the role of anthocoris species (hemiptera: anthocoridae) in the integrated control of the damson-hop...
TRANSCRIPT

Ann. appl. Biol. (1981), 97, 143-153 Printed in Great Britain
143
The role of Anthocoris species (Hemiptera: Anthocoridae) in the integrated control of the damson-hop aphid (Phorodon humuli)
BY C. AVELING* Hop Research Department, Wye College, Wye, Kent, TN25 5AH, UK
(Accepted 22 September 1980)
SUMMARY
Field studies in 1974, 1975 and 1976 investigated integrated control of the damson-hop aphid Phorodon humuli on hops. After the aphicidal effects of an early-season soil drench of mephosfolan had declined, natural enemies controlled the aphids for the remainder of the season. Anthocorid bugs, particularly Anthocoris nemoralis, were the most abundant predators. In each year a rapid decline in aphid numbers occurred in mid- to late-July, coinciding with the peak numbers of fourth and fifth instar larvae and adults, the most voracious anthocorid stages. Aphids in the cones remained under control for the rest of the season in 1974 and 1975, and increased in 1976 but damaging numbers did not develop. When predators were excluded by caging mephosfolan-treated bines, high aphid densities developed on the leaves, and the cones were heavily infested. Plants not treated with an insecticide were almost completely defoliated by late-July. Heavily infested ‘missed bines’, due to uneven uptake of mephosfolan, attracted large numbers of anthocorids, which later dispersed into the surrounding plants.
I N T R O D U C T I O N
Phorodon humuli (Schrank) is the most serious pest of hops (Humulus lupulus L.) in England. In addition to reductions in yield caused by direct feeding damage to the plant, the cone crop is devalued by the presence of aphids and by the discoloration caused by sooty moulds developing in the aphids’ honeydew. At present P. humuli is controlled by regular applications of pesticides and, since the aphids are very difficult to control with chemicals once they enter the relatively protected environment of the cones, the aim of control measures is to eliminate the aphid before cone formation (Zohren, 1970).
Since the 1960s the increasing resistance of P. humuli to some insecticides in England (Muir, 1979) and in other parts of Europe (Hrdy, 1970) has highlighted the long-term problem of continuing to rely on the prophylactic use of insecticides for control of P. humuli. It is therefore important to develop an integrated approach to control, in which other measures are also applied against the pest, in order to relieve the selection pressure for resistance to chemicals (Smallman, 1965).
Naturally-occurring predators, particularly anthocorid bugs, are known to be important in regulating P. humuli populations on hops (Campbell, 1978). Field studies from 1974 to 1976 investigated the possibility of integrating chemical control, in the form of an early-season soil drench of mephosfolan, with biological control by natural enemies during the rest of the season. Foliar sprays of broad-spectrum insecticides pere either not used, or used only in June when few natural enemies were present. The studies were made at Wye College and in several commercial hop gardens in east Kent and the Weald. Populations of all parasites, pathogens and specific predators were monitored, particular attention being paid to the r61e of anthocorid bugs.
* Present address: c/o ‘Garreg Wen’, Bracken Close, Wonersh Park, Guildford, Surrey GU5 O Q S .
0 198 1 Association of Applied Biologists

144 C . A V E L I N G
M A T E R I A L S AND M E T H O D S
Sampling procedure Random number tables were used to select the first leaf to be sampled and to determine the
string (which normally supports two hop bines) on which sampling should begin. Leaves were counted systematically down each string in turn, and every fortieth leaf (plus lateral when extended) was removed. Later in the season the laterals were so long that only every third leaf was sampled on each lateral and an estimate made for the complete lateral. The internodes were also inspected and all insects present were counted. In I974 the leaves were placed in Polythene bags and the insects counted in the laboratory, but in the following two seasons all insects were counted in the field. Counts were made of adult alates and apterae of P. humuli on all the leaves for each sample. In addition records were made of:
1. the number, stage and species of all predators; 2. the number of parasitised aphids (‘mummies’) with parasites not yet emerged; 3. the number of aphids attacked by entomophthorous fungi.
This sampling procedure underestimated the numbers of parasitised aphids since it did not include those that may have dropped from the plant.
