9.6 (100 μl/well) at 4 °C overnight, blocked with5% skimmed dry milk in PBS (pH 7.2) for 2 h at 37 °C and washed withPBS-T (0.05% Tween 20). Strips were incubated at 37 °C for 1 h with
Fig. 1. Effect of P. betel extracts on plaque forming cells (PFC) and haemagglutinating antibosheep red blood cells (1×107) were plated on 1% agarose bottom layer in a petri dish and cohaemagglutinating antibody titre assay intraperitoneal immunisation of mice was carriedmean±SE, pb0.05⁎⁎; pb0.01 ⁎⁎⁎.
primary antibody), washed and secondary antibody (rabbit anti-mouse IgG-HRP) was added. Substrate (o-phenylenediamine, OPD)and H2O2 (Sigma–Aldrich, USA) were added and optical density wasdetermined at 492 nm in an ELISA plate reader (TECAN infinite M200)after stopping the reaction with 2.5 M H2SO4.
2.7.4. Lymphoblast transformation test (LTT)Splenocyte suspension was prepared from the spleens of above
groups of mastomys, washed and cultured (5×106/ml) in 96-welltissue culture plate (Nunc, Denmark) in the presence of T cell mitogenCon A (2.5 μg/ml), B cell mitogen LPS (2.5 μg/ml) and B. malayi adultworm soluble somatic antigen (BmAg, 10 μg/ml) in triplicate wells[25]. Cultures from untreated infected animals served as control whilewells devoid of stimulus served as blank. Cultures were incubated at37 °C in 5% CO2 in air for 120 h, pulsed with 0.5 μCi /well of [3H]-thymidine (BARC, India), harvested after 18 h of pulsing on glass fibrefilters (Whatman). Radioactive incorporation was measured in aliquid scintillation β-counter and results are expressed as stimulationindices (SI) calculated as the ratio of mean counts perminute (cpm) ofstimulated and unstimulated culture.
2.7.5. Nitric oxide (NO) estimationNitrite accumulation in culture supernatants of peritoneal macro-
phages was used as an indicator of NO production as determined by
dy titre (HA) of mice. [a] Spleen cells (1×106 cells/ml) isolated frommice along withmplement was added. The plaques formed are expressed as counts/1×106 cells. [b] Forout with 0.1 ml of 1×107 SRBC on day 9 of initiation of treatment. The bars represents

Fig. 2. Effect of P. betel extracts on delayed type hypersensitivity (DTH) reaction in SRBC immunized mice. SRBC immunized (on day 9) mice were challenged on day 15 with 20 μl of2×108 SRBC suspension in the right-hind foot pad and left foot pad received PBS. Difference in both the foot pad swelling was measured at 24 h, 48 h and 72 h. The figure shows thedata collected at 48 h revealing maximal swelling. Bar represents mean±SE, pb0.05⁎⁎; pb0.01⁎⁎⁎.
Fig. 3. Effect of P. betel extracts on T and B lymphocyte proliferation in vitro. T-/B-cell mitogens, Con-A [a] and LPS [b] were added at an optimal concentration of 2.5 μg/ml and theresults are expressed as stimulation indices (S.I.). Bar represents mean±SE, pb0.05⁎⁎; pb0.01⁎⁎⁎.
720 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
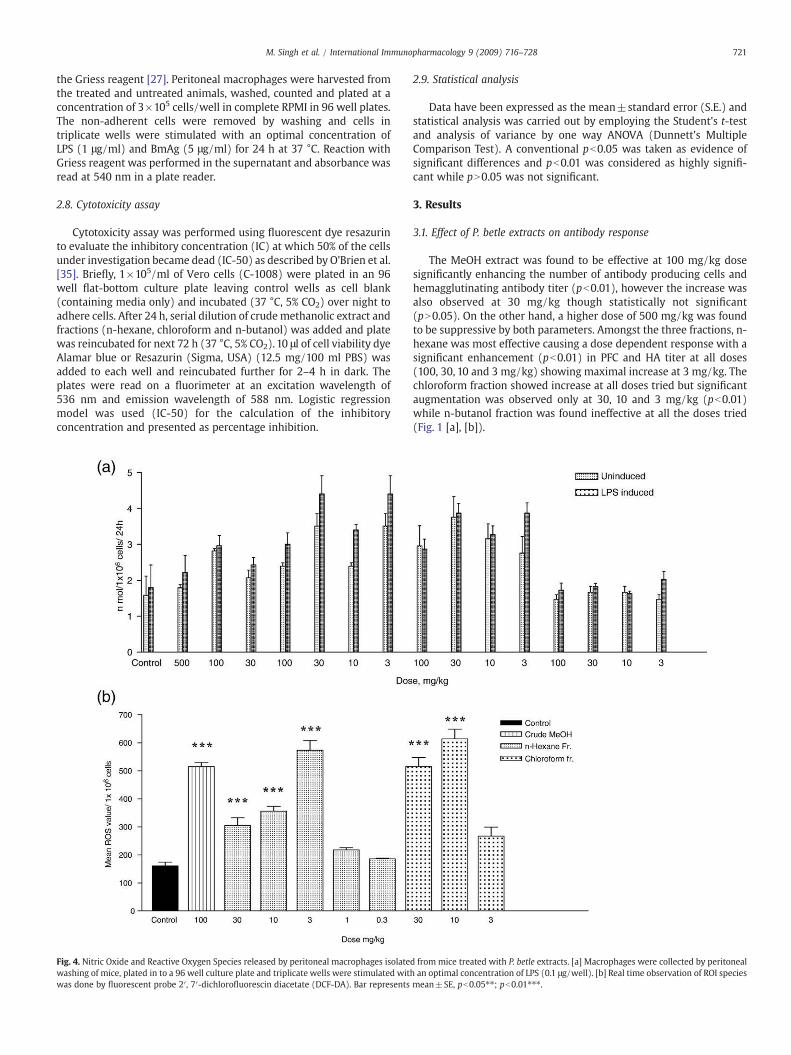
721M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
the Griess reagent [27]. Peritoneal macrophages were harvested fromthe treated and untreated animals, washed, counted and plated at aconcentration of 3×105 cells/well in complete RPMI in 96 well plates.The non-adherent cells were removed by washing and cells intriplicate wells were stimulated with an optimal concentration ofLPS (1 μg/ml) and BmAg (5 μg/ml) for 24 h at 37 °C. Reaction withGriess reagent was performed in the supernatant and absorbance wasread at 540 nm in a plate reader.
