the pathway for mhcii-mediated presentation of endogenous
TRANSCRIPT

IntroductionThe location of antigens in distinct intracellular compartmentsof antigen-presenting cells (APCs) influences their proteolyticprocessing as well as access to major histocompatibilitycomplex (MHC) molecules and presentation to T cells. MHCIand MHCII molecules are directed to different intracellularcompartments for sampling peptide antigens (Neefjes et al.,1990; Peters et al., 1991). It is probable that pathways by whichMHC molecules acquire peptides will have a significant impacton generation of peptide diversity that will be ultimately sensedby the T cell receptor. Thus, understanding the pathwaysinvolved in peptide generation and loading is an extremelyimportant issue from an immunological perspective. This isespecially relevant for the dissection of the mechanism ofgeneration of an immune response against foreign proteins anddevelopment of tolerance against self proteins.
Peptides generated in the cytosol by proteasomaldegradation are generally thought to be presented on MHCI.These peptides are first transported across the endoplasmicreticulum (ER) membrane by the transporter associated withpresentation (TAP), for loading on to MHCI in the ER lumen(Rock and Goldberg, 1999; York and Rock, 1996). Peptide-MHCI interaction is expected to take place during MHCIassembly, and is required for MHCI export out of the ER(Cresswell et al., 1999). In contrast, the generation and loading
of peptides on MHCII molecules which present to the otherclass of T cells, namely the CD4 T cells, is believed to takeplace in endosomes and involves peptides derived fromproteins and other antigens delivered from the outside viaendocytic mechanisms. The apparent dichotomy between thesources of peptides for loading onto MHCI and MHCII isreinforced because newly synthesized MHCII molecules areprotected from peptide loading in the ER and Golgi by theinvariant chain (Ii) (Cresswell, 1994; Cresswell, 1996) whichalso sorts associated MHCII molecules from the Golgi to endo-lysosomal compartments (Bakke and Dobberstein, 1990).
Immunocytochemistry, subcellular fractionation andimmunoelectron microscopy approaches have identifiedMIIC or CIIVs as the canonical loading compartments fornewly synthesized MHC class II (Neefjes, 1999; Peters et al.,1991). These are sites where MHCII molecules accumulate,and it is often argued that MIICs (or CIIVs) are a collectionof late endocytic compartments that contain the necessaryproteins for efficient peptide loading of MHCII molecules(Geuze, 1998; Neefjes, 1999). It is in the acidic milieu ofthese endosomal compartments that Ii-peptide exchange isfacilitated by the endosomally localized peptide exchangecatalyst H-2M in mice or HLA-DM in humans (Martin et al.,1996; Miyazaki et al., 1996). Peptide-MHCII complexes mayalso be generated when cell surface MHCII molecules recycle
4219
Antigen-presenting cells (APCs) are expected to presentpeptides from endocytosed proteins via majorhistocompatibility complex (MHC) class II (MHCII)molecules to T cells. However, a large proportion ofpeptides purified from MHCII molecules are derived fromcytosolic self-proteins making the pathway of cytosolicpeptide loading onto MHCII of critical relevance in theregulation of immune self-tolerance. We show that peptidesderived from cytoplasmic proteins either introduced orexpressed in the cytoplasm are first detectable as MHCII-peptide complexes in LAMP-1+ lysosomes, prior to theirdelivery to the cell surface. These peptide-MHC complexesare formed in a variety of APCs, including peritonealmacrophages, dendritic cells, and B cells, and are able toactivate T cells. This process requires invariant chain (Ii)-
dependent sorting of MHCII to the lysosome and theactivity of the molecular chaperone H-2M. This pathway isindependent of the ER resident peptide transportercomplex TAP and does not take place by cross-presentationfrom neighbouring cells. In conjunction with our earlierresults showing that these peptides are derived by cytosolicprocessing via the proteasome, these observations provideevidence for a ubiquitous route for peptide transport intothe lysosome for the efficient presentation of endogenousand cytoplasmic proteins to CD4 T cells.
Supplemental data available online
Key words: Antigen presentation, MHCII, Cytosolic processing,Dendritic cells
Summary
The pathway for MHCII-mediated presentation ofendogenous proteins involves peptide transport to theendo-lysosomal compartmentAadish Dani 1, Ashutosh Chaudhry 2, Paushali Mukherjee 2, Deepa Rajagopal 2, Sumeena Bhatia 2,Anna George 2, Vineeta Bal 2, Satyajit Rath 2,* and Satyajit Mayor 1,*1National Centre for Biological Sciences, UAS-GKVK Campus, Bellary Road, Bangalore 560065, India2National Institute of Immunology, Aruna Asaf Ali Road, New Delhi 110067, India*Authors for correspondence (e-mail: [email protected]; [email protected])
Accepted 22 April 2004Journal of Cell Science 117, 4219-4230 Published by The Company of Biologists 2004doi:10.1242/jcs.01288
Research Article

4220
to early endosomes (Pathak and Blum, 2000; Pinet and Long,1998).
A breakdown of the MHCI and MHCII peptide-compartment sampling dichotomy (MHCI, intracellular versusMHCII, extracellular) is apparent from the finding that a largeproportion of peptides extracted from purified MHCII appearto be from endogenous sources (Chicz et al., 1993; Rudenskyet al., 1991a), and many of them are derived from self-cytosolicproteins (Dongre et al., 2001). This suggests that this mode ofsampling pathway is likely to be important in establishing self-tolerance in CD4 T cells, a breakdown in which can lead toautoimmune disease.
We have previously shown that cytosolically deliveredproteins as well as an endogenous trans-membrane protein, I-Eα, are efficiently presented in a MHCII-restricted manner toCD4 T cells (Mukherjee et al., 2001); other examples of thistype of presentation have also been reported (Bonifaz et al.,1999; Lich et al., 2000). The exact nature of the peptide-MHCII complexes differ from those generated via endosomalprocessing routes in their ability to be presented to specificsubsets of T-cell clones (Barlow et al., 1998). Data from ourlaboratories has showed that intracellular proteins are likely tobe processed cytosolically by the proteasome for thegeneration of peptides to be presented on MHCII (Mukherjeeet al., 2001).
A specific peptide-MHCII complex, Eα52-68-I-Ab, derivedfrom a peptide fragment of amino acids 52-68 oftransmembrane protein I-Eα (Eα52-68), is detectable in sub-cellular fractions containing MIIC-like compartments of a Bcell line (Rudensky et al., 1994). These compartments, whichalso contain HLA-DM and lysosome-associated-membraneprotein 1 (LAMP-1), are multivesicular and multilamellar.Disruption of endosomal acidification using lysosomotropicagents such as chloroquine, or Golgi transport via monensin,blocks the presentation of cytosolic, secreted or ER residentforms of ovalbumin (OVA) and HEL proteins transientlyexpressed in APCs (Bonifaz et al., 1999; Malnati et al., 1992;Michalek et al., 1992). Together, these data indicate a role forendosomes and biosynthetic transport in the formation ofpeptide-MHCII complexes derived from endogenous proteins,similar to the requirement for exogenous proteins. However, adisruption of lysosomal pH affects several lysosomal functionssuch as lysosomal proteases, peptide loading on MHCII andintracellular transport (Clague et al., 1994; Mellman et al.,1986), making the effects of such treatments on antigenpresentation difficult to interpret. Furthermore, intracellularorganelles where peptides from endogenous cytosolic proteinsare loaded on MHCII remain unidentified.
We have examined the pathway of MHCII-restrictedpresentation of cytosolically introduced protein, constitutiveendogenous trans-membrane protein and cytoplasmicallyexpressed protein. All these proteins contain the peptidesequence Eα52-68. We have localized the subcellularcompartment where the peptide-MHC complex (Eα52-68-I-Ab)forms in a variety of APCs, including bone marrow deriveddendritic cells (BMDCs), macrophages or peritoneal exudatecells (PECs) and a macrophage cell line, BMC-2. Our data areconsistent with MHCII-restricted presentation of cytosolic andendogenous proteins originating with peptide transport directlyinto late endosomal compartments colocalized with LAMP-1and cathepsin-D, for loading onto MHCII molecules.
Materials and MethodsMiceThe mouse strains and genotypes used – C3H/HeJ (H-2k), C57BL/6(H-2b), B10.A(5R) (H-2i5), H-2Mα–/–, Ii–/– (from the JacksonLaboratory, Bar Harbor, USA) and Tap1–/– (gift from Dr L. van Kaer,Vanderbilt University, Nashville, USA) – were bred in the smallanimal facility of the National Institute of Immunology (New Delhi,India) and the animal facility of the National Centre for BiologicalSciences (Bangalore, India), and used for experiments when 6-12weeks old. All experiments were done with the approval of theInstitutional Animal Ethics Committees.
