the pharmacology of batrachotoxin. iii. effect on … pharmacology of... · 2009-08-08 · -40...
TRANSCRIPT

529
THE JOURNAL OF PHARMACOLOCY AND EXPERIMENTAL THERAPEUTICS
Copyright 0 i97i by The Williams & Wilkins Co.
Vol. 176, No. 3Piintedin U.S.A.
THE PHARMACOLOGY OF BATRACHOTOXIN. III. EFFECT ONTHE HEART PURKINJE FIBERS’
P. M. HOGAN’ AND E. X. ALBUQUERQUE
Deportment of Pharmacology, Schools of Medicine and Dentistry, State lnzversity
of New York at Buffalo, Buffalo, New York
Accepted for publication November 12, 1970
ABSTRACT
HOGAN. P. M. AND E. X. ALBUQUERQUE: The pharmacology of batrachiotoxin. III. Effecton the heart Purkinje fibers. J. Pharmacol. Exp. Ther. 176: 529-537, 1971. The effects
of batrachotoxin (BTX) on the electrogenic properties of Purkinjc fibers isolated fromthe dog heart were studied with intracellular techniques. BTX caused a depolarization of
the Purkinje fiber membrane and in a few cases the membrane potential reached valuesof +20 mV. Application of anodal hyperpolarizing current, 1 aiM Na-sucrose solution or
tetrodotoxin to the Purkinje fiber previously depolarized by BTX, restored the mcmii-
brane potential to values similar to control conditions. High extracellu!ar calcium con-
centration prolonged the time course of the BTX effect but did not prevent it. Prior tothe onset of membrane depolarization, BTX caused an increase in the duration of the
plateau and negative afterpotential of the action potential. Simultaneous with the de-polarization induced by BTX, a decremise in the upstroke velocity and overshoot of the
spike potential occurred together with an increase in the rate of (lepolarization duringthe diastolic interval. When the membrane was depolarized to values less than -20 mV.excitability was lost. These effects of BTX on heart Purkinje fibers were antagonized bytetrodotoxin. The results raise the possibility of the existence of functionally different
sodium channels. It is concluded that BTX acts on the heart Purkinje fiber to selectively
increase sodium permeability.
Batrachotoxin (BTX) has been reported to
cause a large variety of ventricular arrhythmias
in addition to a complete block of atrioventricu-
lar conduction; the predominant effect was the
activation of multifocal ventricular ectopic beats
interrupted by transient ventricular tachyca rdia
or fibrillation (Kayaalp et a!., 1970). A terminal
ventricular fibrillation leading to death of the
animal occurred in one hour or less after the ia.
injection of BTX. It was suggested that the
mechanism of action of BTX on cardiac tissue
was at least in part correlated with the ability
of the poison to cause membrane depolarization
Received for publication July 30, 1970.‘in part by U.S. Public Health Serv-
ice Grants NB-08233 (E.X.A.), Heart Associationof Western New York, Inc. 9505-4580 and U.S.Public Health Sen-ice, National Heart and LungInstitute HE 12780-01 (P.M.H.).
2 Present address: Department of Physiology,Schools of Medicine and Dentistry, State Univer-sitv of New York at Buffalo. Buffalo. N.Y. 14214.
Send reprint requests to: Dr. E. X. Albuquer-que. Department of Pharmacology. Schools ofMedicine and Dentistry. State University of NewYork at Buffalo, 122 Capen Hall, Buffalo. N.Y.14214.
due to a selective increase in sodium permeability
(Kayaalp et al., 1970: Albuquerque et al., 1971).
The objective of the present work was to
analyze in sonic detail the effects of BTX on the
elect rogenic properties of heart Purkinje fibers
of young a(lult (logs. Based on the assumption
that the effect of BTX on Purkinje fibers was
essentially similar to that observed by Xarahashi
et al. (1970) on the giant axon of the squid
Loligo pealii and by E. X. Albuquerque and M.
Sasa (manuscript in preparation) on thie lobster
and crayfish giant axons, particular attention
was given to the action of the poison on the
resting membrane and action potentials of Pur-
kinje fibers. Our results demonstrate that BTX is
a highily toxic compound. Its toxicity was mani-
fested by a complete membrane depolarization
of the Purkinje fibers; however, prior to the de-
polarization significant alteration of the action
potential generat ing mechanism was observed.
These effects of BTX on Purkinje fibers were
antagonize(l by tetrodotoxin (TTX). The possi-
bihitv of two sodium channels functionally differ-
ent is discussed.

