the peruvian el tor strains of vibrio cholerae o1 have a distinct ... · 1 the peruvian el tor...
TRANSCRIPT

1
The Peruvian El Tor strains of Vibrio cholerae O1 have a distinct region in the Vibrio
seventh pandemic island-II that differentiates them from the seventh pandemic strains
of other continents
Suraia Nusrin,1
Ana I. Gil, 2
* N. A. Bhuiyan, 1
Ashrafus Safa, 1
Masahiro Asakura, 3
C. F. Lanata,2 E.R. Hall,
4 H. Miranda,
5 B. Huapaya,
6 C. Vargas G,
7 M.A. Luna ,
8 D.A.
Sack, 1
Shinji Yamasaki3 and G. Balakrish Nair
1
1Enteric Microbiology Unit, Laboratory Sciences Division, ICDDR,B, Bangladesh;
2Instituto
de Investigación Nutricional, Lima, Peru; 3Graduate School of Life and Environmental
Sciences, Osaka Prefecture University, Sakai, Osaka , Japan; 4Naval Medical Research
Center Detachment, Lima, Peru; 5Instituto de Medicina Tropical e Infectologia, Facultad de
Medicina, Universidad Nacional de Trujillo, Trujillo, Peru; 6Instituto Nacional de Salud,
Lima Peru; 7CEPIS/SDE/OPS/OMS, Lima Peru;
8Oficina General de Epidemiología, Lima
Peru.
Word count:
Abstract: 253
Full Text without Abstract, References and Acknowledgement: 4277
*Correspondence to:
Ana I Gil, Investigadora Principal, Instituto de Investigacion Nutricional, Av. La Molina
1885, Lima 12, Peru, A.P. 18-0191, Lima – 18, Peru; Tel (+51-1) 349-6023; Fax (+51-1)
3496025; Email [email protected]

2
Abstract
A collection of environmental and clinical strains of Vibrio cholerae O1 isolated from
the beginning of the epidemic in 1991 to 2003 from multiple locations in Peru were examined
using a multi locus virulence gene profiling strategy. The overall results of this screening
indicated that the Peruvian strains were similar to the reference El Tor strain (N16961) of the
seventh pandemic with the only striking difference being the negative result by PCR for
VC0512 and VC0514 in the VSP-II gene cluster of all the Peruvian strains. This difference in
VSP-II was stably conserved in the Peruvian strains and this pattern was not observed in the
seventh pandemic El Tor V. cholerae O1 strains examined from Asia, Africa, Australia and
Europe. The comparison of the nucleotide sequences and other parameters of VSP-II and
flanking regions of one Peruvian strain (PERU-130) with that of VSP-II of N16961
confirmed the PCR results indicating that ORFs 20 to 24 of the Peruvian strain showed low
DNA homology (46.6%) to the corresponding region encompassing VC0511 to VC0515 of
NI6961. Based on the stable difference in VSP-II, we conclude that the Peruvian El Tor O1
strains are quite distinct from the seventh pandemic Eurasian and African strains, suggesting
that, although temporally associated with the seventh pandemic, it arose independently
probably from a nonhuman environmental source. The difference in the VSP-II lends a
distinctive stable molecular signature to the Peruvian strains that could form the basis of
tracking the origin of these strains and, therefore, of the Latin American pandemic.

3
Introduction
In 1991, after the absence of more than a century, Vibrio cholerae O1 biotype El Tor
entered into South America and caused explosive epidemics along the coast of Peru. Cholera
occurred simultaneously at different sites along the Peruvian coast, over a distance of more
than 1200 kms (32). By the end of 1992, 19 countries in Latin America had reported more
that 730,000 cholera cases and 6,300 deaths (15, 29). The epidemic, thereafter, spread to the
rest of south and Central America from Mexico to Argentina (34).
Since the entry of the El Tor biotype into Latin America, an intensive effort has been
made to understand the origin of these strains. Molecular typing methods indicate that
isolates from the 1991 Latin American epidemic are clonal and represent an extension of the
seventh pandemic strain of El Tor biotype in the Western hemisphere (36). More recent
studies have documented relatively high rate of genetic changes as shown by changing
serotypes, electrophoretic types (ET), ribotypes and PFGE types among the Latin American
strains (8, 29, 11). The current understanding is that there are a mélange of different
molecular types among the Latin American strains of V. cholerae O1 closely related to the
Asian and African seventh pandemic strains.
The past few years have witnessed many advances in our understanding of the
genome and genetics of V. cholerae. We now know that the cholera toxin (CT) genes are
encoded by a filamentous bacteriophage CTXΦ (37) and the toxin-coregulated pilus (TCP),
an important colonization factor, acts as the receptor for CTXΦ (18, 16). V. cholerae, like
other members of the genus Vibrio, has two circular chromosomes (35) and the whole
genome sequence of an El Tor V. cholerae O1 strain N16961 has shown them to be 2.96 Mb
and 1.07 Mb, respectively, in size (13). Comparative genomic analysis using a DNA
microarray showed differences in gene content between the sixth (classical biotype) and the
current seventh (El Tor biotype) pandemic strains of V. cholerae O1 and identified two

4
genomic regions designated as the Vibrio Seventh Pandemic island-I (VSP-I) and VSP-II that
are unique to seventh pandemic El Tor strains (10). VSP-I and VSP-II showed several
properties of pathogenicity islands, and are believed to be involved with the pandemic
propensity of the seventh pandemic El Tor strains. Although they share common lineage (27),
the classical strains that caused the first six pandemics are different from the El Tor strains
responsible for the on-going seventh pandemic, in that they lack the VSP-I and VSP-II (10).
In light of the new information on the molecular aspects of V. cholerae, we decided to
re-examine the strains of V. cholerae O1 isolated from the beginning of the Peruvian
epidemic in 1991 to 2003 from different locations in Peru using a PCR based multilocus
virulence gene profiling strategy. We report the identification of a distinctive stable
molecular signature in the Peruvian strains in the VSP-II not seen in other seventh pandemic
strains of V. cholerae O1 El Tor that could form the basis of tracking the origin of these
strains and, therefore, of the Latin American pandemic.
Materials and Methods
Bacterial strains.
Sixty strains of V. cholerae O1 (48 clinical and 12 environmental) isolated from Lima,
Trujillo, Lambayeque, Cajamarca, Arequipa, Ayacucho, Loreto and Ucayali in Peru from
1991 to 2003 were examined in this study. Twenty three V. cholerae O1 El Tor strains from
10 countries as shown in Table 1 were used in this study for comparison. The countries
included Australia, Bangladesh, India, Maldives, Malaysia, Myanmar, Mozambique, Macao,
Germany, and Zambia representing four continents (Asia, Africa, Australia and Europe).
V.cholerae O1 strain O395 of the classical biotype and the O1 El Tor strain N16961 were
used as standard reference strains representing each biotype. The 60 strains of V. cholerae O1

