the pathogenesis of helicobacter pylori–induced gastro-duodenal diseases
TRANSCRIPT

ANRV268-PM01-03 ARI 16 December 2005 15:25
The Pathogenesis ofHelicobacter pylori–InducedGastro-Duodenal DiseasesJohn C. AthertonWolfson Digestive Diseases Centre and Institute of Infections, Immunity, andInflammation, University of Nottingham, Nottingham NG7 2UH, United Kingdom;email: [email protected]
Annu. Rev. Pathol. Mech. Dis.2006. 1:63–96
First published online as aReview in Advance onOctober 3, 2005
The Annual Review ofPathology: Mechanisms ofDisease is online atpathmechdis.annualreviews.org
doi: 10.1146/annurev.pathol.1.110304.100125
Copyright c© 2006 byAnnual Reviews. All rightsreserved
1553-4006/06/0114-0063$20.00
Key Words
Helicobacter pylori, gastric adenocarcinoma, duodenal ulcer, gastriculcer, gastric MALT lymphoma, reflux esophagitis
AbstractHelicobacter pylori is the main cause of peptic ulceration, distal gas-tric adenocarcinoma, and gastric lymphoma. Only 15% of thosecolonized develop disease, and pathogenesis depends upon strainvirulence, host genetic susceptibility, and environmental cofactors.Virulence factors include the cag pathogenicity island, which in-duces proinflammatory, pro-proliferative epithelial cell signaling;the cytotoxin VacA, which causes epithelial damage; and an ad-hesin, BabA. Host genetic polymorphisms that lead to high-levelpro-inflammatory cytokine release in response to infection increasecancer risk. Pathogenesis is dependent upon inflammation, a Th-1acquired immune response and hormonal changes including hy-pergastrinaemia. Antral-predominant inflammation leads to in-creased acid production from the uninflamed corpus and predis-poses to duodenal ulceration; corpus-predominant gastritis leads tohypochlorhydria and predisposes to gastric ulceration and adenocar-cinoma. Falling prevalence of H. pylori in developed countries hasled to a falling incidence of associated diseases. However, whetherthere are disadvantages of an H. pylori-free stomach, for exampleincreased risk of esosphageal adenocarcinoma, remains unclear.
63
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
DISEASES CAUSED BYHELICOBACTER PYLORI
Helicobacter pylori colonizes more than halfthe world’s population and is the main causeof peptic ulceration, gastric adenocarcinoma,and gastric lymphoma. Approximately 80%of peptic ulcer cases are caused by H. py-lori; most of the rest are caused by nons-teroidal anti-inflammatory drugs (NSAIDs).When H. pylori is the etiological factor, itstreatment heals ulcers and prevents their re-currence. Gastric adenocarcinoma is the sec-ond highest cause of cancer deaths world-wide. The nearly one million deaths per yearare due to a combination of high incidence,aggressive disease course, and lack of effec-tive treatment options. H. pylori causes dis-tal but not proximal gastric adenocarcinoma,distal being the much more common form.H. pylori also causes B cell mucosa-associatedlymphoid tissue (MALT) lymphoma of thestomach. These lymphomas can undergohigh-grade transformation but, remarkably,when low-grade, usually resolve following H.pylori treatment. Finally, H. pylori infection isnegatively associated with more severe formsof reflux esophagitis and its sequelae Barrett’sesophagus and esophageal adenocarcinoma.There is much argument about whether thisnegative association is causal.
There has been recent interest in whetherH. pylori may cause or be a risk factor for hu-man diseases outside the upper gastrointesti-nal tract. These include idiopathic thrombo-cytopenic purpura [which appears to improvewith H. pylori treatment (1, 2)], various skindiseases, liver diseases (although these havebeen associated more commonly with Heli-cobacters other than H. pylori), and cardio-vascular and cerebrovascular disease (3, 4).Although interesting and potentially impor-tant, the causality of these associations is un-proven, so the various theories on patho-genesis of these diseases are not discussedfurther.
NATURAL HISTORY OFHELICOBACTER PYLORIINFECTION
H. pylori colonization usually occurs in child-hood, but infection persists lifelong in the ab-sence of treatment. In the rare cases wherecolonization first occurs in adults, it can causea profound gastritis with hypochlohydria, epi-gastric discomfort, and nausea (5, 6). Whetherchildhood colonization causes symptoms orchanges in gastric acidity is unknown. H. py-lori persistence is central to pathogenesis; ul-cers occur mainly in mid- or late adulthoodafter many years of infection and inflam-mation, and gastric adenocarcinoma occursin late adulthood after an even longer pe-riod of chronic inflammation and epithelialdamage.
Colonization of and Persistencein the Gastric Niche
H. pylori is the only human bacterium to per-sistently inhabit the gastric mucus (Figures 1and 2) (7). Elucidation of its complete genomesequence has allowed construction in silico ofits metabolic pathways and other aspects of itsbiology (8, 9) and postgenomic techniques us-ing whole genome strategies and microarraytechnology (for example, see Reference 10)have speeded experimentation. This and otherwork show that factors important for colo-nization include motility (Figure 1), environ-mental sensing, chemotaxis (11), iron acquisi-tion (12), and acid resistance. Acid resistanceis crucial in the gastric niche, and more than300 genes are acid-regulated or acid-affected(13, 14). One of these genes encodes the mostabundant protein of H. pylori, urease, whichhydrolyzes urea to ammonia and carbon diox-ide. This leads to a rise in cytoplasmic pH,which buffers the periplasm and allows sur-vival in acidic conditions. Urea entry into thecytoplasm is regulated by a unique acid-gatedurea channel (UreI), allowing instantaneous
64 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.
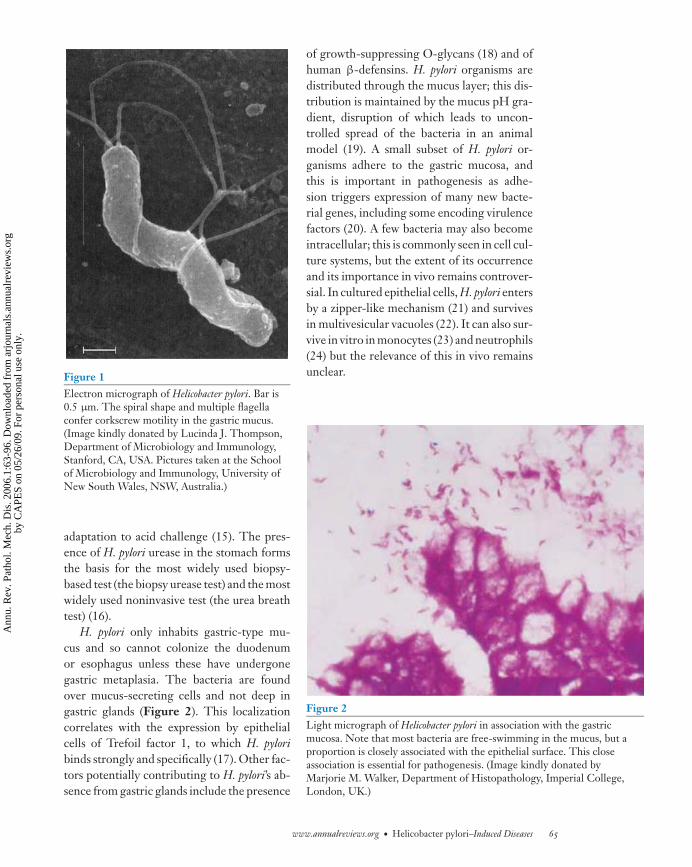
ANRV268-PM01-03 ARI 16 December 2005 15:25
Figure 1Electron micrograph of Helicobacter pylori. Bar is0.5 μm. The spiral shape and multiple flagellaconfer corkscrew motility in the gastric mucus.(Image kindly donated by Lucinda J. Thompson,Department of Microbiology and Immunology,Stanford, CA, USA. Pictures taken at the Schoolof Microbiology and Immunology, University ofNew South Wales, NSW, Australia.)
adaptation to acid challenge (15). The pres-ence of H. pylori urease in the stomach formsthe basis for the most widely used biopsy-based test (the biopsy urease test) and the mostwidely used noninvasive test (the urea breathtest) (16).
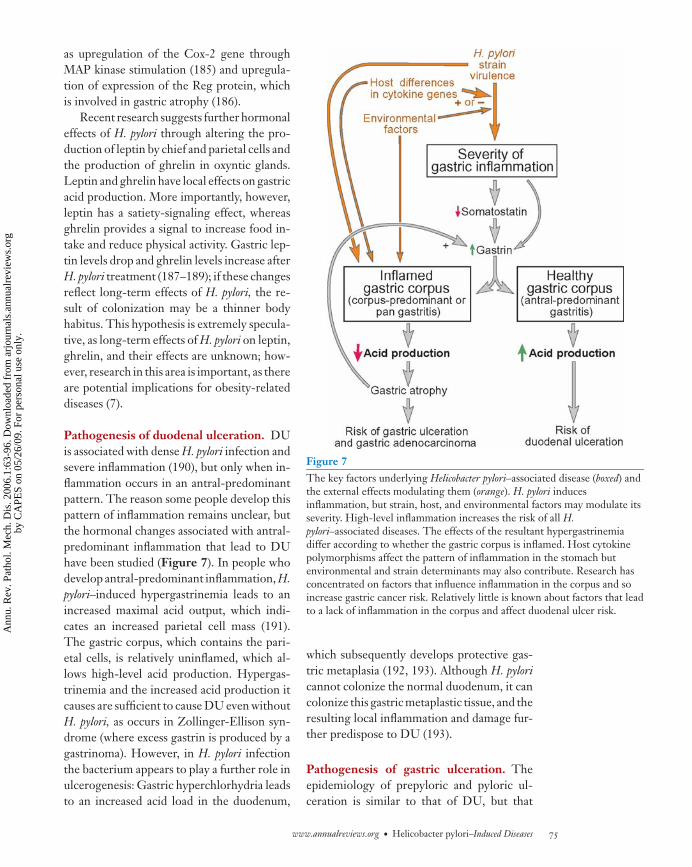
H. pylori only inhabits gastric-type mu-cus and so cannot colonize the duodenumor esophagus unless these have undergonegastric metaplasia. The bacteria are foundover mucus-secreting cells and not deep ingastric glands (Figure 2). This localizationcorrelates with the expression by epithelialcells of Trefoil factor 1, to which H. pyloribinds strongly and specifically (17). Other fac-tors potentially contributing to H. pylori’s ab-sence from gastric glands include the presence
of growth-suppressing O-glycans (18) and ofhuman β-defensins. H. pylori organisms aredistributed through the mucus layer; this dis-tribution is maintained by the mucus pH gra-dient, disruption of which leads to uncon-trolled spread of the bacteria in an animalmodel (19). A small subset of H. pylori or-ganisms adhere to the gastric mucosa, andthis is important in pathogenesis as adhe-sion triggers expression of many new bacte-rial genes, including some encoding virulencefactors (20). A few bacteria may also becomeintracellular; this is commonly seen in cell cul-ture systems, but the extent of its occurrenceand its importance in vivo remains controver-sial. In cultured epithelial cells, H. pylori entersby a zipper-like mechanism (21) and survivesin multivesicular vacuoles (22). It can also sur-vive in vitro in monocytes (23) and neutrophils(24) but the relevance of this in vivo remainsunclear.
Figure 2Light micrograph of Helicobacter pylori in association with the gastricmucosa. Note that most bacteria are free-swimming in the mucus, but aproportion is closely associated with the epithelial surface. This closeassociation is essential for pathogenesis. (Image kindly donated byMarjorie M. Walker, Department of Histopathology, Imperial College,London, UK.)
www.annualreviews.org • Helicobacter pylori–Induced Diseases 65
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
Helicobacter pylori–InducedInflammation and Evasionof the Immune Response
All strains of H. pylori induce a markedimmune response with local neutrophil, lym-phocyte and other inflammatory cell infil-tration, and both local and systemic anti-body production and cell-mediated responses.This is important in pathogenesis but inef-fective at clearing the infection. Many H. py-lori proteins, particularly urease (25, 26) anda bacterioferritin named HP-NAP (H. pylorineutrophil-activating protein) (27, 28), con-tribute to the genesis of inflammation. How-ever, inflammation is more intense and diseasemore likely if the H. pylori strain expresses spe-cific virulence factors.
Although H. pylori does cause inflamma-tion, the bacteria has both minimized theextent to which it provokes an inflamma-tory response and developed mechanisms forevading it. It has evolved its lipopolysaccha-ride and flagellin to minimize recognition bythe innate immune system’s toll-like recep-tors (TLR4 and 5, respectively) (29–31). Ithas also evolved to interfere with local im-mune responses; for example, it inhibits nitricoxide production by macrophages (32) anddownregulates chemokine receptors on neu-trophils (33). H. pylori also directly blocks im-mune cell products that kill bacteria; for exam-ple, it produces enzymes, such as methioninesulfoxide reductase, that combat bacteriocidaloxidative stress (34). Finally, there is increas-ing evidence that the organism influences theacquired immune response to render it inef-fective in H. pylori clearance; this point is dis-cussed later.
Adaptation and Genetic Diversity
H. pylori strains acquire DNA through hori-zontal acquisition from other strains more fre-quently than do any other bacterial species.This leads to immense genetic diversity (35).
Strains adapt within individual stomachs byexchange, loss, and gain of DNA (36); onestudy has estimated a mean of 60 small DNAimports per year (37). This genetic variationlikely allows adaptation to different niches,including perhaps different gastric microen-vironments and different individual humanhosts (38). Adaptation and generation of di-versity also appears important for persistence:In an animal model, mutations that block ge-netic adaptation lead to a polarized T helper(Th)-1-type response rather than a Th2 re-sponse, which in turn leads to early clearanceof infection (39).
PATHOGENESIS: WHY DO ONLYSOME INFECTIONS RESULT INDISEASE?
All strains of H. pylori persist lifelong andall cause gastric inflammation. However, only15% of infections result in peptic ulcera-tion and only 0.5%–2% in gastric adenocar-cinoma. Who develops disease depends uponthree factors: (a) the virulence of the infect-ing H. pylori strain, (b) the type and extentof the host immune response to infection,and (c) modulating cofactors such as smok-ing and diet. H. pylori virulence factors maydamage epithelial cells directly or stimulatethem to produce proinflammatory cytokines,thus inducing inflammation. Infections caus-ing high-level inflammation increase the riskof all H. pylori–induced diseases. Which dis-ease develops depends upon the type and dis-tribution of inflammation in the stomach. Thedeterminants of the distribution of inflamma-tion in the stomach are unclear, but host ge-netics have a proven role, and environmentaland strain factors may also be important. Peo-ple with H. pylori–induced duodenal ulcers areless likely than others to develop gastric ade-nocarcinoma later in life (40), implying thatH. pylori infection predisposes an individualto one or another of these diseases, but notusually to both.
66 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
Bacterial Virulence Factors
The cytotoxin-associated gene pathogeni-city island and cytotoxin-associated geneA. The name of the cytotoxin-associatedgene (cag) pathogenicity island (PAI) is mis-leading because (a) the vacuolating cytotoxingene, vacA, is on a different part of the H. py-lori genome, and (b) both the expression andtoxin activity of VacA are independent of thecag PAI. PAIs are large sections of DNA thatare acquired by bacterial species and renderthem pathogenic. The cag PAI, which con-tains approximately 30 genes, was acquired byH. pylori distantly in its evolution from an un-known source and inserted into its glutamateracemase gene (41). Strains may either possessthe whole PAI (cag+), lack it entirely (cag-), orhave parts of it deleted; these latter strains areusually phenotypically similar to cag- strains(42). Proportions of cag+, cag−, and interme-diate strains vary in different countries, but inall countries cag+ strains predominate and inmany, such as Japan and China, they are vir-tually ubiquitous. In large disease associationstudies, one of the PAI genes, cagA, or, alter-natively, a serological response to the CagAprotein, has been used as a marker of thecag PAI. cagA+ strains are more commonlyassociated with both peptic ulceration andgastric adenocarcinoma than cagA- strains,which are rarely, if ever, disease-associated(43–46). The high prevalence of cagA+ strainsin countries like Japan may contribute to thehigh incidence of H. pylori–associated dis-eases, such as gastric adenocarcinoma, in thesecountries.
