the roles of zinc and metallothionein in …
TRANSCRIPT

THE ROLES OF ZINC AND METALLOTHIONEIN IN HEPATOCYTE
ANTIOXIDANT DEFENSE
By
LOUIS LICHTEN
A THESIS PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE
UNIVERSITY OF FLORIDA

Copyright 2004
by
Louis Lichten

This thesis is dedicated to my father, and my grandparents. Without their love and support, this would never have been possible.

ACKNOWLEDGMENTS
I would like to thank Kelly Clayton for always encouraging me to do my best. I
would also like to thank my lab mates for all their help. I thank Ray Blanchard for his
wisdom in answering all my questions, and Jeff Bobo for always being willing to help me
with my experiments. I thank Cal Green for his help with solving experimental
problems. I would also like to thank Juan Liuzzi for helping when I needed it. I greatly
appreciate Steve Davis’s help from the very beginning of this project. I would also like
to thank Dr. Gregory, and Dr. Kilberg for their help and participation on my thesis
committee. Finally, I would like to thank Dr. Cousins for giving me the opportunity to
work on this project.
iv

TABLE OF CONTENTS page ACKNOWLEDGMENTS ................................................................................................. iv
LIST OF TABLES............................................................................................................ vii
LIST OF FIGURES ......................................................................................................... viii
ABSTRACT....................................................................................................................... ix
CHAPTER 1 INTRODUCTION ........................................................................................................1
2 LITERATURE REVIEW .............................................................................................4
Zinc ...............................................................................................................................4 Interrelations of Zinc and Metallothionein ...................................................................4 Zinc and Metallothionein as Cellular Antioxidants......................................................6 Metallothionein, Nitric Oxide, and Oxidative Stress....................................................7
3 MATERIALS AND METHODS ...............................................................................10
Animals.......................................................................................................................10 Hepatocyte Isolation ...................................................................................................10 Culture Period.............................................................................................................11 Treatment Period. .......................................................................................................12 Cytotoxicity Assays. ...................................................................................................12 Data Analysis..............................................................................................................15
4 RESULTS...................................................................................................................16
Lactate Dehydrogenase Assay....................................................................................16 Metallothionein Genotype Differences.......................................................................17 Zinc Supplementation.................................................................................................19 Altered Genotype and Zinc Supplementation.............................................................20
5 DISCUSSION.............................................................................................................27
APPENDIX CYTOTOXICITY EXPERIMENTS ...........................................................32
v

LIST OF REFERENCES...................................................................................................33
BIOGRAPHICAL SKETCH .............................................................................................37
vi

LIST OF TABLES
Table page 4-1 LD50 responses due to altered metallothionein genotype. ......................................19
4-2 LD50 responses to toxicants of hepatocytes from MT KO, CK, CT, and MT Tg mice as a result of zinc status.......................................................................24
4-3 Summary of LD50 responses between the metallothionein genotypes after pretreatment by zinc.........................................................................................26
vii

LIST OF FIGURES
Figure page 4-1 Influence of SNAP on LDH activity.. ......................................................................16
4-2 Viability due to altered metallothionein genotype.. .................................................18
4-3 Effect of zinc status on subsequent toxicant induced viability changes in control knockout and metallothionein knockout hepatocyes.. .................................22
4-4 Effect of zinc status on subsequent toxicant induced viability changes in control transgenic and metallothionein transgenic hepatocyes.. ..............................23
4-5 Differences in hepatocyte viability due to altered MT genotypes and zinc pretreatment..............................................................................................................25
A-1 Differences in CT and Tg viability due to altered MT genotypes and zinc pretreatment..............................................................................................................32
viii