5
from Peru, the 23 strains from other countries and the two reference strains (Table 1) were
sub-cultured on gelatin agar plates, and stored at –80°C in Luria Bertini broth containing 25%
glycerol for further study.
Serotyping and Biotyping.
The serogroup of all the strains was reconfirmed by using polyvalent O1 and monoclonal
Inaba and Ogawa antisera prepared at ICDDR,B. Biotyping of the Peruvian strains was
performed using polymyxin B susceptibility (50 U), chicken cell (erythrocyte) agglutination
(CCA) and sensitivity to group IV classical and group 5 El Tor phages as described
previously (22).
Genomic DNA Isolation.
For extraction of genomic DNA, cells were harvested from 3 ml of overnight culture in LB
broth (Miller). The harvested cells were subjected to alkaline lysis by 10% SDS in the
presence of TE buffer (10mM Tris-HCl; 1mM EDTA, pH 8.0). The cells were then treated
with freshly prepared Proteinase K (final concentration 100 µg/ml in 0.5% SDS) incubated at
37°C for 1 h. After incubation, 1.0% CTAB/NaCl (Cetyl trimethyl ammonium bromide in
0.7M NaCl) was added followed by incubation for 10 min at 65°C. RNA was removed by
treating with RNase (final concentration 100µg/ml) at 37°C for 1h. This was followed by
phenol chloroform extraction and precipitation of the nucleic acid in the presence of
isopropanol (5). Excess salt was removed by 70% alcohol wash and the nucleic acid was air-
dried, resuspended in sterile TE buffer. The purity of the DNA was assayed using a
spectrophotometer (Gene Quant, England) that self calculates the ratio of optical densities at
260 and 280 nm and the DNA was stored at -20°C for subsequent PCR analysis.
PCR analysis.

6
Confirmation of strain identity: The identity of the strains examined in this study was
confirmed by the V. cholerae species-specific ompW PCR (25) and by a multiplex PCR used
to detect O-antigen biosynthesis genes (wbe or wbf) and for the cholera toxin gene ctxA (14).
The PCR reagents and kits were obtained from Invitrogen™. The biotype of all the strains
was further confirmed by genetic traits using PCR assays targeting the tcpA (classical or El
Tor variant) (17) and by the type of rstR gene that regulates the replication and integration of
the CTXΦ in the V. cholerae genome (9). These PCRs were performed using the primers and
procedures described previously (26).
Multilocus virulence gene profiling: We first used a multilocus virulence gene profiling that
scanned for 9 virulence associated genes and/or gene clusters and a house-keeping gene in the
genome of 20 representative Peruvian V. cholerae isolates and a reference strain each of the
classical (O395) and El Tor (N16961) using 31 sets of PCR primers and conditions described
previously (4, 27, 30, 24, 31). PCR was performed in 20 µl reaction mixture as follows: an
initial denaturation step at 96 °C for 1 min followed by 30 cycles of denaturation at 94°C for
30s, primer annealing at 45 to 58°C for 30s, 1 to 4 min of primer extension at 72°C and 7 min
of final extension at 72°C for one cycle. Amplicons were separated by agarose gel
electrophoresis (1%) in 0.5X Tris-Borate-EDTA buffers. The PCR products were analyzed
by electrophoresis in 1% agarose gels, stained with ethidium bromide, visualized under UV
light and recorded by a gel documentation system (Gel Doc™ 2000, BioRad). The PCR
products were sized with standard molecular weight markers and documented.
Analysis of VSP-II region. PCR was used to assay the VSP-II region in the genome of the
83 V. cholerae O1 isolates using previously designed 7 sets of PCR primers following
conditions described previously (27). Further, the GeneFisher software (available at
http://www.genefisher.de/) was utilized to design 5 sets of primers in this study (Table 2) to
amplify ORFs VC0511, VC0512, VC0513, VC0514 and VC0515. The primers of VC0511

7
and VC0512 were designed from the sequence that flanked the region of each of the ORFs.
The other 3 sets of primers for the ORFs VC0513, VC0514 and VC0515 were designed from
within each of the ORFs. PCR was performed in 20 µl reaction mixture as follows: an initial
denaturation step at 95°C for 3 min followed by 30 cycles of denaturation at 95°C for 1 min,
primer annealing at 50ºC to 54°C for 1 min and primer extension at 72°C for 1 min (Table 2).
Nucleotide sequence analysis of VSP-II region.
Several regions of the VSP-II region in PERU-130 strain were amplified by PCR using Ex
Taq (Takara Bio Inc. Shiga, Japan) and several primers newly designed in this study and
described previously (27) but with different combinations as shown in Table 2. PCR products
were purified by QIAquick PCR Purification Kit according to the manufacturer’s instruction
(QIAGEN GmbH, Hilden, Germany) and sequenced by using ABI Prism BigDye Terminator
Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster, CA, USA) and primers
designed on the basis of the sequence of the VSP-II region in V. cholerae O1 El Tor strain
N16961. The PCR product obtained by using LA-Taq (Takara Bio Inc. Shiga, Japan) and the
primer set VSPII LaU and VSPII LaR was digested by Sau3AI. The digests were ligated into
the BamHI site of pBluescript SKII (-) and the ligation mixture was transformed into E. coli
JM109. The recombinant E. coli strains were cultured in L-broth including ampicillin (100
µg/ml) and aliquot was boiled in TE buffer. The boiled template was amplified by M13
forward and reverse primers. The PCR products were purified by QIAquick PCR Purification
Kits (QIAGEN GmbH) and sequenced by using ABI Prism BigDye Terminator Cycle
Sequencing Ready Reaction Kit (Applied Biosystems, Foster, CA, USA) and M13 forward
and reverse primers or sequencing primer designed by the obtained sequence, if necessary.
The reactions were conducted in a GeneAmp 9700 thermal cycler in accordance with the
manufacturer’s instruction (Applied Biosystems). Nucleotide sequence was determined using
an ABI PRISM 3100-Avant Genetic Analyzer (Applied Biosystems, Forster City, CA, USA).

8
Sequence assembly and analysis.
The sequences obtained in this study were analyzed using DNA Lasergene software package
(DNASTAR Inc., Madison, WI, USA). Homology searches were performed using BLAST
program, made available by DNA Data Bank Japan (DDBJ). The sequence of VSP-II in
PERU-130 strain analyzed in this study has been registered in DDBJ (accession number
AB300724).
Nucleotide sequence of ctxB subunit.
To determine the nucleotide sequence of the ctxB subunit of CT, PCR amplification of ctxB
gene of 3 Peruvian strains of V. cholerae O1 El Tor (PERU-044, PERU-130 and PERU-296
isolated in 1995, 1991 and 2003, respectively) was performed in a 25µL reaction mixture in
an automated Peltier thermal cycler (PTC-200, M. J. Research). PCR primers and conditions
were as previously described (21). PCR products were purified with a Microcon centrifugal
filter device (Millipore Corporation, Bedford, Mass.) and sequenced using an ABI PRISM
BigDye Terminator Cycle Sequencing Reaction kit (Perkin-Elmer Applied Biosystems,
Foster City, CA, USA) on ABI PRISM 310 automated sequencer. The chromatogram
sequencing files were inspected using Chromas 2.23 (Technelysium, Queensland, Australia).
Nucleotide sequences of the reference strains were compared with the corresponding
sequences of the N16961 El Tor (GenBank Accession No. NC-002505) and 569B classical
(GenBank Accession No. U25679) strain retrieved from Genebank using Basic Local
Alignment Search Tool. Multiple sequence alignments were developed using CLUSTALX
1.81.13.
Pulsed Field Gel Electrophoresis.
Intact agarose-embedded chromosomal DNA from the isolates of V. cholerae was prepared
and PFGE was performed using a contour-clamped homogeneous electric field (CHEF-