E �1
530 HOGAN AND ALBUQUERQUE Vol. 176
METHODS. Animal and preparation. Hearts wereexcised from young adult dogs anesthetized with
sodium pentobarbital (30 mg/kg) injected iv. andwere placed immediately in oxygenated Tyrode’s
solution. Purkinje fibers were obtained from the
ventricles by removing the papillary muscles withattached false tendons. This preparation wasmounted by means of stainless-steel insect pins to
a paraffin block in the tissue bath, and it was
bathed continuously with Tyrode’s solution equil-ibrated with a gas mixture containing 95% O�-
5% CO2.Recording details. The resting membrane poten-
tial and action potential of the heart Purkinje fiber
were recorded by means of conventional glassmicro-electrode techniques. The micro-electrodes
were filled with 3 M KC1, and they had a resist-ance of 10 to 20 megohms. The potentials were
continuously monitored on a Tektronix (type 502)
oscilloscope; representative potentials were dis-played on a Tektronix (type 565) oscilloscope andphotographed with a Grass (model C4) kymo-graph camera. The resting membrane potentialwas continuously recorded on a polygraph. Rou-tine calibration was done by applying 100 mV rec-
tangular pulses between the indifferent electrode
and recording amplifier.
The maximum velocity of the action potentialupstroke was determined by two methods. The first.method involved the graphic display of the action
potential upstroke recorded at a ral)id oscilloscopesweep from which the velocity of the upstrokecould be determined. The second method used afirst. derivative (R = 1 megohm, C = 100 pico-
______#Fic. 1. Diagrammatic representation of a typi-
cal action potential of the heart Purkinje fiber. Thetechnique used to measure the various membranepotentials is indicated. A, resting membrane poten-tial (maximum diastolic potential); B, amplitudeof action potential; C, amplitude of overshoot; D,time to repolarize to -60 mY; E, amplitude ofoutput signal from the electronic differentiatorwhich is proportional to the rate of rise of the ac-tion potential upstroke (dV/dt). Time and voltagecalibrations are indicated.
farmids) circuit for electrical differentiation of theupstroke of the action potential.
Figure 1 shows the features of the Purkinje fiber
potentials measured in this study. Since Purkinjefibers do not exhibit a steady resting potential, the
resting membrane potential was taken as the maxi-
mum level of polarization reached after each ac-tiomi potential. In order to detect a chiange in the
duration of the action potential the time requiredto repolarize to -60 mY was measured. This memis-
urement was made because -60 mY is the approxi-mate minimal level of transmembrane potentialnecessary to elicit conducted action potentials in
Purkinje fibers (Weidman. 1955a; Hoffman et a!.,
1957).The preparation was stimulated by means of a
Tektronix (type 161) pulse generator which dcliv-
erect a rectangular pulse of 1 msec duration at. aregular rate (95/mm) through a silver bipolarelectrode in contact with the endocardial surface.In some experiments depolarizing and hyperpolar-izing current pulses were introduced through asecond micro-electrode and the distance betweenthe recording and stimulating electrodes was keptat 150 to 200 �.
Solution and drugs. The Tyrode’s solution hadthe following composition (mihimolar concentra-
tion): NaC1, 137; KC1. 2.7; CaCl,, 2.7; MgCl,.
0.5; NaH,P04, 1.8; NaHCO3, 25.0; and dextrose,5.5. The pH of the solution was 7.2 to 7.3. Thetemperature of the solution was 37#{176}C.
One millimolar Na-Tyrode’s solution was pre-
pared by isosmotic replacement of sodium by su-crose. When the calcium chloride concentration
was increased to 162 mM, the NaH,P04 was re-100 duced to 0.5 mM and a pH of 72 was kept through-
.!�. out the experiments.BTX and TTX were applied to the preparation
by adding them in appropriate concentration to
normal Tyrode’s solution. For each experiment I
mY mg of crystalline TTX (Sankyo Co. Ltd., Tokyo,Japan) was dissolved in 10 ml of distilled water.From this further dilution was made in physio-logic solution.
RESULTS. Effects of batrachotoxin on the rest-
ing membrane potential of heart Purkinje fibers.
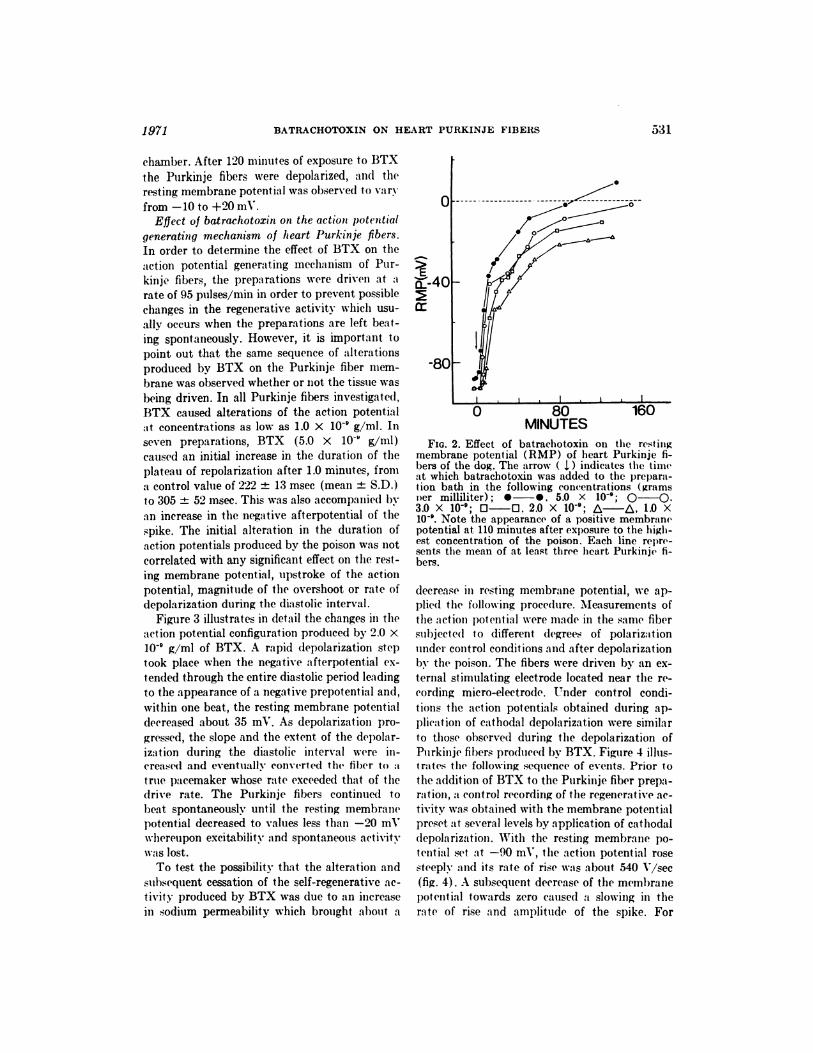
BTX, at concentrations of 1.0 to 5.0 X 10� g/ml,
causes a complete and irreversible depolarization
in all heart Purkinje fibers investigated. The
time course and degree of depolarization pro-