Abstract of Thesis Presented to the Graduate School
of the University of Florida in Partial Fulfillment of the Requirements for the Degree of Master of Science
THE ROLES OF ZINC AND METALLOTHIONEIN IN HEPATOCYTE ANTIOXIDANT DEFENSE
By
Louis Lichten
August 2004
Chair: Robert Cousins Major Department: Food Science and Human Nutrition
Zinc and metallothionein are involved in cellular antioxidant defense, but the
basic mechanisms are not known. Zinc is a biologically redox inert metal, that has been
associated with protection from various oxidant producing species. However, zinc
induces the cysteine-rich, metal-binding protein, metallothionein, through interaction
with the transcription factor MTF-1. Metallothionein may have a role in preventing
oxidative stress, by quenching free radicals and other oxidants. Therefore, it is hard to
distinguish between protection due to zinc, and protection due to induction of
metallothionein.
Hence, metallothionein null (KO) mice and metallothionein transgenic (Tg) mice
provide a unique opportunity to study altered states of metallothionein expression. These
mouse models were used to determine the effects of zinc supplementation and/ or
metallothionein gene expression on the susceptibility to oxidative and nitrosative stress.
Metallothionein KO mice were significantly less viable than control (CK) mice after
ix

tertiary-butyl hydroperoxide (tBH) and s-nitroso n-acetyl penicillamine (SNAP)
treatments. Zinc supplementation (40 µM) had no effect on viability of KO hepatocytes.
The presence of metallothionein in CK hepatocytes demonstrated protection at
baseline and after zinc supplementation. However, overexpression of metallothionein
provided no further protection against tBH or SNAP. These results are in contrast to a
direct antioxidant role for metallothionein in murine hepatocytes.
x

CHAPTER 1 INTRODUCTION
Zinc is an essential mineral nutrient, possessing various biological functions
including catalytic and structural roles in metalloenzymes and metalloproteins, as well as
regulatory roles in synaptic signaling and gene expression (Cousins 1996). If zinc is
absent from a biological system, an organism becomes more susceptible to oxidative
insult. Zinc-deprived animals have demonstrated either increased free-radical production
or enhanced injury from exposure to oxidative stress (reviewed in Powell 2000). The
antioxidant mechanism of zinc protection is uncertain, but may relate to (1) zinc as a
structural factor for copper-zinc superoxide dismutase, (2) the ability of zinc to protect
sulfhydryl groups from oxidation, and (3) the ability of zinc to antagonize the catalytic
properties of redox-active transition metals (reviewed in Powell 2000). A final possible
antioxidant mechanism may be the ability of zinc to induce metallothionein (MT).
Metallothionein is a cysteine-rich protein induced by zinc through the metal
responsive transcription factor-1 (MTF-1), and interaction with metal response elements
in the metallothionein promoter (Andrews 2000). Interestingly, metallothionein is also
induced by oxidants through antioxidant response elements and/ or MTF-1 (Andrews
2000). At the protein level, metallothionein may be protective against oxidants by
scavenging free radicals. This antioxidant potential was first realized by the quenching of
the hydroxyl radical by metallothionein (Thornalley and Vasak 1985). Further, many
studies have shown metallothionein to be protective against a variety of oxidative insults:
radiation exposure (Matsubara 1987), anticancer drugs (Satoh et al. 1988), carbon
1

2
tetrachloride (Schroeder and Cousins, 1991), S-nitroso N-acetyl penicillamine (SNAP)
(Schwartz et al. 1995), peroxynitrite (Cai et al. 2000), and ethanol toxicity (Zhou et al.
2002). The mechanism of protection by metallothionein may be the result of interaction
of an oxidant with the redox-active zinc-sulfur ligands of metallothionein, causing
reduction of the oxidant and subsequent release of zinc (Jacob et al 1998). This
mechanism allows metallothionein to be considered as a possible link between cellular
zinc and the redox state of the cell (reviewed in Maret 2000).
To look at the efficacy of metallothionein protection, metallothionein gene
knock-out (KO), and metallothionein transgenic (TG) mice can be used. In a study by
Davis et al. (2001), expression of metallothionein protected against carbon
tetrachloride-induced hepatotoxicity, but metallothionein overexpression did not provide
further protection. These results are inconsistent with direct antioxidant activity of
metallothionein. Therefore, experiments were designed to more directly assess the
antioxidant activity of metallothionein by treatment of primary hepatocyte cultures (from
these genotypes) with tertiary-butyl hydroperoxide (tBH), and a nitric oxide donor
(SNAP). Current studies utilizing the same study design and a redox-insensitive method
to detect cell viability demonstrate the protection provided by metallothionein against
oxidative and nitrosative stress.
• Hypothesis 1: Basal increase of metallothionein expression in the transgenic mice will provide isolated hepatocytes increased protection from nitric oxide and/or tBH, while the ablation of metallothionein in the knock-out hepatocytes will be deleterious.
• Hypothesis 2: Supplementation of hepatocyte cultures with zinc prior to nitric
oxide or tBH exposure will further enhance the viability of transgenic cells, through induction of metallothionein.