2.8. Cytotoxicity assay
Cytotoxicity assay was performed using fluorescent dye resazurinto evaluate the inhibitory concentration (IC) at which 50% of the cellsunder investigation became dead (IC-50) as described by O'Brien et al.[35]. Briefly, 1×105/ml of Vero cells (C-1008) were plated in an 96well flat-bottom culture plate leaving control wells as cell blank(containing media only) and incubated (37 °C, 5% CO2) over night toadhere cells. After 24 h, serial dilution of crudemethanolic extract andfractions (n-hexane, chloroform and n-butanol) was added and platewas reincubated for next 72 h (37 °C, 5% CO2). 10 μl of cell viability dyeAlamar blue or Resazurin (Sigma, USA) (12.5 mg/100 ml PBS) wasadded to each well and reincubated further for 2–4 h in dark. Theplates were read on a fluorimeter at an excitation wavelength of536 nm and emission wavelength of 588 nm. Logistic regressionmodel was used (IC-50) for the calculation of the inhibitoryconcentration and presented as percentage inhibition.
Fig. 4. Nitric Oxide and Reactive Oxygen Species released by peritoneal macrophages isolatewashing of mice, plated in to a 96 well culture plate and triplicate wells were stimulated witwas done by fluorescent probe 2′, 7′-dichlorofluorescin diacetate (DCF-DA). Bar represents
2.9. Statistical analysis
Data have been expressed as the mean±standard error (S.E.) andstatistical analysis was carried out by employing the Student's t-testand analysis of variance by one way ANOVA (Dunnett's MultipleComparison Test). A conventional pb0.05 was taken as evidence ofsignificant differences and pb0.01 was considered as highly signifi-cant while pN0.05 was not significant.
3. Results
3.1. Effect of P. betle extracts on antibody response
The MeOH extract was found to be effective at 100 mg/kg dosesignificantly enhancing the number of antibody producing cells andhemagglutinating antibody titer (pb0.01), however the increase wasalso observed at 30 mg/kg though statistically not significant(pN0.05). On the other hand, a higher dose of 500 mg/kg was foundto be suppressive by both parameters. Amongst the three fractions, n-hexane was most effective causing a dose dependent response with asignificant enhancement (pb0.01) in PFC and HA titer at all doses(100, 30, 10 and 3 mg/kg) showing maximal increase at 3 mg/kg. Thechloroform fraction showed increase at all doses tried but significantaugmentation was observed only at 30, 10 and 3 mg/kg (pb0.01)while n-butanol fraction was found ineffective at all the doses tried(Fig. 1 [a], [b]).
d from mice treated with P. betle extracts. [a] Macrophages were collected by peritonealh an optimal concentration of LPS (0.1 μg/well). [b] Real time observation of ROI speciesmean±SE, pb0.05⁎⁎; pb0.01⁎⁎⁎.

722 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
3.2. Effect of P. betle extract on cellular immune response
The hypersensitivity response characterized by foot pad swelling wasevident after 24, 48 and72hof injectionwith a suspensionof 2×108 SRBCinto the right hindpawand the readings at 48hwere found tobemaximal
Fig. 5. CD3+/CD4+, CD3+/CD8+ and CD19+ T- and B-cell populations in mice treated with[b] CD3+/CD8+ T cell population in mice fed with crude methanol, n-hexane and chloropb0.01⁎⁎⁎.
and thus included in the graph. The MeOH extract brought aboutsignificant increase in the foot pad swelling at 100 (pb0.01) and30mg/kg(pb0.05) (Fig. 2) but not at 500 mg/kg. N-hexane fraction was found todisplay increased cellular immune response which was significant at3mg/kg(pb0.05). Chloroformandn-butanol fractions alsobrought about
P. betle extracts by Flow cytometry. [a] CD3+ gated population of CD4+ T cell subsets,form fraction [c] The CD19+ B cell population. Bar represents mean±SE, pb0.05⁎⁎;
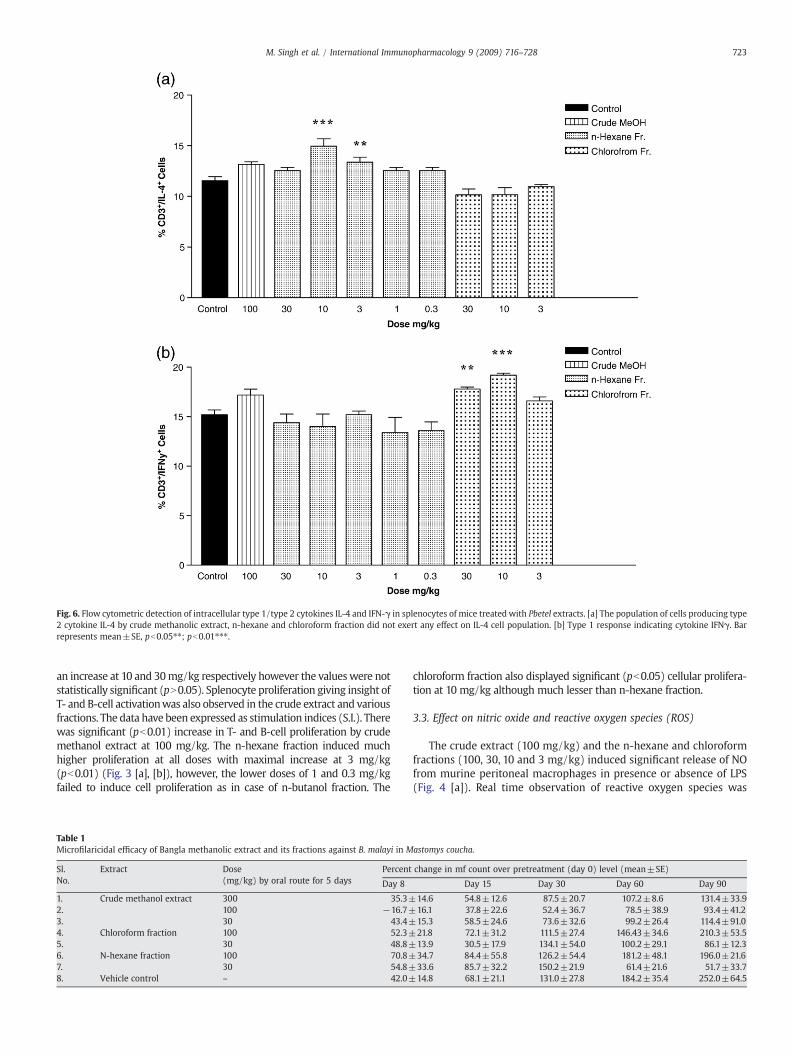
Fig. 6. Flow cytometric detection of intracellular type 1/type 2 cytokines IL-4 and IFN-γ in splenocytes of mice treated with Pbetel extracts. [a] The population of cells producing type2 cytokine IL-4 by crude methanolic extract, n-hexane and chloroform fraction did not exert any effect on IL-4 cell population. [b] Type 1 response indicating cytokine IFNγ. Barrepresents mean±SE, pb0.05⁎⁎; pb0.01⁎⁎⁎.