duced by BTX were found to be dose-dependent
(fig. 2). The decrease in resting membrane po-
tential of the Purkinje fibers at the above men-
tioned concentrations occurred about 2.5 minutes
after the poison had reached the experimental
�-40
cr
-80 -
I �ll
1971 BATRACHOTOXIN ON HEART PURKINJE FIBERS 531
chamber. After 120 minutes of exposure to BTX
the Purkinje fibers were depolarized, and the
resting membrane potential was observed to vary
from -10 to +20 mV.
Effect of batrachotoxin on the action potential
generating mechanism of heart Purkinje fibers.
In order to determine the effect of BTX on the
action potential generating mechanism of Pur-
kinje fibers, the preparations were driven at a
rate of 95 pulses/mm in order to prevent possible
changes in the regenerative activity which usu-
ally occurs when the preparations are left beat-
ing spontaneously. However, it is important to
point out that the same sequence of alterations
produced by BTX on the Purkinje fiber macrn-
brane was observed whether or not the tissue was
being driven. In all Purkinje fibers investigated,
BTX caused alterations of the action potential
at concentrations as low as 1.0 X 10� g/ml. In
seven preparations, BTX (5.0 X 10#{176}g/ml)
caused an initial increase in the duratiomi of the
plateau of repolarization after 1.0 minutes, from
a control value of 222 ± 13 msec (mean ± S.D.)
to 305 ± 52 msec. This was also accompanied by
an increase in the negative afterpotential of the
spike. The initial alteration in the duration of
action potentials produced by the poison was not
correlated with any significant effect on the rest-
ing membrane potential, upstroke of the actiomi
potential, magnitude of the overshoot or rate of
depolarization during the diastolic interval.
Figure 3 illustrates in detail the changes in the
action potential configuration produced by 2.0 x10�� g/ml of BTX. A rapid depolarization step
took place when the negative afterpotential ex-
tended through the entire diastolic period leading
to the appearance of a negative prepotential and,
within one beat, the resting membrane potential
decreased about 35 mY. As depolarization pro-
gressed, the slope and the extent of the depolar-
ization during the diastolic interval were in-creased and eventually converted the fiber to :1
true pacemaker whose rate exceeded that of the
drive rate. The Purkinje fibers continued to
beat spontaneously until the resting membrane
potential decreased to values less than -20 mY
��‘hereupon excitability and spontaneous activity
was lost.
To test the possibility that. the alteration and
subsequent cessation of the self-regenerative ac-
tivity produced by BTX was due to an increase
in sodium permeability which brought about a
0 80 160MINUTES
Fia. 2. Effect of batrachotoxin on the rest ingmembrane potential (RMP) of heart Purkinje fi-bers of the dog. The arrow ( 1.) indicates the timeat which batrachotoxin was added to the prepara-tion bath in the following concentrations (gramsncr milliliter); #{149}-�, 5.0 X l0’; Q-�,3.0 X 10�; 0-0, 2.0 X 10�; � 1.0 X10�. Note the appearance of a positive membranepotential at 110 minutes after exposure to the high-est concentration of the poison. Each line repre-sents the mean of at least thiree heart Purkinje fi-hers.
decrease in resting membrane potential, we ap-
plied the following proce(lure. Measurements of
the action potential were made in the same fiber
subjected to different degrees of polarization
umndei’ control conditions and after depolarization
by the poison. The fibers were driven by an ex-
ternal stimulating electrode located near the re-
cording micro-electrode. Under control condi-
tions the action potentials obtained during ap-
plicat ion of cathodal depolarization were similar
to those observed during the depolarization of
Purkinje fibers produced by BTX. Figure 4 illus-trates thie following sequence of events. Prior to
the addition of BTX to the Purkinje fiber prepa-
ration, a control recording of the regenerative ac-
tivity was obtained with the membrane potential
preset at several levels by application of cathodal
depolarization. With the resting membrane po-
tential set at -90 mY, the action potential rose
steeply and its rate of rise was about 540 V/sec
(fig. 4). A subsequent decrease of the membrane
potential towards zero caused a slowing in therate of rise and amplitude of the spike. For

600
0
300
0
0
#{163}
5”
#{163} 5
#{163}
A.