3
The specific aim of these experiments is to determine whether the presence of
metallothioenein will protect primary hepatocytes from tBH or nitric oxide-induced
oxidative stress, as measured by cell viability.

CHAPTER 2 LITERATURE REVIEW
Zinc
Zinc is an essential mineral nutrient that is required for many biological roles
including catalytic, structural, and regulatory roles (Cousins 1996). Zinc functions
catalytically in more than 60 enzymes, such as metallopeptidases (Turner et al. 1996).
Metalloenzymes are considered to be zinc-dependent when removal of zinc causes
reduction in activity, and replacement of zinc restores activity. Structurally, greater than
1% of the genome codes for proteins containing zinc finger or similar domains, where
zinc serves a structural role (Chesters 1992). Typically, zinc acts to stabilize the tertiary
structure of metalloproteins (Vallee et al. 1991). Zinc is typically coordinated by four
amino acid side chains: usually the sulfur atom of cysteine, or sometimes histidine. The
regulatory role of zinc includes the binding of the metal to specific transcription factors,
allowing binding of the transcription factor to metal responsive elements in the promoters
of metal responsive genes (reviewed in Andrews 2000). Zinc binding may also have a
regulatory role, through activation of signal transduction pathways (reviewed in Maret
2000).
Interrelations of Zinc and Metallothionein
In vivo, zinc metabolism is controlled homeostatically, through mechanisms
related to absorption and excretion (via zinc transporters), and intracellular proteins such
as metallothionein. Metallothioneins are small (6-7 kDa), cysteine-rich
heavy-metal-binding proteins, and can bind up to seven zinc atoms (reviewed in Cousins
4

5
1996). Approximately 5 to 10% of the total cellular zinc is found in complex with
metallothionein under normal physiological conditions (Andrews 2001).
Metallothioneins are the most abundant heavy-metal-binding proteins in the body
(reviewed in Andrews 2000). Metallothioneins are thought to function in the
homeostasis of zinc through involvement in zinc absorption, tissue distribution, and
protection against acute stress (Schroeder and Cousins 1991b). During periods of acute
stress, there is a decrease in plasma zinc levels, along with an increase in tissue zinc,
particularly in the liver (reviewed in Cousins 1996). This reduction in plasma zinc is
directly related to the changes in kinetics of zinc metabolism, which lead to increased
binding of the metal to metallothionein (Dunn and Cousins 1991). In support of a link
among zinc, metallothionein, and regulation of zinc metabolism, results of experiments
with endotoxin-challenged metallothionein null mice (MT KO) show no decrease in
plasma zinc (Philcox et al. 1995). Therefore, metallothionein plays a crucial role in
plasma-zinc clearance and tissue-zinc uptake.
Metallothionein KO mice lack functional expression of metallothionein, and
allow zinc metabolism to be studied in the absence of metallothionein (Davis et al. 1998).
Transgenic mice have been developed that have approximately 55 additional copies of
the metallothionein-I gene in their genome (Palmiter et al. 1993). Metallothionein
transgenic mice provide a model for studying the effect of exaggerated metallothionein
levels on zinc metabolism.
Metallothionein is transcriptionally regulated by zinc, through metal responsive
elements in the promoter region of the gene (Andrews 2000). However, the
metallothionein gene is also transcriptionally regulated by glucocorticoid hormones and