723M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
an increase at 10 and 30mg/kg respectively however the valueswere notstatistically significant (pN0.05). Splenocyte proliferation giving insight ofT- and B-cell activationwas also observed in the crude extract and variousfractions. The data have been expressed as stimulation indices (S.I.). Therewas significant (pb0.01) increase in T- and B-cell proliferation by crudemethanol extract at 100 mg/kg. The n-hexane fraction induced muchhigher proliferation at all doses with maximal increase at 3 mg/kg(pb0.01) (Fig. 3 [a], [b]), however, the lower doses of 1 and 0.3 mg/kgfailed to induce cell proliferation as in case of n-butanol fraction. The
Table 1Microfilaricidal efficacy of Bangla methanolic extract and its fractions against B. malayi in M
Sl.No.
Extract Dose(mg/kg) by oral route for 5 days
Percent
Day 8
1. Crude methanol extract 300 35.3±2. 100 −16.7±3. 30 43.4±4. Chloroform fraction 100 52.3±5. 30 48.8±6. N-hexane fraction 100 70.8±7. 30 54.8±8. Vehicle control – 42.0±
chloroform fraction also displayed significant (pb0.05) cellular prolifera-tion at 10 mg/kg although much lesser than n-hexane fraction.
3.3. Effect on nitric oxide and reactive oxygen species (ROS)
The crude extract (100 mg/kg) and the n-hexane and chloroformfractions (100, 30, 10 and 3 mg/kg) induced significant release of NOfrom murine peritoneal macrophages in presence or absence of LPS(Fig. 4 [a]). Real time observation of reactive oxygen species was
astomys coucha.
change in mf count over pretreatment (day 0) level (mean±SE)
Day 15 Day 30 Day 60 Day 90
14.6 54.8±12.6 87.5±20.7 107.2±8.6 131.4±33.916.1 37.8±22.6 52.4±36.7 78.5±38.9 93.4±41.215.3 58.5±24.6 73.6±32.6 99.2±26.4 114.4±91.021.8 72.1±31.2 111.5±27.4 146.43±34.6 210.3±53.513.9 30.5±17.9 134.1±54.0 100.2±29.1 86.1±12.334.7 84.4±55.8 126.2±54.4 181.2±48.1 196.0±21.633.6 85.7±32.2 150.2±21.9 61.4±21.6 51.7±33.714.8 68.1±21.1 131.0±27.8 184.2±35.4 252.0±64.5

Fig. 7. Worm recovery (white bars) and percentage of recovered female wormssterilized (gray bars) on day 90 after treatment with crude methanol extract,chloroform and n-hexane fractions of P. betle administered at different doses. Percentfemale worm sterilization was assessed over that of vehicle treated control animals. Barrepresents mean±SE, pb0.05⁎⁎; pb0.01⁎⁎⁎.
724 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
thereby carried out in cells of mice using fluorescent probe 2′, 7′-dichlorofluorescin diacetate (DCF-DA) only at the effective dosesevolved during above experimentations (crude extract-100 mg/kg, n-hexane and chloroform fractions- 30, 10, 3, 1 and 0.3 mg/kg). 100 mg/kg of crude extract and n-hexane fraction and 30,10 and 3mg/kg of n-hexane fraction led to an increase (pb0.01) in ROS content whilechloroform fraction showed a remarkable (pb0.01) increase at 30 and10 mg/kg dose. (Fig. 4 [b]).
3.4. Effect onB- and T-cell sub-populations and cytokines byflowcytometry
3.4.1. ImmunophenotypingThe methanol extract and its two fractions (n-hexane and
chloroform) caused up-regulation of CD3+/CD4+ population in thespleen with maximum efficacy shown by methanol extract and n-hexane fraction Fig. 5[a]. In contrast, CD3+/CD8+ cells were up-regulated after treatment with chloroform fraction while the crudeMeOH extract and n-hexane fraction caused its down-regulationexcept at 3 mg/kg dose of n-hexane fraction (Fig. 5[b]). The CD19+
cell population also increased in all extracts with a significant
Fig. 8. Filaria-specific IgG levels in the sera of mastomys belonging to uninfected untreated nfraction and n-hexane fraction at 100 mg/kg, orally for 5 days) on days 0, 15, 30, 60, and 90adult soluble somatic antigen. Bar represents mean±SE at 1:800 serum dilution. pb0.01⁎⁎
increase in n-hexane fraction (pb0.01) at 10 mg/kg concentration(Fig. 5[c]).
3.4.2. Intracellular cytokines by FACSThe population of CD3+ cells producing type 2 cytokine (IL-4)
increased after treatment with crude methanolic extract and n-hexane fraction (pb0.01), latter was statistically significant at 3 mg/kg while chloroform fraction did not show any such effect (Fig. 6[a]).On the other hand, the type 1 cytokine (IFNγ) was observed to beincreased significantly (pb0.05) by chloroform fraction (10 mg/kg)while crude methanolic extract led to marginal increase in IFNγ level(Fig. 6[b]).