S�-A #{149}
#{149}5.#{149}o
#{149}#{149}S.#{149}.#{149}
0 -80-40Em(mV)
Fia. 4. Effect of batrachotoxin omu the relation-ship between the upstroke velocity (cl1� It) of theaction potential of heart Purkinje fibers and se-lected levels of membrane potential (Em). Thefibers were polarized to the desired level of mem-brane potential before an(l during exposure to ha-trachotoxin (2.0 X 10� g/ml) by passimig a currentthrough a second intracellular mnicroelectrode. Thesymbols indicate the values obtained before (#{149})and after exposure to batrachotoxin for 2.5 (0);5.0 (A); 7.5 (M; and 10.0 minutes (O�.
FI(;. 3. Effect of hat rachot OXlfl on t he tct iOU P0-
tent ial amid its first derivat we recoulid fi’omu t heheart Purkinje fiber of the (log. The records shownare the control (C) and at tIme I imiies iIi(li(at ed (luir-ing exposure to batrachotoxin (2.0 X 10#{176}g’mnl).All records are from the samn( fiber I he prep:mra-
ion was (triven b�’ an external stimiiuihuis applied at
the rate of 95 pulses’min. Ijmper vertical scale.100 mY ; lower vertical scale, 425 V sec horizon-tal scale. 200 msec.
532 HOGAN AND ALBUQUERQUE Uol. 176
example, a depolarization of 23 mY from the
control membrane potential reduced the rate of
rise of t lie spike to :mhout 270 V, sec wit Ii simul-
taneous disappearance of the overshoot. When
BTX was admitted to the l)at hung fluid amid de-
l)olariza t ion had begun, a nodal hyper�)ola rizationof the fiber restored the inemnbrane potential to
the coiit rot level. This method caused an increase
in the (lepolarizing effect of BTX, as revealed by
the fact that while complete depolarization by
the poison in nonpolarized preparations took
about. 120 minutes, in the present case the mciii-
brane l)otential reached values from -10 to 0
mV in 10 to 15 minutes (fig. 5). Omi all occasions,
however, ��huen the membrane potential was re-
store(l by anodal hvperpolarization, the rate of
rise and the amplitude of the action potential
were similar to control (figs. 4 and 5). It is
interesting to note that. after five mimiuites of cx-
postire to BTX (2.0 x 10#{176}g ml), �vhien a (he-
polarization of only 20 mY had occurred, an in-crease in the upstroke velocity of the spike po-
teiitial could be Oh)served if an anodal hvper-
polarization brought the membrane potential tocontrol levels. On many occasioiis during the
action of BTX i�’hien the membrane potential was
set at -SO mY, the rate of rise of the regenera-
tive activity was increased by 35’� fig. 4).
With ammodal hyperpolarization oiie can show

1971 B.�TRACHOTOXIX OX IIEA RT PURKI XJE FIBERS 533
FIG. 5. Effect of anodal hyperpolarization on the heart Purkinje fiber depolarized by batrachotoxin.Record A is the control action potential; records B, C and 1) are the action potentials obtained duringexposure to batrachotoxin (2.0 X 10_i g/m.l) at 5, 10 and 15 minutes, respectively. The arrows (�indicate the beginning of anodal hyperpolarization. The second segment of records B, C amid D arediscontinuous with the first but taken in close temporal sequence. Vertical scale, 100 mY; horizontal
scale, 150 msec.
that the disappearance of the action potential
induced by BTX was directly correlated with a
decrease in membrane potential. This method
was rather unsuitable when the membrane po-
tential was reduced to about 50% from control;
under this condition damage to the tissue and
electrodes was encountered due to the necessity
of passing large hyperpolarizing currents. In a
few cases even when the membrane potential was
as low as -10 mY, anodal hyperpolarization al-
lowed the appearance of spike potentials quanti-
tatively similar to the control spikes (fig. 5).
Effects of batrachotoxin on ionic mechanisms
involved in action potential generating mecha-
nisms. The depolarization of Purkinje fibers
caused by BTX is compatible with the suggestion
that the resting sodium conductance is increased
(Albuquerque et at., 1971; Warnick et al., 1971),
Several experiments were therefore carried out
in which the external sodium concentration was
altered prior to and after the action of BTX on
the Purkinje fiber membrane.