Table 1. Details of trials, 1974-76. (C = Copper oxychloride; D = Dinocap; S = Sulphur; Z = Zineb; Df = Dicofol; P = Pyrazophos; T = Tetradifon; * = until 8 /7 /74; ** =.from
15/7 /74; *** = until 5 /8 /75; #=from 12/8/75)
Plot
Wye a
Wye b
Wye c
Wye d
Selling
WYe
WY e
Boughton
Harefield
Horsmonden
Goudhurst
Date Treatment Rate
17/6/74 Dimefox 0.57 g a.i. soil drench /plant
Mephosfolan 0.5 g a.i. soil drench /plant
11/6/74 Mephosfolan 1.0 g a.i. soil drench /plant
11/6/74 Methidathion 504 g a.i. foliar spray /ha
Mephosfolan 1 .O g a.i. soil drench /plant - - 1974
10/6/74 Mephosfolan 1 .O g a.i. soil drench
18/6/75 Mephosfolan soil drench
8/6/76 Mephosfolan soil drench
9/6/76 Mephosfolan soil drench
28/5/76 Mephosfolan soil drench
/plant 1 .O g a.i.
/plant 1 .O g a.i.
/plant 1 .O g a.i.
/plant 1 .O g a.i.
/plant
9/6/76 Mephosfolan I .O g a.i. soil drench /plant
21/6/76 Acephate 922 g a.i. foliar spray /ha
Mephosfolan 1.0 g a.i. soil drench /plant
Plot Garden Size Size (ha) (ha)
0.03 1-4
0.03 1.4
0.03 1.4
0.1 2.1
1 .o 1 .o
0.5*”* 5.2 0*17# 0.5 5.2
0.7 1.8
0.7 2.7
0.8 4.9
0.5 4.2
No. Acaricides strings and
sampled Fungicides Variety
120* C,D,S, x7 Cobbs 60’.
WYe
WYe
- 40
40 Challenger
Challenger -
60
60
4 0
40
40
4 0
40
4 0
-
z,s, x12
C, D, S, x 6
D, S, x 9
D,S, x 9 Df x 3 C, Z, D,
Df,T x 2 Df,T x 3
Z x 16 Df x l
P, S, x13
WYe Challenger
Canterbury Golding
WY e Northdown
W w Northdown
WYe Challenger
Cobbs
WYe
W e Challenger
Target

Anthocorids in integrated control of hop aphid 145
On each sampling date from mid-August onwards, three cones from each of 30 strings/plot in 1974, and 10 cones from each of 20 strings/plot in 1975 and 1976, were taken at random and all stages of aphids and predators found within the cones were counted in the laboratory.
Trial plots Table 1 gives details of the trials. Drenches were applied to the soil around the crown of each
plant and, incidentally, to the base of the bine. They were applied either by a hand lance or by a tractor-mounted Phillips applicator.
Exclusion cage studies Twelve predator exclusion cages (Campbell, 1978) were used in 1975 and 1976. The cages
were made of Terylene netting, with 400 apertures/cm*, and were 2 m long with a diameter of 0.5 m. They were fitted over the strings and sealed at both ends. In 1975 eight predator exclusion cages and four open-ended controls were fitted over randomly chosen strings on 4 August. All predators that could be found were removed from the exclusion cages. Each week 10% of the foliage present was sampled in the exclusion and open cages and also on four uncaged strings. After cone formation 10 conedstring were sampled weekly. In 1976 the cages were fitted on 13 July.
Although cages slightly altered the microclimate of the bines due to increased shading (Campbell, 1978) there was no difference in the rate of development of aphids in open-ended cages and on uncaged plants in 1975, confirming the results of Campbell (1978) and Buxton (1974). Therefore in 1976 open-ended cages were not used, only uncaged strings.
‘Banker’ plants In 1975 a few plants were deliberately left untreated and these became heavily infested with
aphids, and therefore subsequently with good numbers of anthocorids; because they were potentially a source of adult anthocorids for the surrounding plants these were termed ‘banker’ plants. The plot comprised 540 plants in 30 rows, of which 10 plants (in the first 15 rows) were ‘banker’ plants.