9
Mapper) apparatus (Bio-Rad) following the standardized PulseNet Pulsed-Field Gel
Electrophoresis protocol recently developed for subtyping of V. cholerae (6). Genomic DNA
was digested with NotI enzyme (10 U/µl stock, Invitrogen). The restriction fragments were
separated in 1% SeaKam Gold agarose in 0.5X Tris-borate-EDTA buffer. V. cholerae O1 El
Tor biotype (N16961) and the strain 569B of the classical biotype were used as reference
strains. Salmonella enterica serotype Braenderup strain H9812 digested with 40 Units of
XbaI (Invitrogen) was used as the molecular size marker in lanes 1, 8 and 15. Following
electrophoresis, the gels were stained in ethidium bromide solution (50 µg/ml) for 20 to 30
minutes and destained with reagent grade water. Images were captured using a Gel Doc 2000
and Gel Doc XR systems (Bio-Rad).
Results
The 60 Peruvian and 23 strains from 10 other countries were reconfirmed to be V.
cholerae O1 by phenotypic traits and by serotyping. All the strains were positive for V.
cholerae species specific ompW and for the O1 wbe and belonged to the El Tor biotype (data
not shown). Among the 60 strains of V. cholerae O1 from Peru, two strains (PERU-067 and
PERU-189) were negative for tcpA of both the El Tor and classical type while others were
positive for tcpA of the El Tor type (Table 1). All Peruvian strains including the El Tor
reference strain N16961 were positive for rstR2 (repressor gene of the CTX prophage of the
El Tor biotype) whereas the classical reference strain O395 was positive for rstR1 (repressor
gene of the CTX prophage of the classical biotype). The serotype and genotype of the 60
Peruvian strains and the 23 strains of V. cholerae O1 from other countries are shown in Table
1. Four strains from other countries were negative for ctxA. The strains from Mozambique
were rstR1 positive as reported previously (1) and the strains GP7 from Myanmar and GP156
from Australia were positive for both rstR1 and rstR2. A recent study has shown that all the

10
El Tor strains of V. cholerae O1 isolated from 2001 in Bangladesh produce classical cholera
toxin (23). We therefore examined the nucleotide sequence of the ctxB gene of 3
representative Peruvian El Tor strains of V. cholerae O1 and found that the strains possess
DNA sequences identical to the El Tor type of ctxB which was identical to the ctxB sequence
of El Tor reference strain N16961 and differed from that of the classical reference strain
O395.
Of the 60 Peruvian strains, 20 (listed in Table 3) were selected representing different
year and location of isolation in Peru for the preliminary multilocus virulence gene profiling
that involved examining for 9 virulence associated genes and/or gene clusters and for a
house-keeping gene. For purposes of comparison, we also included a reference strain each of
the classical (O395) and El Tor (N16961) biotypes. The presence or absence of the various
genes was scored by PCR with specific primers using DNA extracted from cultured strains.
All 20 strains from Peru showed the presence of all genes comprising VSP-I, MSHA and
RTX gene clusters (Table 3). The four individual loci namely, hlyA, pilE, tlc and intl4 and the
rstC of RS1 were also present in all the Peruvian strains examined. Among the 9 ORFs of
VSP-II examined in the preliminary genetic screen, two ORFs namely VC0512 and VC0514
were consistently negative by PCR in all the 20 Peruvian strains tested, present in the El Tor
reference strain N16961 but absent in the classical reference strain O395 (classical biotype
strains lack the VSP-I and VSP-II). Apart from this, two Peruvian strains (PERU-067 and
PERU-189) lacked the genes of the VPI-I region examined and one strain PERU-097 was
negative for the ctxAB genes and concluded to be non toxigenic. The overall results of the
preliminary genetic screen indicated that all the Peruvian strains were similar to the reference
El Tor strain of the seventh pandemic with the only striking difference being the negative
result for VC0512 and VC0514 in the VSP-II gene cluster in all of the Peruvian strains.

11
Our next objective was to examine the negative PCR results for ORFs 512 and 514 in
VSP-II in further detail in the 60 Peruvian strains and in the 23 strains of V. cholerae O1
isolated from 10 different countries (Table 4). We examined all the 60 Peruvian strains for 12
ORFs of VSP-II and found that 51 out of the 60 Peruvian strains were negative for VC0511
and of these 20 were positive for VC0516 and 31 were negative for VC0516. The remaining
9 strains were positive for VC0511 but negative for VC0516. The other ORFs of VSP-II
region, that is, VC0490, VC0493, VC0498, VC0502, VC0504 and VC0517 were present in
all the Peruvian strains and only strains PERU-044 was negative for VC0490 and PERU-001
was negative for VC0493. VC0512, VC0513, VC0514 and VC0515 were negative by PCR in
all the 60 Peruvian strains but present in all other strains examined from different countries
except in three strains from Australia. The complete VSP-II region was absent in the
Australian strains, which was similar to the classical reference strain of V. cholerae (O395)
used in this study.
To understand the differences between the VSP-II of the Peruvian and the prototype
seventh pandemic V. cholerae O1 El Tor strain, PCR was performed to amplify the region
corresponding to the VSP-II of the Peruvian strain and to sequence. When primer sets, 498F
and 490R, 490F and 493R, 493F and 498R, 502F and 504R, 47U and 517R, were used,
expected sizes of the PCR products were obtained as shown in Fig. 1. When primer sets,
504F and 512R, 512F and 516R and 514F and 517R, were used, however, no amplicon were
obtained. Therefore, we newly designed PCR primer set, VSPII LaU and VSPII LaR on the
basis of the sequence obtained from PCR products amplified by 502F and 504R, and by 47U
and 517R, to amplify the regions where no amplification was observed by previously
reported PCR primer sets and we sequenced 31,299 bp comprising the entire VSP-II and
flanking regions of PERU-130 which was negative by PCR for VC0511 and for VC0513 to
VC0516. The comparison of the nucleotide sequences and other parameters of VSP-II and