Genes in the cag PAI encode proteins thatcomprise a type IV secretion system (T4SS)used for translocating bacterial products di-rectly into the host cell cytoplasm (41). TheT4SS can be visualized microscopically as asheathed, rigid needle linking H. pylori to theepithelial cell (47, 48). The protein CagA istranslocated through this needle into epithe-lial cells (29, 49–52), where it is phospho-rylated on tyrosine residues by host cell Srcfamily kinases and stimulates cell-signaling
pathways (53, 54) (Figure 3). The most obvi-ous effect in vitro is stimulation of cell-shapechange such that the cell elongates and devel-ops long processes. This has been termed thehummingbird phenotype (49, 69). These andother non-cag-dependent (49, 70) cytoskeletalchanges may be important in forming closeinteractions between bacteria and cells (49,71). Interestingly, one study has shown thathummingbird changes are dependent uponCagA-induced activation of the hepatocytegrowth factor receptor c-Met, which is anoncogene (56). A second major effect of CagAis stimulation of transcription factors involvedin multiple cell functions, including cellularproliferation, through activation of c-Fos andc-Jun (62, 65). Unregulated CagA signalingcan lead to apoptosis (67), but activated (phos-phorylated) CagA inhibits the Src kinase thatphosphorylates it, thus creating a negative-feedback loop and perhaps minimizing suchuncontrolled effects (54, 67). In vivo, cag+ H.pylori is associated with epithelial prolifera-tion, but it is unclear whether it is predom-inantly pro- (72) or anti-apoptotic (73). It isimportant to determine which is the case; adisrupted balance in favor of apoptosis mayaffect epithelial restitution and ulcer healing,whereas uncontrolled proliferation may pre-dispose to carcinogenesis. A final function ofCagA, which is not dependant upon its tyro-sine phosphorylation, is to disrupt apical junc-tional complexes between epithelial cells, re-sulting in loss of barrier function (55). Thismay benefit the bacteria by allowing nutrienttransfer and could also be pathogenic throughdestroying the epithelial cell barrier.
Most strains express the CagA protein,but the protein differs in structure betweenstrains. Firstly, the CagA protein may havebetween zero and five active tyrosine phos-phorylation sites; more sites lead to higherlevels of CagA phosphorylation in epithelialcells and more profound cytoskeletal change(59, 74). Strains with more sites are more com-monly isolated from people with precancerousgastric changes (e.g., atrophy and intestinalmetaplasia) and with gastric cancer (74–76).
www.annualreviews.org • Helicobacter pylori–Induced Diseases 67
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
Figure 3CagA signaling and effects in epithelial cells. The green oval is the CagA protein. Red Ps representtyrosine phosphorylation. Dashed black lines show activation where precise pathways are unclear.Dashed blue lines and blue boxes show end effects of CagA on the epithelial cell. CagA is translocatedthrough a bacterial type IV secretion system directly into the epithelial cytoplasm, where it isphosphorylated on tyrosine residues by host cell Src kinases. Phosphorylated CagA binds to and activatesSHP-2 phosphatase (Src homology 2–containing protein-tyrosine phosphatase-2), which leads towidespread dephosphorylation of host proteins and multiple cellular effects. These effects aredownregulated, however, by inhibition by phosphorylated CagA of the Src kinases that activated it.Other effects of Src kinases are also downregulated, leading to cytoskeletal changes. CagA binds to thezona occludens 1 protein (ZO-1), affecting junctional complexes and epithelial integrity. It transactivatesthe epidermal growth factor receptor (EGFR) and similar receptors leading to extracellularsignal-regulated protein kinase (ERK) signaling and ultimately to activator protein-1 (AP-1)–mediatedpro-proliferative effects. ERK and JNK (c-Jun N-terminal kinase) MAP (mitogen-activated protein)kinase pathways are also activated by phosphorylated CagA. Finally, CagA associates with the c-Metreceptor as an adaptor protein and this is necessary for epithelial cell motility. Effects differ somewhatbetween different cell lines, and it is not clear which effects are most pertinent in vivo (53–68).
Secondly, CagA from East Asian strains has adifferent binding site for the signaling proteinSHP-2 (see Figure 3) and binds this proteinmore avidly, thus inducing higher levels of cy-toskeletal change. Such strains are associatedwith higher levels of inflammation and atro-
phy and are more likely to be isolated frompatients with gastric cancer (77).
The main neutrophil-attracting chemo-kine in H. pylori gastritis is interleukin (IL)-8, and cag+ strains induce higher levelsof IL-8 production in epithelial cells than
68 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
cag– strains. This effect is dependent upon ex-pression of at least mainly intact cag-encodedT4SS, but is not dependent upon CagAtranslocation and phosphorylation (66, 78).Contact between the cag-encoded T4SS andepithelial cells allows soluble components ofthe bacterial peptidoglycan cell wall, knownas muramyl dipeptides, to enter the cell; thesepeptides are detected by Nod1, part of thecell’s system for recognizing intracellular bac-teria (79). This detection then stimulates sig-naling through nuclear factor κB (NFκB) ac-tivation and nuclear translocation (79–81),which in turn leads to transcription of variousgenes, including those encoding IL-8, humanβ-defensin 2 (82), anti-apoptotic factors (83),and matrix metalloproteinases 7 and 9 (81, 84).The importance of the cag PAI for inflamma-tion and disease has been demonstrated in theMongolian gerbil model. H. pylori–infectedgerbils develop gastric ulcers, precancerousgastric changes, and ultimately, in somecases, gastric adenocarcinoma. Infection withan isogenic mutant strain unable to make theT4SS results in none of these effects (85).
In summary, the cag PAI is needed fordisease, but not all cag+ strains are disease-associated. The cag-encoded T4SS induces in-flammation independently of CagA, and thisinflammation is necessary for disease. Strainsthat deliver more active CagA are more com-monly associated with precancerous changesand cancer. Unfortunately, however, this asso-ciation between strain pathogenicity and dis-ease is complicated by the fact that strains canevolve to change their pathogenicity. The sizeand importance of this effect is undetermined,but individual examples have been found ofstrains losing the cag PAI or gaining or losingCagA tyrosine phosphorylation sites withinindividual stomachs (86, 87). Such evolutioncould potentially benefit H. pylori by allowingadaption to a changing gastric niche. How-ever, the implications for pathogenesis remainunclear.
The vacuolating cytotoxin. Unlike the cagPAI, the vacuolating cytotoxin gene, vacA, is
present in all strains. However, it is polymor-phic, varying most markedly in two regionstermed the mid- and signal regions (88, 89)(Figure 4). The main signal region types ares1 and s2, and the mid-region types are m1and m2. The vacA gene may comprise anycombination of signal and mid-region types,although the s2/m1 combination is rare (89,90). The importance of type-s2 vacA is thatits product is virtually nontoxic (89, 91) andamong s1-type vacA, s1/m1 is toxic for a widerrange of cells than s1/m2 (91, 92). In ob-servational studies in humans, vacA s2-typestrains are rarely associated with disease (93–96). Although both vacA s1/m1- and s1/m2-type strains may be disease-associated, whereboth are common, patients with gastric ade-nocarcinoma usually have the s1/m1-type ofvacA (97–99). In Japan, s1/m1-type vacA isubiquitous, perhaps contributing to the highincidence of gastric adenocarcinoma. In thefew instances where strains are nontoxic inJapan, this is owing usually to inactivating mu-tations in s1/m1-type vacA rather than the oc-currence of other types (100).
vacA is translated into a pre-protoxin,which is an autotransporter that undergoes N-and C-terminal cleavage during bacterial se-cretion (101) (Figure 4). The mature secretedtoxin comprises two subunits, which remainassociated. Both are required for toxin activity,and the larger p58 subunit mediates bindingof the toxin to the epithelial cell. Studies of thetoxin and its effects on cells have concentratedon in vitro effects of the most active (andmost disease-associated) s1/m1 form. VacAcan be transferred to epithelial cells eitherby secretion or by contact-dependent transfer,which may be a more efficient method (102).It can bind to several epithelial cell recep-tors: (a) receptor protein tyrosine phosphatase(RPTP) β and α (103–105), (b) the epider-mal growth factor receptor (EGFR) (106),and (c) a glycosylphosphatidylinositol (GPI)-anchored protein associated with lipid rafts(107, 108). In vitro, VacA is internalized byendocytosis (108). Both internalization andsubsequent effects are dependent upon acid
www.annualreviews.org • Helicobacter pylori–Induced Diseases 69
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
Figure 4Polymorphism in the vacuolating cytotoxin protein, VacA, between Helicobacter pylori strains. The vacAgene varies most markedly in the signal region (encoding the signal peptide), which may be type s1 or s2,and in the mid-region (encoding part of the p58 binding subunit), which may be type m1 or m2. Threetypes of toxin are commonly found: s1/m1, s1/m2, and s2/m2 (the s2/m1 combination occurs but israre).
activation of VacA, making the toxin par-ticularly well suited to the stomach (109,110).
VacA is a pore-forming toxin (111), andthis action underlies many of its other ef-fects, including vacuolation (112) (Figure 5).VacA forms hexameric pores, which are selec-tive to anions and small neutral molecules, in-cluding urea. This urea permease activity maybe an important aid to acid survival throughproviding a substrate for urease (113). Whensurface pores are endocytosed, they becomeprocessed into late-endosomal compartments(114), which then undergo osmotic swellingto become large acidic vacuoles. Whereas thisvacuolation is the most obvious effect of VacAin vitro, is not marked in vivo. This fact hasled to interest in other VacA functions. Oneimportant effect may be increased paracellu-lar permeability and potential nutrient acqui-sition (115). Such a role would fit well with the
Figure 5Atomic force microscopy image of the vacuolatingcytotoxin, VacA, on an artificial lipid membrane.The “flowers” represent anion-selectivemembrane pores. The line drawing in the topright corner shows a VacA hexamer with adiameter of 28 nm. (Reproduced with permissionfrom Reference 111. Copyright 1999 NationalAcademy of Sciences USA.)
70 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
expression of vacA in nutrient-depleted con-ditions such as iron starvation (116). Anotherinteresting effect of VacA is induction of apop-tosis (117, 118) at least partly through a novelmechanism of toxin-induced pore formationin mitochondria that allows cytochrome c re-lease (119, 120). However, high concentra-tions are needed for this effect; consequently,there is some doubt about its importancein vivo (120). More subtle effects include in-terference with cell signaling, as VacA stimu-lates p38 and ERK1/2 MAP kinases (121).
VacA causes epithelial damage in mousemodels both when given orally as a singleagent (101) and when delivered by a toxi-genic strain of H. pylori during gastric in-fection (122). Interestingly, mice null for theVacA receptor RPTPβ do not develop ul-cers (123) and work in vitro shows that, al-though cells lacking this receptor can still takeup VacA and undergo vacuolation, both cellsignaling and detachment from a basementmembrane are blocked (123). Although VacAis not needed for proinflammatory signalingby epithelial cells, it may induce cytokine re-lease from other cell types [for example, mastcells (124)]. However, recent interest has fo-cused more on the immunosuppressive ratherthan the immunostimulatory effects of VacA.In vitro, VacA can interfere with antigen pre-sentation, which occurs in compartments thatare similar to late endosomes (125). Moresurprisingly, it can specifically inhibit T cellactivation and proliferation (126–128). Thisobservation has led to speculation that VacAmay help prevent clearance of highly host-interactive (cag+) H. pylori strains. In sup-port of this (or at least some gain of functionfor possessing both the cag PAI and an activeform of VacA), possession of the cag PAI andexpression of an active s1 form of the cyto-toxin are closely associated features of H. pyloristrains.
Some forms of VacA are nontoxic and lessoften associated with disease (Figure 4). Thes2 form has a short N-terminal peptide ex-tension on the mature protein. This extensionblocks vacuolating activity (91, 129), leads to
slower, less effective pore formation (130), anddoes not cause apoptosis (118). The s1/m1form is fully active, but the sl/m2 form bindsto and vacuolates a narrow range of epithe-lial cell types; this activity is a direct con-sequence of changes in the binding subunitencoded by the vacA m region (91, 92). Lessactive forms of vacA are maintained in theH. pylori population, but their benefit toH. pylori is unclear. Interestingly, H. pylori canevolve through recombination to change itstoxicity (131), potentially allowing adaptationand changing levels of host interaction. How-ever, the frequency with which this recombi-nation occurs and its importance in pathogen-esis remain undefined.
Helicobacter pylori adhesion and bloodgroup antigen-binding adhesin A. Al-though most H. pylori organisms are freelivingin gastric mucus, a small proportion adhereto epithelial cells. Adhesion is, at least in partthrough binding to blood group antigens, ex-pressed on gastric epithelial cells. The mostimportant of these antigens is Lewis b (132).Lewis b expression in the stomach is associ-ated with higher H. pylori density; in stomachsnot expressing Lewis b, expression of otherblood group antigens such as Lewis x andLewis a also increase bacterial density, but to alesser extent (133). Transgenic mice express-ing human Lewis b develop gastritis and gas-tric ulcer, whereas wild-type mice do not, con-firming the importance of both Lewis b andbacterial-epithelial adhesion for disease (134).Lewis x binding is mediated by the H. pylorisialic-acid-binding adhesin, SabA (135), butresearch has concentrated on Lewis b bind-ing through the blood group antigen-bindingadhesin (BabA) (136). Strains possessing thegene babA2, which encodes active BabA, areassociated with increased epithelial prolifera-tion and inflammation and also increased riskfor duodenal ulcer, gastric atrophy, intestinalmetaplasia, and gastric adenocarcinoma (137–139). Lewis b binding can change by muta-tion through several mechanisms. For exam-ple, babA2 can be lost during experimental
www.annualreviews.org • Helicobacter pylori–Induced Diseases 71
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
infection in Macaque monkeys (140). In vitro,if babA2 is disrupted, bacteria can regain theLewis b binding phenotype by recombininga nonexpressed homolog, babA1, into anotherlocus, babB, where it is then expressed (141,142). There is good evidence that binding ofH. pylori has evolved in specific human popu-lations to adapt to that population: In indige-nous South American populations, which areblood group O, H. pylori strains have evolvedBabA to bind blood group O antigens best(38).
Outer inflammatory protein A. Outer in-flammatory protein A (OipA) is an outermembrane protein of H. pylori that con-tributes to the induction of IL-8 productionby epithelial cells (143). It is present in allstrains, but variable numbers of dinucleotiderepeats in its 5′ region mean that an activeprotein may or may not be produced becauseof a common mutational adaption system inbacteria known as slipped strand mispairing(143). “On” status is associated in vivo withincreased bacterial density, increased IL-8 lev-els in the gastric mucosa, increased neutrophilinfiltration and duodenal ulceration, but notwith atrophy (143).