AntigensA recombinant fusion protein (GST-Εα52-68-myc) consisting of GST,aa residues 52-68 of the mouse I-Eα sequence (Eα52-68) and theoligopeptide sequence of the c-myc protein recognized by the 9E10monoclonal antibody was made in Escherichia colifrom the plasmidpGEX-2T with IPTG-inducible expression and was affinity purifiedover glutathione-Sepharose beads (data not shown). Ovalbumin(Sigma, St Louis, USA) was maleylated with maleic anhydride(Sigma) at alkaline pH as previously described (Mukherjee et al.,2001), dialysed against PBS and the degree of maleylation wasestimated as the loss of free ε-amino groups measured bytrinitrobenzenesulfonic acid. Maleylated proteins were used onlywhen maleylation was >90%. All proteins were dialysed against PBSto remove small degradation products and this was additionallyconfirmed for the lack of presentation to T cells by fixed APCs (datanot shown) before use. The Εα52-68-GFP recombinant DNA was madeby synthesizing an oligonucleotide containing a start codon andsequence encoding for amino acids (aa) 52-68 of the I-Eα sequence,with flanking BglII and BamHI restriction endonuclease sites. Thiswas cloned in frame with eGFP in eGFP-N3 vector (Clontech-BDBiosciences, Palo Alto, CA, USA), and confirmed by sequencing.
Cell lines and primary cell culturesThe MHCI (H-2Kb)-restricted OVA-specific T cell transfectant line B3was a gift from Drs K. Hogquist and M. Bevan (University ofWashington, Seattle, USA). The MHCII (I-Ab)-restricted OVA-specificT cell hybridoma 13.8 was generated from OVA-immunized C57BL/6mice and characterized (data not shown). The I-Ab-restricted T cellhybridoma specific for an I-Eα peptide (aa 52-68; Εα52-68), 1H3.1, andthe I-Ab-expressing macrophage cell line, BMC-2 were gifts from DrC. A. Janeway, Yale University, New Haven, USA.
All macrophage cells unless otherwise mentioned were plastic-adherent peritoneal resident macrophages or PECs. Non-adherentmouse bone marrow cells were cultured with either monocyte colonystimulating factor (M-CSF; 30% L929 fibroblast-conditioned mediumas M-CSF source) or recombinant granulocyte-monocyte colonystimulating factor (GM-CSF; Pharmingen, San Diego, CA, USA),respectively, for 9 days, with growth factor replenishment at 3, 5 and7 days. Tightly adherent cells were excluded from the GM-CSF-containing cultures on day 7. Live cells growing out on day 9 wereused as a source of bone marrow-derived DCs (DC).
AntibodiesMonoclonal antibodies; Y-Ae (anti-Εα52-68-I-Ab), Y3P (anti-I-Ab)were detected using a goat anti-mouse IgG-Fc-specific polyclonalantibody (Jackson ImmunoResearch, USA) conjugated with Alexa-488 or Alexa 568 fluorophores (Molecular Probes). Polyclonalantibodies; rabbit anti-TGN38 (gift from Dr K. Stanley (HeartResearch Institute, Sydney, Australia), rabbit anti-ERp57 (Dr U. Tatu,IISc Bangalore, India), rabbit anti-mannose-6-phospate receptor(MPR-300, from Dr Peter Schu, Georg-August-University,Goettingen, Germany) were detected with a polyclonal donkey anti-
Journal of Cell Science 117 (18)

4221Lysosomal loading of cytosolic peptides on MHCII
rabbit IgG (Jackson) conjugated with Alexa-568 flurophore. Rat anti-mouse LAMP-1 (BD-Pharmingen, USA) was used as a primaryantibody labelled with Alexa-568 fluorophore.
Transfections and cytoplasmic loadingsCells were transfected using Fugene 6™ (Roche, Indianapolis, IN)according to the manufacturer’s protocol, with 6-10 µg of plasmidDNA for 2×106 cells and stained for Y-Ae antibodies. Apart fromindicated cell lines, primary macrophages and DCs grown from mousebone marrow were also transfected using similar protocols.
To assess for the involvement of inhibitors of PI 3-kinase in thegeneration of peptide-MHC complexes, BMC-2 cells were transfectedwith Εα52-68-GFP construct and 2 hours post-transfection were treatedwith titrating doses of PI 3-kinase inhibitor, LY294002 (1 µM-50µM), or autophagy inhibitor, 3-methyladenine (1 mM-10 mM). 20hours post-transfection, cells were stained with Y-Ae to determinesurface levels of Eα52-68-IAb complexes.
Proteins were delivered into the cytosol using osmotic lysis ofpinosomes as described previously (Mukherjee et al., 2001). Briefly,APCs were incubated in hypertonic serum-free Dulbecco’s modifiedEagle’s medium (DMEM) with 0.5 M sucrose, 10% polyethyleneglycol 800, and 10 mM Hepes containing antigenic protein for 10minutes at 37°C, followed by washing and incubation in isotonicserum-free DMEM for 5 minutes. Cells were again washed andchased further in serum containing DMEM before being fixed withparaformaldehyde. APCs were chased for 15 minutes (after the 10minute pulse), for detection of intracellular peptide-MHC complexesand chased for 1 hour when used to stimulate T cells. Post-fixation,APCs were washed, counted and used in T cell stimulation assays.Exogenous pulses of protein were done similarly, except in isotonicDMEM, thus, resembling a fluid phase uptake protocol.
Proteins were also delivered into the cytosol using the Chariot™transfection reagent according to the manufacturer’s instructions.Briefly, 1 mg of GST-Εα52-68-myc was incubated with 10 µl ofChariot™ reagent for 30 minutes to allow complex generation. Thismixture was then overlayered on 1-2×106 cells that were kept on ice.The cells were incubated on ice for a further 15 minutes before thecomplexes were washed off with PBS and the cells were shifted to37°C. Loading of streptavidin conjugated to FITC (SA-FITC) wasdone in the same way except that the concentration was 50 µg/ml.
Imaging and image processingPECs were adhered on glass coverslip bottom dishes, immediatelyafter removing from the animal. After antigen pulsing, cells were fixedwith 2% paraformaldehyde (15 minutes), permeabilized with 0.1%saponin (20 minutes) and stained with antibodies. Confocal imagingwas carried out on a Bio-Rad MRC-1024 confocal microscope (Bio-Rad Microsciences, UK) equipped with factory set dichroics and aKrypton-Argon laser. Images were processed using MetaMorphsoftware (Universal Imaging, PA, USA) and corrected for cross-talkonly in the images in Fig. 4 and Fig. 7 (E and F). For this purposesingle Alexa-488 labelled cells were scanned at the same settings oflaser power and gain as the sample to be imaged, and images of greenfluorescence and spillover in the red channel were collected; underthese conditions there was no spillover fluorescence of the fluorophoreimaged in the red channel into the green. The percentage/fractionspillover was calculated by dividing the pixel value for the red imageby that for the green and multiplying by 100. This was of the order 3-10%. This fraction of the signal in the green channel was subtractedfrom the image in the red-channel and the corrected images were usedfor further analyses. Cross-talk correction was not required for theother images since under the imaging conditions used there was nodetectable cross-talk. Images were acquired on a Nikon TE 300inverted microscope, 60×, 1.4 NA objective lens, using the Bio-Radacquisition software as described in detail elsewhere (Sabharanjak et
al., 2002). Wide-field fluorescence microscopy was done using similaroptics and images collected via a charge-coupled device camera(Princeton Instruments, Princeton, NJ, USA) using MetaMorphacquisition software (Universal Imaging, West Chester, PA, USA). Allimages were finally processed in Adobe Photoshop software for output.
Flow cytometryBMC-2 cells were transfected with eGFP or Εα52-68-GFP usingFugene6 according to manufacturer’s instructions, and were stainedwith Y-Ae antibody on ice and analysed on a BD-LSR flow cytometer(Becton Dickinson).
Antigen presentation assaysMacrophages in suspension were either exogenously pulsed orcytosolically loaded with various antigens. They were allowed toprocess antigens at 37°C for 1 hour and then fixed with 1%paraformaldehyde (Sigma) for 1 minute, washed and used as APCsin T cell stimulation assays. To assess bystander presentation, H-2k
macrophage APCs (Ag+) were first pulsed with antigen as above.After rigorous washing, these APCs were subsequently incubatedwith an equal number of unpulsed H-2b APCs (–Ag) prior to fixationand subsequent use as mixed APCs in T cell stimulation assays.