3.5. Antifilarial activity of P. betle extracts against Brugia malayi in vivo
Based on the immunostimulatory findings, the antifilarial activityof crude methanol extract, chloroform and n-hexane fractions wasevaluated against human lymphatic filariid B. malayi in rodent hostMastomys coucha.
3.5.1. Effect on microfilaraemiaCrude MeOH extract led to suppression in microfilaraemia
(Table 1) as compared to the untreated control groups. This activitywas most noticeable at the dose of 100 mg/kg. Both the fractions at30 mg/kg dose led to suppressed microfilaraemia on day 90 ascompared to that seen in untreated controls (Table 1); however, thedifference in microfilarial density in case of n-hexane fraction wasstatistically significant (pb0.05) as compared to controls.
3.5.2. Adulticidal (macrofilaricidal) efficacyWhen the crude MeOH extract was given at 300 mg/kg, the
adulticidal activity was found low (25.9%). At a dose of 100 mg/kg,crude MeOH extract caused reduction in adult worm recovery whencompared to untreated control animals. At a dose of 30 mg/kg,significant reduction in adult worm recovery was exhibited by n-hexane (57.9%, pb0.05) while crude MeOH and chloroform fractionshowed moderate adulticidal activity (Fig. 7).
3.5.3. Female worm sterilizationSignificant difference was observed in percent of female worm
sterilized in the animal group treated with crude MeOH (pb0.01)when administered at a dose of 100 mg/kg. The treatment with
ormals, infected untreated and P. betle treated groups (crude MeOH extract, chloroformafter initiation of treatment. IgG titre was determined by indirect ELISA using B. malayi⁎; Analysis done by one way ANOVA, Dunnetts' Multiple comparison test.
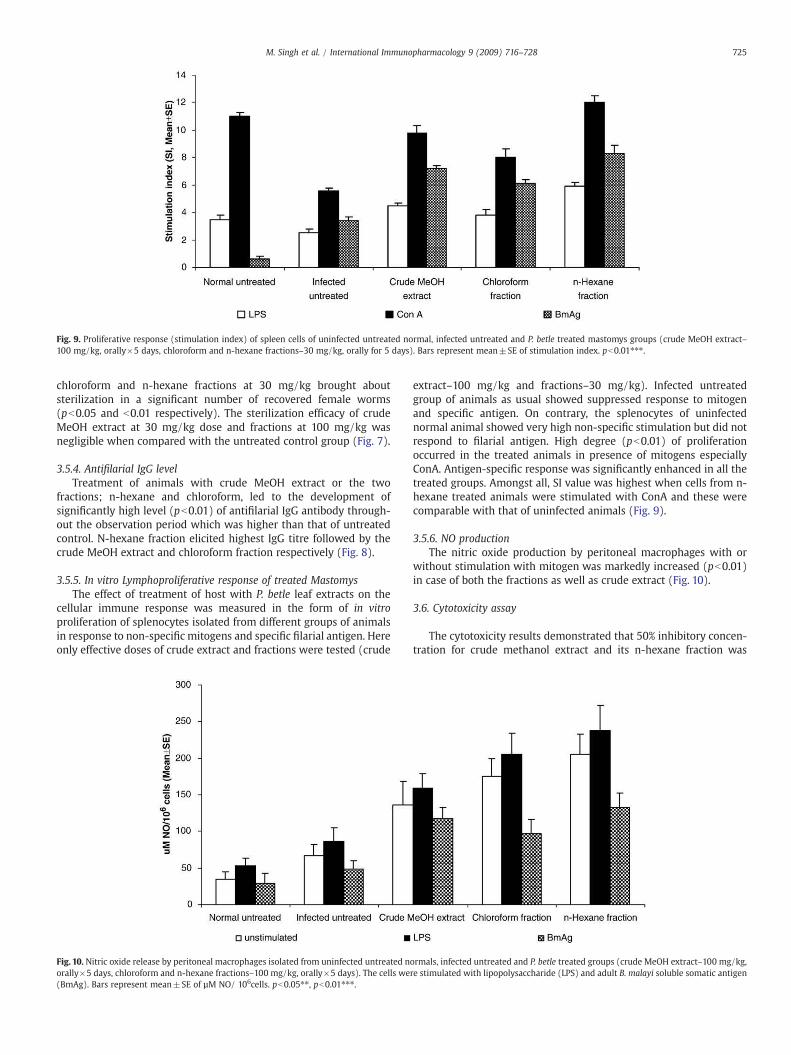
Fig. 9. Proliferative response (stimulation index) of spleen cells of uninfected untreated normal, infected untreated and P. betle treated mastomys groups (crude MeOH extract–100 mg/kg, orally×5 days, chloroform and n-hexane fractions–30 mg/kg, orally for 5 days). Bars represent mean±SE of stimulation index. pb0.01⁎⁎⁎.
725M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
chloroform and n-hexane fractions at 30 mg/kg brought aboutsterilization in a significant number of recovered female worms(pb0.05 and b0.01 respectively). The sterilization efficacy of crudeMeOH extract at 30 mg/kg dose and fractions at 100 mg/kg wasnegligible when compared with the untreated control group (Fig. 7).
3.5.4. Antifilarial IgG levelTreatment of animals with crude MeOH extract or the two
fractions; n-hexane and chloroform, led to the development ofsignificantly high level (pb0.01) of antifilarial IgG antibody through-out the observation period which was higher than that of untreatedcontrol. N-hexane fraction elicited highest IgG titre followed by thecrude MeOH extract and chloroform fraction respectively (Fig. 8).