Induced-by-contact-with-epithelium ge-ne A and Helicobacter pylori methylases.Contact of H. pylori with epithelium inducestranscription of the induced-by-contact-with-epithelium gene (iceA1), and presence of thisgene rather than a distant homologue, iceA2,has been associated with duodenal ulceration(DU) and gastric adenocarcinoma (144, 145).The role of iceA1 in pathogenesis is not ob-vious, as it does not usually encode a func-tional protein and has homology with a re-striction endonuclease gene in other bacteria.As restriction endonucleases act as bacterialdefense agents that break down foreign bac-terial DNA, they are not usually associatedwith pathogenesis. Consequently, disease as-sociations with the iceA genes may be chancefindings. However, an alternative possibilityis that expression of the methylase hpy1M,
which accompanies iceA1 (all restriction en-zymes are associated with a specific methy-lase to protect self-DNA from the restrictionenzyme activity), may be important. Methy-lases have a demonstrated role in virulencein Salmonella typhimurium (146). In H. pylori,the presence of iceA1 leads to higher expres-sion of its cognate methylase gene, hpyIM,than does the presence of iceA2 (147). AnotherH. pylori methylase gene, hpyIIIM, can also bedifferentially transcribed depending upon thegene in the immediate upstream locus (148).However, in this case, methylase function islost via the replacement of hpyIIIM by theH. pylori restriction endonuclease replacinggene A (hrg); this genotype has been weaklyassociated with cancer in Asian populations(148, 149). Further studies are needed to con-firm whether methylation has a direct role inH. pylori virulence.
Host Genetic Polymorphisms andHelicobacter pylori–Induced Disease
Polymorphisms in human cytokine genes af-fect the level of cytokine production by cellsafter contact with H. pylori. Specific poly-morphisms in the IL-1β gene and the IL-1receptor-antagonist gene (IL-1RN) lead to in-creased gastric mucosal levels of IL-1β in in-dividuals infected with H. pylori (150, 151) andincreased levels of inflammation (150–154).The polymorphisms also increase the risk ofgastric atrophy, achlorhydria, intestinal meta-plasia (151, 152, 154, 155), and distal gas-tric adenocarcinoma (46, 153, 155–158). Theeffect of these polymorphisms on duodenal ul-cer risk is uncertain; one report showed an in-creased risk with proinflammatory polymor-phisms (159), but another showed a reducedrisk (152). Such a reduced risk would be logi-cal for people with an atrophic, hypochlorhy-dric stomach. Although polymorphisms af-fecting IL-1β levels are best studied, othersalso appear important. Tumor necrosis fac-tor α (TNFa) polymorphisms have not beendemonstrated to affect gastric mucosal levelsof TNFα (151), but they have been shown to
72 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
affect gastric inflammation and cancer risk insome (153, 157) but not all (151, 158) stud-ies. Polymorphisms affecting the immuno-suppressive cytokine IL-10 affect mucosallevels and may lead to colonization by morevirulent H. pylori strains (151); they alsoincrease gastric cancer risk (157). IL-1β,TNFα, and H. pylori all induce IL-8 secre-tion by epithelial cells; an IL-8 polymorphismcausing increased IL-8 production in vitrohas been associated with an enhanced risk ofgastric adenocarcinoma and gastric ulcer in aJapanese population (160).
Carriage of multiple proinflammatorypolymorphisms increases the level of gastricinflammation associated with H. pylori infec-tion and the risk of atrophy, intestinal meta-plasia, and gastric carcinoma (153, 157). Asexpected, infection with H. pylori strainspossessing known virulence factors furtherincreases these risks (46, 154). Individualswith both multiple proinflammatory cytokinepolymorphisms and the most virulent H. pyloristrains can have up to fiftyfold increased riskof gastric cancer as compared with individualslacking these factors.
Importance of the Immune Responsein Pathogenesis
H. pylori–induced inflammation is central tothe pathogenesis of all H. pylori–associateddiseases. It is increased by proinflammatorycytokine polymorphisms and by bacterial vir-ulence factors such as the cag PAI and OipA.It is also modulated by T helper cells, whichcontrol the type of response and its cy-tokine profile. Acquired immune responses tomicro-organisms can be predominantly Th-1(mainly cell mediated) or Th-2 (mainly an-tibody dependent). H. pylori stimulates bothresponses, although gastric mucosal cytokineprofiles suggest that the Th-1 response pre-dominates (Figure 6) (161, 162). The Th-1response is important in the pathogene-sis of H. pylori–induced disease. Mice ei-ther genetically prone to or manipulatedto have a Th-1 response (for example, IL-
4 null mice) develop more intense H. py-lori gastritis (163, 164). T cell transfer ex-periments show that Th-1 cells are directlyresponsible (163). Mice genetically manipu-lated to have a Th-1 response develop in-flammation, epithelial apoptosis, and disrup-tion of cell organization in gastric glandseven in the absence of H. pylori (165). Incontrast, mice deficient in T cell responsesdo not show these changes. Even infusionof the Th-1 cytokine interferon-γ (IFN-γ)alone induces gastric inflammation and at-rophy, although effects are enhanced withhelicobacter coinfection (166). In addition,
Figure 6The central role of the T helper (Th)-cellresponse to Helicobacter pylori infection. Th-1,Th-2, and regulatory T cells are activated, but thebalance of the response varies between peopleowing to unknown factors. Studies in animalmodels and supportive data in humans (see text)show that a strong Th-1 response increasesseverity of gastritis and cancer risk. Both H. pyloriand the host downregulate this Th-1 response.This potentially benefits H. pylori by allowingmore dense colonization and benefits the host bydownregulating mucosal inflammation.
www.annualreviews.org • Helicobacter pylori–Induced Diseases 73
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
IFN-γ–deficient mice have less inflammationand less atrophy when infected with heli-cobacter (164, 167, 168), a result that furtherconfirms the importance of IFN-γ.
H. pylori appears to induce a Th-1 responseby interaction with dendritic cells. In vitro,H. pylori coculture with dendritic cells inducesmaturation and activation of high-level IL-12 production, characteristic of a Th-1 re-sponse. This effect is at least partially de-pendent upon the presence of the cag PAI(169, 170). Once matured, dendritic cells arealso resistant to H. pylori–induced apoptosis;this resistance potentially allows these cellsto fulfil their T helper–directing function(171).
An unregulated Th-1 response is poten-tially deleterious to H. pylori, as bacterial den-sity is reduced (163, 172–174). H. pylori ap-pears to have adapted to downregulate theTh-1 response it induces, possibly to avoidclearance. H. pylori strains expressing Lewisantigens on their lipopolysaccharide bind thedendritic cell-specific intracellular adhesionmolecule-3 (ICAM3)-grabbing nonintegrin(DCSIGN) lectin on gastric dendritic cellsand thus block Th-1 cell development. Inter-estingly, Lewis-negative variants do not bindDCSIGN; these strains induce a strong Th-1response, which suggests rapid adaption ofH. pylori to modulate the T helper–cell bal-ance (175). H. pylori also activates Cox-2 inmononuclear cells, which reduces Th-1 polar-ization (176). Regulatory T cells may down-regulate the Th-1 response: T-memory-cellsfrom H. pylori–infected people proliferate andproduce IFN-γ less effectively than thosefrom H. pylori–uninfected people; this phe-nomenon is dependent upon regulatory Tcell activity, as depletion of regulatory T cellsabolishes the effect and addition of regula-tory T cells induces it (177). Finally, the Th-1 response can be downregulated by externalfactors. The Helicobacter felis–infected mousemodel has been immunologically manipulatedby experimental helminth coinfection; thisleads to increased H. pylori density, but re-duced Th-1 cytokine levels and no gastric at-
rophy (172). It has been suggested that thehigh incidence in Africa of parasitic infections,which suppress Th-1 responses, may explainthe so-called African enigma—the low inci-dence of H. pylori–associated disease in Africadespite high H. pylori prevalence.
The Role of Hormonal Changesand Acid Homeostasis in thePathogenesis of Ulcers andGastric Adenocarcinoma
Helicobacter pylori–induced hormonalchanges and their importance in dis-ease. H. pylori infection is associated withreduced numbers of somatostatin-producingD cells in the stomach and reduced so-matostatin production (178–180). This maybe partly due to suppression of somato-statin release by inflammatory mediators,including nitric oxide (181). Hypergastrine-mia develops because gastrin productionfrom G cells is released from its usualnegative feedback by somatostatin; a di-rect stimulatory action of cytokines onG cells may also contribute (Figure 7) (179).In a mouse model, the Th-1 cytokine IFN-γinduces hypergastrinemia and reduces so-matostatin levels (182). The gastritis inducedin this model is reversed by administering thesomatostatin analogue octreotide, implyingthat Th-1 responses drive inflammationthrough hormonal changes. Direct evidenceof the importance of hypergastrinemia in dis-ease comes from the gastrin-overexpressingINS-GAS mouse model. These mice haveincreased acid secretion, but in later life de-velop reduced acid secretion, gastric atrophy,and gastric adenocarcinoma (183). Thesephenomena occur even in the absence ofhelicobacter infection, although its presenceconsiderably speeds the process. In the H.pylori–infected gerbil model, the course ofhypergastrinemia development correlateswith epithelial cell proliferation, which mayunderlie the rapid progression to atrophy(184). Hypergastrinemia has a number ofother potentially prooncogenic effects such
74 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.
ANRV268-PM01-03 ARI 16 December 2005 15:25
as upregulation of the Cox-2 gene throughMAP kinase stimulation (185) and upregula-tion of expression of the Reg protein, whichis involved in gastric atrophy (186).
Recent research suggests further hormonaleffects of H. pylori through altering the pro-duction of leptin by chief and parietal cells andthe production of ghrelin in oxyntic glands.Leptin and ghrelin have local effects on gastricacid production. More importantly, however,leptin has a satiety-signaling effect, whereasghrelin provides a signal to increase food in-take and reduce physical activity. Gastric lep-tin levels drop and ghrelin levels increase afterH. pylori treatment (187–189); if these changesreflect long-term effects of H. pylori, the re-sult of colonization may be a thinner bodyhabitus. This hypothesis is extremely specula-tive, as long-term effects of H. pylori on leptin,ghrelin, and their effects are unknown; how-ever, research in this area is important, as thereare potential implications for obesity-relateddiseases (7).
Pathogenesis of duodenal ulceration. DUis associated with dense H. pylori infection andsevere inflammation (190), but only when in-flammation occurs in an antral-predominantpattern. The reason some people develop thispattern of inflammation remains unclear, butthe hormonal changes associated with antral-predominant inflammation that lead to DUhave been studied (Figure 7). In people whodevelop antral-predominant inflammation, H.pylori–induced hypergastrinemia leads to anincreased maximal acid output, which indi-cates an increased parietal cell mass (191).The gastric corpus, which contains the pari-etal cells, is relatively uninflamed, which al-lows high-level acid production. Hypergas-trinemia and the increased acid production itcauses are sufficient to cause DU even withoutH. pylori, as occurs in Zollinger-Ellison syn-drome (where excess gastrin is produced by agastrinoma). However, in H. pylori infectionthe bacterium appears to play a further role inulcerogenesis: Gastric hyperchlorhydria leadsto an increased acid load in the duodenum,
Figure 7The key factors underlying Helicobacter pylori–associated disease (boxed) andthe external effects modulating them (orange). H. pylori inducesinflammation, but strain, host, and environmental factors may modulate itsseverity. High-level inflammation increases the risk of all H.pylori–associated diseases. The effects of the resultant hypergastrinemiadiffer according to whether the gastric corpus is inflamed. Host cytokinepolymorphisms affect the pattern of inflammation in the stomach butenvironmental and strain determinants may also contribute. Research hasconcentrated on factors that influence inflammation in the corpus and soincrease gastric cancer risk. Relatively little is known about factors that leadto a lack of inflammation in the corpus and affect duodenal ulcer risk.
which subsequently develops protective gas-tric metaplasia (192, 193). Although H. pyloricannot colonize the normal duodenum, it cancolonize this gastric metaplastic tissue, and theresulting local inflammation and damage fur-ther predispose to DU (193).
Pathogenesis of gastric ulceration. Theepidemiology of prepyloric and pyloric ul-ceration is similar to that of DU, but that
www.annualreviews.org • Helicobacter pylori–Induced Diseases 75
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
of H. pylori–induced ulceration elsewhere inthe stomach is more similar to that of gastricadenocarcinoma. Unlike DU, gastric ulcera-tion (GU) is associated with a pan-gastriticinflammation pattern and reduced or normalacid production. The pathogenesis of GUis poorly understood, but GUs arise mostcommonly at the transitional zone betweenantrum and corpus on the lesser gastric curve.This area in particular can be densely col-onized by H. pylori and become heavily in-flamed (194); it is likely that the heavy inflam-mation and epithelial damage at this site leaddirectly to ulceration.
Pathogenesis of gastric adenocarcinoma.Gastric adenocarcinoma is associated withpan-gastritis or corpus-predominant gastritis(Figure 7). Autoimmune gastritis also has thispattern of inflammation and is an independentrisk factor for distal gastric adenocarcinoma.Why H. pylori–induced inflammation has apan-gastritic or corpus-predominant patternin some people when it is antral-predominantin others is unknown. One possibility is that,like autoimmune gastritis, it may be caused byimmune effectors with specificity for the gas-tric proton pump ATPase. In support of this,T cells have been identified that cross-reactbetween H. pylori proteins and the ATPase(195). Pan-gastritis is associated with reducedacid secretion, despite hypergastrinemia (196)(Figure 7). This association is due partly toa loss of gastric glands (atrophy). However,H. pylori treatment increases acid productionrapidly in some people with pan-gastritis andhypochlorhydria (although not usually to nor-mal levels) (196). This result implies a di-rect suppressive effect of H. pylori–induced in-flammatory mediators or products. IL-1β is aknown acid suppressant; it, TNFα (197), and,in a recent report, the Th-1 cytokines IFN-γand IL-12 (198) suppress acid production inisolated rabbit parietal cells. Acid suppressionby auto-antibodies against the proton pump,increased cytokine production in the gastriccorpus, or a combination of these factors fur-ther increase hypergastrinemia and so may ac-
celerate development of gastric atrophy andintestinal metaplasia. Proton pump inhibitors(PPIs), drugs used for the profound suppres-sion of acid, both induce a pan-gastritic orcorpus-predominant pattern of inflammationand speed atrophy and intestinal metaplasiawhen used in H. pylori–infected people (199).However, there is no evidence so far that use ofthese drugs has increased gastric cancer inci-dence. Once acid production is reduced, otherbacteria may colonize the stomach and causefurther inflammation that may contribute tohypergastrinemia; interestingly, in a mousemodel, PPI-induced hypergastrinemia couldbe reversed fully by treatment with antibiotics(200).
Adenocarcinoma can arise in the atrophicstomach regardless of whether the atrophy isinduced by H. pylori or by autoimmune mech-anisms. This implies that the role of H. py-lori is principally in inducing atrophy, which isthen in itself a precancerous state. This is sup-ported by a large H. pylori eradication study,in which H. pylori eradication appeared to pre-vent later cancer development in patients thatdid not have atrophy and intestinal metapla-sia, but not in patients who already had thesepremalignant conditions (201). Also, H. pyloridensity is much reduced in the atrophic stom-ach, so direct H. pylori–induced effects thereshould also be reduced.
Carcinogenesis in the environment of gas-tric atrophy is likely owing to reactive oxygenand nitrogen species secondary to the accom-panying inflammation, and possibly partlycaused by overgrowth of other bacteria in thenow achlorhydric stomach (200). In a mousemodel, mutations in epithelial cell DNA arecharacteristic of such oxidative damage (202),and inducible nitric oxide synthase–deficientmice are less likely to develop cancer follow-ing H. pylori infection and chemical carcino-gen challenge than normal mice (203). Also,chemoprevention trials using antioxidants inpatients with atrophy or intestinal metapla-sia show some regression of these conditions(204), and high ascorbic acid levels, whichare antioxidant, are associated with lower
76 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
levels of gastric cancer development in hu-mans (205).