T cell lines (1-10×104 cells/well) were stimulated with titratedconcentrations of antigen-pulsed APCs in triplicate cultures in 200 µlof DMEM with 10% FCS, antibiotics, L-glutamine and 0.5 mM 2-MEin 96-well flat-bottom plates (Falcon, Franklin Lakes, USA) asindicated. Culture supernatants were collected 24-36 hours later andused to estimate the IL-2 induced by stimulating the IL-2-dependentcell line CTLL-2 (1×104 cells/well) with them, incubating for 24 hours,and pulsing the plates with 0.5 µCi per well of [3H]thymidine (NENLife Science, Boston, USA) for 12-16 hours to measure the proliferativeresponses. Plates were harvested onto glass-fibre filters for scintillationcounting (Betaplate; Wallac, Turku, Finland). Data are shown asproliferation (mean±s.e.m.) observed in triplicate CTLL-2 cultures. IL-2 secreted in the culture supernatant was also estimated by an IL-2-specific cytokine ELISA (R&D Systems, Minneapolis, USA), and thedata are plotted as Absorbance Units at 450 nm or 490 nm. Alternately,stimulation of the T cell line 13.8, which carries a transfected constructcapable of expressing β-galactosidase under the consensus IL-2-promoter, was also assayed by detecting the induction of expression ofintracellular β-galactosidase in them by an enzyme detection assay oncell lysates, data for which are plotted as absorbance units at 570 nm.
ResultsCytoplasmically expressed proteins are capable ofMHCII-restricted presentation of the same epitope asendogenously expressed proteins or proteins loaded viaosmotic lysis of pinosomesPreviously (Mukherjee et al., 2001), we detected peptide-MHCcomplexes derived from proteins introduced into the cytoplasmof diverse APC via osmotic lysis of pinosomes using peptide-MHCII-specific T cell lines, 1H3.1 [for the Εα52-68-I-Ab
complex (Murphy et al., 1992)] and 13.8 [a MHCII (I-Ab)-restricted OVA-specific T cell hybridoma]. In the workpresented here we have used a monoclonal antibody, Y-Ae,directed against a specific MHC-peptide complex, Εα52-68-I-Ab
complex (Rudensky et al., 1991b), to study the formation ofMHC-peptide complexes in intracellular compartments. BothY-Ae and 1H3.1 T cells are specific for the Eα52-68-I-Ab
complex, a major self peptide-MHCII complex in cellsexpressing I-E and I-Ab (Rudensky et al., 1991a). In addition,since we had used osmotic lysis of pinosome protocol to deliver

4222
proteins into the cytoplasm, we wished to rule out the possibilitythat these peptide-MHC complexes detected thus far are fromlocations other than the cytosol. For example, these peptide-MHCII complexes could arise by incomplete release of proteins
into the cytoplasm (during osmotic lysis of pinosomes) or byendosomal degradation of the transmembrane proteins (in cellsexpressing I-Ab and I-Eα). For this purpose we expressed Eα52-
68eGFP, a fusion protein containing the Eα52-68 fragment of I-Eα in the cytoplasm of I-Ab expressing APCs, the BMC-2 cellline. Y-Ae staining was detected by flow cytometry specificallyin cells that expressed Eα52-68-eGFP but not in cells expressingonly eGFP (Fig. 1A). This is similar to the situation in whichthe Y-Ae+ complexes are detected on primary macrophage cellspulsed either cytosolically by osmotic lysis of pinosomes orexogenously via the fluid phase with GST-Eα52-68-myc fusionprotein (Fig. 1B). The complexes generated in BMC-2 cellsderived by transfected Eα52-68-eGFP were capable of elicitinga T cell response from 1H3.1 T cells (Fig. 1C), similar to thatobtained from GST-Eα52-68-myc fusion protein delivered viathe fluid phase (Fig. 1C) or cytoplasmically delivered viaosmotic lysis of pinosomes (Mukherjee et al., 2001). Thus,expression of recombinant Eα52-68containing fusion proteins inthe cytosol of APCs specifically leads to presentation-competent peptide-MHCII molecules, confirming that peptidesderived from cytosolically located proteins are also processedand presented on MHCII.
MHCII-restricted presentation of cytosolic antigen is notmediated through extra-cellular peptide regurgitationThe detection of peptides derived from endogenouslyexpressed proteins or cytoplasmically introduced proteinssuggest two possibilities for loading onto MHCII. Peptidesfrom the cytoplasm may be regurgitated onto bystander APCs,alternatively peptides may be loaded onto autonomous MHCIIby hitherto uncharacterized mechanisms. To distinguishbetween these possibilities we investigated whether peptidesfrom cytosolic proteins are delivered into the extra-cellularmedium, and subsequently bind MHCII either at the cellsurface or in a conventional endosomal compartment. Such apathway has been demonstrated in some instances involvingMHCI-mediated presentation (Harding and Song, 1994).
To examine the role of bystander presentation (or cross-presentation), macrophages from mice of MHCII haplotype H-2k were cytosolically loaded with native OVA via osmotic lysisof pinosomes as previously described (Moore et al., 1988;Mukherjee et al., 2001). They were then co-incubated withbystander macrophages from mice with a different MHCIIhaplotype (H-2b) and the response of the OVA-specific H-2b-restricted 13.8 T cell line was tested (Fig. 2). No T cellstimulation was observed on bystander APCs [H-2b APCs(–Ag) incubated along with H-2k APCs (+Ag)], in contrastto the result when H-2b APCs were themselves loadedcytosolically with OVA (compare circles with squares inFig. 2). These results show that regurgitation (bystanderpresentation or cross-presentation) is unlikely to account forthe processing of cytosolic proteins for MHCII-restrictedpresentation via bystander APCs.
Peptide-MHC complexes from cytosolically locatedproteins as well as an endogenous trans-membraneanchored protein accumulate in a late lysosomalcompartmentWe next used the monoclonal antibody Y-Ae to investigate the
Journal of Cell Science 117 (18)
Fig. 1.Peptide-MHCII complexes are formed and presented to Tcells from a cytosolically expressed as well as from a cytosolicallydelivered protein. (A) BMC-2 cells in 6-well dishes were transfectedwith 10 µg of either Eα52-68-eGFP or eGFP (control) DNA andstained 24 hours later with the Y-Ae antibody. Fluorescence of GFPand Y-Ae was analysed by flow cytometry. Data are shown as two-colour plots and as single-parameter graphs for Y-Ae fluorescence ofappropriately gated eGFP+ and eGFP– cells as indicated. Shadedarea under curve, untransfected cells; thin lined curves, eGFPtransfected cells; thick lined curves, Eα52-68-eGFP transfected cells.(B) BMC-2 cells pulsed cytosolically (via osmotic pinosome lysismethod) or exogenously via the fluid phase with 5 mg/ml GST-Eα52-68-myc fusion protein were stained with Y-Ae after 1 hour andanalysed by flow cytometry. Shaded curve, unstained cells; thin line,unloaded cells; thick line, exogenously loaded cells; grey line,cytosolically loaded cells. (C) Responses of the 1H3.1 T cell line totitrating numbers of BMC-2 cells (APCs/well), transfected withEα52-68-eGFP DNA (squares) or eGFP DNA (triangles), orexogenously pulsed with GST-Eα52-68-myc fusion protein (circles)for 24 hours are shown as measured by IL-2 ELISA.

4223Lysosomal loading of cytosolic peptides on MHCII
intracellular localization of Eα52-68 peptide-I-Ab complexes.When the H-2b macrophage cell line BMC-2 transientlyexpresses Eα52-68-eGFP fusion protein, we find that Y-Ae+
complexes colocalized with LAMP-1, a lysosomal marker(Fig. 3A; Y-Ae staining was not detected in cells expressingeGFP alone; Fig. 3B). In macrophages from H-2b C57Bl/6mice that were loaded cytosolically via osmotic lysis ofpinosomes (Fig. 3C) or exogenously via fluid phase uptake(Fig. 3D) with the fusion protein GST-Εα52-68-myc, Y-Ae+
complexes were colocalized with intracellular compartmentscontaining LAMP-1.