When the Purkinje fibers were treated with 1
mM Na-sucrose solution prior to the addition
of BTX, a hyperpolarization of 2 to 5 mV of the
fiber membrane was observed; upon washing the
preparation with normal Tyrode’s solution, the
resting membrane potential returned to the con-
trol level, i.e., about -90 rnV. In these experi-
Inents, six preparations were exposed to 2.0 X
10#{176}g/ml of BTX for about 15 mintites, the
membrane was depolarized to values of the order
of -25.7 ± 5.3 mY whereas the control resting
membrane potential was -87.7 ± 5.9 mY. Wash-
ing of these preparations with 1 mM Na-sucrose
solution restored the membrane potential to
-86.6 ± 5.3 mV. When the Purkinje fibers were
exposed to normal Tyrode’s after being treated
with 1 mM Na-sucrose solution, the resting
membrane potential returned in about 20 seconds
to the depolarized state with values of -18.9 ±
3.9 mY (fig. 6). It is important to mention thatprolonged exposure to 1 mM Na-sucrose solution
caused a considerable deterioration of the prep-
aration. Therefore, the exposure to the Na#{176}-
deficient solution was never for more than 10
minutes. This time proved to be sufficiently long
to allow a complete recovery of the resting mem-
brane potential of the Purkinje fibers depolarized
by BTX.
Another way in which one could study the par-
ticipation of sodium ions on the regenerative
activity is by using TTX. This toxin interferes
with the self-regenerative activity by selectively

BTX IlmMNaI N�TyrC
-50
0 20
MINUTES
9
0�
ii
-100
FIG. 6. Effect of low extracellular sodium con-centration on the resting membrane potential ofthe heart Purkinje fiber previously exposed to ba-trachotoxin (BTX). The resting membrane poten-tial (RMP) before exposure to batrachotoxin(2.0 X 10 g/ml) was -85 mV and after 15 min-utes about -25 mY. Normal Tyrode’s solution (N-Tyr).
blocking passive sodium permeability. The con-
centration of TTX that effectively blocks the ac-
tion potential generating mechanisms of the
squid giant axons and lobster stretch receptor
(Narahashi et a!., 1964; Albuquerque and
Grampp, 1968) is rather ineffective on heart
Purkinje fibers (Dudel et a!., 1967). For instance,
1.5 X 10� g/ml of TTX did not block the action
potential of the Purkinje fiber; however, a mem-
brane hyperpolarization of 2 to 5 mY was ob-