Other mechanisms of carcinogenesis havealso been either demonstrated or impliedin the atrophic stomach. Parietal cells ex-press the Sonic hedgehog protein, which isthe main organizer of cell polarity in gas-tric pits. Sonic hedgehog expression is lost inatrophy and intestinal metaplasia (206), andmice engineered to have parietal cell ablationdevelop epithelial disorganisation and ampli-fication of epithelial progenitors. This disrup-tion potentially exposes stem cells to muta-genic agents (207). The intestinal metaplasiathat develops in the atrophic stomach com-prises a widespread metaplastic cell lineagein both rodents and man and can be rec-ognized by its expression of Trefoil factor 2(208). The contention that carcinoma devel-ops directly from intestinal metaplasia is sup-ported by a transgenic mouse model express-ing the Cdx-2 protein. These mice developextensive intestinal metaplasia in the stom-ach and malignant changes within the intesti-nal metaplastic epithelium (209). A startlingand important recent finding is that the meta-plastic and neoplastic cells arising in thestomach of the H. pylori–infected mouse ap-pear to be derived from bone marrow; thisfinding has important implications for car-cinogenesis in the stomach and elsewhere(210).
Although cancer can arise in the atrophicstomach without H. pylori, there is some evi-dence that direct interaction of H. pylori withthe epithelium may confer an additive or al-ternative risk. Firstly, the less common dif-fuse type of gastric adenocarcinoma, which isnot addressed in the studies mentioned aboveand often affects younger adults (211), canarise directly in inflamed nonatrophic mu-cosa. Secondly, in animal models such as thehypergastrinemic INS-GAS mouse, carcino-genesis is speeded by helicobacter infection.Thirdly, human trials have shown that H. py-lori treatment in patients with atrophy and in-testinal metaplasia may slow the progressionof changes, if not halt them (204, 212). Finally,
HELICOBACTER PYLORI AND GASTRICLYMPHOMA
The uninfected stomach has no mucosa-associated lymphoidtissue (MALT), so its acquisition depends upon infection withhelicobacter species, usually H. pylori. The risk factors for itstransformation into low-grade extranodal B cell lymphoma ofMALT (MALT lymphoma) are poorly defined: cag+ strains donot increase risk markedly (237–240), but low-grade MALTlymphomas do usually arise in the context of a pan-gastriticinflammation pattern (241). In vitro, proliferation of clonalB cells is driven by the presence of non-neoplastic T cells,whose effects in turn are dependant upon the presence ofH. pylori (242). H. pylori also appears to act through directantigen stimulation of tumor cells (243). In humans, remark-ably, eradication of H. pylori frequently leads to regressionof these low-grade lymphomas (244, 245). The natural his-tory of MALT lymphomas is that they are only very slowlyprogressive, but they sometimes undergo high-grade trans-formation to diffuse large-cell B cell lymphomas. These lym-phomas have mutations consistent with oxidative damage andare more commonly associated with cag+ H. pylori strain in-fection and a more inflamed stomach (238, 239). High-gradelymphomas rarely regress following H. pylori treatment. Re-cently, a subgroup of low-grade B cell MALT lymphomas (ap-proximately 25%) have been found to have a translocationfrom chromosome 11 to 18 (246, 247). This causes a specificfusion between the activator protein-12 (AP-12) and MALT-1genes that creates a new gene (248) whose product stimulatesNFκB signaling (249). This is known to be anti-apoptoticand so promotes cell survival. T(11;18)+ lymphomas are rela-tively locally invasive but rarely undergo high-grade trans-formation; most importantly, they are unresponsive to H.pylori treatment (250, 251). Like t(11;18)+ lymphomas atother sites in the body, these lymphomas arise in associa-tion with a neutrophil infiltrate, and, as expected from thisfact, they are usually associated with cag+ H. pylori infection(252).
recent reports suggest that H. pylori may sur-vive and express virulence factors intracellu-larly in metaplastic, dysplastic, and neoplasticepithelial cells in vivo, although these reportsneed further confirmation (213).
Following the theory that direct onco-genic effects of H. pylori do contribute to
www.annualreviews.org • Helicobacter pylori–Induced Diseases 77
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
carcinogenesis in vivo, several potential mech-anisms exist. As described earlier, CagA sig-naling is strongly pro-proliferative, and thiseffect may not be balanced by increased apop-tosis: Although H. pylori has proapoptotic ef-fects on cells (214, 215), it also has anti-apoptotic effects including NFκB activation,reduction in p27 levels (216), and downregu-lation of the TNF-related apoptosis-inducingligand (TRAIL) system (217). Whether pro-liferation and anti-apoptotic effects have a di-rect role in epithelial transformation is un-clear, but they do appear important in thepathogenesis of gastric atrophy: Fas-deficientmice, which have impaired apoptosis, developextensive and severe atrophy (217). Other di-rect effects of H. pylori on gastric epithelialcells that have potential carcinogenic impor-tance are its stimulation of reactive oxygenspecies (ROS) and oxidative damage (218)and its impairment of DNA mismatch repair(219). However, H. pylori also downregulatesthe oxidative damage it causes by inducing en-zymes that scavenge ROS (220), and the hostminimizes ROS-induced DNA damage by ex-pressing the oxidative repair proteins Ape-1 and Rel-1 (221). That H. pylori needs toprotect itself against oxidative damage sug-gests that the generation of ROS may be ahost defense system for inhibiting H. pylorimetabolism or otherwise damaging H. pylori(222, 223).
Protection by Helicobacter pylori againstsevere gastro-esophageal reflux and itscomplications. The incidence of gastro-esophageal reflux disease (GERD) and itscomplications, Barrett’s esophagus (intestinalmetaplasia in the esophagus), and esophagealadenocarcinoma, is rising rapidly in devel-oped countries. Over the same time period,the incidences of gastric adenocarcinoma andpeptic ulceration are falling, mimicking thefalling prevalence of H. pylori infection (224).Epidemiological observations support a linkbetween H. pylori and protection against se-vere GERD. Reflux esophagitis is rare incountries where H. pylori infection is com-
mon, such as China and Japan. Many associa-tion studies have shown that people with com-plicated reflux disease, including esophagealadenocarcinoma, are less likely than othersto have H. pylori infection (225, 226). Morerecently, a case control study nested in acohort established in 1964 has shown thatH. pylori–positive subjects are less likelythan H. pylori–negative subjects to developesophageal adenocarcinoma (odds ratio 0.37)(227).
If the negative association between H. py-lori and GERD complications is causal, themost likely mechanism is through effects ofH. pylori on gastric acid secretion. Althoughsome H. pylori–infected individuals have in-creased acid secretion, a reduction is morecommon, and thus, at a population level, ab-sence of H. pylori should lead to higher meanacid secretion. In patients with GERD, therefluxate would on average be more acidic,which would increase damage caused andthereby increase the population risk of com-plications, including esophageal adenocarci-noma. Support for this idea comes fromstudies showing that cag+ H. pylori infec-tion, which has more profound effects onacid secretion, is associated with less severeesophagitis and a larger reduction in bothBarrett’s esophagus and esophageal cancerrisk (228–230) [although not all reports con-firm this observation (227)]. Further supportcomes from the greater reduction in risk seenin people with corpus-predominant gastritis,atrophy, intestinal metaplasia, and low acid se-cretion (225, 231, 232). Evidence from inter-vention studies is less clearcut; after H. py-lori treatment, some people show a reduc-tion, some show no effect, and some showan increase in reflux symptoms and GERDendoscopic severity. However, such hetero-geneity between studies may be predicted, aseffects of H. pylori treatment on acid secretionwould be expected to differ between individu-als and populations depending upon pretreat-ment gastric inflammation pattern, acid secre-tion, parietal cell mass, and presence and levelof gastric atrophy.
78 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
HELICOBACTER PYLORI/HUMANCOEVOLUTION:IMPLICATIONS FOR DISEASEAND ITS MANAGEMENT
H. pylori has colonized humans for at least2500 years, and likely since we first evolved(7, 37); other mammals have their own spe-cialized indigenous gastric helicobacters, im-plying differential evolution of these bacteriato suit different niches. H. pylori has adaptedto persist, and this adaptation involved ma-nipulation of both human acquired immunityand gastric physiology. Human gastric physi-ology has also likely evolved to coexist with H.pylori. In many developing countries (proba-bly representing the history of humanity) H.pylori is ubiquitous and gastric acid secretionis, on average, reduced. One could regard theacid production in the modern H. pylori–freestomach as abnormally high, or at least newfor humans. This could contribute to compli-cations of GERD.
That humans have not evolved to clearH. pylori infection has led to speculation thatthe bacterium may confer some benefit. If so,this would most likely be in childhood, as thisis when H. pylori is usually acquired. Severalstudies have addressed whether H. pylori mayprotect against other infections, and a nega-tive association has been shown with gastroen-teritis (233). Mechanisms and even causalityare unclear, but an intriguing possibility isthat an increased acid barrier in some peoplemay protect against bacterial infections. As inthe INS-GAS hypergastrinemic mouse (234),perhaps H. pylori+ children in previous gen-erations had increased acid production, whichprotected them from infectious disease; re-duced acid secretion and its complications inpostreproductive adulthood would be evolu-tionarily neutral.
Whether H. pylori confers evolutionarybenefit to humans, it is unlikely to conferevolutionary disadvantages. Gastric adeno-carcinoma occurs after human reproductiveyears and DU is a disease of the late nineteenthand twentieth centuries (235, 236). Why
duodenal ulcers arose at this time is unclear.However, as H. pylori infection is likely tohave been ubiquitous before this point, and H.pylori virulence and human genetics are un-likely to have changed during the timescale,environmental changes are the mostlikely cause. One possibility is that themodern stomach produces more acid becauseit is healthier, possibly owing to increasednutrition, lack of gastric irritants, or perhapsH. pylori infection at a later age. The in-crease in acid production may subsequentlypredispose humans to antral-predominantinfection/inflammation and DU. The adventof smoking may also have been a contributoryfactor.
The future burden of H. pylori–associateddiseases is unclear. In many developed coun-tries H. pylori prevalence is falling. This re-duction is mirrored by a fall in incidence ofpeptic ulceration and gastric adenocarcinomawith a concomitant rise in reflux esophagitisand its complications, including esophagealadenocarcinoma (224). However, on the ba-sis of current trends, esophageal adenocarci-noma is unlikely to approach previous lev-els of gastric adenocarcinoma in mankind’s H.pylori–ubiquitous history. In developing coun-tries, H. pylori–associated diseases remain ma-jor medical problems, and worldwide, the bur-den of these problems is increasing becauseof an aging world population. With mod-ernization, H. pylori prevalence in developingcountries is likely to decrease eventually, butwhether, before that, we will see an increasein ulcer incidence as occurred in developedcountries in the past century is unclear. Vac-cination is the main hope for rapid interven-tion, and perhaps prevention of the consider-able morbidity from peptic ulceration and thenearly one million deaths per year from gas-tric cancer. Vaccine research is ongoing, butan effective vaccine has still not been devel-oped. Preventing H. pylori infection is likelyto do more good than harm, but research isneeded before intervention to ensure that thisis the case.
www.annualreviews.org • Helicobacter pylori–Induced Diseases 79
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
SUMMARY POINTS
1. H. pylori infection causes peptic ulceration, gastric adenocarcinoma, and gastric lym-phoma. It may protect against complications of gastro-esophageal reflux disease.
2. Pathogenesis depends upon persistent infection over decades.
3. Which infections result in disease depends upon bacterial virulence, host geneticsusceptibility, and environmental factors.
4. Strains possessing the cag PAI are more pro-inflammatory, more pro-proliferative,and more likely to cause disease.
5. The vacuolating cytotoxin VacA causes epithelial damage, and H. pylori strains ex-pressing more active forms of VacA are more likely to cause disease.
6. Host proinflammatory cytokine polymorphisms predispose H. pylori–infected peopleto gastric adenocarcinoma.
7. H. pylori induces gastric atrophy, which may progress to adenocarcinoma even ifH. pylori is treated.
8. H. pylori and humans have co-evolved and co-adapted. The absence of H. pylori changeshuman gastric physiology.
FUTURE DIRECTIONS/UNRESOLVED ISSUES
1. How does H. pylori persist throughout human life?
2. What are the mechanisms underlying gastric carcinogenesis?
3. Can an effective human vaccine be developed?
4. Are there adverse consequences for an H. pylori–free human population?
RELATED RESOURCES
1. Atherton JC. 2004. Understanding Helicobacter pylori. In: Molecular Insights Into Gas-troenterology, ed. D Adams, TT MacDonald, pp. 51–60. London: BMJ PublishingGroup
2. Isaacson PG, Du MQ. 2004. Timeline - MALT lymphoma: from morphology tomolecules. Nat. Rev. Cancer 4(8):644–53
3. Merrell DS, Falkow S. 2004. Frontal and stealth attack strategies in microbial patho-genesis. Nature 430(6996):250–56
4. Peek RM Jr, Blaser MJ. 1999. Helicobacter pylori strain-specific genotypes and modu-lation of the gastric epithelial cell cycle. Cancer Res. 59(24):6124–31
ACKNOWLEDGMENTS
I would like to thank Miss Emma Bradley for all her help with the preparation of this manuscript.
80 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
LITERATURE CITED1. Franchini M, Veneri D. 2004. Helicobacter pylori infection and immune thrombocy-
topenic purpura: an update. Helicobacter 9:342–462. Jackson S, Beck PL, Pineo GF, Poon MC. 2005. Helicobacter pylori eradication: novel
therapy for immune thrombocytopenic purpura? A review of the literature. Am. J. Hematol.78:142–50
3. Pellicano R, Fagoonee S, Rizzetto M, Ponzetto A. 2003. Helicobacter pylori and coronaryheart disease: Which directions for future studies? Crit. Rev. Microbiol. 29:351–59
4. Gasbarrini A, Carloni E, Gasbarrini G, Chisholm SA. 2004. Helicobacter pylori andextragastric diseases - other Helicobacters. Helicobacter 9:57–66
5. Morris A, Nicholson G. 1987. Ingestion of Campylobacter-Pyloridis causes gastritis andraised fasting gastric Ph. Am. J. Gastroenterol. 82:192–99
6. Graham DY, Alpert LC, Smith JL, Yoshimura HH. 1988. Iatrogenic Campylobacter-Pylori infection is a cause of epidemic achlorhydria. Am. J. Gastroenterol. 83:974–80
7. Blaser MJ, Atherton JC. 2004. Helicobacter pylori persistence: biology and disease. J.Clin. Invest. 113:321–33
Publication of thecomplete genomesequence ofH. pylori providedimmediate insightsinto its biology andhas revolutionizedresearch methods.
8. Tomb JF, White O, Kerlavage AR, Clayton RA, Sutton GG, et al. 1997. Thecomplete genome sequence of the gastric pathogen Helicobacter pylori. Nature
388:539–479. Alm RA, Ling LSL, Moir DT, King BL, Brown ED, et al. 1999. Genomic-sequence
comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori.Nature 397:176–80
10. Kavermann H, Burns BP, Angermuller K, Odenbreit S, Fischer W, et al. 2003. Identifi-cation and characterization of Helicobacter pylori genes essential for gastric colonization.J. Exp. Med. 197:813–22
11. Terry K, Williams SM, Connolly L, Ottemann KM. 2005. Chemotaxis plays multipleroles during Helicobacter pylori animal infection. Infect. Immun. 73:803–11
12. Waidner B, Greiner S, Odenbreit S, Kavermann H, Velayudhan J, et al. 2002. Essentialrole of ferritin Pfr in Helicobacter pylori iron metabolism and gastric colonization. Infect.Immun. 70:3923–29
13. Wen Y, Marcus EA, Matrubutham U, Gleeson MA, Scott DR, et al. 2003. Acid-adaptivegenes of Helicobacter pylori. Infect. Immun. 71:5921–39
14. McGowan CC, Necheva AS, Forsyth MH, Cover TL, Blaser MJ. 2003. Promoter analysisof Helicobacter pylori genes with enhanced expression at low pH. Mol. Microbiol. 48:1225–39
Explained howH. pylori couldsurvive sudden acidexposure in thestomach.