12
flanking regions of PERU-130 with that of VSP-II of N16961 are shown in Figure 1 and
Table 5. The 26.5 kb (26,519 bp) VSP-II of PERU-130 contained 24 ORFs. The N16961 26.9
kb (26,866 bp) also contained 24 ORFs (28). In the VSP-II DNA shared between PERU-130
and N16961, four regions could be identified based on extent of similarity. These were ORFs
2 to 14 of PERU-130 (100% similarity), ORFs 15 to 19 (94.4% similarity), ORFs 20 to 24
(46.8% similarity) and ORF 25 (89% similarity) (Figure 1). The sequencing data confirmed
our PCR results indicating that ORFs 20 to 24 of the Peruvian strain were, indeed, different
from VC0511 to VC0515 of NI6961.
Each of the ORFs of PERU-130 was analyzed using the BLAST program (2). The
hypothetical proteins encoded by ORFs 20 and 21 showed 46.7% and 33.2% homology to
proteins of Azoarcus sp. BH72, a mutualistic N2-fixing endophyte of rice and other grasses
(19). ORF 22 representing another hypothetical protein that showed 46.7% homology to a
protein encoded by a gene designated as Mmcs_4842 of Mycobacterium sp. MCS (Table 5).
The transposase orfAB, subunit A represented by ORF 23 of PERU-130 showed 87.2%
homology to similar protein encoded by VCA0372 of NI6961 while transposase orfAB
subunit B representing ORF24 showed 83.6% and 86.1% homology to similar proteins of
Nitrococcus mobilis Nb-231 and VCA0371 of V. cholerae N16961, respectively (Table 5).
However, ORF23 and 24 DNA (1214 bp) have high homology to V. cholerae serogroup
O103 insertion sequences ISalg (DNA identity = 98.8%) reported earlier (33). ORF25 of
PERU-130 shared close homology to the phage integrase of V. vulnificus YJ016 (98.5%) and
V. cholerae (93.7%) strain N16961. The 5’ flanking region (ORF1) and the 3’ flanking region
(ORF26) of PERU-130 were identical to VC0489 and VC0517 of strain N16961.
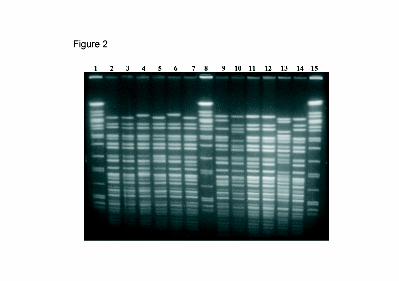
PFGE analysis of 10 representative Peruvian strains isolated between 1991 and 1996
showed that the strains were not clonal and there were 8 different PFGE profiles (Figure 2)
indicating the genetic diversity among these strains.

13
Discussion
The VSP-I and VSP-II are unique to the seventh pandemic El Tor biotype strains of V.
cholerae O1 and absent in strains of the classical biotype. The role of VSP-I and VSP-II in
the El Tor biotype is not known (27) but their exclusive presence among the El Tor strains is
responsible for some of the distinctive properties of this biotype (10) and may contribute in
some way to its fitness and survival (27). The VSP-II is a 29.6 kb genomic island
encompassing 24 predicted ORFs (VC0490 to VC0516) whose functions include DNA repair
and methyl-accepting chemotaxis proteins, a group of hypothetical proteins and a
bacteriophage-like integrase adjacent to a tRNA gene (28). By utilizing a multilocus
virulence gene profiling approach, we discovered that the Peruvian El Tor strains of V.
cholerae O1 have an intact VSP-I but by PCR all strains were consistently negative for
adjacent ORFs of the VSP-II namely VC0512 (methyl-accepting chemotaxis protein),
VC0513 (transcriptional regulator), VC0514 (methyl-accepting chemotaxis protein) and
VC0515 (hypothetical protein). At the time of its discovery, the VSP-II was described as a
7.5 kb region encompassing 8 ORFs (VCO490 to VCO497). Later, O’Shea et. al. (28)
demonstrated that the 7.5 kb VSP-II region is part of a 29.6 kb island that encompasses ORFs
VC0490-VC0516. In the initial study of Dizejman et al. (10), a Peruvian strain C6709 was
included but the absence of ORFs although identified went unnoticed since at that time these
ORFs were not included as a part of the VSP-II region.
The Latin American isolates, at the time of its introduction, were clonal and were
described as ET4 by multi locus enzyme electrophoresis (MEE); the seventh pandemic strains
in Asia and Africa belonged to ET3 (36). Later studies showed at least two distinct clones
easily distinguishable by multilocus enzyme electrophoresis (MEE), ribotype, PFGE pattern
and antimicrobial resistance pattern (11). By ribotyping, the Latin American epidemic was
caused by strains of ribotype 5, which were isolated from several other geographical locations

14
but could be differentiated from the Latin American strains by other molecular typing
techniques (29). The evolution of V. cholerae O1 biotype El Tor strains isolated in Lima,
Peru from 1991 to 1995 showed the continuous, and more frequent occurrence than
previously, of genetic changes in the Latin American cholera epidemic strains (8). The
Peruvian strains that we examined in this study were isolated from the beginning of the
epidemic in 1991 through 2003. The PFGE analysis of the Peruvian strains from 1991 to
1996 in this study showed a variety of PFGE profiles indicating genetic diversity among
these strains. Despite the genetic diversity, changing serotypes and different years and
locations of isolation, the VSP-II region of the Peruvian strains isolated for more than a
decade was stably conserved and different from the seventh pandemic strains isolated in other
continents.
The complete nucleotide sequence analysis of VSP-II showed little difference in the
overall size of the VSP-II of PERU-130 and NI6961 and the flanking regions were
conserved. Of the 25 ORFs in the VSP-II of PERU-130, 20 showed extensive homology with
the corresponding ORFs of N16961. Only one region comprising ORFs 20 to 24 of PERU-
130 was different from the corresponding VSP-II region constituting VC0511 to VC0515 of
the prototype El Tor strain N16961. The presence of hypothetical proteins homologous to
those present in the nitrogen-fixing endophyte symbiotic Azoarcus sp. in the VSP-II region
(ORF 20 and 21) indicates that the progenitor of the Peruvian V. cholerae strains may have
acquired these genes from microorganisms in the environment. The extensive DNA
homology of ORFs 23 and 24 of the VSP-II of PERU-130 to V. cholerae serogroup O103
insertion sequence ISalg is interesting since ISalg is distributed in 69% of the V. cholerae
non-O1 non-O139 strains and has extensive homology to the IS element of Vibrio
alginolyticus and IS911 of Shigella dysenteriae (12, 33). We are not sure if these five

15
divergent ORFs of PERU-130 assembled together or were independent acquisitions but the
presence of ISalg suggests genetic rearrangements or horizontal transfer.
The origin and the source of the Latin American strains of V. cholerae O1 in Peru
remains a mystery. The ‘single introduction hypothesis’ states that the Latin American strains
may have originated as a result of emptying of contaminated bilge waters of ships arriving
from cholera endemic areas of Asia or Africa (20). The difference in the VSP-II between the
Peruvian strains and those from Asia and Africa does not support this hypothesis. Another,
less accepted hypothesis, consistent with simultaneous appearance of the disease in a number
of coastal Peruvian cites within a few days, relates to the sudden and massive expansion of a
preexisting small population of pathogenic cholera vibrios on zoo- or phytoplankton in
Peruvian coastal waters (7). Once introduced into the coastal communities in concentrations
large enough for human infection to occur, cholera spread by the well-known means of
contaminated water and food (32). Our studies indicate that the Peruvian strains may be
genetic derivatives of V. cholerae O1 from the coastal environs of Peru because the VSP-II of
1991 environmental Peruvian strains examined in this study were similar to all the clinical
Peruvian strains and different from strains isolated from other countries and continents. Seas
et. al. (32) also postulate that cholera vibrios, autochthonous to the aquatic environment, were
present in multiple coastal locations, and resulted from environmental conditions that existed
during an El Niño phenomenon.
From a functional viewpoint, we do not know the significance of the difference
between VSP-II of the Peruvian strains and those isolated from other continents. However,
from an identity point of view, this difference in the VSP-II lends a unique molecular
signature to the Peruvian strains that could form the basis of tracking the origin of the
Peruvian strains and therefore the Latin American pandemic. Based on the stable difference
in VSP-II, we conclude that the Peruvian El Tor O1 strains are quite distinct from the seventh

16
pandemic Eurasian and African strains, suggesting that, although temporally associated with
the seventh pandemic, it arose independently probably from a nonhuman environmental
source. Optimally, we should have examined strains from other Latin American countries but
did not do so since we had none in our collection. Further studies are required to understand
what the difference means functionally.