served. This concentration of TTX was applied
to the fiber for 15 minutes prior to the addition
of BTX (1.0 X 10� g/ml) to the TTX-Tyrode’s
solution. Within the scope of our experiments,
this concentration of TTX was sufficient to pre-
vent the effects of BTX on the spike activity and
resting membrane potential for as long as 30
minutes. After 20 minutes exposure to BTX and
TTX, the preparation was washed with normal
Tyrode’s solution; in two minutes depolarization
began and after 20 minutes resting membrane
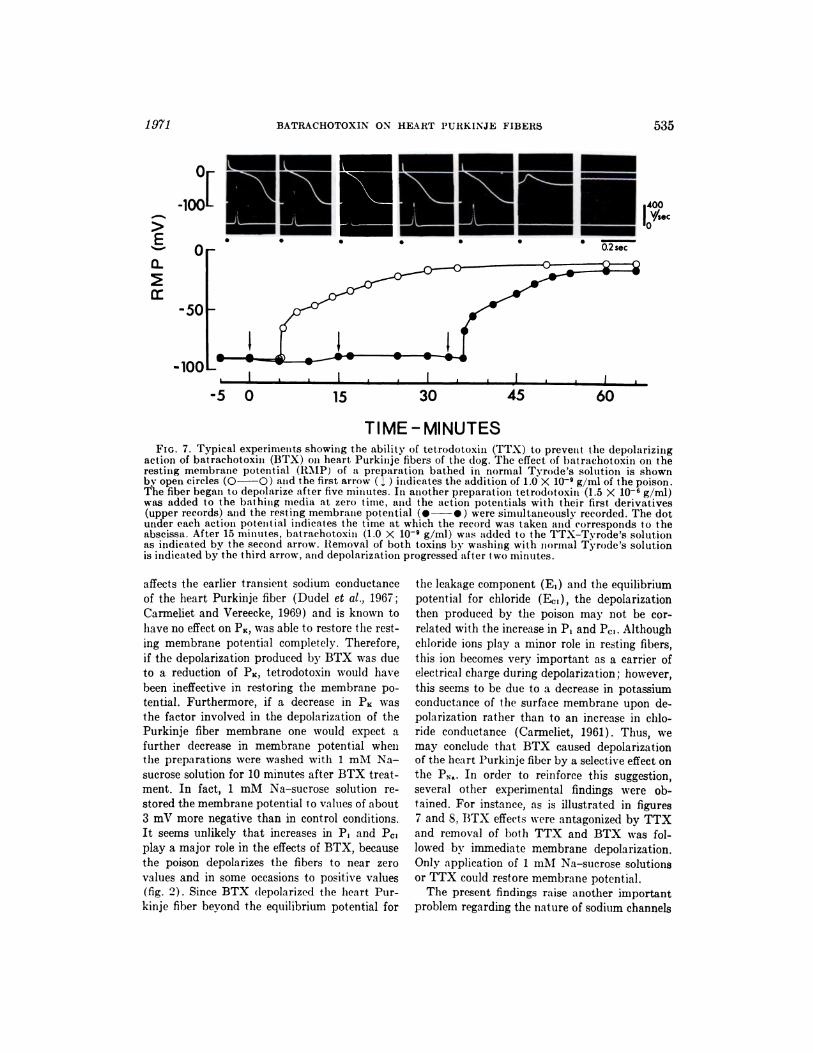
potential had decreased to -15 mV (fig. 7). Con-
centrations of TTX below 1.5 x 106 g/ml pro-
longed the onset and time course of BTX effects,
but did not prevent the effects of the poison on
534 HOGAN AND ALBUQUERQUE Vol. 176
Purkinje fibers. In other experiments, when a
preparation previously depolarized by BTX was
subjected to TTX (1.5 x 10� g/ml), a complete
repolarization of the fiber membrane to control
levels was recorded. In a few cases the repolar-
ized level exceeded the control level. As the rest-
ing membrane potential increased, the upstroke
velocity, amplitude and duration of the action
potential approached those values recorded in
control conditions. However, under such condi-
tions, the terminal phase of the spike potential
was prolonged and a typical negative afterpo-
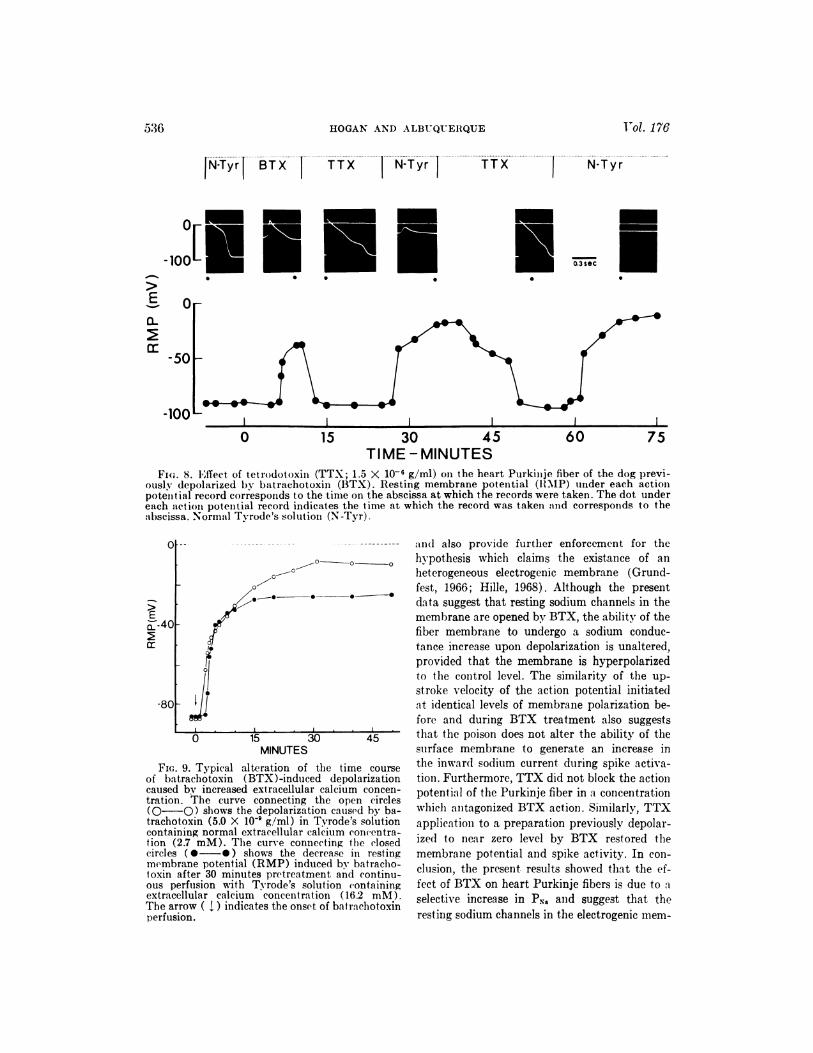
tential could be observed (fig. 8).
Calcium ions have been shown to affect mem-
brane permeability to other ions and to exert a
membrane “stabilizing” effect on Purkinje fibers
#{149}j (Weidmann, 1955b). The possibility that high
4 0 extracellular calcium might antagonize the de-
polarizing effect of BTX was therefore tested.
In the experiment illustrated by figure 9, Pur-
kinje fibers were pretreated for 30 minutes with
Tyrode’s solution containing 16.2 mM Ca�. Dur-
ing continued exposure of the preparation to 16.2
mM Ca�-Tyrode’s solution, BTX (2.0 X 10�
g/ml) was applied to the Purkinje fibers. The
presence of high external calcium concentrations
in the physiologic solution delayed the onset and
extent of depolarization produced by BTX but
did not prevent the action of the poison. It is
therefore likely that the action of calcium on the
fiber membrane may be the result of a stabiliz-
ing action of the ion rather than any competitive
interaction with BTX.