3.5.5. In vitro Lymphoproliferative response of treated MastomysThe effect of treatment of host with P. betle leaf extracts on the
cellular immune response was measured in the form of in vitroproliferation of splenocytes isolated from different groups of animalsin response to non-specific mitogens and specific filarial antigen. Hereonly effective doses of crude extract and fractions were tested (crude
Fig. 10. Nitric oxide release by peritoneal macrophages isolated from uninfected untreated norally×5 days, chloroform and n-hexane fractions–100 mg/kg, orally×5 days). The cells we(BmAg). Bars represent mean±SE of µM NO/ 106cells. pb0.05⁎⁎, pb0.01⁎⁎⁎.
extract–100 mg/kg and fractions–30 mg/kg). Infected untreatedgroup of animals as usual showed suppressed response to mitogenand specific antigen. On contrary, the splenocytes of uninfectednormal animal showed very high non-specific stimulation but did notrespond to filarial antigen. High degree (pb0.01) of proliferationoccurred in the treated animals in presence of mitogens especiallyConA. Antigen-specific response was significantly enhanced in all thetreated groups. Amongst all, SI value was highest when cells from n-hexane treated animals were stimulated with ConA and these werecomparable with that of uninfected animals (Fig. 9).
3.5.6. NO productionThe nitric oxide production by peritoneal macrophages with or
without stimulation with mitogen was markedly increased (pb0.01)in case of both the fractions as well as crude extract (Fig. 10).
3.6. Cytotoxicity assay
The cytotoxicity results demonstrated that 50% inhibitory concen-tration for crude methanol extract and its n-hexane fraction was
ormals, infected untreated and P. betle treated groups (crude MeOH extract–100 mg/kg,re stimulated with lipopolysaccharide (LPS) and adult B. malayi soluble somatic antigen

726 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
184.24 μg/ml while chloroform and n-butanol fractions exhibited IC-50 of 73.85 and 233.73 μg/ml respectively showing very lowcytotoxicity.
4. Discussion
The concept of immunomodulation relates to non-specific activa-tion of the function and efficiency of macrophages, granulocytes,complement, natural killer cells and lymphocytes, and also to theproduction of various effector molecules generated by activated cells[36]. In the past, a number of drugs with plant or mineral origin havebeen advocated as means of immunomodulation for various diseaseconditions in humans [2,3,37,38]. The present findings are the firstever reported on immunomodulatory properties of P. betle extract andits fractions in BALB/c mice. The crude extract and the two fractionswere found to be highly efficient in augmenting the T- and B-cellimmune responses and enhanced macrophage function. In our earlierstudy, the leaf extract of widely consumed Bangla Mahoba (femaleplant) demonstrated antiparasitic activity in vitro [19]. The presentstudy reports the promising in vivo antifilarial efficacy in P. betleMeOH extract and its n-hexane and chloroform fractions in anexperimental model of lymphatic filariasis complementing theimmunomodulatory activity of plant leaves. Sincemost of the parasiticinfections including filariasis are associated with immunosuppression[39,40], use of immunostimulant like P. betle as immunoprophylacticand/or chemotherapy adjunct would help in controlling the infection.
The crude methanol extract and fractions (n-hexane and chloro-form) were found to stimulate the humoral immune response asdisplayed by an increase in both antibody level and the number ofantibody forming cells apart from accentuating the cellular immuneparameters. The release of nitric oxide in appropriate magnitudeserves as a key-signalling molecule in various physiological processesand has also been found to be protective against parasitic infection likefilariasis [26]. A significant rise in nitric oxide and reactive oxygenspecies content in the peritoneal macrophages of treated mice (crudemethanol, n-hexane and chloroform)was observed. The leaf extract ofP. betle has earlier been reported to scavenge free radicals involved inthe initiation and propagation steps of oxidative damage [5,10,11]; itshydroxychavicol and eugenol constituents are known to be non-mutagenic and anti-carcinogenic which has also been attributed to itsanti-oxidant property [7] while other molecules like piperbetol,piperol A and B have been shown to inhibit platelet aggregation [41].
The alcoholic extract of P. betle leaves is known to exert potentanti-leishmanial, anti-amoebic, anti-giardial, anti-microbial and lar-vicidal properties [42–46]. The ethanolic leaf extract of P. betle at100 mg kg−1 has earlier been shown to possess IL-12 inhibitoryproperty in rat, thus indicating the ability of leaf extract to down-regulate Th1 pro-inflammatory responses [47]. The murine bonemarrowand splenocyte cellular proliferation has also been found to betriggered by P. betle leaf extract [48]. The major active principleallylpyrocatechol (APC) isolated from P. betle leaves is found to targetthe inflammatory response of macrophages via inhibition of iNOS,COX-2 and IL-12 p40 via down regulation of the NF-κB pathway,indicating a possible therapeutic potential of APC in inflammationassociated disorders [49]. Also a pharmaceutical formulation consist-ing of lyophilized extracts of Piper betle and Murraya koeniggi is a Th2inhibitor suppressing the IL4 and leukotrine synthesis and thus usefulfor treatment of respiratory conditions in humans [50]. The ethanolleaf extract also significantly reduces histamine and GM-CSF, inhibi-tion of eotaxin and IL-8 secretion in TNF-α and IL-4-induced allergicreaction thus divulging the role of P. betle in control of allergic diseasesthrough inhibition of production of allergic mediators.
P. betle is being widely consumed amongst human populationworldwide with about 15–20 million people in India on a regular basis[49] besides those in other countries of the world which may includeover 2 billion consumers [51]. P. betel leaves are used in a number of
Indian traditional remedies as stomach ailments, infections, curing ofworms, and as aphrodisiac and anti-carcinogen [52]. It is also used as ageneral tonic, antibiotic and as a stimulant for chewing purpose incombination with the betel nut (Areca catechu) to treat bad breath orhalitosis which is attributed to the presence of allylpyrocatechol in theleaves [53]. In Malaysia, Philippines, Thailand, Indonesia and China,P. betle is used in the treatment of headaches, arthritis, toothache,constipation, congestion and to boost lactation. Leaves fight againstmouth bacteria Streptococcus viridans, S. aureus, S. mutans in humansand have potential application as natural food preservative [54]. Theessential oil found in the leaves possesses antiprotozoan and antifungalproperties apart from being effective against typhoid, cholera andtuberculosis, which needs proper evaluation and exploitation [4].