In APCs from B10.A(5R) mice, the Eα52-68 peptide isderived from endogenously expressed trans-membraneanchored I-Eα protein and is loaded onto I-Ab molecules(Rudensky et al., 1991b). The localization of endogenousY-Ae-+ peptide-MHCII complexes in macrophages fromB10.A(5R) mice was also found to be predominantlyperinuclear, colocalized with LAMP-1 (Fig. 4A) and alysosomal lumenal protease cathepsin-D (Fig. 4C). Thesemolecules have been earlier shown to be marking MHCIIpeptide-loading compartments in LB27.4, a B-lymphoblastoid cell line (Geuze, 1998; Rudensky et al.,1994). Late endosomes and lysosomes are distinguished by
Fig. 2.MHCII-mediated presentation of cytosolically introducedprotein is not the result of peptide regurgitation and subsequentbystander or cross presentation. IL2 secretion was monitored todetermine specific TCR activation of the OVA-specific MHCII (H-2b)-restricted T cell line 13.8, to C57BL/6 (H-2b) and/or C3H/HeJ(H-2k) macrophages cytosolically loaded with 10 mg/ml OVA (+Ag).Bystander presentation was assessed by incubating T cell line 13.8with C3H/HeJ (H-2k) macrophages cytosolically loaded withantigens (+Ag) along with unloaded (–Ag) C57BL/6 (H-2b)macrophages. IL2-secretion was monitored by a proliferation assayas described in Materials and Methods.
Fig. 3.Peptide-MHCII complexes derived fromcytosolic proteins are formed in lysosomalcompartments in different APCs. (A,B) BMC-2cells transfected as in Fig. 2A with Eα52-68-eGFP(A) or eGFP (B) were fixed 12 hours aftertransfection, permeabilized and stained with Y-Aeand anti-LAMP-1 antibodies. (C,D) C57Bl/6macrophages were pulsed cytosolically (C) orexogenously via the fluid phase (D) with GST-Eα52-68-myc protein (5 mg/ml), chased for 15minutes, fixed, permeabilized and stained with Y-Ae and anti-LAMP-1 antibodies. Images are singlerepresentative confocal sections. All images on thevery right are respective overlays of the stainedimages of A, B, C and D. Scale bar, 10 µm.

4224
the presence of the mannose-6-phosphate receptor, MPR(Griffiths et al., 1988). MPR traffics from the Golgi to thelate endosomes, delivering mannosylated lysosomal lumenalenzymes, and is rapidly recycled back to the Golgi (Hunzikerand Geuze, 1996; Kornfeld, 1992). The steady statedistribution of the MPR is predominantly towards lateendosomes and hence it serves as a late endosomal marker.Endogenously generated Y-Ae+ complexes were notcolocalized with MPR; neither antibodies raised againstMPR-300 (Fig. 4B) or MPR-46 (data not shown) exhibitedsignificant colocalization with the Y-Ae+ structures. Thesedata locate the main site of accumulation of the Eα52-68-peptide MHCII complex to the late-lysosomal system in theseAPCs.
We next examined the localization of Y-Ae+ complexes inbone marrow-derived I-Ab+ dendritic cells either endogenouslyexpressing I-Eα (from B10.A(5R) mice) or derived fromcytoplasmically introduced GST-Εα52-68-myc. In both casesintracellular Y-Ae reactivity was detected mainly in LAMP-1-positive structures (Fig. 5).
These data show that peptide-MHCII complexes derivedfrom three different sources, cytosolic (expressed or introducedvia osmotic lysis of pinosomes), constitutively expressed-transmembrane proteins (I-Eα), or delivered via the fluid phase(or receptor mediated pathway using maleylated proteins; datanot shown), are localized in a large perinuclear compartmentthat colocalize extensively with LAMP-1 in a variety of APCs,including dendritic cells.
Rapid formation of peptide-MHCII complexes fromcytosolic proteins in macrophagesTo investigate the site of formation of peptide-MHCII complexesderived from cytosolic proteins, we used a pulse chase protocol.To deliver a short pulse of GST-Eα52-68-myc fusion protein intothe cytosol of APCs we used the osmotic lysis of pinosomesprotocol (Fig. 6B) or the Chariot™ method (Fig. 6C,D), and forexploiting the exogenous route we used the fluid phase pathway.C57Bl/6 macrophages were thus loaded with GST-Eα52-68fusion protein and allowed to process the antigen for varioustimes (the chase; Fig. 6A,B), fixed and stained with the Y-Aeantibody to detect formation of intracellular peptide-MHCIIcomplexes. We found that in all cases, Y-Ae+ peptide-MHCIIcomplexes were formed rapidly, being detectable within 5-10minutes of the start of the chase (Fig. 6A-C); the predominantdistribution was in a large perinuclear compartment colocalizedwith LAMP-1. In the case of exogenously delivered protein, Y-Ae+ complexes were seen in some smaller peripheralcompartments prior to their detection in the perinuclear LAMP-1+ compartment at extremely early times in the chase (Fig. 6A).However, cytosolically derived Y-Ae complexes were alwaysdetected only in large perinuclear compartments (Fig. 6B,C).With subsequent times of chase (20 minutes), the distribution ofthe exogenously and cytosolically derived complexes wasindistinguishable, with only a perinuclear localization seen.Further chase (60 minutes) resulted in the emptying of theinternal compartments, concomitant with the appearance of thepeptide-MHC complexes on the cell surface. Thus, peptidesfrom exogenous as well as cytosolically delivered protein arerapidly processed and loaded on MHCII within a similar timescale inside the macrophage APC, and trafficked to the cellsurface in identical fashion. The peptide-MHC complexesgenerated in such a pulse-chase paradigm are also competent tobe presented efficiently to MHCII-restricted T cells [Fig. 1C (seeMukherjee et al., 2001)].
Journal of Cell Science 117 (18)
Fig. 4.Peptide-MHC class II complexes derived from trans-membrane anchored I-Eα localize to late lysosomal compartments ofAPCs. Macrophages from B10A. (5R) mice that express I-Ab and I-Eα were fixed, permeabilized and stained with Y-Ae and antibodiesrecognizing LAMP-1, MPR (MPR-300) and cathepsin-D. Y-Aecomplexes colocalize with LAMP-1 (A) and cathepsin-D (C), but notwith MPR (B). Images are single representative confocal sections.All images on the very right are respective overlays of the stainedimages of A, B and C. Scale bar, 10 µm.
Fig. 5.Peptide-MHC class II complexes derived from endogenoustransmembrane or cytosolic protein are localized to lysosomalcompartments in bone marrow-derived dendritic cells. GM-CSFcultured BM-DCs from B10A.(5R) (A) or C57Bl/6 (B) mice wereeither directly fixed (A) or first pulsed cytosolically with GST-Eα52-68-myc protein, chased, fixed and then stained for Y-Ae, anti-LAMP-1 antibodies. Images are single representative confocalsections. All images on the very right are respective overlays of thestained images of A and B. Scale bar, 10 µm.

4225Lysosomal loading of cytosolic peptides on MHCII
To preclude any complication arising from the method ofosmotic lysis of pinosomes used to deliver proteins into thecytosol of APCs, we examined whether this methodology waslikely to disrupt endosomal trafficking in the APC. For thispurpose we preceded a 10-minute pulse of exogenouslydelivered GST-Εα52-68-myc fusion protein to C57Bl/6 APCswith an osmotic lysis procedure marked by FITC-conjugateddextran, F-Dex (Supplemental text, http://jcs.biologists.org/supplemental/). Uptake of Cy3-mBSA (used to monitor theefficiency of endocytosis after the osmotic lysis protocol) wasunaffected by the osmotic lysis protocol. The formation of Y-Ae+ complexes from exogenously delivered GST-Εα52-68-mycfusion protein was also unaffected; these were detected in asimilar perinuclear location at identical times of chase insidemacrophages in treated and untreated cells (see supplementaldata Fig. S1, http://jcs.biologists.org/supplemental/). Thus, theosmotic lysis method does not appear to have any effect onoverall endosomal trafficking and antigen presentationproperties of the APC.
The use of the Chariot™ reagent to deliver proteins intothe cytoplasm of cells as a short pulse (Fig. 6C; see alsosupplemental data Fig. S2) further validates the results of theosmotic lysis protocol (Fig. 6B). The time course of generationof these complexes is remarkably similar to the osmotic stress-induced pinosome lysis protocol (Fig. 6D), confirming thevalidity of the different means used to deliver proteins into thecytoplasm of cells. Together these results show that peptides
from cytoplasmic proteins are rapidly loaded onto MHCII inLAMP-1 compartments prior to their delivery to the cellsurface.
Role of invariant chain in MHCII-restricted presentationof cytosolic proteinA major factor involved in determining the availability ofMHCII molecules for peptide loading is the invariant chain (Ii).Invariant-chain-bound MHCII molecules intersect theendocytic pathway. The Ii chain is degraded by endosomallyresident proteases, the CLIP (Class-II-associated invariant-chain peptide) is replaced and MHCII molecules bind peptidespresent in the endosomal lumen. Ii is thought to prevent newlysynthesized MHCII molecules from being stably occupied byendogenous peptides present in the endoplasmic reticulumlumen (Cresswell, 1996). Ii also affects the intracellulartransport and distribution of MHCII molecules by changing theefficiency of their egress from the ER and delivery toendosomal compartments (Bertolino and Rabourdin-Combe,1996). Thus, in Ii-deficient mouse APCs there is a decrease inthe MHCII-mediated presentation of many but not allexogenous antigens (Viville et al., 1993).