6
specific cytokines. This allows for the possibility that these metalloproteins plays a role
in inflammatory and stress-related responses (reviewed in Davis and Cousins 2000).
Zinc and Metallothionein as Cellular Antioxidants
The oxidation of many different cellular constituents is involved in the
pathogenesis of an array of diseases (Gutteridge and Halliwell 2000). Cellular oxidative
stress occurs when the antioxidant defense system in the cell becomes overwhelmed.
There are many different forms of oxidants, including reactive oxygen species (H2O2, O2-,
and OH), reactive nitrogen species (NO and ONOO-), and carbon centered radicals
(e.g., .CCl3). These oxidative species can damage lipids, proteins, and nucleic acids
(Farber 1994). When oxidative stressors are introduced into cells, specific cellular
antioxidant defense mechanisms are present to quench or reduce the radical mediated
damage that may occur. These defense mechanisms involve antioxidant nutrients such as
tocopherols and ascorbate, as well as the cellular antioxidants glutathione and superoxide
dismutase (Yu 1994). If damage does occur, there are ways for the cell to repair itself.
For instance, oxidized bases are removed from DNA to prevent further damage, oxidized
lipid membranes are reduced by the action of glutathione peroxidase, and damaged
proteins are committed to a proteasome degradation pathway. If these cellular defenses
are overcome, and oxidative stress occurs in great amounts, the cell or cells may not
survive. Therefore, establishing a way to reduce oxidative stress is important.
Supplemental zinc provides additional protection against certain radicals (Blain et
al. 1998). Zinc is a potent inducer of metallothionein expression. Consequently, the
antioxidant protection attributed to zinc may be due to metallothionein induction.
Metallothionein expression is also induced in response to oxidative stress (reviewed in

7
Davis and Cousins 2000). Therefore, there is a great degree of correlation between
increased metallothionein synthesis and protection from oxidative stress. However, this
protection is not always observed. In studies using metallothionein knock out and
metallothionein transgenic mice, carbon tetrachloride induced hepatotoxicity was reduced
initially by the presence of metallothionein in control compared to knock out mice (Davis
et al. 2001). Later time points showed no difference between genotypes in response to
carbon tetrachloride. Also, metallothionein transgenic and transgenic control mice did
not differ in the levels of hepatotoxicity, despite a large degree of difference in hepatic
metallothionein and zinc content. Further examination of zinc and metallothionein as
cellular antioxidants in cells from this mouse model led to differing results. In two
independent studies, overexpresssion of metallothionein was found to be ineffective at
increasing viability, and may actually be deleterious (Davis et al. unpublished data).
These results are a remarkable contrast from the apparent protective effects of
metallothionein observed with carbon tetrachloride treated rat hepatocytes (Schroeder
and Cousins, 1990), and numerous other reports describing a protective effect of this
protein.
While many experiments have shown metallothionein and zinc to have
cytoprotective roles, some have not. This disparity in protection must be investigated
further utilizing a different oxidative stressor, such as nitric oxide.
Metallothionein, Nitric Oxide, and Oxidative Stress
During hepatic inflammation or endotoxemia, hepatocytes can respond to
cytokine or bacterial LPS stimulation by activating the inducible form of nitric oxide
synthase, which can generate a large amount of nitric oxide from arginine (Nüssler et al.
1993). This production of NO has been shown to be both hepatoprotective and cytotoxic