R E S U L T S
Season 19 74 In plot a at Wye dimefox, a systemic organophosphorus aphicide which had become
increasingly less effective against P. humuli, was used in order to maintain a high aphid population. The populations of aphids and predators under these circumstances could then be compared with the other sites at Wye and Selling where a single application of mephosfolan, a more effective systemic organophosphorous aphicide, was used.
Fig. 1 shows numbers of aphids, anthocorids and other predators in plot a at Wye. Predators, mainly anthocorids, arrived at least a month after the first aphids, by which time a large aphid population had built up. Three anthocorid species were identified - Anthocoris nemoraIis (Fab.), A. confusus Reuter (both recorded as A. nemoralis because of the difficulty of identifying A . confusus larvae (Sands, 1957)), and A . nemorum (L.). Despite a half-strength mephosfolan application on 30 June aphid numbers did not begin to fall rapidly until after 5 July, by which time a large predator population had developed. The poor effectiveness of the dimefox was probably due to a combination of aphid resistance and dry soil preventing adequate uptake of insecticide.
On the untreated plot d at Wye the plants were completely defoliated by mid-June before a predator population was established, while on the mephosfolan-treated plots at Wye (b and c)
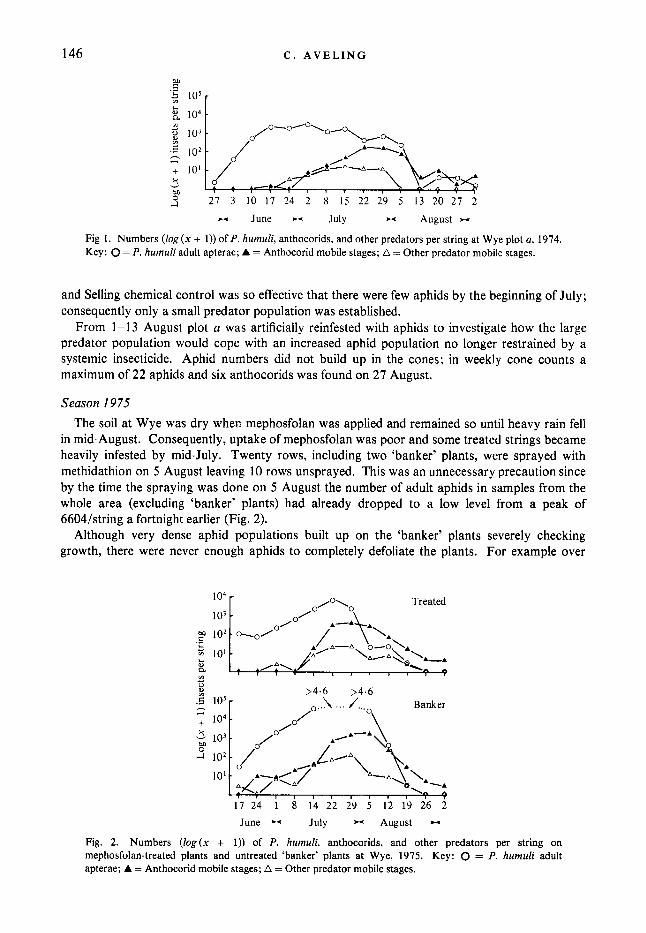
146
M .-
B 104 I 0 l C . A V E L I N G
5 27 3 10 17 24 2 8 15 22 29 5 13 20 27 2
H June M July H August H
Fig 1. Numbers ( log (x + 1)) of P. humuli, anthocorids, and other predators per string at Wye plot a, 1974. Key: 0 = P. humuli adult apterae; A = Anthocorid mobile stages; A = Other predator mobile stages.
and Selling chemical control was so effective that there were few aphids by the beginning of July; consequently only a small predator population was established.
From 1-13 August plot a was artificially reinfested with aphids to investigate how the large predator population would cope with an increased aphid population no longer restrained by a systemic insecticide. Aphid numbers did not build up in the cones; in weekly cone counts a maximum of 22 aphids and six anthocorids was found on 27 August.
Season 19 75 The soil at Wye was dry when mephosfolan was applied and remained so until heavy rain fell
in mid-August. Consequently, uptake of mephosfolan was poor and some treated strings became heavily infested by mid-July. Twenty rows, including two ‘banker’ plants, were sprayed with methidathion on 5 August leaving 10 rows unsprayed. This was an unnecessary precaution since by the time the spraying was done on 5 August the number of adult aphids in samples from the whole area (excluding ‘banker’ plants) had already dropped to a low level from a peak of 6604/string a fortnight earlier (Fig. 2).