15. Weeks DL, Eskandari S, Scott DR, Sachs G. 2000. A H+-gated urea channel: thelink between Helicobacter pylori urease and gastric colonization. Science 287:482–85
16. Atherton JC, Blaser MJ. 2004. Chapter 154: Helicobacter pylori infections. In Harrison’sPrincipals of Internal Medicine, ed. DL Kasper, E Braunwald, AS Fauci, SL HAuser, DLLongo, JL Jameson, pp. 886–89 . New York: McGraw-Hill. 16th ed.
17. Clyne M, Dillon P, Daly S, O’Kennedy R, May FEB, et al. 2004. Helicobacter pyloriinteracts with the human single-domain trefoil protein TFF1. Proc. Natl. Acad. Sci. USA101:7409–14
18. Kawakubo M, Ito Y, Okimura Y, Kobayashi M, Sakura K, et al. 2004. Natural antibioticfunction of a human gastric mucin against Helicobacter pylori infection. Science 305:1003–6
www.annualreviews.org • Helicobacter pylori–Induced Diseases 81
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
19. Schreiber S, Konradt M, Groll C, Scheid P, Hanauer G, et al. 2004. The spatial orien-tation of Helicobacter pylori in the gastric mucus. Proc. Natl. Acad. Sci. USA 101:5024–29
20. Kim N, Marcus EA, Wen Y, Weeks DL, Scott DR, et al. 2004. Genes of Helicobacterpylori regulated by attachment to AGS cells. Infect. Immun. 72:2358–68
21. Kwok T, Backert S, Schwarz H, Berger J, Meyer TF. 2002. Specific entry of Helicobacterpylori into cultured gastric epithelial cells via a zipper-like mechanism. Infect. Immun.70:2108–20
22. Amieva MR, Salama NR, Tompkins LS, Falkow S. 2002. Helicobacter pylori enter andsurvive within multivesicular vacuoles of epithelial cells. Cell. Microbiol. 4:677–90
23. Rittig MG, Shaw B, Letley DP, Thomas RJ, Argent RH, et al. 2003. Helicobacter pylori-induced homotypic phagosome fusion in human monocytes is independent of the bacterialvacA and cag status. Cell. Microbiol. 5:887–99
24. Odenbreit S, Gebert B, Puls J, Fischer W, Haas R. 2001. Interaction of Helicobacterpylori with professional phagocytes: role of the cag pathogenicity island and translocation,phosphorylation and processing of CagA. Cell. Microbiol. 3:21–31
25. Ohta T, Shibata H, Kawamori T, Iimuro M, Sugimura T, et al. 2001. Marked reductionof Helicobacter pylori-induced gastritis by urease inhibitors, acetohydroxamic acid andflurofamide, in Mongolian gerbils. Biochem. Biophys. Res. Commun. 285:728–33
26. Harris PR, Ernst PB, Kawabata S, Kiyono H, Graham MF, et al. 1998. Recombinant He-licobacter pylori urease activates primary mucosal macrophages. J. Infect. Dis. 178:1516–20
27. Satin B, Del Giudice G, Della Bianca V, Dusi S, Laudanna C, et al. 2000. The neutrophil-activating protein (HP-NAP) of Helicobacter pylori is a protective antigen and a majorvirulence factor. J. Exp. Med. 191:1467–76
28. Tonello F, Dundon WG, Satin B, Molinari M, Tognon G, et al. 1999. The Helicobacterpylori neutrophil-activating protein is an iron-binding protein with dodecameric struc-ture. Mol. Microbiol. 34:238–46
29. Backhed F, Rokbi B, Torstensson E, Zhao Y, Nilsson C, et al. 2003. Gastric mucosalrecognition of Helicobacter pylori is independent of Toll-like receptor 4. J. Infect. Dis.187:829–36
30. Smith MF, Mitchell A, Li GL, Ding S, Fitzmaurice AM, et al. 2003. Toll-like receptor(TLR) 2 and TLR5, but not TLR4, are required for Helicobacter pylori-induced NF-kappa B activation and chemokine expression by epithelial cells. J. Biol. Chem. 278:32552–60
31. Gewirtz AT, Yu Y, Krishna US, Israel DA, Lyons SL, et al. 2004. Helicobacter pyloriflagellin evades toll-like receptor 5-mediated innate immunity. J. Infect. Dis. 189:1914–20
32. Gobert AP, McGee D, Akhtar M, Mendz GL, Newton JC, et al. 2001. Helicobacterpylori arginase inhibits nitric oxide production by eukaryotic cells: a strategy for bacterialsurvival. Gut 49:429
33. Schmausser B, Andrulis M, Endrich S, Lee SK, Josenhans C, et al. 2004. Expressionand subcellular distribution of toll-like receptors TLR4, TLR5 and TLR9 on the gastricepithelium in Helicobacter pylori infection. Clin. Exp. Immunol. 136:521–26
34. Alamuri P, Maier RJ. 2004. Methionine sulphoxide reductase is an important antioxidantenzyme in the gastric pathogen Helicobacter pylori. Mol. Microbiol. 53:1397–406
Shows that H.
pylori strainsrecombine morefrequently than anyother bacterialspecies, allowingmaintenance ofenormous geneticdiversity. 35. Suerbaum S, Smith JM, Bapumia K, Morelli G, Smith NH, et al. 1998. Free recom-
bination within Helicobacter pylori. Proc. Natl. Acad. Sci. USA 95:12619–24
82 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
36. Israel DA, Salama N, Krishna U, Rieger UM, Atherton JC, et al. 2001. Helicobacterpylori genetic diversity within the gastric niche of a single human host. Proc. Natl. Acad.Sci. USA 98:14625–30
37. Falush D, Kraft C, Taylor NS, Correa P, Fox JG, et al. 2001. Recombination and mutationduring long-term gastric colonization by Helicobacter pylori: estimates of clock rates,recombination size, and minimal age. Proc. Natl. Acad. Sci. USA 98:15056–61
38. Aspholm-Hurtig M, Dailide G, Lahmann M, Kalia A, Ilver D, et al. 2004. Functionaladaptation of BabA, the H. pylori ABO blood group antigen binding adhesin. Science305:519–22
39. Robinson K, Loughlin MF, Potter R, Jenks PJ. 2005. Host adaptation and immune mod-ulation are mediated by homologous recombination in Helicobacter pylori. J. Infect. Dis.191:579–87
40. Hansson LE, Nyren O, Hsing AW, Bergstrom R, Josefsson S, et al. 1996. The risk ofstomach cancer in patients with gastric or duodenal ulcer disease. New Engl. J. Med.335:242–49
Characterized thecag pathogenicityisland, nowrecognized as themost importantH. pylori virulencefactor.
41. Censini S, Lange C, Xiang ZY, Crabtree JE, Ghiara P, et al. 1996. cag, a pathogenic-ity island of Helicobacter pylori, encodes type I-specific and disease-associatedvirulence factors. Proc. Natl. Acad. Sci. USA 93:14648–53
42. Nilsson C, Sillen A, Eriksson L, Strand ML, Enroth H, et al. 2003. Correlation betweencag pathogenicity island composition and Helicobacter pylori-associated gastroduodenaldisease. Infect. Immun. 71:6573–81
43. Crabtree JE, Taylor JD, Wyatt JI, Heatley RV, Shallcross TM, et al. 1991. MucosalIga recognition of Helicobacter pylori 120-kDa protein, peptic ulceration, and gastricpathology. Lancet 338:332–35
44. Nomura AMY, Perez-Perez GI, Lee J, Stemmermann G, Blaser MJ. 2002. Relation be-tween Helicobacter pylori cagA status and risk of peptic ulcer disease. Am. J. Epidemiol.155:1054–59
45. Blaser MJ, Perezperez GI, Kleanthous H, Cover TL, Peek RM, et al. 1995. Infectionwith Helicobacter pylori strains possessing caga is associated with an increased risk ofdeveloping adenocarcinoma of the stomach. Cancer Res. 55:2111–15
46. Figueiredo C, Machado JC, Pharoah P, Seruca R, Sousa S, et al. 2002. Helicobacter pyloriand interleukin 1 genotyping: an opportunity to identify high-risk individuals for gastriccarcinoma. J. Natl. Cancer Inst. 94:1680–87
47. Rohde M, Puls J, Buhrdorf R, Fischer W, Haas R. 2003. A novel sheathed surface organelleof the Helicobacter pylori cag type IV secretion system. Mol. Microbiol. 49:219–34
48. Tanaka J, Suzuki T, Mimuro H, Sasakawa C. 2003. Structural definition on the surface ofHelicobacter pylori type IV secretion apparatus. Cell. Microbiol. 5:395–404
49. Segal ED, Cha J, Lo J, Falkow S, Tompkins LS. 1999. Altered states: involvement ofphosphorylated CagA in the induction of host cellular growth changes by Helicobacterpylori. Proc. Natl. Acad. Sci. USA 96:14559–64
50. Asahi M, Azuma T, Ito S, Ito Y, Suto H, et al. 2000. Helicobacter pylori CagA proteincan be tyrosine phosphorylated in gastric epithelial cells. J. Exp. Med. 191:593–602 Shows how the
CagA protein istranslocated intoepithelial cellswhere it stimulatesspecific signalingpathways.
51. Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, et al. 2000. Translocationof Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science
287:1497–50052. Stein M, Rappuoli R, Covacci A. 2000. Tyrosine phosphorylation of the Helicobacter
pylori CagA antigen after cag-driven host cell translocation. Proc. Natl. Acad. Sci. USA97:1263–68
www.annualreviews.org • Helicobacter pylori–Induced Diseases 83
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
53. Stein M, Bagnoli F, Halenbeck R, Rappuoli R, Fantl WJ, et al. 2002. c-Src/Lyn kinasesactivate Helicobacter pylori CagA through tyrosine phosphorylation of the EPIYA motifs.Mol. Microbiol. 43:971–80
54. Selbach M, Moese S, Hauck CR, Meyer TF, Backert S. 2002. Src is the kinase ofthe Helicobacter pylori CagA protein in vitro and in vivo. J. Biol. Chem. 277:6775–78
55. Amieva MR, Vogelmann R, Covacci A, Tompkins LS, Nelson WJ, et al. 2003. Disruptionof the epithelial apical-junctional complex by Helicobacter pylori CagA. Science 300:1430–34
56. Churin Y, Al-Ghoul L, Kepp O, Meyer TE, Birchmeier W, et al. 2003. Helicobacterpylori CagA protein targets the c-Met receptor and enhances the motogenic response. J.Cell. Biol. 161:249–55
57. El-Etr SH, Mueller A, Tompkins LS, Falkow S, Merrell DS. 2004. Phosphorylation-independent effects of CagA during interaction between Helicobacter pylori and T84polarized monolayers. J. Infect. Dis. 190:1516–23
58. Higashi H, Nakaya A, Tsutsumi R, Yokoyama K, Fujii Y, et al. 2004. Helicobacter pyloriCagA induces Ras-independent morphogenetic response through SHP-2 recruitment andactivation. J. Biol. Chem. 279:17205–16
59. Higashi H, Tsutsumi R, Fujita A, Yamazaki S, Asaka M, et al. 2002. Biological activity ofthe Helicobacter pylori virulence factor CagA is determined by variation in the tyrosinephosphorylation sites. Proc. Natl. Acad. Sci. USA 99:14428–33
60. Hirata Y, Maeda S, Mitsuno Y, Tateishi K, Yanai A, et al. 2002. Helicobacter pylori CagAprotein activates serum response element-driven transcription independently of tyrosinephosphorylation. Gastroenterology 123:1962–71
61. Keates S, Sougioultzis S, Keates AC, Zhao DZ, Peek RM, et al. 2001. cag plus Helicobacterpylori induce transactivation of the epidermal growth factor receptor in AGS gastricepithelial cells. J. Biol. Chem. 276:48127–34
62. Meyer-ter-Vehn T, Covacci A, Kist M, Pahl HL. 2000. Helicobacter pylori activatesmitogen-activated protein kinase cascades and induces expression of the proto-oncogenesc-fos and c-jun. J. Biol. Chem. 275:16064 –72
63. Mitsuno Y, Maeda S, Yoshida H, Hirata Y, Ogura K, et al. 2002. Helicobacter pyloriactivates the proto-oncogene c-fos through SRE transactivation. Biochem. Biophys. Res.Commun. 291:868–74
64. Mimuro H, Suzuki T, Tanaka J, Asahi M, Haas R, et al. 2002. Grb2 is a key mediator ofHelicobacter pylori CagA protein activities. Mol. Cell. 10:745–55
65. Naumann M, Wessler S, Bartsch C, Wieland B, Covacci A, et al. 1999. Activationof activator protein 1 and stress response kinases in epithelial cells colonized by He-licobacter pylori encoding the cag pathogenicity island. J. Biol. Chem. 274:31655–62
66. Selbach M, Moese S, Meyer TF, Backert S. 2002. Functional analysis of the Helicobacterpylori cag pathogenicity island reveals both VirD4-CagA-dependent and VirD4-CagA-independent mechanisms. Infect. Immun. 70:665–71
67. Tsutsumi R, Higashi H, Higuchi M, Okada M, Hatakeyama M. 2003. Attenuation ofHelicobacter pylori CagA-SHP-2 signaling by interaction betwwen CagA and C-terminalSrc kinase. J. Biol. Chem. 278:3664–70
68. Yamazaki S, Yamakawa A, Ito Y, Ohtani M, Higashi H, et al. 2003. The CagA proteinof Helicobacter pylori is translocated into epithelial cells and binds to SHP-2 in humangastric mucosa. J. Infect. Dis. 187:334–37
84 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
69. Moese S, Selbach M, Kwok T, Brinkmann V, Konig W, et al. 2004. Helicobacter py-lori induces AGS cell motility and elongation via independent signaling pathways. Infect.Immun. 72:3646–9
70. Bebb JR, Letley DP, Rhead JL, Atherton JC. 2003. Helicobacter pylori supernatants causeepithelial cytoskeletal disruption that is bacterial strain and epithelial cell line dependentbut not toxin VacA dependent. Infect. Immun. 71:3623–27
71. Segal ED, Falkow S, Tompkins LS. 1996. Helicobacter pylori attachment to gastric cellsinduces cytoskeletal rearrangements and tyrosine phosphorylation of host cell proteins.Proc. Natl. Acad. Sci. USA 93:1259–64
72. Moss SF, Sordillo EM, Abdalla AM, Makarov V, Hanzely Z, et al. 2001. Increased gastricepithelial cell apoptosis associated with colonization with cagA plus Helicobacter pyloristrains. Cancer Res. 61:1406–11
73. Peek RM Jr, Moss SF, Tham KT, Perez-Perez GI, Wang S, et al. 1997. Helicobacterpylori cagA+ strains and dissociation of gastric epithelial cell proliferation from apoptosis.J. Natl. Cancer Inst. 89:863–68
74. Argent RH, Kidd M, Owen RJ, Thomas RJ, Limb MC, et al. 2004. Determinants and con-sequences of different levels of CagA phosphorylation for clinical isolates of Helicobacterpylori. Gastroenterology 127:514–23
75. Yamaoka Y, El-Zimaity HM, Gutierrez O, Figura N, Kim JG, et al. 1999. Relationship be-tween the cagA 3’ repeat region of Helicobacter pylori, gastric histology, and susceptibilityto low pH. Gastroenterology 117:342–49
76. Azuma T, Yamakawa A, Yamazaki S, Fukuta K, Ohtani M, et al. 2002. Correlation betweenvariation of the 3’ region of the cagA gene in Helicobacter pylori and disease outcome inJapan. J. Infect. Dis. 186:1621–30
77. Azuma T, Yamazaki S, Yamakawa A, Ohtani M, Muramatsu A, et al. 2004. Associationbetween diversity in the Src homology 2 domain-containing tyrosine phosphatase bindingsite of Helicobacter pylori CagA protein and gastric atrophy and cancer. J. Infect. Dis.189:820–27
78. Fischer W, Puls J, Buhrdorf R, Gebert B, Odenbreit S, et al. 2001. Systematic muta-genesis of the Helicobacter pylori cag pathogenicity island: essential genes for CagAtranslocation in host cells and induction of interleukin-8. Mol. Microbiol. 42:1337–48
79. Viala J, Chaput C, Boneca IG, Cardona A, Girardin SE, et al. 2004. Nod1 responds topeptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat. Immun.5:1166–74
80. Maeda S, Yoshida H, Ogura K, Mitsuno Y, Hirata Y, et al. 2000. H. pylori activatesNF-kappa B through a signaling pathway involving I kappa B kinases, NF-kappa B-inducing kinase, TRAF2, and TRAF6 in gastric cancer cells. Gastroenterology 119:97–108
81. Mori N, Sato H, Hayashibara T, Senba M, Geleziunas R, et al. 2003. Helicobacter pyloriinduces matrix metalloproteinase-9 through activation of nuclear factor kappaB. Gastroen-terology 124:983–92
82. Wada A, Ogushi K, Kimura T, Hojo H, Mori N, et al. 2001. Helicobacter pylori-mediatedtranscriptional regulation of the human beta-defensin 2 gene requires NF-kappa B. Cell.Microbiol. 3:115–23
83. Yanai A, Hirata Y, Mitsuno Y, Maeda S, Shibata W, et al. 2003. Helicobacter pyloriinduces antiapoptosis through nuclear factor-kappa B activation. J. Infect. Dis. 188:1741–51
www.annualreviews.org • Helicobacter pylori–Induced Diseases 85
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
84. Bebb JR, Letley DP, Thomas RJ, Aviles F, Collins HM, et al. 2003. Helicobacter pyloriupregulates matrilysin (MMP-7) in epithelial cells in vivo and in vitro in a Cag dependentmanner. Gut 52:1408–13
85. Ogura K, Maeda S, Nakao M, Watanabe T, Tada M, et al. 2000. Virulence factors ofHelicobacter pylori responsible for gastric diseases in Mongolian gerbil. J. Exp. Med.192:1601–10
86. Kersulyte D, Chalkauskas H, Berg DE. 1999. Emergence of recombinant strains of He-licobacter pylori during human infection. Mol. Microbiol. 31:31–43
87. Aras RA, Lee Y, Kim SK, Israel D, Peek RM, et al. 2003. Natural variation in populationsof persistently colonizing bacteria affect human host cell phenotype. J. Infect. Dis. 188:486–96
88. Cover TL, Tummuru MK, Cao P, Thompson SA, Blaser MJ. 1994. Divergence of geneticsequences for the vacuolating cytotoxin among Helicobacter pylori strains. J. Biol. Chem.269:10566–73
Describes geneticvariation in thevacuolatingcytotoxin gene,vacA; this underliesdifferential toxicityof H. pylori strains.