We therefore examined the role of Ii in presentation ofcytosolic proteins on MHCII, using macrophage APCs from Ii-deficient or wild-type H-2b mice. For both exogenous andcytosolic pathways, the presentation of native OVA was lower
Fig. 6.Rapid formation ofpeptide-MHC class IIcomplexes derived fromcytosolically delivered proteinin endosomal compartmentsand their subsequentappearance on the cell surfaceof APCs. (A,B) Macrophagesfrom C57Bl/6 mice werepulsed either exogenously inisotonic medium (A) orcytosolically in hypertonicmedium (B) for 10 minuteswith 5 mg/ml GST-Eαprotein. Pulsed protein waswashed off and cells werechased for different timesindicated, fixed-permeabilizedand stained with the Y-Aeantibody to detect formationof peptide-MHC class IIcomplexes. (C,D) BMC-2cells incubated with 5 mg/mlGST-Eα52-68-myc proteincoupled to the Chariot™reagent for 15 minutes at 0°C,were washed, chased for 15minutes at 37°C, fixed-permeabilized and co-stainedfor LAMP-1 and Y-Aeantibodies to detect theintracellular Y-Ae+complexes formed at thistime. Right image is an overlay of the stained images in C. (D) A time course of detection by flow cytometry of intracellular Y-Ae+ complexesformed in BMC-2 cells after pulsing by the indicated methods and chased for different lengths of time. Scale bars, 10 µm.
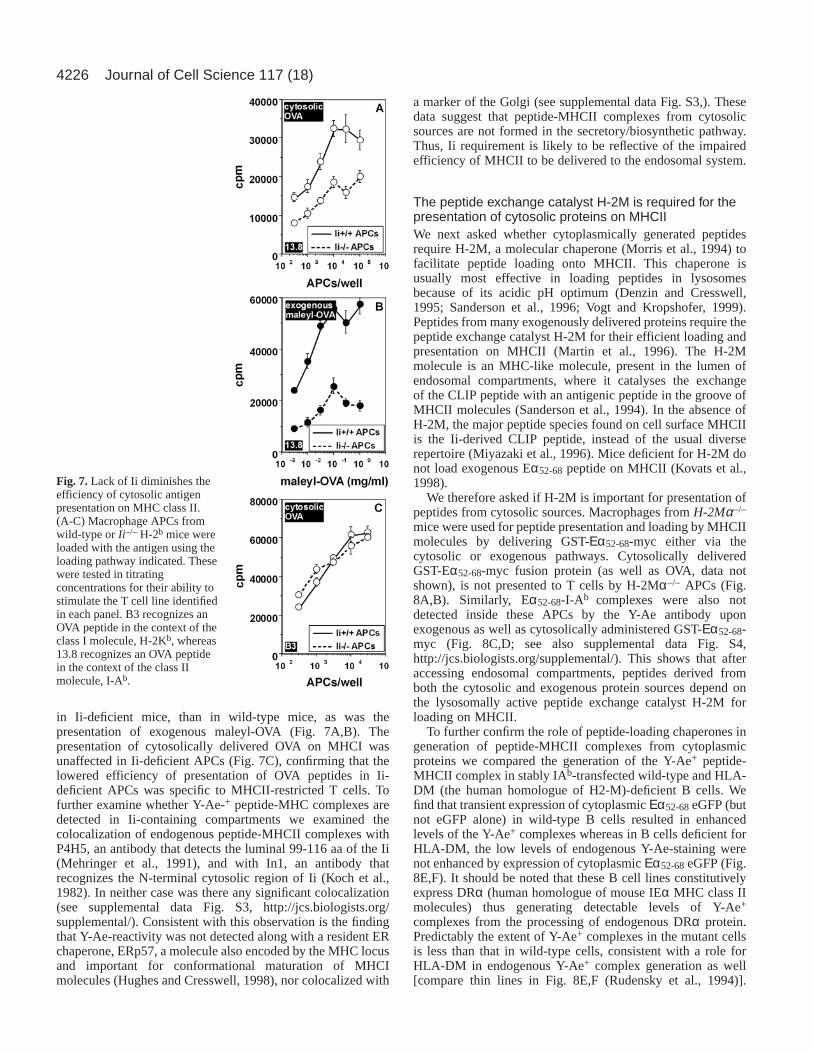
4226
in Ii-deficient mice, than in wild-type mice, as was thepresentation of exogenous maleyl-OVA (Fig. 7A,B). Thepresentation of cytosolically delivered OVA on MHCI wasunaffected in Ii-deficient APCs (Fig. 7C), confirming that thelowered efficiency of presentation of OVA peptides in Ii-deficient APCs was specific to MHCII-restricted T cells. Tofurther examine whether Y-Ae-+ peptide-MHC complexes aredetected in Ii-containing compartments we examined thecolocalization of endogenous peptide-MHCII complexes withP4H5, an antibody that detects the luminal 99-116 aa of the Ii(Mehringer et al., 1991), and with In1, an antibody thatrecognizes the N-terminal cytosolic region of Ii (Koch et al.,1982). In neither case was there any significant colocalization(see supplemental data Fig. S3, http://jcs.biologists.org/supplemental/). Consistent with this observation is the findingthat Y-Ae-reactivity was not detected along with a resident ERchaperone, ERp57, a molecule also encoded by the MHC locusand important for conformational maturation of MHCImolecules (Hughes and Cresswell, 1998), nor colocalized with
a marker of the Golgi (see supplemental data Fig. S3,). Thesedata suggest that peptide-MHCII complexes from cytosolicsources are not formed in the secretory/biosynthetic pathway.Thus, Ii requirement is likely to be reflective of the impairedefficiency of MHCII to be delivered to the endosomal system.
The peptide exchange catalyst H-2M is required for thepresentation of cytosolic proteins on MHCIIWe next asked whether cytoplasmically generated peptidesrequire H-2M, a molecular chaperone (Morris et al., 1994) tofacilitate peptide loading onto MHCII. This chaperone isusually most effective in loading peptides in lysosomesbecause of its acidic pH optimum (Denzin and Cresswell,1995; Sanderson et al., 1996; Vogt and Kropshofer, 1999).Peptides from many exogenously delivered proteins require thepeptide exchange catalyst H-2M for their efficient loading andpresentation on MHCII (Martin et al., 1996). The H-2Mmolecule is an MHC-like molecule, present in the lumen ofendosomal compartments, where it catalyses the exchangeof the CLIP peptide with an antigenic peptide in the groove ofMHCII molecules (Sanderson et al., 1994). In the absence ofH-2M, the major peptide species found on cell surface MHCIIis the Ii-derived CLIP peptide, instead of the usual diverserepertoire (Miyazaki et al., 1996). Mice deficient for H-2M donot load exogenous Eα52-68 peptide on MHCII (Kovats et al.,1998).
We therefore asked if H-2M is important for presentation ofpeptides from cytosolic sources. Macrophages from H-2Mα–/–
mice were used for peptide presentation and loading by MHCIImolecules by delivering GST-Εα52-68-myc either via thecytosolic or exogenous pathways. Cytosolically deliveredGST-Eα52-68-myc fusion protein (as well as OVA, data notshown), is not presented to T cells by H-2Mα–/– APCs (Fig.8A,B). Similarly, Eα52-68-I-Ab complexes were also notdetected inside these APCs by the Y-Ae antibody uponexogenous as well as cytosolically administered GST-Εα52-68-myc (Fig. 8C,D; see also supplemental data Fig. S4,http://jcs.biologists.org/supplemental/). This shows that afteraccessing endosomal compartments, peptides derived fromboth the cytosolic and exogenous protein sources depend onthe lysosomally active peptide exchange catalyst H-2M forloading on MHCII.
To further confirm the role of peptide-loading chaperones ingeneration of peptide-MHCII complexes from cytoplasmicproteins we compared the generation of the Y-Ae+ peptide-MHCII complex in stably IAb-transfected wild-type and HLA-DM (the human homologue of H2-M)-deficient B cells. Wefind that transient expression of cytoplasmic Εα52-68eGFP (butnot eGFP alone) in wild-type B cells resulted in enhancedlevels of the Y-Ae+ complexes whereas in B cells deficient forHLA-DM, the low levels of endogenous Y-Ae-staining werenot enhanced by expression of cytoplasmic Εα52-68eGFP (Fig.8E,F). It should be noted that these B cell lines constitutivelyexpress DRα (human homologue of mouse IEα MHC class IImolecules) thus generating detectable levels of Y-Ae+
complexes from the processing of endogenous DRα protein.Predictably the extent of Y-Ae+ complexes in the mutant cellsis less than that in wild-type cells, consistent with a role forHLA-DM in endogenous Y-Ae+ complex generation as well[compare thin lines in Fig. 8E,F (Rudensky et al., 1994)].