Although very dense aphid populations built up on the ‘banker’ plants severely checking growth, there were never enough aphids to completely defoliate the plants. For example over
.- M
L Y
L W a * * .- x h - + Y
cl
v
$
17 24 1 8 14 22 29 5 12 19 26 2 June H July H August H
Fig. 2. Numbers ( log(x + 1)) of P. humuli, anthocorids, and other predators per string on mephosfolan-treated plants and untreated ‘banker’ plants at Wye, 1975. Key: 0 = P. humuli adult apterae; A = Anthocorid mobile stages; A = Other predator mobile stages.

Anthocorids in integrated control of hop aphid 147
40000 adult apterae per string were recorded on 22 and 29 July (Fig. 2). Aphid numbers declined on the ‘banker’ plants, reaching a low level a week after control was achieved on the treated plants. Later secondary growth occurred on the ‘banker’ plants with the formation of a few cones on some strings.
Fig. 2 shows that predators, predominantly anthocorids, were attracted first to the ‘banker’ plants and later large populations developed on both treated and ‘banker’ plants. Their numbers declined as the aphid populations declined in early August.
Season 1976 Because of the drought in 1976 the soil was exceptionally dry before and after mephosfolan
was applied, except in the two irrigated commercial gardens, Horsmonden and Goudhurst. Uptake of mephosfolan at Wye and Harefield was poor; aphid numbers increased rapidly during June, reaching maxima of 5054 and 8776 adult apteraehtring respectively at the end of June (Fig. 3). However uptake in the non-irrigated garden at Boughton was better despite the similar dry conditions. A maximum of 2262 adult apterae/string was recorded in this garden compared with a maximum of 1712/string in the irrigated garden at Horsmonden. These aphid numbers were not considered serious by the growers in contrast to the numbers at Wye and Harefield (see later). In the irrigated garden at Goudhurst, which received a foliar spray of acephate the day before mephosfolan was applied on 22 June, only three adult apterae were found during the season.
At Wye and Harefield most plants examined during the third week of June were heavily infested with P. humuli, but many of these were heavily infested only in the top third of the bine. Possibly lack of water prevented translocation of mephosfolan to the top of the plant. In the other gardens, as in previous years, the ‘missed bine’ phenomenon was also observed (the
- BOUGHTON
‘ 0
/O---O--O,o HAREFIELD
\
HORSMONDEN 1 0 4
15 22 29 6 13 20 27 3 10 17 24 31 June y July H August *
Fig. 3. Numbers (log (x + 1)) of P. hurnuli, and fourth and fifth instar larvae and adult anthocorids, per string at Boughton, Harefield and Horsmonden, 1976. Key: 0 = P. hurnuli adult apterae; A = fourth and fifth instar larvae and adult anthocorids.

148 C . A V E L I N G
occasional heavily infested bine due to poor mephosfolan uptake). At Horsmonden the percentage of ‘missed bines’ (including those heavily infested at the top of the bine only) was estimated at 3% on 1 July. Although the sampling procedure gives a good estimate of the relative numbers of aphids and predators, it occasionally gave a misleadingly high estimate of the numbers per string, when a sample in which most plants supported a low number of aphids included a ‘missed bine’.
On 25 June the Wye plot was sprayed with methidathion to prevent permanent aphid damage to the plants. It was hoped that, by spraying at the end of June, only a few natural enemies would be killed, and those arriving in early July would be able to prevent any further aphid build-up.
As in previous years the most abundant predators were anthocorid bugs, and their numbers increased in all gardens in response to the aphid build-up. A rapid aphid population decline occurred between 13 and 20 July followed closely by a decrease in anthocorid numbers (Fig. 3).