17
Acknowledgements
This study was supported by grants provided to AG (IIN, Lima, Peru) and GBN
(ICDDR,B, Dhaka, Bangladesh) from The Academy of Sciences for the Developing World
(formerly the Third World Academy of Sciences) and by International Cooperation Research
Grant to SY (OPU, Osaka, Japan) and GBN (ICDDR,B, Dhaka, Bangladesh) from the
Ministry of Health, Labor, and Welfare, Japan. We also wish to thank Dr. Cesar Munayco
and Biol. Tania H Alarcon for graciously providing the environmental strains of V. cholerae
from Peru

18
References
1. Ansaruzzaman, M., N.A. Bhuiyan, G.B. Nair, D.A. Sack, M. Lucas, J.L. Deen, J.
Ampuero, C.L.Chaignat, and Mozambique Cholera Vaccine Demonstration
Project Coordination Group. 2004. Cholera in Mozambique, variant of Vibrio
cholerae. Emerg. Infect. Dis. 10:2057-2059.
2. Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, and
D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein
database search programs. Nucleic Acids Res. 25: 3389–3402.
3. Chen, C.Y., K.M. Wu, Y.C. Chang, C.H. Chang, H.C. Tsai, T.L. Liao, Y.M. Liu,
H.J. Chen, A.B. Shen, J.C. Li, T.L. Su, C.P. Shao, C.T. Lee, L.I. Hor, and S.F.
Tsai. 2003. Comparative genome analysis of Vibrio vulnificus, a marine pathogen.
Genome Res. 13:2577-87.
4. Chow, K. H., T. K. Ng, K. Y. Yuen, and W. C. Yam. 2001. Detection of RTX toxin
gene in Vibrio cholerae by PCR. J. Clin. Microbiol. 39: 2594–2597.
5. Chowdhury, N. R., S. Chakraborty, T. Ramamurthy, M. Nishibuchi, S.
Yamasaki, Y. Takeda, and G. B. Nair. 2000. Molecular evidence of clonal Vibrio
parahaemolyticus pandemic strains. Emerg. Infect. Dis. 6: 631-636.
6. Cooper, K.L., C.K. Luey, M. Bird, J. Terajima, G.B. Nair, K.M. Kam, E.
Arakawa, A. Safa, D.T. Cheung, C.P. Law, H. Watanabe, K. Kubota, B.
Swaminathan, and E.M. Ribot. 2006. Development and validation of a PulseNet
standardized pulsed-field gel electrophoresis protocol for subtyping of Vibrio
cholerae. Foodborne Pathog. Dis.3: 51-58
7. Craig, J. P. 1996. Cholera: outlook for the twenty-first century. Caduceus. 12: 25-42.

19
8. Dalsgaard, A., M.N. Skov, O. Serichantalergs, P. Echeverria, R. Meza, D.N.
Taylor. 1997. Molecular evolution of Vibrio cholerae O1 strains isolated in Lima,
Peru, from 1991 to 1995. J. Clin. Microbiol. 35:1151– 1156.
9. Davis, B. M., H. H. Kimsey, W. Chang, and M. K. Waldor.1999. The Vibrio
cholerae O139 Calcutta bacteriophage CTXφ is infectious and encodes a novel
repressor. J. Bacteriol. 181: 6779–6787.
10. Dziejman, M., E. Balon, D. Boyd, C. M. Fraser, J. F. Heidelberg, and J. J.
Mekalanos. 2002. Comparative genomic analysis of Vibrio cholerae: genes that
correlate with cholera endemic and pandemic disease. Proc. Natl. Acad. Sci. U S A.
99: 1556–1561.
11. Evins, G.M., D.N. Cameron, J.G. Wells, K.D. Greene, T. Popovic, S. Giono-
Cerezo, I.K. Wachsmuth, and R.V. Tauxe. 1995. The emerging diversity of the
electrophoretic types of Vibrio cholerae in the Western hemisphere. J. Infect. Dis.
172:173-179.
12. Hayashi, M., J. Fujii, and T. Unemoto. 1997. Genetic defect of the sodium pump-
defective mutant nap-1 from the marine Vibrio alginolyticus. Biochem. Mol. Biol.
Intl. 41:41-47.
13. Heidelberg J. F., J. A. Eisen, W. C. Nelson, and 23 other authors. 2000. DNA
sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 406:
477–483.
14. Hoshino, K., S. Yamasaki, A. K. Mukhopadhyay, S. Chakraborty, A. Basu, S. K.
Bhattacharya, G. B. Nair, T. Shimada, and Y. Takeda. 1998. Development and
evaluation of a multiplex PCR assay for rapid detection of toxigenic Vibrio cholerae
O1 and O139. FEMS Immunol. Med. Microbiol .20: 201–207.

20
15. Kaper, J. B., J. G. Morris, M. M. Jr, and Levine. 1995. Cholera. Clin. Microbiol.
Rev. 8: 48–86.
16. Karaolis, D.K., J.A. Johnson, C.C. Bailey, E.C. Boedeker, J.B. Kaper, and
P.R.Reeves. 1998. A Vibrio cholerae pathogenicity island associated with epidemic
and pandemic strains. Proc. Natl. Acad. Sci. U S A. 95:3134-3139.
17. Keasler, S. P., and R. H. Hall. 1993. Detecting and biotyping V. cholerae O1 with
multiplex polymerase chain reaction. Lancet. 341: 1661.
18. Kovach, M.E., M.D. Shaffer, and K.M. Peterson. 1996. A putative integrase gene
defines the distal end of a large cluster of ToxR-regulated colonization genes in
Vibrio cholerae. Microbiology. 142:2165-2174.
19. Krause, A., A. Ramakumar, D. Bartels, and 19 other authors. 2006. Complete
genome of the mutualistic, N2-fixing grass endophyte Azoarcus sp. Strains BH72.
Nature Biotech. 24:1385-1391.
20. McCarthy, S. A., and F. M. Khambaty. 1994. International dissemination of
epidemic Vibrio cholerae by cargo ship ballast and other nonpotable waters. Appl.
Env. Microbiol. 60: 2597-2601.
21. Mitra, R. K., R. K. Nandy, T. Ramamurthy, S. K. Bhattacharya, S. Yamasaki,T.
Shimada, S. Toshio, Y. Takeda, and G. B. Nair. 2001. Molecular characterization
of rough variants of Vibrio cholerae isolated from hospitalised patients with
diarrhoea. J. Med. Microbiol. 50:268–276.
22. Nair, G. B., S. M. Faruque, N. A. Bhuiyan, M. Kamruzzaman, A. K. Siddique,
and D. A. Sack. 2002. New variants of Vibrio cholerae O1 biotype El Tor with
attributes of the classical biotype from hospitalized patients with acute diarrhea in
Bangladesh. J. Clin. Microbiol. 40:3296–3299.