DIscussIoN. In the heart Purkinje fibers of
the dog BTX at concentrations of 1.0 to 5.0 x10� g/ml caused complete depolarization of the
fiber membrane and rendered it inexcitable. The
depolarization produced by BTX was irreversible
and persisted after removal of the poison from
the bathing fluid by washing the preparation
with normal Tyrode’s solution.
Several factors may be responsible for the de-
polarization of Purkinje fibers produced by
BTX. First, there is the possibility of an in-
crease in sodium permeability (P�j; secondly,
there is the possibility of a decrease in potassium
permeability (PK), an increase in chloride per-
meability (P�1) and an increase in the leakage
permeability (P1), or a combination of all of
these factors. The possibility that a decrease in
PK would occur after treating the Purkinje fiber
with BTX can be ruled out because TTX, which
BATRACHOTOXIN ON HEART PURKINJE FIBERS1971
0�
535
0
-100
0
-50
-100I I I - 1 _l
-5 0 15 30 45 60
TIME - MINUTESFIG. 7. Typical experiments showimig the ability of tet.rodotoxiiu (TTX) to prevent the depolarizing
action of batrachotoxin (BTX) on heart Purkinje fibers of the dog. The effect of batrachotoxin omi theresting membrane potential (RMP) of a preparation bathed in normal Tyrode’s solution is shownby open circles (O-O) amid the first arrow (1) imidicates the addition of 1.0 X 10� g/ml of the poison.The fiber began to depolarize after five mimiutes. In another preparatiomi tetrodotoxin (1.5 X 106 g/ml)was added to the baihimig media at zero time, and the action potentials with their first. derivatives(upper records) amid the resting membrane potential (.-.) were simultaneously recorded. The dotunder each action potential indicates the time at which the record was taken amid corresponds to theabscissa. After 15 minutes, batrachotoxin (1.0 X 10� g/ml) was added to the TTX-Tvrode’s solutionas indicated by the second arrow. Removal of both toxins by washing with miormnal Tyrode’s solutionis indicated by the third arrow, and depolarization progressed after two minutes.
affects the earlier transient sodium conductance
of the heart Purkinje fiber (Dudel et al., 1967;
Carmeliet and Vereecke, 1969) and is known to
have no effect on PK, was able to restore the rest-
ing membrane pot.eiitial completely. Therefore,
if the depolarization produced by BTX was due
to a reduction of PK, tetrodotoxin would have
been ineffective in restoring the membrane po-
tential. Furthermore, if a decrease in PK was
the factor involved in the depolarization of the
Purkinje fiber membrane one would expect a
further decrease in membrane potential when
the preparations were washed with 1 mM Na-
sucrose solution for 10 minutes after BTX treat-
ment. In fact, 1 mM Na-sucrose solution re-
stored the membrane potential to values of about
3 mV more negative than in control conditions.
It seems unlikely that increases in Pm and P�1
play a major role in the effects of BTX, because
the poison depolarizes the fibers to near zero
values and in some occasions to positive values
(fig. 2). Since BTX depolarized the heart Pur-
kinje fiber beyond the equilibrium potential for
the leakage component (Es) and the equilibrium
potential for chloride (Efl), the depolarization
then produced by the poison may not be cor-
related with the increase in P� and Pcm. Although
chloride ions play a minor role in resting fibers,
this ion becomes very important as a carrier of
electrical charge during depolarization; however,
this seems to be due to a decrease in potassium
conductance of the surface membrane upon de-
polarization rather than to an increase in chlo-
ride conductance (Carmehiet, 1961). Thus, we
may conclude that BTX caused depolarization
of the heart Purkinje fiber by a selective effect on
the P5k. In order to reinforce this suggestion,
several other experimental findings were ob-
tained. For instance, as is illustrated in figures
7 and 8, BTX effects were antagonized by TTX
and removal of both TTX and BTX was fol-
lowed by immediate membrane depolarization.
Only application of 1 mI’iI Na-sucrose solutions
or TTX could restore membrane potential.
The present findings raise another important
problem regarding the nature of sodium channels
0-
0.3 sec
#{149}50 -
-100 -
0 15 30 45 60 75TIME -MINUTES
Fm;. 8. Effect. of tetrodotoxin (TTX; 1.5 X 10’ g/ml) on the heart Purkimije fiber of the dog previ-ously depolarized by bat rachotoxin (BTX). Resting membrane potential (RMP) under each actionpotemitial record corresponds to the time on the abscissa at which the records were taken. The dot undereach act iou potential record indicates the time at which the record was taken and corresponds to theabscissa. Normal Tyrode’s solution (N-Tyr).
0
>E
�-4C
-8C
1530 45MINUTES
FIG. 9. Typical alteration of time time courseof batrachotoxin (BTX)-induced depolarizationcaused by increased extracellular calcium concen-tration. The curve connecting the open circles(O-O) shows the depolarization caused by ba-trachotoxin (5.0 X l0’ g/ml) in Tyrode’s solutioncontaining normal cxtracellular calcium concentra-lion (2.7 mM). The curve connecting the closedcircles (#{149}-#{149}) shows the decrease in restingmembrane potential (RMP) induced by batracho-toxin after 30 minutes pretreatment and continu-ous perfusion with Tyrode’s solution (‘ontainingextracellular calcium concentration (16.2 mM).The arrow ( �.) indicates the onset of bat rachotoxinperfusion.