Table 2. Numbers of Phorodon humuli in 200 cones from Boughton, Harejeld and Horsmonden in 1976. Percentage of cones aphid infested in parentheses
Gardens.. . Boughton Harefield Horsmonden Datesof r-*-, (-Ap, & sampling Adults Nymphs Adults Nymphs Adults Nymphs
- - - - 3 Aug. 1 6 (1) 9-10Aug. 5 43 (17) 0 1 (1) 0 4 (2)
16-17A~g. 12 80 (23) 0 0 (0) 0 13 ( 5 )
6 Sept. 92 461 (38) 42 213(12) 13 47 ( 5 )
23-24 Aug. 21 295 (32) 0 6 (1) 2 123 (15) 30-31 Aug. 40 275 (18) 3 34 (5) 25 220(16)
From the beginning of August weekly counts were made of insect numbers in 200 cones from each garden (Table 2). In every garden aphid numbers increased slowly until harvest, but in none of the gardens was the level of aphids of concern to the growers although at Boughton small growths of sooty moulds were noted in nine of the cones. Small numbers of anthocorids, particularly Orius majusculus (Reut.) and 0. laevigatus Fieb., were found but their numbers did not increase in response to aphids.
Aphid migration In 1974 aphid migration started on 14 May, compared with 4 June in 1975 and 22 May in
1976. The highest weekly counts for each year in the 12.2 m suction trap at Wye were 602, 171 and 274 in 1974, 1975 and 1976 respectively. In 1976 alates were not recorded in the suction trap after 5 July, which was at least a month earlier than the last records in 1974; in 1975 small numbers of alates were recorded throughout August until the start of the return migration (Aveling, 1977). The late start to migration in 1975 followed by a low influx, compared with other years, was possibly because of cold weather in April which may have killed early spring aphid colonies on Prunus spp.
A nthocorid populations Since A . nemoralis and A . confusus were difficult to separate in the field, in 1974 they were
recorded as A. nemoralis. In 1975 only adults were distinguished, while in 1976 both adults and larvae were distinguished.
In all seasons anthocorid bugs began to migrate to the hops 3 to 4 wk after the first aphids

Anthocorids in integrated control of hop aphid 149
were recorded. This highlighted the need for chemical control of aphids during early June in order to prevent damaging numbers of aphids from developing before the anthocorid populations could become established in late-June and early-July.
A . nemoralis was always the most abundant species and each season A. confusus and A . nemoralis were the early arrivals, with A . nemorum beginning to appear up to 2 wk later. In 1975 and 1976 A. confusus adults were found up to a week before A . nemoralis. In those trials in which aphid numbers built up during June and early July larger populations of larval A. nemoralis also developed. In 1974 and 1975 a rapid aphid population decline occurred during the first week of August while in 1976 the decline occurred 3 wk earlier; in each instance this decline coincided with the peak numbers of the most voracious anthocorid stages (i.e. fourth and fifth instar larvae and adults), suggesting that the anthocorids caused the aphid declines (Fig. 3).
n A. nemoralis t A. confusus 2oo F loo t
- 5 25 c A . nemorum 1 2 3 4 5 A 1 2 3 4 5 A l 2 3 4 5 A
c 5 400 [ A . nemoralis + A. conjiusus c
300 - c-
g 200 - 6 z
100 -
l 2 3 4 5 A - 1 2 3 4 5 A
23 b - 1 2 3 4 5 A
1 2oo t
[ A . nemorum 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A
1 July 8 July 14 July 22 July 29 July 5 August 12 August
- Fig. 4. Changing population structure of anthocorids on mephosfolan-treated and untreated ‘banker’ plants at Wye, 1975. Numbers of each instar (1-5) and adults (A) per string for each sampling date. Blacked sections represent A . confusus adult numbers only.
The changing population structures of the anthocorids in 1975 at Wye and 1976 at Harefield and Boughton, are shown in Figs 4 and 5 . A . nemorum was never numerous because, being a later arrival, the development of its larval population coincided with the aphid population decline. Severe competition for aphid prey took place as aphid numbers dropped; also older A . nemoralis were often seen feeding on young A . nemorum as well as on other beneficial insects such as syrphid larvae and aphid ‘mummies’ containing developing parasites. Inter- and intra-specific competition became so intense after the aphid population decline that many anthocorids either died or emigrated. In all seasons A . nemorum adults were the most common anthocorid on the hop picking machines at harvest.