Theactivatedanddifferentiated lymphocytesplaya critical role againstextracellular pathogens besides being important in antigen presentationand activation of T-cells [55]. The present study established that the crudemethanol extract and its n-hexane and chloroform fractions significantlyenhanced both T- and B-lymphocyte proliferation inpresence ofmitogensat various doses while n-butanol fraction did not show any immunomo-dulation property. To further explore the molecular mechanism behindthe cellular activation by effective extracts, crude extract, n-hexane andchloroform fractions were fed at the most effective doses. The n-hexanefraction which showed activity at 3 mg/kg in initial experiment wasfurther evaluated at lowerdoses to eliminate thepossibility of ‘hook effect’and the maximum efficacy was still shown at 3 mg/kg. An up-regulatedCD3+/CD4+ cell population was observed in mice treated with MeOHextract, n-hexane or chloroform fractions, indicating the expansion of T-helper immune response while CD8+ cytotoxic cells in general showed adecreased trend except in chloroform fractionwheremarginal increase inCD3+/CD8+ counts was noticed. On the other hand, CD19 expressedprimarily on follicular dendritic cells and B-cells were found to beincreased only in case of n-hexane fraction. The activated T lymphocytesare a major source of released cytokines and on the basis of cytokinesecretory profile CD4+T-cells can be categorized as type 1 and type 2. Thepolarization of T-cells into the Th1/Th2 cells is accompanied by secretionof type 1 cytokines such as IL-2 and IFN-γ and type 2 such as IL-4, IL-5 andIL-10. The degree of polarisation of T-cell subsets depends on the strengthand duration of the polarising signals, and involves both cell-intrinsicfactors, as well as exogenous signals [56–58]. Administration of n-Hexanefraction in mice led to a predominant type 2 cytokine response whilechloroform fraction in contrast developed a biased type 1 response. Thecrudemethanol extract, however produced amixed response whichmaybeexplaineddue to thepresenceof thesediversely responding fractions inthe crude extract. The increase in IFN-γ positive T-cells clearly correlateswith the expansion of CD3+/CD8+ cytotoxic T-cells. There is a singlepatent report on stimulation of IFN-γ by human peripheral bloodmononuclear cells when treated with aqueous leaf extract of P. betleindicating Th1 type of immune response [59]. IFN- represents a keymolecule in human anti-parasitic host-defense. Deleterious inflammatoryresponses tometazoanparasites appear to be consistently associatedwitha highly polarized Th2 cytokine profile. The pleiotropic cytokine IL-4 isimportant for the differentiation of naive T helper precursors into Th2effector cells and is considered to be one of the most critical factorsdetermining immunoglobulin (Ig) class switching to IgG1 and IgE.
In the present report, the n-Hexane fraction of P. betlewas found toexert remarkable antifilarial activity as revealed by worm mortalityand high percentage of female worm sterilization. The oral adminis-tration of the P. betle extract and both the fractions (n-hexane andchloroform) at 100 mg/kg for 5 consecutive days in filarial animalmodel mastomys caused significant increase in NO productioncorrelating well with the reduced blood microfilarial density. Theextract and fractions also led to increased T- and B-cell proliferation inpresence of mitogens as well as filaria-specific antigen and increasedIgG antibody production. Moreover, crude methanol extract showingmixed type of immune response in BALB/c caused significant femaleworm sterilization and had moderate action on worm load. In case of

727M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
filarial infection Th2 response has been described to play a protectiverole while antigen-specific proliferative responses (Th1) are severelyimpaired [60]. In a filaria-endemic area, infection-free endemicnormal individuals show hyperimmune-responsiveness to filarialantigens while impaired cellular proliferation is well known duringactive infection as also observed in B. malayi infected mastomys in thepresent study and this unresponsiveness could be reversed bytreatment with crude methanolic extract, n-Hexane and chloroformfractions. This reversal brought about significant antifilarial activityapart from modulating the cell-proliferation and nitric oxide produc-tion. Thus, the immunomodulatory activity of Piper betle helped inovercoming the immunosuppression caused by filarial infection andthe findings may well be applied in case of other parasitic infections.
The immunostimulants usually display suppressive activity if thedose surpasses optimal level [61] as also observed in the presentfindings, and therefore selection of an ideal dose is the key to achievedesired immunostimulation. This is the first ever report on theimmunomodulatory properties of Piper betle leaves which skew theTh1/Th2 balance of immune system and thus may be furtherexploited against various other infections [62]. Though the extrapola-tion of animal data to a clinical setting is not always appropriate due tocertain inherent limitations, yet the interesting results obtained herewarrant further human studies to exploit this plant and extrapolateour results in view of wide human consumption of P. betle leaves.
Acknowledgements
The authors acknowledge University Grants Commission andCouncil of Scientific and Industrial Research (M.S., S.S. and A.D.),Indian Council of Medical Research (V.K.S), New Delhi, India forfinancial assistance in the form of Senior Research Fellowships. Thismanuscript bears CDRI communication no. 7315.
References
[1] Holland BK. Prospecting for drugs in ancient texts. Nature 1994;369:702–5.[2] Newman DJ, Cragg GM, Snader KM. The influence of natural products upon drug
discovery. Nat Prod Rep 2000;17:215–34.[3] Ganju L, Karan D, Chanda Srivastava KK, Sawhney RC, Selvamurthy W. Immunomo-
dulatory effects of agents of plant origin. Biomed Pharmacother 2003;57:296–300.[4] CSIR (Council of Scientific and Industrial Research, New Delhi). The Wealth of
India, vol. 8; 1969. p. 84–94.[5] Guha P, Jain RK. Status Report on Production, Processing and Marketing of Betel
Leaf (Piper betle L.). Kharagpur, India: Agricultural and Food EngineeringDepartment, IIT; 1997.
[6] Norton SA. Betel: consumption and consequences. J Am Acad Dermatol 1998;38:81–8.[7] NagabhushanM, Amonkar AJ, D'Souza AV, Bhide SV. Nonmutagenicity of betel leaf
and its antimutagenic action against environmental mutagens. Neoplasma1987;34:159–67.
[8] Garg SC, Jain R. Biological activity of the essential oil of Piper betle L. J Essent Oil Res1992;4:601–6.
[9] GangulyMS, Chattopadhyay S, Chatterjee S, Mitali. An ethanol extract of Piper betleLinn. mediates its anti-inflammatory activity via down-regulation of nitric oxide. JPharm Pharmacol 2007;59(5):711–8.