536 HOGAN AND ALBUQUERQUE Vol. 176
>
E
a-
N-Tyr� BTXlix N�Tyr TTX N-Tyr
� - amid also provide further enforcement for the
hypothesis which claims the existance of an
heterogeneous electrogenic membrane (Grund-
fest, 1966; Hille, 1968). Although the present
data suggest that resting sodium channels in the
membrane are opened by BTX, the ability of the
fiber membrane to undergo a sodium conduc-
tance increase upon depolarization is unaltered,
provided t.hat the membrane is hyperpolarized
to the coiitrol level. The similarity of the up-
stroke velocity of the action potential initiated
at identical levels of membrane polarization be-
fore and during BTX treatment also suggests
that the poison does not alter the ability of the
surface membrane to generate an increase in
the inward sodium current during spike activa-tion. Furthermore, TTX did not block the action
potential of the Purkinje fiber in a concentration
which antagonized BTX action. Similarly, TTX
application to a preparation previously depolar-
ized to near zero level by BTX restored the
membrane potential and spike activity. In con-
clusion, the present. results showed that the ef-
fect of BTX on heart Purkinje fibers is due to a
selective increase in PN5 and suggest that theresting sodium channels in the electrogenic mem-

1971 BATRACHOTOXIN ON HEART PURKINJE FIBERS 537
brane may be functionally different from the
sodium channels for membrane excitation.
ACKNOWLEDGMENTS. We gratefully acknowl-
edge the helpful discussions and comments of
Dr. J. Daly and Dr. J. E. Warnick as well as
the technical assistance of Miss Dolores Timmer
and Miss Druscilla Latta. The authors were in-
debted to Dr. J. Daly and Dr. B. Witkop for
their generous supply of batrachotoxin used dur-
ing the course of this investigation.
REFERENCES
AisuQuzs�uE, E. X. AND Gu�qau’p, W.: Effects oftetrodotoxin on the slowly adapting stretch re-ceptor neurone of lobster. J. Physiol. (London)195: 141-156 1968.
ALBUQUERQUE, E. X., W�sr�mIcK, J. E. AND SANSONE,F. M.: The pharmacology of batrachotoxin. II.Effect on electrical properties of the mammaliannerve and skeletal muscle membranes. J. Phar-macol. Exp. Ther. 176: 511-528, 1971.
CAIis�uirr, E.: Chloride ions and the membranepotential of Purkinje fibres. J. Physiol. (London)156: 375-388, 1961.
CLEUE’r, E. AND VEREECEK, J.: Adrenaline andthe plateau phase of the cardiac action potential-IMPORTANCE OF � NA� MTh K� CONDUCTANCE.
PfiUgers Arch. Gesemte Physiol. Menschen Tiere313: 300-315, 1969.
DUDEL, J., Pzpxii, K., Rtinsn, R. �n TRAUTWEIN,W.: The effect of tetrodotoxin on the membranecurrent in cardiac muscle (Purkinje fibers). Pfl#{252}-
gers Arch. Gesamte Physiol. Menschen Tiere295: 213-226, 1967.
GRUNDFEST, H.: Heterogeneity of excitable mem-brane: Electrophysiological and pharmacologicalevidence and some consequences. Ann. N.Y.Acad. Sci. 137: 901-949, 1966.
HILLE, B.: Pharmacological modifications of thesodium channels of frog nerve. J. Gen. Physiol.51: 199-219, 1968.
HOFFMAN, B. F., KAO, C. Y. AND SUCKLING. E. E.:Refractoriness in cardiac muscle. Amer. J. Phys-iol. 190: 473-482, 1957.
KAYAALP, S. 0., ALBUQUERQUE, E. X. AND WARNICK,
J. E.: Ganglionic and cardiac actions of ba-trachotoxin. Eur. J. Pharmacol. 12: 10-19, 1970.
NARAHASHI, T., MOORE, J. W. AND ScoTr, W.: Tet-rodotoxin blockage of sodium conductance in-crease in lobster giant axons. J. Gen. Phmysiol.47: 965-974, 1964.
NARAHASHI, T., ALBUQUERQUE, E. X. AND DEGUCHI.T.: Effects of batrachotoxin on ionic conduc-tances on giant axon membranes. Biophys. Soc.Annu. Meet. Abstr. 10: 183A, 1970.
WARNICK, J. E., ALBUQUERQUE, E. X. AND SANSONE,
F. M.: The pharmacology of batrachotoxin.I. Effects on the contractile mechanism and onneuromuscular transmission of mammalian skele-tal muscle. J. Pharmacol. Exp. Ther. 176: 497-510, 1971.
WF.misANN, S.: Effects of the cardiac membranepotential on the rapid availability of the sodium-carrying system. J. Physiol. (London) 127: 213-224, 1955a.
WEIDMANN, S.: Effects of calcium ions and localanesthetics on electrical properties of Purkinjefibres. J. Physiol. (London) 129: 568-582, 1955b.