In 1975 there were two clear peaks, on 22 July and 5 August, in numbers of adult A . nemoralis on both treated and ‘banker’ plants (Fig. 4). There was also a peak in adult A . confusus numbers on 5 August on the ‘banker’ plants only, although no obvious peak in the early part of July. Clearly for both treatments the peak on 5 August comprised adults which, being the progeny of early migrants, had spend their entire larval life of about 4 wk on the hops, while the

150 C . A V E L I N G
80 60
40 40 20 20
M Y
1 lo [ A.confusus c
1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A
~ ~ ~ n e m ~ d , , - Ash - - 1 6 0
40 20 20
8 BOUGHTON 80
- 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A 1 2 3 4 5 A
22June 29June 6July 13 July 20 July 27 July 3 August
Fig. 5. Changing population structure of anthocorids at Harefield and Boughton, 1976. Numbers of each stage per string for each sampling date.
22 July peak probably comprised migrating A . nemoralis adults only. Similarly in 1976 at Harefield, Boughton and Horsmonden, A . nemoralis was able to complete a generation on the hops. Fig. 5 shows an early peak of migrating A . nemoralis adults on 29 June at Harefield and Boughton followed 2 wk later by a second peak comprising recently matured adults. A . nemorum produced only a single peak of migrating adults but because they were late arrivals few second generation adults developed.
Although the ‘banker’ plants in 1975 were a potential source of large numbers of anthocorids, there was no clear evidence that they contributed significantly to P . humuli control in the surrounding plants. With the rapid increase in the number of adult anthocorids on the ‘banker’ plants between 29 July and 5 August it can be estimated that on 5 August there were approximately 3200 anthocorids (100 adults/string from 32 ‘banker’ plant strings) which were potentially capable of dispersing into the surrounding plants. However, aphid numbers were already falling on the treated plants during the previous week, so that the aphids might still have been controlled in the absence of ‘banker’ plants. Nevertheless the level of anthocorids within the plot may have been higher and more prolonged than in the absence of the ‘banker’ plants. This would be particularly useful in the event of a prolonged aphid migration and would ensure the presence of a few anthocorids in the cones where aphid control is critical.
Predator exclusion cage experiments Experiments with cages to exclude predators provided conclusive evidence of the controlling
influence of predators. The cones in the exclusion cages in 1975 (Table 3) and 1976 (Table 4) became increasingly heavily infected by aphids with considerable growth of sooty moulds in the

Anthocorids in integrated control of hop aphid 151
Table 3. Numbers of Phorodon humuli in 80 cones from exclusion cages and untreated controls in 1975 (10 coneslstring). Controls comprised four open-ended cages and four
uncaged strings. Cagesjitted on 4 August
Exclusion cages Open-ended cages Uncaged strings & A
Dates Adults Nymphs Adults Nymphs Adults Nymphs 12 Aug. 41 85 23 78 15 52 19 Aug. 87 608 0 3 0 6
- 26 Aug. 181 1652 0 8 0 1 2 Sept. 870 6193 0 27 0 12
Table 4. Numbers of Phorodon humuli in 80 cones from eight exclusion cages and eighl uncaged control strings in 1976 (10 coneslstring). Cagesjitted on 13 July
Exclusion cages Uncaged strings
& Adults Nymphs
& Dates Adults Nymphs
5 Aug. 5 3 829 1 2 13 Aug. 200 2359 0 4 20 Aug. 668 9654 2 19 27 Aug. 29 10 - 6 5 5
2 Sept. > 10 000 - 18 47
honeydew and stunting of plant growth. In contrast aphid numbers in cones from open cages and on uncaged plants in 1975 dropped to a low level by 19 August and the cones remained clean until harvest. In 1976, although aphid numbers on uncaged bines increased slowly, their numbers remained low. Thus on the last sampling date, there were 18 adult apterae in 80 cones compared with over 10 000 from the exclusion cage cones.
Other predators Other predators found feeding on the aphids in all seasons included Syrphus ribesii L., S .
balteatus DeG., Adalia bipunctata (L), Coccinella undecimpunctata L., C. septempunctata L., Propylea quatuordecimpunctata L., and Chrysopa carnea Stephens. However in all years their numbers were low in comparison to anthocorid numbers.
In 1975 large numbers of C. undecimpunctata were recorded in south-east England (Owen, 1976) during the first week of August, but their contribution towards P. humuli control was probably insignificant because they arrived too late and did not lay eggs.