89. Atherton JC, Cao P, Peek RM, Tummuru MKR, Blaser MJ, et al. 1995. Mosaicismin vacuolating cytotoxin alleles of Helicobacter pylori—association of specific vacAtypes with cytotoxin production and peptic ulceration. J. Biol. Chem. 270:17771–77
90. Letley DP, Lastovica A, Louw JA, Hawkey CJ, Atherton JC. 1999. Allelic diversity ofthe Helicobacter pylori vacuolating cytotoxin gene in South Africa: rarity of the vacAs1a genotype and natural occurrence of an s2/m1 allele. J. Clin. Microbiol. 37:1203–5
91. Letley DP, Rhead JL, Twells RJ, Dove B, Atherton JC. 2003. Determinants of non-toxicityin the gastric pathogen Helicobacter pylori. J. Biol. Chem. 278:26734–41
92. Pagliaccia C, de Bernard M, Lupetti P, Ji X, Burroni D, et al. 1998. The m2 form of theHelicobacter pylori cytotoxin has cell type-specific vacuolating activity. Proc. Natl. Acad.Sci. USA 95:10212–17
93. Atherton JC, Cao P, Peek RM Jr, Tummuru MK, Blaser MJ, et al. 1995. Mosaicism invacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types withcytotoxin production and peptic ulceration. J. Biol. Chem. 270:17771–77
94. Atherton JC, Peek RM Jr, Tham KT, Cover TL, Blaser MJ. 1997. Clinical and patholog-ical importance of heterogeneity in vacA, the vacuolating cytotoxin gene of Helicobacterpylori. Gastroenterology 112:92–99
95. Atherton JC, Cover TL, Papini E, Telford JL. 2001. Vacuolating cytotoxin. In Helicobac-ter Pylori: Physiology and Genetics, ed. HLT Mobley, GL Mendz, SL Hazell, pp. 97–110.Washington: ASM Press.
96. van Doorn LJ, Figueiredo C, Sanna R, Pena S, Midolo P, et al. 1998. Expanding allelicdiversity of Helicobacter pylori vacA. J. Clin. Microbiol. 36:2597–603
97. Kidd M, Lastovica AJ, Atherton JC, Louw JA. 1999. Heterogeneity in the Helicobacterpylori vacA and cagA genes: association with gastroduodenal disease in South Africa? Gut45:499–502
98. Miehlke S, Kirsch C, Agha-Amiri K, Gunther T, Lehn N, et al. 2000. The Helicobacterpylori vacA s1, m1 genotype and cagA is associated with gastric carcinoma in Germany.Int. J. Cancer 87:322–27
99. Figueiredo C, Van Doorn LJ, Nogueira C, Soares JM, Pinho C, et al. 2001. Helicobacterpylori genotypes are associated with clinical outcome in Portuguese patients and showa high prevalence of infections with multiple strains. Scand. J. Gastroenterol. 36:128–35
86 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
100. Ito Y, Azuma T, Ito S, Suto H, Miyaji H, et al. 1998. Full-length sequence analysis of thevacA gene from cytotoxic and noncytotoxic Helicobacter pylori. J. Infect. Dis. 178:1391–98
101. Telford JL, Ghiara P, Dellorco M, Comanducci M, Burroni D, et al. 1994. Gene structureof the Helicobacter pylori cytotoxin and evidence of its key role in gastric disease. J. Exp.Med. 179:1653–58
102. Ilver D, Barone S, Mercati D, Lupetti P, Telford JL. 2004. Helicobacter pylori toxinVacA is transferred to host cells via a novel contact-dependent mechanism. Cell. Microbiol.6:167–74
103. Yahiro K, Wada A, Nakayama M, Kimura T, Ogushi K, et al. 2003. Protein-tyrosinephosphatase alpha, RPTP alpha, is a Helicobacter pylori VacA receptor. J. Biol. Chem.278:19183–89
104. Yahiro K, Niidome T, Kimura M, Hatakeyama T, Aoyagi H, et al. 1999. Activation ofHelicobacter pylori VacA toxin by alkaline or acid conditions increases its binding to a250-kDa receptor protein-tyrosine phosphatase beta. J. Biol. Chem. 274:36693–99
105. Padilla PI, Wada A, Yahiro K, Kimura M, Niidome T, et al. 2000. Morphologic differ-entiation of HL-60 cells is associated with appearance of RPTPbeta and induction ofHelicobacter pylori VacA sensitivity. J. Biol. Chem. 275:15200–6
106. Seto K, Hayashi-Kuwabara Y, Yoneta T, Suda H, Tamaki H. 1998. Vacuolation inducedby cytotoxin from Helicobacter pylori is mediated by the EGF receptor in HeLa cells.FEBS Lett. 431:347–50
107. Schraw W, Li Y, McClain MS, van der Goot FG, Cover TL. 2002. Association of Heli-cobacter pylori vacuolating toxin (VacA) with lipid rafts. J. Biol. Chem. 277:34642–50
108. Kuo CH, Wang WC. 2003. Binding and internalization of Helicobacter pylori VacA viacellular lipid rafts in epithelial cells. Biochem. Biophys. Res. Commun. 303:640–44
109. McClain MS, Schraw W, Ricci V, Boquet P, Cover TL. 2000. Acid activation of Heli-cobacter pylori vacuolating cytotoxin (VacA) results in toxin internalization by eukaryoticcells. Mol. Microbiol. 37:433–42
110. Ricci V, Sommi P, Fiocca R, Necchi V, Romano M, et al. 2002. Extracellular pH modulatesHelicobacter pylori-induced vacuolation and VacA toxin internalization in human gastricepithelial cells. Biochem. Biophys. Res. Commun. 292:167–74
111. Czajkowsky DM, Iwamoto H, Cover TL, Shao ZF. 1999. The vacuolating toxin fromHelicobacter pylori forms hexameric pores in lipid bilayers at low pH. Proc. Natl. Acad.Sci. USA 96:2001–6
112. Szabo I, Brutsche S, Tombola F, Moschioni M, Satin B, et al. 1999. Formation of anion-selective channels in the cell plasma membrane by the toxin VacA of Helicobacter pyloriis required for its biological activity. EMBO J. 18:5517–27
113. Tombola F, Morbiato L, Del Giudice G, Rappuoli R, Zoratti M, et al. 2001. The Heli-cobacter pylori VacA toxin is a urea permease that promotes urea diffusion across epithelia.J. Clin. Invest. 108:929–37
114. Papini E, Debernard M, Milia E, Bugnoli M, Zerial M, et al. 1994. Cellular vacuolesinduced by Helicobacter pylori originate from late endosomal compartments. Proc. Natl.Acad. Sci. USA 91:9720–24
115. Papini E, Satin B, Norais N, de Bernard M, Telford JL, et al. 1998. Selective increase ofthe permeability of polarized epithelial cell monolayers by Helicobacter pylori vacuolatingtoxin. J. Clin. Invest. 102:813–20
116. Merrell DS, Thompson LJ, Kim CC, Mitchell H, Tompkins LS, et al. 2003. Growth phase-dependent response of Helicobacter pylori to iron starvation. Infect. Immun. 71:6510–25
www.annualreviews.org • Helicobacter pylori–Induced Diseases 87
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
117. Kuck D, Kolmerer B, Iking-Konert C, Krammer PH, Stremmel W, et al. 2001. Vacuo-lating cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelialcell line AGS. Infect. Immun. 69:5080–87
118. Cover TL, Krishna US, Israel DA, Peek RM Jr. 2003. Induction of gastric epithelial cellapoptosis by Helicobacter pylori vacuolating cytotoxin. Cancer Res. 63:951–57
119. Galmiche A, Rassow J, Doye A, Cagnol S, Chambard JC, et al. 2000. The N-terminal34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria andinduces cytochrome c release. EMBO J. 19:6361–70
120. Willhite DC, Cover TL, Blanke SR. 2003. Cellular vacuolation and mitochondrial cy-tochrome c release are independent outcomes of Helicobacter pylori vacuolating cyto-toxin activity that are each dependent on membrane channel formation. J. Biol. Chem.278:48204–9
121. Nakayama M, Kimura M, Wada A, Yahiro K, Ogushi K, et al. 2004. Helicobacter pyloriVacA activates the p38/activating transcription factor 2-mediated signal pathway in AZ-521 cells. J. Biol. Chem. 279:7024–28
122. Marchetti M, Arico B, Burroni D, Figura N, Rappuoli R, et al. 1995. Developmentof a mouse model of Helicobacter pylori infection that mimics human disease. Science267:1655–58
123. Fujikawa A, Shirasaka D, Yamamoto S, Ota H, Yahiro K, et al. 2003. Mice deficient inprotein tyrosine phosphatase receptor type Z are resistant to gastric ulcer induction byVacA of Helicobacter pylori. Nat. Genet. 33:375–81
124. de Bernard M, Cappon A, Pancotto L, Ruggiero P, Rivera J, et al. 2005. The Helicobacterpylori VacA cytotoxin activates RBL-2H3 cells by inducing cytosolic calcium oscillations.Cell. Microbiol. 7:191–98
125. Molinari M, Salio M, Galli C, Norais N, Rappuoli R, et al. 1998. Selective inhibitionof Ii-dependent antigen presentation by Helicobacter pylori toxin VacA. J. Exp. Med.187:135–40
126. Gebert B, Fischer W, Weiss E, Hoffmann R, Haas R. 2003. Helicobacter pylori vacuo-lating cytotoxin inhibits T lymphocyte activation. Science 301:1099–102
127. Boncristiano M, Paccani SR, Barone S, Ulivieri C, Patrussi L, et al. 2003. The Helicobacterpylori vacuolating toxin inhibits T cell activation by two independent mechanisms. J. Exp.Med. 198:1887–97
128. Sundrud MS, Torres VJ, Unutmaz D, Cover TL. 2004. Inhibition of primary human Tcell proliferation by Helicobacter pylori vacuolating toxin (VacA) is independent of VacAeffects on IL-2 secretion. Proc. Natl. Acad. Sci. USA 101:7727–32
129. Letley DP, Atherton JC. 2000. Natural diversity in the N terminus of the mature vacuolat-ing cytotoxin of Helicobacter pylori determines cytotoxin activity. J. Bacteriol. 182:3278–80
130. McClain MS, Cao P, Iwamoto H, Vinion-Dubiel AD, Szabo G, et al. 2001. A 12-amino-acid segment, present in type s2 but not type s1 Helicobacter pylori VacA proteins, abol-ishes cytotoxin activity and alters membrane channel formation. J. Bacteriol. 183:6499–508
131. Aviles-Jimenez F, Letley DP, Gonzalez-Valencia G, Salama N, Torres J, et al. 2004. Evo-lution of the Helicobacter pylori vacuolating cytotoxin in a human stomach. J. Bacteriol.186:5182–85
132. Boren T, Falk P, Roth KA, Larson G, Normark S. 1993. Attachment of Helicobacterpylori to human gastric epithelium mediated by blood-group antigens. Science 262:1892–95
88 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
133. Sheu BS, Sheu SM, Yang HB, Huang AH, Wu JJ. 2005. Host gastric Lewis expressiondetermines the bacterial density of Helicobacter pylori in babA2 genopositive infection.Gut 52:927–32
134. Guruge JL, Falk PG, Lorenz RG, Dans M, Wirth HP, et al. 1998. Epithelial attachmentalters the outcome of Helicobacter pylori infection. Proc. Natl. Acad. Sci. USA 95:3925–30
135. Mahdavi J, Sonden B, Hurtig M, Olfat FO, Forsberg L, et al. 2002. Helicobacter py-lori SabA adhesin in persistent infection and chronic inflammation. Science 297:573–78
136. Ilver D, Arnqvist A, Ogren J, Frick IM, Kersulyte D, et al. 1998. Helicobacter pyloriadhesin binding fucosylated histo-blood group antigens revealed by retagging. Science279:373–77
137. Gerhard M, Lehn N, Neumayer N, Boren T, Rad R, et al. 1999. Clinical relevance of theHelicobacter pylori gene for blood-group antigen-binding adhesin. Proc. Natl. Acad. Sci.USA 96:12778–83
138. Prinz C, Schoniger M, Rad R, Becker I, Keiditsch E, et al. 2001. Key importance of theHelicobacter pylori adherence factor blood group antigen binding adhesin during chronicgastric inflammation. Cancer Res. 61:1903–9
139. Yu J, Leung WK, Go MYY, Chan MCW, To KF, et al. 2002. Relationship betweenHelicobacter pylori babA2 status with gastric epithelial cell turnover and premalignantgastric lesions. Gut 51:480–84
140. Solnick JV, Hansen LM, Salama NR, Boonjakuakul JK, Syvanen M. 2004. Modificationof Helicobacter pylori outer membrane protein expression during experimental infectionof rhesus macaques. Proc. Natl. Acad. Sci. USA 101:2106–11
141. Backstrom A, Lundberg C, Kersulyte D, Berg DE, Boren T, et al. 2004. Metastabilityof Helicobacter pylori bab adhesin genes and dynamics in Lewis b antigen binding. Proc.Natl. Acad. Sci. USA 101:16923–28
142. Hennig EE, Mernaugh R, Edl J, Cao P, Cover TL. 2004. Heterogeneity among Heli-cobacter pylori strains in expression of the outer membrane protein BabA. Infect. Immun.72:3429–35
143. Yamaoka Y, Kwon DH, Graham DY. 2000. A M-r 34,000 proinflammatory outer mem-brane protein (oipA) of Helicobacter pylori. Proc. Natl. Acad. Sci. USA 97:7533–38
144. Peek RM, Thompson SA, Donahue JP, Tham KT, Atherton JC, et al. 1998. Adherenceto gastric epithelial cells induces expression of a Helicobacter pylori gene, iceA, that isassociated with clinical outcome. Proc. Assoc. Am. Physicians 110:531–44
145. Kidd M, Peek RM, Lastovica AJ, Israel DA, Kummer AF, et al. 2001. Analysis of iceAgenotypes in South African Helicobacter pylori strains and relationship to clinically sig-nificant disease. Gut 49:629–35
146. Heithoff DM, Sinsheimer RL, Low DA, Mahan MJ. 1999. An essential role for DNAadenine methylation in bacterial virulence. Science 284:967–70
147. Xu Q, Blaser MJ. 2001. Promoters of the CATG-specific methyltransferase gene hpyIMdiffer between iceA1 and iceA2 Helicobacter pylori strains. J. Bacteriol. 183:3875–84
148. Ando T, Aras RA, Kusugami K, Blaser MJ, Wassenaar TM. 2003. Evolutionary historyof hrgA, which replaces the restriction gene hpyIIIR in the hpyIII locus of Helicobacterpylori. J. Bacteriol. 185:295–301
149. Ando T, Wassenaar TM, Peek RM, Aras RA, Tschumi AI, et al. 2002. A Helicobacterpylori restriction endonuclease-replacing gene, hrgA, is associated with gastric cancer inAsian strains. Cancer Res. 62:2385–89
www.annualreviews.org • Helicobacter pylori–Induced Diseases 89
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
150. Hwang IR, Kodama T, Kikuchi S, Sakai K, Peterson LE, et al. 2002. Effect of interleukin1 polymorphisms on gastric mucosal interleukin 1 beta production in Helicobacter pyloriinfection. Gastroenterology 123:1793–803
151. Rad R, Dossumbekova A, Neu B, Lang R, Bauer S, et al. 2004. Cytokine gene poly-morphisms influence mucosal cytokine expression, gastric inflammation, and host specificcolonisation during Helicobacter pylori infection. Gut 53:1082–89
152. Furuta T, El-Omar EM, Xiao F, Shirai N, Takashima M, et al. 2002. Interleukin 1 betapolymorphisms increase risk of hypochlorhydria and atrophic gastritis and reduce risk ofduodenal ulcer recurrence in Japan. Gastroenterology 123:95–105
153. Machado JC, Figueiredo C, Canedo P, Nabais S, Alves CC, et al. 2003. A pro-inflammatory genetic profile increases the risk of chronic atrophic gastritis and gastriccarcinoma. Gastroenterology 125:364–71
154. Rad R, Prinz C, Neu B, Neuhofer M, Zeitner M, et al. 2003. Synergistic effect of Heli-cobacter pylori virulence factors and interleukin-1 polymorphisms for the developmentof severe histological changes in the gastric mucosa. J. Infect. Dis. 188:272–81
The firstdescription ofhuman cytokinepolymorphismsaffecting gastriccancer risk inH. pylori-infectedpeople.