[10] Majumdar B, Chaudhuri SR, Roy A, Bandyopadhyay SK. Potent anti-ulcerogenicactivity of ethanol extract of leaf of Piper betle Linn. by anti-oxidative mechanism.Indian J Clin Biochem 2002;17:49–57.
[11] Bhattacharya S, Subramanian M, Roychowdhury S, Bauri AK, Kamat JP, Chatto-padhyay S, et al. Radioprotective property of the ethanolic extract of Piper betelleaf. J Radiat Res 2005;46:165–71.
[12] Guha P. Betel leaf: the neglected green gold of India. J Hum Ecol 2006;19(2):87–93.[13] Kumar N, Gupta S, Tripathi A. Gender-specific responses of Piper betle L. to low
temperature stress: changes in chlorophyllase activity. Biol Plant 2006;50:705–8.[14] Sarkar A, Sen R, Saha P, Ganguly S, Mandal G, Chatterjee M. An ethanolic extract of
leaves of Piper betle (Paan) Linn mediates its anti-leishmanial activity viaapoptosis. Parasitol Res 2008;102:1249–55.
[15] Wirotesangthong M, Inagaki N, Tanaka H, Thanakijcharoenpath W, Nagai H.Inhibitory effects of Piper betle on production of allergic mediators by bonemarrow-derived mast cells and lung epithelial cells. Int Immunopharmacol2008;8:453–7.
[16] Philip HE, William SB. Identification of fungicidal and nemotocidal components inthe leaves of Piper betle (Piperaceae). J Agric Food Chem 1984;32:1254–6.
[17] Bhide SV, Zariwala MBA, Amonlar AJ, Azuine MA. Chemo-preventive efficacy ofbetel leaf extract against benzo (a) pyrene induced fore-stomach tumors in mice.J Ethnopharmacol 1991;34:207–13.
[18] Kumar N. Betel vine (Piper betle L.) cultivation: a unique case of plant establish-ment under anthropogenically regulatedmicroclimatic conditions. Indian J Hist Sci1999;34:19–32.
[19] Verma A, Kumar N, Ranade SA. Genetic diversity among landraces of a dioeciousvegetatively propagated plant, betel vine (Piper betel L.). J Biosci 2004;29:319–28.
[20] Tripathi S, Singh N, Shakya S, Dangi A, Misra-Bhattacharya S, Dube A, et al. Genderbased differences in phenol and thiocyanate contents and biological activity inPiper betle L. Curr Sci 2008;91(6):746–9.
[21] Nelson DS, Mildenhall P. Studies on cytophillic antibodies. The production by miceof macrophage cytophillic antibodies to sheep erthrocytes, relationship to theproduction of other antibodies and development of delayed type hypersensitivity.Aust J Exp Biol Med Sci 1967;45:113–30.
[22] Puri A, Saxena, Sumati RP, Guru PY, Kulshrestha DK, Saxena KC. Immunostimulantactivity of P. kurroa and protection against L. donovani infection in hamsters.Planta Med 1992;58:519–24.
[23] Jerne NK, Nordin AA. Plaque formation in agar by single antibody-producing cells.Science 1963;140:405–8.
[24] Barua CC, Gupta PP, Patnaik GK, Misra-Bhattacharya S, Goel RK, Kulshrestha DK,et al. Immunomodulatory effect of Albizzia Lebbeck. Pharm Biol 2000;38:161–6.
[25] Misra S, Mukherjee M, Dikshit M, Chatterjee RK. Cellular immune response ofgerbils and mastomys in experimental filariasis. Trop Med Int Hlth 1998;3:124–9.
[26] Gupta R, Bajpai P, Tripathi LM, Srivastava VML, Jain SK, Misra-Bhattacharya S.Macrophages in the development of protective immunity against experimentalBrugia malayi infection. Parasitology 2004;129:1–13.
[27] Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SA.Analysis of nitrate, nitrite and [15N] nitrate in biological fluids. Anal Biochem1982;126:131–8.
[28] Zurgil N, Shafran Y, Afrimzon E, Fixler D, Shainberg A, Deutsch M. Concomitant real-timemonitoringof intracellular reactive oxygen species andmitochondrialmembranepotential in individual living promonocytic cells. J Immunol Methods 2006;20:27–41.
[29] Ash LR, Riley JM. Development of sub periodic B. malayi in the jird (Merionesunguiculatus) with notes on infection in other rodents. J Parasitol 1970;56:969–72.
[30] Misra-Bhattacharya S, Katiyar D, Bajpai P, Tripathi RP, Saxena JK. 4-Methyl-7-(tetradecanoyl)-2H-1-benzopyran-2-one: a novel DNA topoisomerase II inhibitorwith adulticidal and embryo static activity against sub periodic Brugia malayi.Parasitol Res 2004;92:177–82.
[31] Singh U, Misra S, Murthy PK, Katiyar JC, Agarwal A, Sircar AR. Immunoreactivemolecules of Brugia malayi and their diagnostic potential. Serodiagn ImmunotherInfect Dis 1997;8:207–12.
[32] Misra S, Chatterjee RK, Sen AB. Antifilarial action of furazolidone. Indian J Med Res1981;73:725–8.
[33] Misra S, Chatterjee RK, Sen AB. The response of Litomosoides carinii to antifilarialagents in cotton rat Sigmodon hispidus and multimammate rat (Mastomysnatalensis). Indian J Med Res 1984;7:749–52.
[34] Singh U, Misra S, Murthy PK, Katiyar JC, Agarwal A, Sircar AR. Immunoreactivemolecules of Brugia malayi and their diagnostic potential. Serodiagn ImmunotherInfect Dis 1997;8:207–12.
[35] O'Brien J, Wilson I, Orton T, Pognan F. Investigation of the Alamar Blue (resazurin)fluorescent dye for the assessment of mammalian cell cytotoxicity. Euro J Biochem2003;267(267):17): 5421–5426.
[36] Brekhman II, Dardimov IV. New substances of plant origin which increase non-specific resistance. Annu Rev Pharmacol 1969;21:219–26.