21
23. Nair, G.B., F. Qadri, J. Holmgren, A.-M. Svennerholm, A. Safa, N.A. Bhuiyan,
Q.Shafi Ahmed, S.M. Faruque, Y. Takeda and D.A. Sack. 2006. Cholera due to
altered El Tor strains of Vibrio cholerae O1 in Bangladesh. J. Clin. Microbiol.
44:4211-4213.
24. Nair, G.B., A. Safa, N.A. Bhuiyan, S. Nusrin, D. Murphy, C. Nicol, M. Valcanis,
S. Iddings, I. Kubuabola, and H. Vally. 2006. Isolation of Vibrio cholerae O1
strains similar to pre-seventh pandemic El Tor strains during an outbreak of
gastrointestinal disease in an island resort in Fiji. J. Med. Microbiol. 55:1559-1562.
25. Nandi, S., D. Maiti, A. Shaha, and R. K. Bhadra. 2003. Genesis of variants of V.
cholerae O1 biotype El Tor: role of the CTXϕ array and its position in the genome.
Microbiology. 149: 89–97.
26. Nusrin, S., G. Y. Khan, N. A. Bhuiyan, and 9 other authors. 2004. Diverse ctx
phages among toxigenic Vibrio cholerae O1 and O139 strains isolated between 1994
and 2002 in an area where cholera is endemic in Bangladesh. J. Clin. Microbiol. 42:
5854–5856.
27. O’Shea, Y. A., F. J. Reen, A. M. Quirke, and E. F. Boyd. 2004a. Evolutionary
genetic analysis of the emergence of epidemic Vibrio cholerae isolates based on
comparative nucleotide sequence analysis and multilocus virulence gene profiles. J.
Clin. Microbiol. 42:4657–4671.
28. O’Shea, Y.A., S. Finnan, F. J. Reen, J. P. Morrissey, F. O’Gara, and E.F. Boyd.
2004b. The Vibrio seventh pandemic island-II is a 26.9 kb genomic island present in
Vibrio cholerae El Tor and O139 serogroup isolates that shows homology to a 43.4 kb
genomic island in V. vulnificus. Microbiology. 150: 4053–4063.

22
29. Popovic, T., C. Bopp, O. Olsvik, and I.K. Wachsmuth. 1993. Epidemiologic
application of a standardized ribotype scheme for Vibrio cholerae O1. J. Clin.
Microbiol. 31:2474-2482.
30. Rivera, I. N., J. Chun, A. Huq, R. B. Sack, and R. R. Colwell. 2001. Genotypes
associated with virulence in environmental isolates of Vibrio cholerae. Appl. Env.
Microbiol. 67: 2421–2429.
31. Safa, A., N.A. Bhuiyan, S. Nusrin, M. Ansaruzzaman, M. Alam, T. Hamabata, Y.
Takeda, D.A. Sack, and G.B. Nair. 2006. Genetic characteristics of Matlab variants
of Vibrio cholerae O1 that are hybrids between classical and El Tor biotypes. J. Med.
Microbiol. 55:1563-1569.
32. Seas, C., J. Miranda, A.I. Gil, R. Leon-Barua, J. Patz, A. Huq, R.R. Colwell, and
R.B. Sack. 2000. New insights on the emergence of cholera in Latin America during
1991: The Peruvian experience. Am. J. Trop. Med. Hyg. 62:513-517.
33. Sozhamannan, S., Y.K. Deng, M. Li, A. Sulakvedidze, J.B. Kaper, J. A. Johnson,
G.B. Nair, and J.G. Morris, Jr. 1999. Cloning and sequencing of the genes
downstream of the wbf gene cluster of Vibrio cholerae serogroup O139 and analysis
of the junction genes in other serogroups. Infect. Immun. 67:5033-5040.
34. Tauxe R., E. Mintz, and R. Quick. 1995. Epidemic cholera in the New World:
translating field epidemiology into new prevention strategies. Emerg. Infect. Dis. 1:
141-146.
35. Trucksis, M., J. Michalski, Y. K. Deng, and J. B. Kaper. 1998. The Vibrio
cholerae genome contains two unique circular chromosomes. Proc. Natl. Acad. Sci. U
S A. 95: 14464–14469.

23
36. Wachsmuth, I.K., G.M. Evins, P.I. Fields, Ǿ. Olsvik, T. Popovic, C.A. Bopp, J.G.
Wells, C. Carrillo, and P.A. Blake .1993. The molecular epidemiology of cholera in
Latin America. J. Infect. Dis. 167:621– 626.
37. Waldor, M.K. and J.J. Mekalanos. 1996. Lysogenic conversion by a filamentous
phage encoding cholera toxin. Science. 272: 1910-1914.

24
Legend to Figures
Figure 1. Schematic representation of the organization of VSP-II from V. cholerae strains
N16961 (a) and PERU-130 (b). The position and direction of transcription of the ORFs are
indicated by the direction of the arrows. Homology between N16961 and PERU-130 at the
DNA level is described as percentage and is shown between the ‘a’ and ‘b’ panels. The
numbers refer to the genetic organization of the genes. The regions amplified by PCR using
each primer set, described in Table 2, are depicted (c). The solid bar indicates the region
amplified, however, dashed bar indicates the region fail to be amplified. The number below
the arrow indicates the name of the PCR primer used.
Figure 2. NotI-restricted patterns of chromosomal DNA of Peruvian V. cholerae O1 El Tor
strains isolated between 1991 and 1996. Lanes: 1, 8, and 15 Salmonella braenderup strain
H9812 as molecular mass marker; lanes 2, 3,and 5 represent pulsotype A1, PERU-180
(1991), PERU-139 (1992) and PERU-067 (1993) respectively; lane 4 pulsotype B1, PERU-
011 (1993); lane 6 pulsotype B2, PERU-086 (1994); lane 7 pulsotype A2, PERU-115 (1994);
lane 9 pulsotype C, PERU-097 (1995); lane 10 pulsotype D, PERU-132 (1995); lane 11
pulsotype A3, PERU-120 (1996); lane 12 pulsotype A4, PERU-137 (1996); and lanes 13, 14
represent 569B classical and N16961 El Tor strains respectively.