Journal of Cell Science 117 (18)
Fig. 7.Lack of Ii diminishes theefficiency of cytosolic antigenpresentation on MHC class II.(A-C) Macrophage APCs fromwild-type or Ii–/– H-2b mice wereloaded with the antigen using theloading pathway indicated. Thesewere tested in titratingconcentrations for their ability tostimulate the T cell line identifiedin each panel. B3 recognizes anOVA peptide in the context of theclass I molecule, H-2Kb, whereas13.8 recognizes an OVA peptidein the context of the class IImolecule, I-Ab.

4227Lysosomal loading of cytosolic peptides on MHCII
Together, these results confirm that the peptide-loadingchaperone in late endosomes is involved in the generation ofthe peptide-MHCII complex from cytoplasmically expressedproteins.
To ascertain that in H-2Mα–/– mouse cells, MHCII deliveryto the lysosomal compartment or Ii processing is normal, we
immunolocalized the CLIP peptide-MHCII complex (Fig. S4,http://jcs.biologists.org/supplemental/). Predictably we findthat the CLIP compartment was completely colocalized withY3P, an antibody that detects I-Ab-molecules (Janeway et al.,1984) in both H-2Mα–/– and control cells. However, Y-Aereactivity was not detected in H-2Mα–/– cells when GST-Εα52-68-myc was delivered into the cytosol or providedexogenously via the fluid phase. This strongly suggests that inthe absence of H-2M activity in the endosomal system, the Y-Ae+ complex is not efficiently generated from cytoplasmicallylocalized proteins.
The TAP transporter is not required for peptides fromendogenous proteins to be presented on MHCII; theroute of peptide loading on MHCII involves directtransport into an endo-lysosomal compartmentPeptides from endogenous proteins are processed in thecytoplasm and access the ER lumen via the TAP transporter,where they bind MHCI molecules (York and Rock, 1996). TheTAP transporter belongs to the ABC family of transporters andfunctions to efficiently transport peptides for loading on MHCImolecules. We hence examined the possibility that thecytosolic peptides that load on MHCII also gain access to themembrane bound organelles via the TAP transporter.
We used macrophages from Tap1–/– mice (Van Kaer et al.,1992) which are deficient for TAP1. These macrophages areunable to present cytosolic OVA to MHCI-restricted B3 T cells(Fig. 9A). They also showed lower levels of surface MHCI(data not shown), because empty MHCI molecules do not exitthe ER efficiently or last long on the cell surface (Ljunggrenet al., 1990). However, Tap1–/– APCs presented cytosolicallydelivered proteins, GST-Εα52-68-myc and OVA (data notshown) on MHCII to the respective T cells at unalteredefficiencies compared to wild-type cells (compare Fig. 9B,C).Peptide-MHCII complexes from cytosolically delivered GST-Εα52-68-myc are also found localized to LAMP-1+ endosomalcompartments in macrophages from Tap1–/– mice (Fig. 9D). Inaddition, the extent of generation of Y-Ae+ complexes fromtransiently transfected Eα52-68-eGFP in bone marrow-derivedmacrophages from Tap1–/– mice appeared no different fromthose formed in the wild-type mice (Fig. 9E). These data showthat, unlike MHCI-restricted presentation, MHCII-restrictedpresentation of cytosolic antigens is Tap independent.
Cytoplasmic loading onto MHCII is insensitive to potentinhibitors of macroautophagyTo examine whether macroautophagy is a plausible mechanismfor cytoplasmically expressed proteins to gain access to thelumenal peptide binding site of MHCII, BMC-2 cells,transfected with Εα52-68-eGFP were treated with inhibitors ofPI 3-kinase activity, wortmannin (WT), LY294002 or 3-methyladenine (3-MA) (Stromhaug and Klionsky, 2001). Noneof the inhibitors reduced the generation of Y-Ae+- peptide-MHCII complexes (Fig. 10). However, inhibition of PI 3-kinase did induce a vesiculation of the LAMP-1 stainingpattern (data not shown), indicative of inhibition of PI 3-kinaseactivity inside the cell. These results imply the lack ofinvolvement of PI 3-kinase dependent macroautophagy in thisprocess of cytoplasm to lysosome transport.
Fig. 8.H-2M is required for cytosolic antigen presentation on MHCclass II. (A,B) Macrophage APCs from wild-type or H-2Mα–/– H-2b
mice were loaded with OVA, by the fluid phase loading pathway (A,OVA Exogenous) or via osmotic lysis of pinosomes (OVA,Cytosolic) and tested in titrating concentrations for their ability tostimulate the MHCII restricted T cell line 13.8 by monitoring IL2secretion in an ELISA reaction, absorbance at 570 nm, A 570.(C,D) Total cell associated Y-Ae and MHCII I-Ab-specific antibody,Y-3P, fluorescence were quantified from APCs derived from H-2Mα–/– (grey circles) and control C57/Bl6 (black circles) mice, eitherloaded via the fluid phase (C) or by the osmotic lysis of pinosomes(D) show that Y-Ae-positive peptide-MHCII complexes are absent intheH-2Mα–/– mice. (E,F) Human B cell lines (T1-IAb) or its HLA-DM-deficient mutant counterpart (T2-IAb) stably transfected withIAb were transiently transfected with eGFP (thin line) or withEα52-68-eGFP (thick line) as indicated and stained 20 hours later withY-Ae and analysed by flow cytometry after gating for eGFP+ cells.The shaded area under the curve represents isotype control signalsfor the Y-Ae antibody staining.

4228
DiscussionWe present evidence here that a novel peptide transportmechanism is likely to be involved in the translocation ofpeptides derived from cytosolic and endogenous trans-membrane anchored proteins. We have shown earlier thatMHCII-mediated presentation of proteins introduced into thecytosol by osmotic lysis of pinosomes depends on cytosolicproteolytic mechanisms such as the proteasome (Mukherjee etal., 2001). We have examined the route and localisation ofpeptide transport and loading onto MHCII molecules.
It is possible that degraded peptides from dead or dying cellsmay be loaded on surface or recycling MHCII molecules onneighbouring cells. Using bystander APC assays, we show thatthis is not the mode of peptide loading on MHCII forcytosolically delivered proteins. It is hence possible thatloading occurs in the ER lumen after peptide transport fromthe cytosol using mechanisms related to those used by MHCI.However, we find that peptide-MHCII complexes fromendogenous proteins are not formed in the ER or the Golgicomplex. In addition, the TAP transporter is not required forloading of peptides from cytosolic proteins on MHCII.
Peptide derived from proteins delivered into or expressed inthe cytosol, and those derived from trans-membrane anchoredproteins are loaded on MHCII in a large perinuclear
compartment of the APC. This compartment contains markersof lysosomes, LAMP-1 and cathepsin-D, but does not containa marker of the late endosomes (MPR), confirming thelysosomal nature of the compartment where peptide loading onMHCII occurs in these cells.
A major facilitator of peptide loading on MHCII in theendo-lysosomal compartment is the H-2M molecule. Previousreports (Kovats et al., 1998) as well as our data here show thatthe endogenous pathway of MHCII-mediated presentation ofcytosolic proteins is dependent on H-2M, similar to the classicpathway of loading peptides derived from exogenous antigens.Peptides from cytosolic proteins required the peptide-loadingchaperone H-2M or HLA-DM as seen in experiments usingAPCs from mice lacking H-2M or in the human B cell linedeficient in HLA-DM. Together with the almost immediateimmunolocalization of the peptide-MHCII complex in thelysosome after a short cytosolic pulse, these data show thatwhile the peptides from endogenous proteins are generated inthe cytosol of APCs, they are rapidly and possibly directlytransported into lysosomal compartments followed by a H-2M-dependent loading onto MHCII. Our data are thusconsistent with a model of cytoplasmic antigen processingcoupled to rapid transport into the endo-lysosomalcompartment, and suggest a novel mechanism of peptide
Journal of Cell Science 117 (18)
Fig. 9.TAP transporter is notrequired for cytosolic antigenpresentation on MHC class II.(A-C) Responses of the (A) OVA-specific MHC class I-restricted Tcell line, B3Z, and (B,C) the Eα-specific MHC class II-restricted Tcell line, 1H3.1. APCs used wereeither wild-type or TAP-1–/– H-2bmacrophages as indicated. Antigensindicated were loaded eithercytosolically or exogenously andused to stimulate the relevant T celllines in titrating numbers. IL2-secretion was monitored by ELISAas indicated in the text.(D) Macrophage APCs from TAP-1–/– mice were loaded cytosolicallywith GST-Eα52-68-myc and stainedwith the Y-Ae and anti-LAMP-1antibodies. Image on the rightshows overlay; green, Y-Ae; red,anti-LAMP-1. Scale bar, 10 µm.(E) Bone marrow-derivedmacrophages from TAP-1–/– andwild-type mice were transientlytransfected with Eα52-68-eGFP oreGFP and stained for Eα-specificMHCII complexes with Y-Aeantibodies and detected by flowcytometry. Thin lined curve showsthe signal from TAP-1–/–, thicklined curve from wild-type cells andshaded area represent isotypecontrol signals for Y-Ae antibodystaining.