155.El-Omar EM, Carrington M, Chow WH, McColl KEL, Bream JH, et al. 2000.Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Na-
ture 404:398–402156. Machado JC, Pharoah P, Sousa S, Carvalho R, Oliveira C, et al. 2001. Interleukin 1B and
interleukin 1RN polymorphisms are associated with increased risk of gastric carcinoma.Gastroenterology 121:823–29
157. El-Omar EM, Rabkin CS, Gammon MD, Vaughan TL, Risch HA, et al. 2003. Increasedrisk of noncardia gastric cancer associated with proinflammatory cytokine gene polymor-phisms. Gastroenterology 124:1193–201
158. Garza-Gonzalez E, Bosques-Padilla FJ, El-Omar E, Hold G, Tijerina-Menchaca R, et al.2005. Role of the polymorphic IL-1B, IL-1RN and TNF-A genes in distal gastric cancerin Mexico. Int. J. Cancer 114:237–41
159. Hsu PI, Li CN, Tseng HH, Lai KH, Hsu PN, et al. 2004. The Interleukin-1 RN polymor-phism and Helicobacter pylori infection in the development of duodenal ulcer. Helicobacter9:605–13
160. Ohyauchi M, Imatani A, Yonechi M, Asano N, Miura A, et al. 2005. The polymorphisminterleukin 8–2251 A/T influences the susceptibility of Helicobacter pylori related gastricdiseases in the Japanese population. Gut 54:330–35
161. Bamford KB, Fan X, Crowe SE, Leary JF, Gourley WK, et al. 1998. Lymphocytes inthe human gastric mucosa during Helicobacter pylori have a T helper cell 1 phenotype.Gastroenterology 114:482–92
162. D’Elios MM, Manghetti M, De Carli M, Costa F, Baldari CT, et al. 1997. T helper 1effector cells specific for Helicobacter pylori in the gastric antrum of patients with pepticulcer disease. J. Immunol. 158:962–67
163. Mohammadi M, Nedrud J, Redline R, Lycke N, Czinn SJ. 1997. Murine CD4 T-cellresponse to Helicobacter infection: TH1 cells enhance gastritis and TH2 cells reducebacterial load. Gastroenterology 113:1848–57
164. Smythies LE, Waites KB, Lindsey JR, Harris PR, Ghiara P, et al. 2000. Helicobacterpylori-induced mucosal inflammation is Th1 mediated and exacerbated in IL-4, but notIFN-gamma, gene-deficient mice. J. Immunol. 165:1022–29
165. Yamori M, Yoshida M, Watanabe T, Shirai Y, Iizuka T, et al. 2004. Antigenic activationof Th1 cells in the gastric mucosa enhances dysregulated apoptosis and turnover of theepithelial cells. Biochem. Biophys. Res. Commun. 316:1015–21
90 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
166. Cui G, Houghton J, Finkel N, Carlson J, Wang TC. 2003. IFN-gamma infusion inducesgastric atrophy, metaplasia and dysplasia in the absence of Helicobacter pylori infection - arole for the immune response in Helicobacter disease. Gastroenterology 2003:A19
167. Mohammadi M, Czinn S, Redline R, Nedrud J. 1996. Helicobacter-specific cell-mediatedimmune responses display a predominant Th1 phenotype and promote a delayed-typehypersensitivity response in the stomachs of mice. J. Immunol. 156:4729–38
168. Obonyo M, Guiney DG, Harwood J, Fierer J, Cole SP. 2002. Role of gamma interferon inHelicobacter pylori induction of inflammatory mediators during murine infection. Infect.Immun. 70:3295–99
169. Guiney DG, Hasegawa P, Cole SP. 2003. Helicobacter pylori preferentially induces in-terleukin 12 (IL-12) rather than IL-6 or IL-10 in human dendritic cells. Infect. Immun.71:4163–66
170. Kranzer K, Eckhardt A, Aigner M, Knoll G, Deml L, et al. 2004. Induction of matura-tion and cytokine release of human dendritic cells by Helicobacter pylori. Infect. Immun.72:4416–23
171. Galgani M, Busiello I, Censini S, Zappacosta S, Racioppi L, et al. 2004. Helicobacterpylori induces aptosis of human monocytes but not monocyte-derived dendritic cells: roleof the cag pathogenicity island. Infect. Immun. 72:4480–85
This animal studyraises theinterestinghypothesis thatparasitemodulation of theimmune responsemay affect diseaserisk.
172.Fox JG, Beck P, Dangler CA, Whary MT, Wang TC, et al. 2000. Concurrent en-teric helminth infection modulates inflammation and gastric immune responsesand reduces helicobacter-induced gastric atrophy. Nat. Med. 6:536–42
173. Foryst-Ludwig A, Naumann M. 2000. p21-activated kinase 1 activates the nuclear fac-tor kappa B (NF-kappa B)-inducing kinase-Ikappa B kinases NF-kappa B pathway andproinflammatory cytokines in Helicobacter pylori infection. J. Biol. Chem. 275:39779–85
174. Girardin SE, Boneca IG, Carneiro LAM, Antignac A, Jehanno M, et al. 2003. Nod1 detectsa unique muropeptide from Gram-negative bacterial peptidoglycan. Science 300:1584–87
175. Bergman MP, Engering A, Smits HH, van Vliet SJ, van Bodegraven AA, et al. 2004.Helicobacter pylori modulates the T helper cell 1/T helper cell 2 balance through phase-variable interaction between lipopolysaccharide and DC-SIGN. J. Exp. Med. 200:979–90
176. Meyer F, Ramanujam KS, Gobert AP, James SP, Wilson KT. 2003. Cutting edge:cyclooxygenase-2 activation suppresses Th1 polarization in response to Helicobacter py-lori. J. Immunol. 171:3913–17
177. Lundgren A, Suri-Payer E, Enarsson K, Svennerholm AM, Lundin BS. 2003. Helicobacterpylon-specific CD4(+) CD25(high) regulatory T cells suppress memory T-cell responsesto H. pylori in infected individuals. Infect. Immun. 71:1755–62
178. Moss SF, Legon S, Bishop AE, Polak JM, Calam J. 1992. Effect of Helicobacter pylori ongastric somatostatin in duodenal-ulcer disease. Lancet 340:930–2
179. Beales I, Calam J, Post L, Srinivasan S, Yamada T, et al. 1997. Effect of transforminggrowth factor alpha and interleukin 8 on somatostatin release from canine fundic D cells.Gastroenterology 112:136–43
180. Odum L, Petersen HD, Andersen IB, Hansen BF, Rehfeld JF, et al. 1994. Gastrin andsomatostatin in Helicobacter pylori infected antral mucosa. Gut 35:615–18
181. Arebi N, Healey ZV, Bliss PW, Ghatei M, Van Noorden S, et al. 2002. Nitric oxideregulates the release of somatostatin from cultured gastric rabbit primary D-cells. Gas-troenterology 123:566–76
182. Zavros Y, Rathinavelu S, Kao JY, Todisco A, Del Valle J, et al. 2003. Treatment of Heli-cobacter gastritis with IL-4 requires somatostatin. Proc. Natl. Acad. Sci. USA 100:12944–49
www.annualreviews.org • Helicobacter pylori–Induced Diseases 91
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
Thehypergastrinemicmouse describedhere is animportant model ofgastriccarcinogenesis andunderlines the keyrole of gastrin.
183.Wang TC, Dangler CA, Chen D, Goldenring JR, Koh T, et al. 2000. Synergisticinteraction between hypergastrinemia and Helicobacter infection in a mouse modelof gastric cancer. Gastroenterology 118:36–47
184. Peek RM, Wirth HP, Moss SF, Yang MQ, Abdalla AM, et al. 2000. Helicobacter py-lori alters gastric epithelial cell cycle events and gastrin secretion in Mongolian gerbils.Gastroenterology 118:48–59
185. Guo YS, Cheng JZ, Jin GF, Gutkind JS, Hellmich MR, et al. 2002. Gastrin stimulatescyclooxygenase-2 expression in intestinal epithelial cells through multiple signaling path-ways - evidence for involvement of Erk5 kinase and transactivation of the epidermal growthfactor receptor. J. Biol. Chem. 277:48755–63
186. Fukui H, Franceschi F, Penland RL, Sakai T, Sepulveda AR, et al. 2003. Effects of He-licobacter pylori infection on the link between regenerating gene expression and serumgastrin levels in Mongolian gerbils. Lab. Invest. 83:1777–86
187. Azuma T, Suto H, Ito Y, Ohtani M, Dojo M, et al. 2001. Gastric leptin and Helicobacterpylori infection. Gut 49:324–29
188. Breidert M, Miehlke S, Glasow A, Orban Z, Stolte M, et al. 1999. Leptin and its receptorin normal human gastric mucosa and in Helicobacter pylori-associated gastritis. Scand. J.Gastroenterol. 34:954–61
189. Nwokolo CU, Freshwater DA, O’Hare P, Randeva HS. 2003. Plasma ghrelin followingcure of Helicobacter pylori. Gut 52:637–40
190. Atherton JC, Tham KT, Peek RM, Cover TL, Blaser MJ. 1996. Density of Helicobacterpylori infection in vivo as assessed by quantitative culture and histology. J. Infect. Dis.174:552–56
191. Gillen D, El-Omar EM, Wirz AA, Ardill JES, McColl KEL. 1998. The acid response togastrin distinguishes duodenal ulcer patients from Helicobacter pylori–infected healthysubjects. Gastroenterology 114:50–57
192. Hamlet A, Olbe L. 1996. The influence of Helicobacter pylori infection on postpran-dial duodenal acid load and duodenal bulb pH in humans. Gastroenterology 111:391–400
193. Khulusi S, Badve S, Patel P, Lloyd R, Marrero JM, et al. 1996. Pathogenesis of gas-tric metaplasia of the human duodenum: role of Helicobacter pylori, gastric acid, andulceration. Gastroenterology 110:452–58
194. Van Zanten S, Dixon MF, Lee A. 1999. The gastric transitional zones: neglected linksbetween gastroduodenal pathology and Helicobacter ecology. Gastroenterology 116:1217–29
195. Amedei A, Bergman MP, Appelmelk BJ, Azzurri A, Benagiano M, et al. 2003. Molecularmimicry between Helicobacter pylori antigens and H+, K+-adenosine triphosphatase inhuman gastric autoimmunity. J. Exp. Med. 198:1147–56
196. ElOmar EM, Oien K, ElNujumi A, Gillen D, Wirz A, et al. 1997. Helicobacter pyloriinfection and chronic gastric acid hyposecretion. Gastroenterology 113:15–24
197. Beales ILP, Calam J. 1998. Interleukin 1 beta and tumour necrosis factor alpha in-hibit acid secretion in cultured rabbit parietal cells by multiple pathways. Gut 42:227–34
198. Padol IT, Hunt RH. 2004. Effect of Th1 cytokines on acid secretion in pharmacologicallycharacterised mouse gastric glands. Gut 53:1075–81
199. Kuipers EJ, Uyterlinde AM, Pena AS, Hazenberg HJA, Bloemena E, et al. 1995. In-crease of Helicobacter pylori-associated corpus gastritis during acid suppressive therapy-implications for long-term safety. Am. J. Gastroenterol. 90:1401–6
92 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
200. Zavros Y, Rieder G, Ferguson A, Samuelson LC, Merchant JL. 2002. Hypergastrinemiain response to gastric inflammation suppresses somatostatin. Am. J. Physiol. Gastrointest.Liver Physiol. 282:G175–G83
201. Wong BCY, Lam SK, Wong WM, Chen JS, Zheng TT, et al. 2004. Helicobacter py-lori eradication to prevent gastric cancer in a high-risk region of China - a randomizedcontrolled trial. JAMA 291:187–94
202. Touati E, Michel V, Thiberge JM, Wuscher N, Huerre M, et al. 2003. Chronic Helicobac-ter pylori infections induce gastric mutations in mice. Gastroenterology 124:1408–19
203. Nam KT, Oh SY, Ahn B, Kim YB, Jang DD, et al. 2004. Decreased Helicobacter py-lori associated gastric carcinogenesis in mice lacking inducible nitric oxide synthase. Gut53:1250–55
204. Correa P, Fontham ETH, Bravo JC, Bravo LE, Ruiz B, et al. 2000. Chemopreventionof gastric dysplasia: randomized trial of antioxidant supplements and anti-Helicobacterpylori therapy. J. Natl. Cancer Inst. 92:1881–88
205. You WC, Zhang L, Gail MH, Chang YS, Liu WD, et al. 2000. Gastric cancer: Helicobacterpylori, serum vitamin C, and other risk factors. J. Natl. Cancer Inst. 92:1607–12
206. Dimmler A, Brabletz T, Hlubek F, Hafner M, Rau T, et al. 2003. Transcription of sonichedgehog, a potential factor for gastric morphogenesis and gastric mucosa maintenance,is up-regulated in acidic conditions. Lab. Invest. 83:1829–37
207. Syder AJ, Oh JD, Guruge JL, O’Donnell D, Karlsson M, et al. 2003. The impact of parietalcells on Helicobacter pylori tropism and host pathology: an analysis using gnotobioticnormal and transgenic mice. Proc. Natl. Acad. Sci. USA 100:3467–72
208. Nomura S, Baxter T, Yamaguchi H, Leys C, Vartapetian AB, et al. 2004. Spasmolyticpolypeptide expressing metaplasia to preneoplasia in H-felis-infected mice. Gastroenterol-ogy 127:582–94
209. Mutoh H, Sakurai S, Satoh K, Tamada K, Kita H, et al. 2004. Development of gastriccarcinoma from intestinal metaplasia in CDx2-transgenic mice. Cancer Res. 64:7740–47
That gastric cancermay develop frombonemarrow-derivedgastric stem cells ischanging our viewson carcinogenesis.