Table 1. Serotype and genotype of the V. cholerae O1 El Tor biotype strains and
the two reference strains examined in this study
Genotype Country
(number of strains)
Year of
Isolation
Serotype
ctxA tcpA rstRa
Peru (47) 1991-1999, 2003 Ogawa + + 2
Peru (10) 1991-1994 Inaba + + 2
Peru (1) 1991 Inaba + - 2
Peru (1) 1995 Ogawa - + 2
Peru (1) 1993 Ogawa + - 2
Australia (1) 1986 Inaba + + 1
Australia (1) 1965 Inaba - - -
Australia (1) 1979 Inaba + + 1,2
Bangladesh (4) 2004 Ogawa + + 2
India (2) 1975, 1982 Ogawa + + 2
India (1) 1981 Ogawa - + 2
Maldives (1) 1978 Ogawa + + 2
Maldives (1) 1978 Ogawa - + 2
Malaysia (1) 1978 Inaba + + 2
Malaysia (1) 1978 Ogawa + + 2
Myanmar (1) 1970 Ogawa + + 1,2
Macao, China (1) 1970 Inaba - + 2
Germany (1) 1975 Inaba + + 2
Germany (1) 1975 Ogawa + + 2
Zambia (2) 1996, 2003 Ogawa + + 2
Mozambique (3) 2004 Ogawa + + 1
O395 (classical) 1965 Ogawa + + 1
N16961 (El Tor) 1971 Inaba + + 2
a 1, classical biotype rstR; 2 El Tor biotype rstR

Table 2. PCR primer sequences and conditions for analyzing the VSP-II region of V. cholerae O1 El Tor strains isolated in Peru and various other
countries
Primer Sequences (5'-3') PCR condition No. of Product size Reference
designation Denaturation Annealing Extention cycle (bp)
VC0511F1 CTTGCTGCGTACTTAGCA 95ºC, 1 min 54ºC, 1 min 72ºC, 1 min 30 385 This study
VC0511R1 AGTAGCATCGCTCTCGTA
VC0512F1 TCCTGATTGGGAGCGAA 95ºC, 1 min 54ºC, 1 min 72ºC, 1 min 30 1.704 This study
VC0512R1 TCACGCGGGTTATTCCA
VC0513F1 CTGAGGTGTTATATGTTTCG 95ºC, 1 min 54ºC, 1 min 72ºC, 1 min 30 781 This study
VC0513R1 TCAAATTTCCTGACAGTTCC
VC0514F1 GTTTGGGAAGGGTACACA 95ºC, 1 min 52ºC, 1 min 72ºC, 1 min 30 1.651 This study
VC0514R1 GCTCTTCAGCCGCTGA
VC0515F1 GGTGGTGCTGCATGGA 95ºC, 1 min 50ºC, 1 min 72ºC, 1 min 30 1.135 This study
VC0515R1 TCTAAAGCCTCACACCA
VC0489F AGATCAACTACGATCAAGCC 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 5.202 27
VC0490R TGCAGTTGTTGAATGGAC
VC0490F CGTGAAGGGATATAGGAG 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 6.718 27
VC0493R CGCTCTTCTTTCCACGCTTCA
VC0493F AATGCTTCTCAGGGGGGTCTT 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 7.439 27
VC0498R TGCGGCTCCAATGGAGTCTG
VC0502F TCATCAGTTAGCACACGAAC 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 5.808 27
VC0504R AGCCCGAAATGAATCCCAAAA
VC0504F CAGCAAAGGCGGAAGAGGTAG 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 7.616 27
VC0512R CCCTCCACTGCTATTCCG
VC0512F CAGTGGCTTCGCAGAGGA 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 8.384 27
VC0516R TCCTGATGTCTCTCTTGCCG
VC0514F TTATGATCCAAGGAGTAGGG 94ºC, 30 sec 52ºC, 30 sec 72ºC, 4 min 30 6.736 27

Table 3. Results of the genetic screen used for identifying 11 virulence regions and 1 house-keeping gene of Vibrio cholerae O1 strains isolated in Peru between 1991 and 2003
Ho
use
kee
pin
g
gen
e
VSP-I MSHA hlyA VSP-II VPI-I pilE RTX RS1 CTX
Strain
Bio
typ
e
md
h
175
178
180
183
185
398
400
403
406
489
490
493
498
502
504
512
514
516
517
tcp
A
tox
T
acf
B
pil
E
rtxA
rtxC
rstC
rstA
orf
U
zot
ctx
AB
tlc
intl
4
PERU-130 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-139 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-067 El Tor + + + + + + + + + + + + + + + + - - + + - - - + + + + + + + + + +
PERU-115 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-044 El Tor + + + + + + + + + + + - + + + + - - + + E + + + + + + + + + + + +
PERU-097 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + - + +
PERU-120 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-169 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-037 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-299 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-189 El Tor + + + + + + + + + + + + + + + + - - + + - - - + + + + + + + + + +
PERU-200 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-205 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-219 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-306 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-324 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-327 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-328 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-329 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
PERU-330 El Tor + + + + + + + + + + + + + + + + - - + + E + + + + + + + + + + + +
N16961 El Tor + + + + + + + + + + + + + + + + + + + + E + + + + + + + + + + + +
O395 Classical + - - - - - + + + + + - - - - - - - - - C + + + + - - + + + + + +
E for El tor tcpA and C for classical tcpA