4229Lysosomal loading of cytosolic peptides on MHCII
transport for the efficient presentation of endogenous cellularproteins to CD4 T cells.
How then do peptides generated from cytosolic sources entervesicular compartments and reach the endo-lysosomal regionfor binding MHCII? Peptides generated via proteasomaldegradation in the cytosol will have to cross the lysosomalmembrane barrier. We discuss two possibilities. One involvestransport via peptide transporters and the other via multiplemechanisms that come under a umbrella term ‘autophagy’(Cuervo, 2004; Klionsky and Ohsumi, 1999).
While our data shows that TAP, the ER resident peptidetransporter from the ABC family is not involved in this peptidetransport process, TAP is a member of the transmembrane ABCfamily of transporters, and other members of this family may beinvolved. The ABC transporter ABC-1 has been claimed to affect
the transport of the leaderless protein interleukin-1β from thecytosol to the lysosome in monocyte cells (Andrei et al., 1999;Hamon et al., 1997). ABC-B9, another promising candidatetransporter, phylogenetically related to TAP, is localized toLAMP-1-containing lysosomes, although its function is as yetunknown (Zhang et al., 2000). Other as yet unidentified ABCtransporters may also be present on lysosomal membranes.Molecular tools would be helpful in addressing whether thefunction of any of the ABC transporters is necessary intranslocation of proteins or peptides into lysosomes.
The MHCII peptide loading compartments exhibit amultivesicular or multilamellar ultrastructure in many cells.Another mechanism that can result in inclusion of cytoplasmicproteins is the process of autophagy, of which there are manyforms (Cuervo, 2004). The lack of an effect of the selectiveautophagy inhibitor, 3-methyladenine, and inhibitors of PI 3-kinase on the generation of Y-Ae+ complexes, argues againstany role for macroautophagy. An alternative autophagicprocess, described as ‘chaperone mediated autophagy’ (Cuervoand Dice, 1996; Cuervo et al., 2003) is up-regulated understress, serum withdrawal, and involves the selective uptake ofcytosolic proteins into lysosomes. Uptake and processing ofOVA and Εα52-68-GFP (both of which lack KFERQ-likemotifs) via this pathway, argues against this mechanism.This leaves the constitutive (but poorly characterized)micropinocytic pathway wherein cytoplasmic proteins areincluded in multivesicular lysosomes, which could be degradedand presented by MHCII molecules (Klionsky and Ohsumi,1999). Additional, as yet unidentified mechanisms may alsoexist that are either specific to antigen presenting cells, orubiquitous to many cell types, and result in the import ofproteins and peptides directly into lysosomes.
This work was supported in part by grants from the Departmentsof Biotechnology and Science and Technology, Government of India(A.G., S.R., V.B.) and by intra-mural funds from NCBS (S.M.). TheNational Institute of Immunology is supported by the Department ofBiotechnology, Government of India. A.D. was supported by a PhDfellowship from TIFR and a Kanwal Rekhi Fellowship from the TIFREndowment Fund. S.M. is a Senior Research Fellow of the WellcomeTrust (UK), and thanks V. Sriram for critically reading the manuscript,and K. Belur and F. F. Bosphorus for inspiration.
ReferencesAndrei, C., Dazzi, C., Lotti, L., Torrisi, M. R., Chimini, G. and Rubartelli,
A. (1999). The secretory route of the leaderless protein interleukin 1betainvolves exocytosis of endolysosome-related vesicles. Mol. Biol. Cell 10,1463-1475.
Bakke, O. and Dobberstein, B. (1990). MHC class II-associated invariantchain contains a sorting signal for endosomal compartments. Cell 63,707-716.
Barlow, A. K., He, X. and Janeway, C., Jr (1998). Exogenously providedpeptides of a self-antigen can be processed into forms that are recognizedby self-T cells. J. Exp. Med. 187, 1403-1415.
Bertolino, P. and Rabourdin-Combe, C. (1996). The MHC class II-associated invariant chain: a molecule with multiple roles in MHC class IIbiosynthesis and antigen presentation to CD4+ T cells. Crit. Rev. Immunol.16, 359-379.
Bonifaz, L. C., Arzate, S. and Moreno, J. (1999). Endogenous andexogenous forms of the same antigen are processed from different pools tobind MHC class II molecules in endocytic compartments. Eur. J. Immunol.29, 119-131.
Chicz, R. M., Urban, R. G., Gorga, J. C., Vignali, D. A., Lane, W. S. andStrominger, J. L. (1993). Specificity and promiscuity among naturallyprocessed peptides bound to HLA-DR alleles. J. Exp. Med. 178, 27-47.
Fig. 10.Cytoplasmic loading onto MHCII is insensitive to reduction ofPI 3-kinase activity and 3-methyladenine treatment. BMC-2 cells weretransfected and treated with autophagy inhibitors as indicated to theright of each plot (3MA, 10 mM; LY294002, 50 µM). The generationof Y-Ae+ peptide MHCII complexes was assayed by flow cytometry at20 hours post-transfection. The numbers in the top right quadrantindicate that the percentage of eGFP-expressing cells that are alsopositive for Y-Ae+ complexes is unaffected by the different treatmentstested. Numbers in the top left quadrant indicate percentage of eGFP-expressing cells; numbers in the bottom right quadrant indicatepercentage of cells gated for positive Y-Ae signals; numbers in thebottom left quadrant indicate percentage of unstained cells.

4230
Clague, M. J., Urbe, S., Aniento, F. and Gruenberg, J. (1994). VacuolarATPase activity is required for endosomal carrier vesicle formation. J. Biol.Chem. 269, 21-24.
Cresswell, P. (1994). Assembly, transport, and function of MHC class IImolecules. Annu. Rev. Immunol. 12, 259-293.
Cresswell, P. (1996). Invariant chain structure and MHC class II function. Cell84, 505-507.
Cresswell, P., Bangia, N., Dick, T. and Diedrich, G. (1999). The nature ofthe MHC class I peptide loading complex. Immunol. Rev. 172, 21-28.
Cuervo, A. M. (2004). Autophagy: in sickness and in health. Trends Cell Biol.14, 70-77.
Cuervo, A. M. and Dice, J. F. (1996). A receptor for the selective uptake anddegradation of proteins by lysosomes. Science273, 501-503.
Cuervo, A. M., Mann, L., Bonten, E. J., d’Azzo, A. and Dice, J. F. (2003).Cathepsin A regulates chaperone-mediated autophagy through cleavage ofthe lysosomal receptor. EMBO J. 22, 47-59.
Denzin, L. K. and Cresswell, P. (1995). HLA-DM induces CLIP dissociationfrom MHC class II alpha beta dimers and facilitates peptide loading. Cell82, 155-165.
Dongre, A. R., Kovats, S., deRoos, P., McCormack, A. L., Nakagawa, T.,Paharkova-Vatchkova, V., Eng, J., Caldwell, H., Yates, J. R., III andRudensky, A. Y. (2001). In vivo MHC class II presentation of cytosolicproteins revealed by rapid automated tandem mass spectrometry andfunctional analyses. Eur. J. Immunol. 31, 1485-1494.
Geuze, H. J. (1998). The role of endosomes and lysosomes in MHC class IIfunctioning. Immunol. Today19, 282-287.
Griffiths, G., Hoflack, B., Simons, K., Mellman, I. and Kornfeld, S. (1988).The mannose 6-phosphate receptor and the biogenesis of lysosomes. Cell52, 329-341.
Hamon, Y., Luciani, M. F., Becq, F., Verrier, B., Rubartelli, A. andChimini, G. (1997). Interleukin-1beta secretion is impaired by inhibitors ofthe Atp binding cassette transporter, ABC1. Blood90, 2911-2915.