[37] Dahanukar SA, Kulkarni RA, Rege NN. Pharmacology of medicinal plants andnatural products. Indian J Pharmacol 2000;32:S81–S118.
[38] JayathirthaMG, Mishra SH. Preliminary immunomodulatory activities of methanolextracts of Eclipta alba and Centella asiatica. Phytomedicine 2004;11:361–5.
[39] Ottesen EA, Weller PF, Heck L. Specific cellular immune unresponsiveness inhuman filariasis. Immunology 1977;33:413–21.
[40] Lawrence RA, Devaney E. Lymphatic filariasis: parallels between the immunologyof infection in humans and mice. Parasite Immunol 2001;23:353–61.
[41] Zeng HW, Jiang YY, Bian DGJ, Long K, Cghen ZL. Piperbetol, methylpiperbetol,Piperol A, and Piperol B: a new series of highly specific PAf receptor antagonistsfrom Piper betle. Planta Med 1997;63:296–8.
[42] Sarkar A, Sen R, Saha P, Ganguly S, Mandal G, Chatterjee M. An ethanolic extract ofleaves of Piper betle (Paan) Linnmediates its antileishmanial activity via apoptosis.Parasitol Res 2008;102(6):1249–55.
[43] Sharma S, Khan IA, Ali I, Ali F, KumarM, Kumar A, et al. Evaluation of hydroxychavicolfor its antimicrobial, antioxidant and anti-inflammatory activity for its potential useas oral care agent. Antimicrob Agents Chemother 2008, doi:10.1128/AAC.00045-08[Published online ahead of print].
[44] Sawangjaroen N, Subhadhirasakul S, Phongpaichit S, Siripanth C, Jamjaroen K.Sawangjaroen K. The in vitro anti-giardial activity of extracts from plants that areused for self-medication by AIDS patients in southern Thailand. Parasitol Res2005;95(1):17–21.
[45] Sawangjaroen N, Phongpaichit S, Subhadhirasakul S, Visutthi M, Srisuwan N,Thammapalerd N. The anti-amoebic activity of some medicinal plants used byAIDS patients in southern Thailand. Parasitol Res 2006;98(6):588–92.
[46] Wirotesangthong M, Inagaki N, Tanaka H, Thanakijcharoenpath W, Nagai H.Inhibitory effects of Piper betle on production of allergic mediators by bonemarrow-derived mast cells and lung epithelial cells. Int Immunopharmacol2008;8(3):453–7.
[47] Ganguly S, Mula S, Chattopadhyay S, Chatterjee M. An ethanol extract of Piper betleLinn. mediates its anti-inflammatory activity via down-regulation of nitric oxide.Journal of Pharmacy and Pharmacology, 2007; 59(5): 711–718 (8).
[48] Edgardo ET, Zenaida TE. Cytokine-mimetic properties of some philippine food andmedicinal plants. J Med Food 2007;10(2):290–9.

728 M. Singh et al. / International Immunopharmacology 9 (2009) 716–728
[49] Bandyopadhyay S, Roy KC, Roy M, Pal BC, Bhadra R, Das K, Bhattacharaya S. Herbalformulation of a combination of Piper betel and Murrya koenigii extracts forblocking 5 lipoxygenase activity. United States Patent 6413553, 2002.
[50] Jana BL. Improved technology for betel leaf cultivation. A paper presented in the“Seminar-cum-Workshop on Betel leaf Marketing”, held at State cashew nutfarm, Directorate of Agricultural Marketing, Digha, Midnapur (W. B.), India; 1996.
[51] Jeng JH, Chen SY, Liar CH, Tung YY, Lin BR, Hahn LJ, et al. Modulation of plateletaggregation by areca nut and betel leaf ingredients: roles of relative oxygen speciesand cyclooxygenase. Free Radic Biol Med 2002;32(9):860–71.
[52] Wu MT, Wu DC, Hsu HK, Kao EL, Lee JM. Constituents of areca chewing related toesophageal cancer risk in Taiwanese men. Dis Esophagus 2004;17(3):257–9.
[53] Ramji N, Ramji N, Iyer R, Chandrasekaran S. Phenolic antibacterials from Piper betlein the prevention of halitosis. J Ethnopharmacol 2002;83(1–2):149–52.
[54] Jenie BSL, Andarwulan N, Puspitasari-Nienaber NL, Nuraida L. Antimicrobialactivity of Piper betle Linn extract towards foodborne pathogens and food spoilagemicroorganisms. Food Microbiology: General, IFT Annual Meeting - New Orleans,Louisiana 2001;3(1).
[55] Coffmann RL, SeymourWP, Lebman DA, Hiraki DD, Christiansen JA, Shrader B, et al.The role of helper t cell products in mouse b-cell differentiation and isotyperegulation. Immunol Rev 2006;102(1):5–28.
[56] Murphy KM, Reiner SL. The lineage decisions of helper T cells. Nat Rev Immunol2002(2):933–44.
[57] Belardelli F. Role of interferons and other cytokines in the regulation of theimmune response. APMIS 1995;103(3):161–79.
[58] Kidd P. Th1/Th2 balance: the hypothesis, its limitations, and implications forhealth and disease. Altern Med Rev 2003;8:223–46.
[59] Bandyopadhyay S, Pal B, Bhattacharya S, RayM, Roy KC. Use of betel leaf extracts toinduce IFN-gamma production from human peripheral blood T cells and as a Th1type immunomodulator. United States Patent: 2003; 6: 531,166. Council ofScientific and Industrial Research (New Delhi, IN).
[60] Semnani RT, Law M, Kubofcik J, Nutman TB. Filaria-induced immune evasion:suppression by the infective stage of Brugia malayi at the earliest host-parasiteinterface. J Immunol 2004;172:6229–38.
[61] Calabrese EJ, Baldvin LA. Hormesis: u-shaped dose responses and their centralityin toxicology. Trends Pharmacol Sci 2001;22:285–8.
[62] Tawill S, Le GL, Ali F, Blaxter M, Allen JE. Both free-living and parasitic nematodesinduce a characteristic Th2 response that is dependent on the presence of intactglycans. Infect Immun 2004:398–407.