TABLE 4. Comparison of VSP-II region of the Peruvian V. cholerae O1 El Tor isolates with
the isolates from different countries possessing different ORFs of VSP-II region
VSP-II
Pla
ce o
f
isola
tion
No. of
str
ain
s
VC
O4
90
VC
O4
93
VC
O4
98
VC
O5
02
VC
O5
04
VC
O5
11
VC
O5
12
VC
O5
13
VC
O5
14
VC
O5
15
VC
O5
16
VC
O5
17
Peru 30 + + + + + - - - - - - +
Peru 19 + + + + + - - - - - + +
Peru 9 + + + + + + - - - - - +
Peru 1 - + + + + - - - - - + +
Peru 1 + - + + + - - - - - - +
Australia 3 - - - - - - - - - - - +
Bangladesh 4 + - - + + + + + + + + +
Myanmar 1 + + + + + + + + + + + +
India 3 + + + + + + + + + + + +
Maldives 2 + + + + + + + + + + + +
Malaysia 2 + + + + + + + + + + + +
Mozambique 3 + + + + + + + + + + + +
Macao, China 1 + + + + + + + + + + + +
Germany 2 + + + + + + + + + + + +
Zambia 1 + + + + + + + + + + + +
Zambia 1 + - - + + + + + + + + +
Classical (O395) 1 - - - - - - - - - - - +
El Tor (N16961) 1 + + + + + + + + + + + +
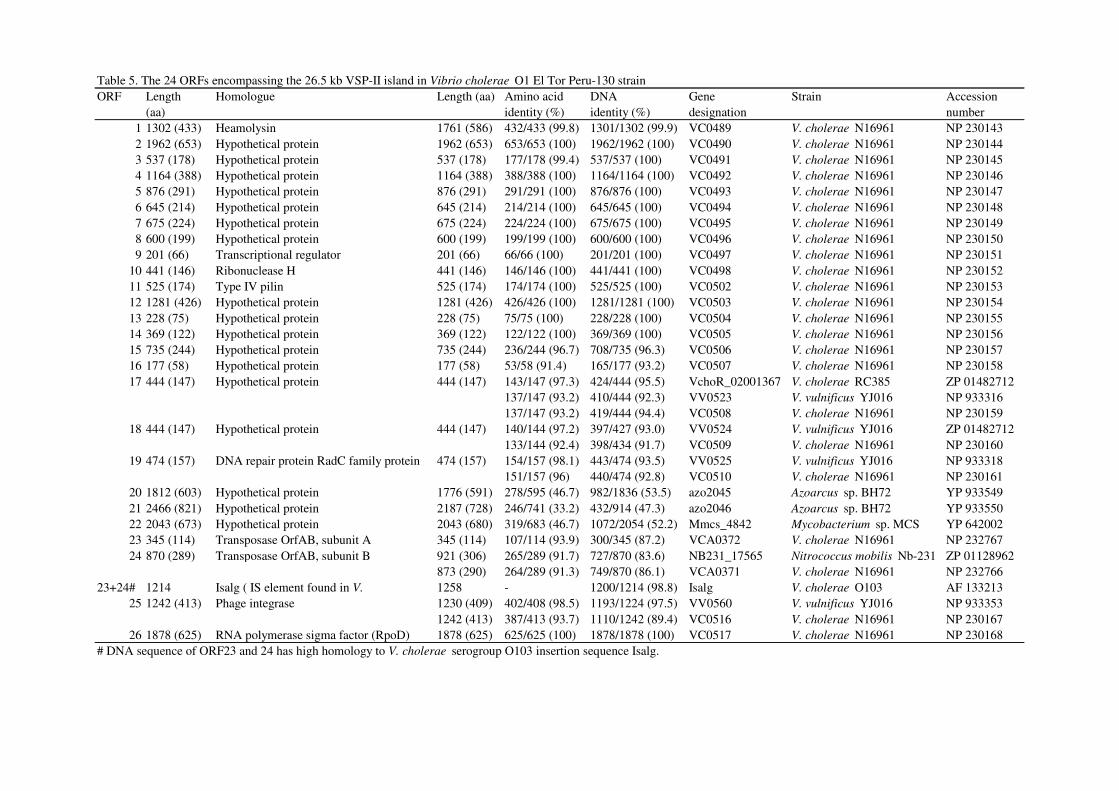
Table 5. The 24 ORFs encompassing the 26.5 kb VSP-II island in Vibrio cholerae O1 El Tor Peru-130 strain
ORF Length Homologue Length (aa) Amino acid DNA Gene Strain Accession
(aa) identity (%) identity (%) designation number
1 1302 (433) Heamolysin 1761 (586) 432/433 (99.8) 1301/1302 (99.9) VC0489 V. cholerae N16961 NP 230143
2 1962 (653) Hypothetical protein 1962 (653) 653/653 (100) 1962/1962 (100) VC0490 V. cholerae N16961 NP 230144
3 537 (178) Hypothetical protein 537 (178) 177/178 (99.4) 537/537 (100) VC0491 V. cholerae N16961 NP 230145
4 1164 (388) Hypothetical protein 1164 (388) 388/388 (100) 1164/1164 (100) VC0492 V. cholerae N16961 NP 230146
5 876 (291) Hypothetical protein 876 (291) 291/291 (100) 876/876 (100) VC0493 V. cholerae N16961 NP 230147
6 645 (214) Hypothetical protein 645 (214) 214/214 (100) 645/645 (100) VC0494 V. cholerae N16961 NP 230148
7 675 (224) Hypothetical protein 675 (224) 224/224 (100) 675/675 (100) VC0495 V. cholerae N16961 NP 230149
8 600 (199) Hypothetical protein 600 (199) 199/199 (100) 600/600 (100) VC0496 V. cholerae N16961 NP 230150
9 201 (66) Transcriptional regulator 201 (66) 66/66 (100) 201/201 (100) VC0497 V. cholerae N16961 NP 230151
10 441 (146) Ribonuclease H 441 (146) 146/146 (100) 441/441 (100) VC0498 V. cholerae N16961 NP 230152
11 525 (174) Type IV pilin 525 (174) 174/174 (100) 525/525 (100) VC0502 V. cholerae N16961 NP 230153
12 1281 (426) Hypothetical protein 1281 (426) 426/426 (100) 1281/1281 (100) VC0503 V. cholerae N16961 NP 230154
13 228 (75) Hypothetical protein 228 (75) 75/75 (100) 228/228 (100) VC0504 V. cholerae N16961 NP 230155
14 369 (122) Hypothetical protein 369 (122) 122/122 (100) 369/369 (100) VC0505 V. cholerae N16961 NP 230156
15 735 (244) Hypothetical protein 735 (244) 236/244 (96.7) 708/735 (96.3) VC0506 V. cholerae N16961 NP 230157
16 177 (58) Hypothetical protein 177 (58) 53/58 (91.4) 165/177 (93.2) VC0507 V. cholerae N16961 NP 230158
17 444 (147) Hypothetical protein 444 (147) 143/147 (97.3) 424/444 (95.5) VchoR_02001367 V. cholerae RC385 ZP 01482712
137/147 (93.2) 410/444 (92.3) VV0523 V. vulnificus YJ016 NP 933316
137/147 (93.2) 419/444 (94.4) VC0508 V. cholerae N16961 NP 230159
18 444 (147) Hypothetical protein 444 (147) 140/144 (97.2) 397/427 (93.0) VV0524 V. vulnificus YJ016 ZP 01482712
133/144 (92.4) 398/434 (91.7) VC0509 V. cholerae N16961 NP 230160
19 474 (157) DNA repair protein RadC family protein 474 (157) 154/157 (98.1) 443/474 (93.5) VV0525 V. vulnificus YJ016 NP 933318
151/157 (96) 440/474 (92.8) VC0510 V. cholerae N16961 NP 230161
20 1812 (603) Hypothetical protein 1776 (591) 278/595 (46.7) 982/1836 (53.5) azo2045 Azoarcus sp. BH72 YP 933549
21 2466 (821) Hypothetical protein 2187 (728) 246/741 (33.2) 432/914 (47.3) azo2046 Azoarcus sp. BH72 YP 933550
22 2043 (673) Hypothetical protein 2043 (680) 319/683 (46.7) 1072/2054 (52.2) Mmcs_4842 Mycobacterium sp. MCS YP 642002
23 345 (114) Transposase OrfAB, subunit A 345 (114) 107/114 (93.9) 300/345 (87.2) VCA0372 V. cholerae N16961 NP 232767
24 870 (289) Transposase OrfAB, subunit B 921 (306) 265/289 (91.7) 727/870 (83.6) NB231_17565 Nitrococcus mobilis Nb-231 ZP 01128962
873 (290) 264/289 (91.3) 749/870 (86.1) VCA0371 V. cholerae N16961 NP 232766
23+24# 1214 Isalg ( IS element found in V. 1258 - 1200/1214 (98.8) Isalg V. cholerae O103 AF 133213
25 1242 (413) Phage integrase 1230 (409) 402/408 (98.5) 1193/1224 (97.5) VV0560 V. vulnificus YJ016 NP 933353
1242 (413) 387/413 (93.7) 1110/1242 (89.4) VC0516 V. cholerae N16961 NP 230167
26 1878 (625) RNA polymerase sigma factor (RpoD) 1878 (625) 625/625 (100) 1878/1878 (100) VC0517 V. cholerae N16961 NP 230168
# DNA sequence of ORF23 and 24 has high homology to V. cholerae serogroup O103 insertion sequence Isalg.