Harding, C. V. and Song, R. (1994). Phagocytic processing of exogenousparticulate antigens by macrophages for presentation by class I MHCmolecules. J. Immunol. 153, 4925-4933.
Hughes, E. A. and Cresswell, P. (1998). The thiol oxidoreductase ERp57 isa component of the MHC class I peptide-loading complex. Curr. Biol. 8,709-712.
Hunziker, W. and Geuze, H. J. (1996). Intracellular trafficking of lysosomalmembrane proteins. BioEssays18, 379-389.
Janeway, C. A., Jr, Conrad, P. J., Lerner, E. A., Babich, J., Wettstein, P. andMurphy, D. B. (1984). Monoclonal antibodies specific for Ia glycoproteinsraised by immunization with activated T cells: possible role of T cell boundIa antigens as targets of immunoregulatory T cells. J. Immunol. 132, 662-667.
Klionsky, D. J. and Ohsumi, Y. (1999). Vacuolar import of proteins andorganelles from the cytoplasm. Annu. Rev. Cell Dev. Biol. 15, 1-32.
Koch, N., Koch, S. and Hammerling, G. J. (1982). Ia invariant chain detectedon lymphocyte surfaces by monoclonal antibody. Nature299, 644-645.
Kornfeld, S. (1992). Structure and function of the mannose 6-phosphate/insulinlike growth factor II receptors. Annu. Rev. Biochem. 61, 307-330.
Kovats, S., Grubin, C. E., Eastman, S., deRoos, P., Dongre, A., van Kaer,L. and Rudensky, A. Y. (1998). Invariant chain-independent function of H-2M in the formation of endogenous peptide-major histocompatibilitycomplex class II complexes in vivo. J. Exp. Med. 187, 245-251.
Lich, J. D., Elliott, J. F. and Blum, J. S. (2000). Cytoplasmic processing isa prerequisite for presentation of an endogenous antigen by majorhistocompatibility complex class II proteins. J. Exp. Med. 191, 1513-1524.
Ljunggren, H. G., Stam, N. J., Ohlen, C., Neefjes, J. J., Hoglund, P.,Heemels, M. T., Bastin, J., Schumacher, T. N., Townsend, A., Karre, K.et al. (1990). Empty MHC class I molecules come out in the cold. Nature346, 476-480.
Malnati, M. S., Marti, M., LaVaute, T., Jaraquemada, D., Biddison, W.,DeMars, R. and Long, E. O. (1992). Processing pathways forpresentation of cytosolic antigen to MHC class II-restricted T cells. Nature357, 702-704.
Martin, W. D., Hicks, G. G., Mendiratta, S. K., Leva, H. I., Ruley, H. E.and van Kaer, L. (1996). H2-M mutant mice are defective in the peptideloading of class II molecules, antigen presentation, and T cell repertoireselection. Cell 84, 543-550.
Mehringer, J. H., Harris, M. R., Kindle, C. S., McCourt, D. W. and Cullen,S. E. (1991). Characterization of fragments of the murine Ia-associatedinvariant chain. J. Immunol. 146, 920-927.
Mellman, I., Fuchs, R. and Helenius, A. (1986). Acidification of theendocytic and exocytic pathways. Annu. Rev. Biochem. 55, 663-700.
Michalek, M. T., Benacerraf, B. and Rock, K. L. (1992). The class II MHC-restricted presentation of endogenously synthesized ovalbumin displaysclonal variation, requires endosomal/lysosomal processing, and is up-regulated by heat shock. J. Immunol. 148, 1016-1024.
Miyazaki, T., Wolf, P., Tourne, S., Waltzinger, C., Dierich, A., Barois, N.,Ploegh, H., Benoist, C. and Mathis, D. (1996). Mice lacking H2-Mcomplexes, enigmatic elements of the MHC class II peptide-loadingpathway. Cell 84, 531-541.
Moore, M. W., Carbone, F. R. and Bevan, M. J. (1988). Introduction ofsoluble protein into the class I pathway of antigen processing andpresentation. Cell 54, 777-785.
Morris, P., Shaman, J., Attaya, M., Amaya, M., Goodman, S., Bergman,C., Monaco, J. J. and Mellins, E. (1994). An essential role for HLA-DMin antigen presentation by class II major histocompatibility molecules.Nature368, 551-554.
Mukherjee, P., Dani, A., Bhatia, S., Singh, N., Rudensky, A. Y., George,A., Bal, V., Mayor, S. and Rath, S. (2001). Efficient presentation of bothcytosolic and endogenous transmembrane protein antigens on MHC class IIis dependent on cytoplasmic proteolysis. J. Immunol. 167, 2632-2641.
Murphy, D. B., Rath, S., Pizzo, E., Rudensky, A. Y., George, A., Larson,J. K. and Janeway, C. A., Jr (1992). Monoclonal antibody detection of amajor self peptide. MHC class II complex. J. Immunol. 148, 3483-3491.
Neefjes, J. (1999). CIIV, MIIC and other compartments for MHC class IIloading. Eur. J. Immunol. 29, 1421-1425.
Neefjes, J. J., Stollorz, V., Peters, P. J., Geuze, H. J. and Ploegh, H. L.(1990). The biosynthetic pathway of MHC class II but not class I moleculesintersects the endocytic route. Cell 61, 171-183.
Pathak, S. S. and Blum, J. S. (2000). Endocytic recycling is required for thepresentation of an exogenous peptide via MHC class II molecules. Traffic1, 561-569.
Peters, P. J., Neefjes, J. J., Oorschot, V., Ploegh, H. L. and Geuze, H. J.(1991). Segregation of MHC class II molecules from MHC class I moleculesin the Golgi complex for transport to lysosomal compartments. Nature349,669-676.
Pinet, V. M. and Long, E. O. (1998). Peptide loading onto recycling HLA-DR molecules occurs in early endosomes. Eur. J. Immunol. 28, 799-804.
Rock, K. L. and Goldberg, A. L. (1999). Degradation of cell proteins andthe generation of MHC class I-presented peptides. Annu. Rev. Immunol. 17,739-779.
Rudensky, A., Preston-Hurlburt, P., Hong, S. C., Barlow, A. and Janeway,C. A., Jr (1991a). Sequence analysis of peptides bound to MHC class IImolecules. Nature353, 622-627.
Rudensky, A., Rath, S., Preston-Hurlburt, P., Murphy, D. B. and Janeway,C. A., Jr (1991b). On the complexity of self. Nature353, 660-662.
Rudensky, A. Y., Maric, M., Eastman, S., Shoemaker, L., DeRoos, P. C.and Blum, J. S. (1994). Intracellular assembly and transport of endogenouspeptide-MHC class II complexes. Immunity1, 585-594.
Sabharanjak, S., Sharma, P., Parton, R. G. and Mayor, S. (2002). GPI-anchored proteins are delivered to recycling endosomes via a distinct cdc42-regulated, clathrin-independent pinocytic pathway. Dev. Cell2, 411-423.
Sanderson, F., Kleijmeer, M. J., Kelly, A., Verwoerd, D., Tulp, A., Neefjes,J. J., Geuze, H. J. and Trowsdale, J. (1994). Accumulation of HLA-DM,a regulator of antigen presentation, in MHC class II compartments. Science266, 1566-1569.
Sanderson, F., Thomas, C., Neefjes, J. and Trowsdale, J. (1996).Association between HLA-DM and HLA-DR in vivo. Immunity4, 87-96.
Stromhaug, P. E. and Klionsky, D. J. (2001). Approaching the molecularmechanism of autophagy. Traffic 2, 524-531.
Van Kaer, L., Ashton-Rickardt, P. G., Ploegh, H. L. and Tonegawa, S.(1992). TAP1 mutant mice are deficient in antigen presentation, surface classI molecules, and CD4-8+ T cells. Cell 71, 1205-1214.
Viville, S., Neefjes, J., Lotteau, V., Dierich, A., Lemeur, M., Ploegh, H.,Benoist, C. and Mathis, D. (1993). Mice lacking the MHC class II-associated invariant chain. Cell 72, 635-648.
Vogt, A. B. and Kropshofer, H. (1999). HLA-DM – an endosomal andlysosomal chaperone for the immune system. Trends Biochem. Sci. 24, 150-154.
York, I. A. and Rock, K. L. (1996). Antigen processing and presentation bythe class I major histocompatibility complex. Annu. Rev. Immunol. 14, 369-396.
Zhang, F., Zhang, W., Liu, L., Fisher, C. L., Hui, D., Childs, S., Dorovini-Zis, K. and Ling, V. (2000). Characterization of ABCB9, an ATP bindingcassette protein associated with lysosomes. J. Biol. Chem. 275, 23287-23294.
Journal of Cell Science 117 (18)