Survey of non-crop vegetation Since areas of uncultivated land are important sources of beneficial insects, the vegetation
around hop gardens at Wye was examined each year in early spring. Many female anthocorids, particularly A . nemorum, were found overwintering under the bark of sycamore trees (Acer pseudoplatanus L.). They were later found feeding among the early aphid colonies such as the sycamore aphid, Drepanosiphum platanoides (Schr.), and the nettle aphid, ~ i c r o l o p h u m evansi (Theobald). Each spring a large larval generation developed, particularly on nettles, and first-generation adults appeared during late June. During July the adults migrated to a wide range of summer host plants, including nearby hops. However, many nettle patches examined adjacent to other hop gardens were not infested with aphids and contained no anthocorids. Furthermore although A . nemoralis was the most numerous anthocorid on hops in all seasons,

152 C . A V E L I N G
overwintered adults or first generation larvae were rarely found on the nettles or on beech and hawthorn hedges (cf: Anderson, 1962) which surrounded most of the hop gardens.
D I S C U S S I O N A N D C O N C L U S I O N S
Predation by anthocorids On all sites studied, anthocorids were more abundant than other predatory insects. Three
species of these, particularly A . nemoralis, were mainly responsible for the rapid decline in aphid numbers in July/August each year. This decline always coincided with the peak numbers of fourth and fifth instar larvae and adult anthocorids, i.e. the most voracious life stages. It is crucial to control aphids whilst the hop cones are forming, and anthocorids were the only predators found in the cones. When excluded, as in the cage experiments, aphid infestation of the cones was severe.
In the 1976 trials, anthocorids controlled the aphids well in advance of cone formation and their numbers then declined markedly. As a result a few aphids entered the cones and developed to an appreciable infestation, although not at a level to cause concern to the growers. In contrast, on other farms where contact insecticides toxic to predators were sprayed, aphid infestation in the cones was severe. Ideally, therefore, control of the aphids by natural enemies should not occur too far in advance of cone formation; it is important that anthocorids are fairly abundant at this growth stage.
There was little evidence that A . nemoralis -the most numerous anthocorid on hops in all 3 yr - came mainly from vegetation adjacent to hop gardens, e.g. nettles, beech and hawthorn hedges. Both A . nemorum and A . nemoralis show considerable flight activity (Southwood, 1960) and can probably disperse over considerable distances. Nevertheless, where hops are grown intensively the role of non-crop habitats as sources of anthocorids needs further investigation.
Chemical control Mephosfolan soil drenches. Uptake of mephosfolan from the soil is good when the drench is
applied to young, rapidly-growing plants immediately before or after rainfall. Dry weather occurred during June in all 3 yr and this apparently restricted the uptake of mephosfolan by some bines, resulting in a variable proportion of ‘missed bines’ which became heavily infested. The latter can be useful in integrated control as they become ‘banker’ plants, i.e. sources of adult anthocorids for surrounding plants in early August when the cones are forming.
Nevertheless, means of ensuring more uniform uptake of mephosfolan - possibly by trickle irrigation - are needed so that the proportion of ‘banker’ plants can be closely controlled. Since totally untreated bines are defoliated by aphids in most seasons, it will probably be necessary to treat potential ‘banker’ plants with a reduced rate of mephosfolan, perhaps a quarter dosage. Further work is needed to determine what proportion of the bines as ‘banker’ plants is effective and acceptable to growers, possible about 1%. These plants pose no threat of aphid infestation to the rest of the garden since P. humuli does not produce vagrants.
Foliar sprays of contact insecticides. The trial at Wye in 1976 showed that when a foliar spray of a contact insecticide was used before the end of June few predators were present and those arriving later were able to multiply and control the aphids. Clearly it is not desirable to eliminate the aphids in July whilst anthocorids are establishing natural control. Thus, means are needed of obtaining partial control by insecticides of low toxicity to anthocorids. Work on control of P. humuli with juvenile hormone analogues is of interest (Hrdy & Zelany, 1973), but the effects of these compounds on beneficial insects need further evaluation.