210.Houghton J, Stoicov C, Nomura S, Rogers AB, Carlson J, et al. 2004. Gastric canceroriginating from bone marrow-derived cells. Science 306:1568–71
211. Lauren P. 1965. 2 Histological main types of gastric carcinoma-diffuse and so-calledintestinal-type carcinoma—an attempt at a histo-clinical classification. Acta Pathol. Micro-biol. Scand. 64:31–49
212. Leung WK, Lin SR, Ching JYL, To KF, Ng EKW, et al. 2004. Factors predicting progres-sion of gastric intestinal metaplasia: results of a randomised trial on Helicobacter pylorieradication. Gut 53:1244–49
213. Semino-Mora C, Doi SQ, Marty A, Simko V, Carlstedt I, et al. 2003. Intracellular andinterstitial expression of Helicobacter pylori virulence genes in gastric precancerous in-testinal metaplasia and adenocarcinoma. J. Infect. Dis. 187:1165–77
214. Wagner S, Beil W, Westermann J, Logan RPH, Bock CT, et al. 1997. Regulation ofgastric epithelial cell growth by Helicobacter pylori: evidence for a major role of apoptosis.Gastroenterology 113:1836–47
215. Ashktorab H, Frank S, Khaled AR, Durum SK, Kifle B, et al. 2004. Bax translocation andmitochondrial fragmentation induced by Helicobacter pylori. Gut 53:805–13
216. Eguchi H, Carpentier S, Kim SS, Moss SF. 2004. p27(kip1) regulates the apoptotic re-sponse of gastric epithelial cells to Helicobacter pylori. Gut 53:797–804
217. Jones NL, Day AS, Jennings H, Shannon PT, Galindo-Mata E, et al. 2002. Enhanceddisease severity in Helicobacter pylori-infected mice deficient in Fas signaling. Infect.Immun. 70:2591–97
www.annualreviews.org • Helicobacter pylori–Induced Diseases 93
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
218. Obst B, Wagner S, Sewing KF, Beil W. 2000. Helicobacter pylori causes DNA damagein gastric epithelial cells. Carcinogenesis 21:1111–15
219. Kim JJ, Tao H, Carloni E, Leung WK, Graham DY, et al. 2002. Helicobacter pyloriimpairs DNA mismatch repair in gastric epithelial cells. Gastroenterology 123:542–53
220. Smoot DT, Elliott TB, Verspaget HW, Jones D, Allen CR, et al. 2000. Influence ofHelicobacter pylori on reactive oxygen-induced gastric epithelial cell injury. Carcinogenesis21:2091–95
221. Ding SZ, O’Hara AM, Denning TL, Dirden-Kramer B, Mifflin RC, et al. 2004. He-licobacter pylori and H2O2 increase AP endonuclease-1/redox factor-1 expression inhuman gastric epithelial cells. Gastroenterology 127:845–58
222. Nagata K, Yu H, Nishikawa M, Kashiba M, Nakamura A, et al. 1998. Helicobacter py-lori generates superoxide radicals and modulates nitric oxide metabolism. J. Biol. Chem.273:14071–73
223. O’Rourke EJ, Chevalier C, Pinto AV, Thiberge JM, Ielpi L, et al. 2003. Pathogen DNAas target for host-generated oxidative stress: role for repair of bacterial DNA damage inHelicobacter pylori colonization. Proc. Natl. Acad. Sci. USA 100:2789–94
224. el-Serag HB, Sonnenberg A. 1998. Opposing time trends of peptic ulcer and reflux disease.Gut 43:327–33
225. Yamaji Y, Mitsushima T, Ikuma H, Okamoto M, Yoshida H, et al. 2001. Inverse back-ground of Helicobacter pylori antibody and pepsinogen in reflux oesophagitis comparedwith gastric cancer: analysis of 5732 Japanese subjects. Gut 49:335–40
226. Ye WM, Held M, Lagergren J, Engstrand L, Blot WJ, et al. 2004. Helicobacter pyloriinfection and gastric atrophy: risk of adenocarcinoma and squamous-cell carcinoma of theesophagus and adenocarcinoma of the gastric cardia. J. Natl. Cancer Inst. 96:388–96
227. de Martel C, Llosa AE, Farr SM, Friedman GD, Vogelman JH, et al. 2005. Helicobacterpylori infection and the risk of development of esophageal adenocarcinoma. J. Infect. Dis.191:761–67
228. Warburton-Timms VJ, Charlett A, Valori RM, Uff JS, Shepherd NA, et al. 2001. Thesignificance of cagA(+) Helicobacter pylori in reflux oesophagitis. Gut 49:341–46
229. Chow WH, Blaser MJ, Blot WJ, Gammon MD, Vaughan TL, et al. 1998. An inverserelation between cagA+ strains of Helicobacter pylori infection and risk of esophagealand gastric cardia adenocarcinoma. Cancer Res. 58:588–90
230. Vaezi MF, Falk GW, Peek RM, Vicari JJ, Goldblum JR, et al. 2000. CagA-positivestrains of Helicobacter pylori may protect against Barrett’s esophagus. Am. J. Gastroenterol.95:2206–11
231. El-Serag HB, Sonnenberg A, Jamal MM, Inadomi JM, Crooks LA, et al. 1999. Corpusgastritis is protective against reflux oesophagitis. Gut 45:181–85
232. Koike T, Ohara S, Sekine H, Iijima K, Abe Y, et al. 2001. Helicobacter pylori infectionprevents erosive reflux oesophagitis by decreasing gastric acid secretion. Gut 49:330–34
233. Rothenbacher D, Blaser MJ, Bode G, Brenner H. 2000. Inverse relationship betweengastric colonization of Helicobacter pylori and diarrheal illnesses in children: results of apopulation-based cross-sectional study. J. Infect. Dis. 182:1446–49
234. Wang G, Ge ZM, Rasko A, Taylor DE. 2000. Lewis antigens in Helicobacter pylori:biosynthesis and phase variation. Mol. Microbiol. 36:1187–96
235. Baron JH, Sonnenberg A. 2001. Period- and cohort-age contours of deaths from gastricand duodenal ulcer in New York 1804–1998. Am. J. Gastroenterol. 96:2887–91
236. Susser S. 1962. Civilisation and peptic ulcer. Lancet 1:115–18
94 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
237. deJong D, vanderHulst RWM, Pals G, vanDijk WC, vanderEnde A, et al. 1996. Gastricnon-Hodgkin lymphomas of mucosa-associated lymphoid tissue are not associated withmore aggressive Helicobacter pylori strains as identified by CagA. Am. J. Clin. Pathol.106:670–75
238. Peng HZ, Ranaldi R, Diss TC, Isaacson PG, Bearzi I, et al. 1998. High frequency ofCagA+ Helicobacter pylori infection in high-grade gastric MALT B-cell lymphomas. J.Pathol. 185:409–12
239. Delchier JC, Lamarque D, Levy M, Tkoub EM, Copie-Bergman C, et al. 2001. Heli-cobacter pylori and gastric lymphoma: high seroprevalence of CagA in diffuse large B-celllymphoma but not in low-grade lymphoma of mucosa-associated lymphoid tissue type.Am. J. Gastroenterol. 96:2324–28
240. Lehours P, Menard A, Dupouy S, Bergey B, Richy F, et al. 2004. Evaluation of theassociation of nine Helicobacter pylori virulence factors with strains involved in low-gradegastric mucosa-associated lymphoid tissue lymphoma. Infect. Immun. 72:880–88
241. Miehlke S, Yu J, Schuppler M, Frings C, Kirsch C, et al. 2001. Helicobacter pylori vacA,iceA, and cagA status and pattern of gastritis in patients with malignant and benign gas-troduodenal disease. Am. J. Gastroenterol. 96:1008–13
242. Hussell T, Isaacson PG, Crabtree JE, Spencer J. 1993. The response of cells from low-grade B-cell gastric lymphomas of mucosa-associated lymphoid-tissue to Helicobacterpylori. Lancet 342:571–74
243. Du M, Diss TC, Xu C, Peng H, Isaacson PG, et al. 1996. Ongoing mutation in MALTlymphoma immunoglobulin gene suggests that antigen stimulation plays a role in theclonal expansion. Leukemia 10:1190–97
Proved that theselymphomas areH. pylori driven,and led to effectivetreatment byH. pylori
eradication.
244.Wotherspoon AC, Doglioni C, Diss TC, Pan LX, Moschini A, et al. 1993. Re-gression of primary low-grade B-cell gastric lymphoma of mucosa-associatedlymphoid-tissue type after eradication of Helicobacter pylori. Lancet 342:575–77
245. Fischbach W, Goebeler-Kolve ME, Dragosics B, Greiner A, Stolte M. 2004. Long termoutcome of patients with gastric marginal zone B cell lymphoma of mucosa associatedlymphoid tissue (MALT) following exclusive Helicobacter pylori eradication therapy: ex-perience from a large prospective series. Gut 53:34–37
246. Ott G, Katzenberger T, Greiner A, Kalla J, Rosenwald A, et al. 1997. Thet(11;18)(q21;q21) chromosome translocation is a frequent and specific aberration in low-grade but not high-grade malignant non-Hodgkin’s lymphomas of the mucosa-associatedlymphoid tissue (MALT-) type. Cancer Res. 57:3944–48
247. Auer IA, Gascoyne RD, Connors JM, Cotter FE, Greiner TC, et al. 1997. t(11;18)(q21;q21) is the most common translocation in MALT lymphomas. Ann. Oncol. 8:979–85
248. Dierlamm J, Baens M, Wlodarska I, Stefanova-Ouzounova M, Hernandez JM, et al. 1999.The apoptosis inhibitor gene API2 and a novel 18q gene, MLT, are recurrently rearrangedin the t(11;18)(q21;q21) associated with mucosa-associated lymphoid tissue lymphomas.Blood 93:3601–9
249. Lucas PC, Yonezumi M, Inohara N, McAllister-Lucas LM, Abazeed ME, et al. 2001.Bcl10 and MALT1, independent targets of chromosomal translocation in MALT lym-phoma, cooperate in a novel NF-kappa B signaling pathway. J. Biol. Chem. 276:19012–19
250. Liu HX, Ruskon-Fourmestraux A, Lavergne-Slove A, Ye HT, Molina T, et al. 2001.Resistance of t(11;18) positive gastric mucosa-associated lymphoid tissue lymphoma toHelicobacter pylori eradication therapy. Lancet 357:39–40
www.annualreviews.org • Helicobacter pylori–Induced Diseases 95
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

ANRV268-PM01-03 ARI 16 December 2005 15:25
251. Liu HX, Ye HT, Ruskone-Fourmestraux A, de Jong D, Pileri S, et al. 2001. t(11;18)is a marker for all stage gastric MALT lymphomas that will not respond to H. pylorieradication. Blood 98:767A
252. Ye HT, Liu HX, Attygalle A, Wotherspoon AC, Nicholson AG, et al. 2003. Variablefrequencies of t(11;18)(q21;q21) in MALT lymphomas of different sites: significant asso-ciation with CagA strains of H. pylori in gastric MALT lymphoma. Blood 102:1012–18
96 Atherton
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

Contents ARI 7 December 2005 18:14
Annual Reviewof Pathology:Mechanisms ofDisease
Volume 1, 2006Contents
FrontispieceMorris J. Karnovsky � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xii
A Pathologist’s OdysseyMorris J. Karnovsky � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Immunobiology and Pathogenesis of Viral HepatitisLuca G. Guidotti and Francis V. Chisari � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �23
The Pathogenesis of Helicobacter pylori–InducedGastro-Duodenal DiseasesJohn C. Atherton � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �63
Molecular Pathology of Malignant GliomasDavid N. Louis � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �97
Tumor Stroma and Regulation of Cancer DevelopmentThea D. Tlsty and Lisa M. Coussens � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 119
Neurodegenerative Diseases: New Concepts of Pathogenesis andTheir Therapeutic ImplicationsDaniel M. Skovronsky, Virginia M.-Y. Lee, and John Q. Trojanowski � � � � � � � � � � � � � � � � � � 151
The Endothelium as a Target for InfectionsGustavo Valbuena and David H. Walker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 171
Genetic Regulation of Cardiogenesis and Congenital Heart DiseaseDeepak Srivastava � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 199
Regulation of Lung Inflammation in the Model of IgGImmune-Complex InjuryHongwei Gao, Thomas Neff, and Peter A. Ward � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 215
Integrative Biology of Prostate Cancer ProgressionScott A. Tomlins, Mark A. Rubin, and Arul M. Chinnaiyan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 243
KSHV Infection and the Pathogenesis of Kaposi’s SarcomaDon Ganem � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 273
vi
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.

Contents ARI 7 December 2005 18:14
Inflammation and AtherosclerosisGoran K. Hansson, Anna-Karin L. Robertson, and Cecilia Söderberg-Nauclér � � � � � � � � � 297
Lung Cancer PreneoplasiaIgnacio I. Wistuba and Adi F. Gazdar � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 331
Pathogenesis of Nonimmune GlomerulopathiesChristopher Kwoh, M. Brendan Shannon, Jeffrey H. Miner, and Andrey Shaw � � � � � � � � 349
Spectrum of Epstein-Barr Virus–Associated DiseasesJ.L. Kutok and F. Wang � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 375
Calcium in Cell Injury and DeathZheng Dong, Pothana Saikumar, Joel M. Weinberg,
and Manjeri A. Venkatachalam � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 405
Genetics of Soft Tissue TumorsMatt van de Rijn and Jonathan A. Fletcher � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 435
Severe Sepsis and Septic Shock: The Role of Gram-NegativeBacteremiaRobert S. Munford � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 467
Proteases in Parasitic DiseasesJames H. McKerrow, Conor Caffrey, Ben Kelly, P’ng Loke, and Mohammed Sajid � � � � 497
INDEX
Subject Index � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 537
Contents vii
Ann
u. R
ev. P
atho
l. M
ech.
Dis
. 200
6.1:
63-9
6. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by C
APE
S on
05/
26/0
9. F
or p
erso
nal u
se o
nly.