Economic injury levels and monitoring methods Low numbers of P. humuli must be present on hops in late June and July to allow anthocorids
to establish. Therefore an important area for future research is the determination of economic

Anthocorids in integrated control of hop aphid 153
injury levels, which in respect of the effect of aphid damage on yield will probably differ according to whether attack occurs in June, July or August (the last month involves infestations in the cones). Another aspect concerns the possible effect on the content of alpha-acids in the cones; these acids are important to the brewing industry and influence the market value of hops. However, Aveling (1977) showed that alpha-acid content was unlikely to be affected by the numbers of aphids necessary to attract and establish anthocorids.
More work is also needed to develop monitoring methods for the aphid and for anthocorids which are fairly quick and reliable and suitable for use by field advisors and growers. Methods for estimating aphid infestation might be based on the proportion of ‘missed bines’. However a difficulty will be to decide what constitutes a ‘missed bine’ as for example in the present study when a number of plants were heavily infested only in the top of the bine. Monitoring of anthocorids will require being able to recognise young larvae (first to third instar), old larvae (fourth and fifth instar), and adults, and sampling should be confined to ‘missed bines’ where large anthocorid populations would be expected to develop. Growers should anticipate good control by predators if young larvae predominate in the anthocorid population no less than 3-4 wk before expected cone formation, and large anthocorid larvae predominate no less than one to two weeks before cone formation. When sampling for anthocorids it would be important to check not only the leaves, but also between the bine and string. Once the different anthocorid stages can be recognised this sampling method should be easy to carry out.
I thank my supervisor Dr D. S . Madge and Dr R. A. Neve for their interest and advice. A Research Scholarship was provided by Arthur Guinness, Son & Co. Ltd, to whom I am grateful.
R E F E R E N C E S
ANDERSON, N. H. ( 1 9 6 2 ) . Bionomics of six species of Anthocoris (Heteroptera: Anthocoridae) in England. Transactions of the Royal Entomological Society of London 1 1 4 , 67-95.
AVELING, c. ( 1 9 7 7 ) . The Biology of Anthocorids (Heteroptera: Anthocoridae) and their role in the integrated control of the damson-hop aphid (Phorodon humuli (Schrank)). Ph.D. Thesis, University of London.
BUXTON, J. H. ( 1 9 7 4 ) . The biology of the European earwig, ForJicula auricularia (L.), with reference to its predatory activities on the damson-hop aphid, Phorodon humuli (Schrank). Ph.D. Thesis, University of London.
CAMPBELL, c . A. M. ( 1 9 7 8 ) . Regulation of the damson-hop aphid (Phorodon humuli (Schrank)) on hops (Humulus lupulus L.) by predators. Journal of Horticultural Science 53, (3) 235-242.
H R D ~ , I. ( 1 9 7 0 ) . Hop aphid outbreak and resistance. Chmelarstui 43, 1 6 6 - 1 7 7 and 1 8 1 - 1 8 3 . H R D ~ I. & Z E L A N ~ , J. ( 1 9 7 3 ) . Effects of juvenoids on the population density of Phorodon humuli in a cage
experiment (Homoptera: Aphididae). Acta Entomologica Bohemoslouaca 70,386-389. MUIR, R. C. ( 1 9 7 9 ) . Insecticide resistance in damson-hop aphid Phorodon humuli in commercial hop
gardens in Kent. Annals of Applied Biology 92, 1-9. OWEN, D. (1976). Ladybird, ladybird, fly away home. New Scientist 7 1 , 6 8 6 - 6 8 7 . SANDS, W. A. ( 1 9 5 7 ) . The immature stages of some British Anthocoridae (Hemiptera). Transactions of
SMALLMAN, B. N. ( 1 9 6 5 ) . Integrated insect control. Australian Journal of Science 28,230-234. SOUTHWOOD, T. R. E. ( 1 9 6 0 ) . The flight activity of Heteroptera. Transactions of the Royal Entomological
ZOHREN, E. ( 1 970). Moglichkeiten einer integrierten Bekampfung von Hopfenschadlingen. Zeitschrft f i r
the Royal Entomological Society of London 1 0 9 , 2 9 5 - 3 10.
Society of London 112, 1 7 3 - 2 2 0 .
Angewandte Entomologie 6 5 , 4 1 2 - 4 1 9 .
(Received I3 August 1979)