8
(Wink and Mitchell 1998). The paradoxical effects of nitric oxide may relate to its ability
to interact with O2- to form peroxynitrite, which is highly reactive and may account for
part of nitric oxide-induced cytotoxicity (Fu et al. 2001). The susceptibility of cells to
peroxynitrite and nitric oxide is primarily dependent upon their reducing capacity (thiol
content) (Kim et al. 1999). Metallothionein has been shown, in vitro, to react directly
with peroxynitrite to prevent lipoprotein and DNA damage caused by this reactive
nitrogen species (Cai et al. 2000). However, the protection by metallothionein in this
study was not dose dependent. Increasing levels of the protein did not provide further
protection against DNA damage, or LDL oxidation. Nonetheless, metallothionein over
expression has been documented to be protective against SNAP (NO-donor) - induced
killing of, and DNA single-strand breaks in NIH 3T3 cells (Schwarz et al. 1995).
Further in vitro studies have elucidated a link between the cellular redox state and
metal ion homeostasis (Jacob et al. 1998, reviewed in Maret 2000). The metal thiolate
clusters of metallothionein possess the unique ability to function as a redox unit, therefore
it has the potential to be involved in a redox sensitive signaling pathway (Pearce et al.
2000). Indeed, nitric oxide has been shown to S-nitrosylate metallothionein and release
zinc from cultured pulmonary artery endothelial cells as confirmed by fluorescence
protein modified metallothionein that undergoes conformational changes in the presence
of nitric oxide (Pearce et al., 2000). These alterations in cellular zinc homeostasis may
lead to the protective effects of metallothionein against nitric oxide toxicity (Schwarz et
al. 1995).
Overall, the data on the protective effects provided by zinc and metallothionein
against oxidative stress is not definitive. Therefore, further characterization of the

9
interaction of zinc and/or metallothionein to oxidative insult, is necessary. The
pathogenesis of many liver diseases may begin with oxidative injury (Gutteridge and
Halliwell, 2000). For that reason, hepatocytes provide a good model to understand the
possible role of zinc as an antioxidant. Consequently, the goal of this study is to
elucidate the role of zinc and metallothionein as hepatic cytoprotective agents, utilizing
the murine model of altered metallothionein gene expression (i.e. MT-KO and MT-TG
mice).

CHAPTER 3 MATERIALS AND METHODS
Animals
All breeding of MT-KO and MT-Tg mice was done in-house from founder mice
purchased from the Jackson Laboratory, Bar Harbor, ME. The C57BL/6 mice were
crossed with SJL mice, possessing multiple copies of the murine metallothionein-1 gene,
to create the line of metallothionein overexpressing mice (Palmiter et al. 1993). The
control transgenic mice were created by backcrossing against the C57BL/6 strain. The
MT-KO genotype was developed by crossing 129/SvCPJ mice MT (-/-), with C57BL/6
mice (Masters 1997). The control KO mice are 129S3/SvImJ mice. Six- to eight-week
old male mice were used in all experiments. Mice were fed a modified commercial
rodent diet (Harland TEKLAD, TEKLAD Rodent Diet (W) 8604) and given tap water.
Hepatocyte Isolation
Liver perfusions were alternated among all four genotypes. Mice were anesthetized
with an intraperitoneal injection of sodium pentobarbital (60 mg/kg). The isolation of
liver cells involves many steps. First, the liver was infused with a calcium-free solution
containing 0.142 M NaCl, 0.0067 M KCl, and 0.01 M HEPES buffer (pH 7.3), via the
subhepatic inferior vena cava (modification of Schroeder and Cousins 1991a). Initial
flow rate for the calcium-free solution was set at 2 mL/min. As the liver began to
blanche, an incision was made in the portal vein, allowing perfusate to freely flow
without recirculation. After opening the rib cage, a hemostat was then placed on the
inferior vena cava superior to the diaphragm. The flow rate of calcium-free solution was
10

11
then increased to 8 mL/min for approximately 10-12 min. The subsequent step involved
perfusion of a solution containing 0.0667 M NaCl, 0.0067 M KCl, 0.00476 M CaCl2,
0.10 M HEPES buffer (pH 7.3), and 0.04% collagenase (Sigma type IV collagenase, C-
5138) at a flow rate of 7 mL/min for 14 min. Upon completion of digestion, the liver was
excised rapidly from the body cavity, and transferred to a sterile glass culture dish
containing approximately 15 mL of the collagenase solution. The culture dish was then
transferred to a sterile cell culture hood.
Prior to liberation of hepatocytes, the gall bladder was carefully removed.
Incisions were then made in each of the liver lobes to allow easy access to the
hepatocytes. Finally, using a cell scraper, cells were released from the liver lobes gently,
and as quickly as possible.
Culture Period
After liberation, the liver cells were passed through a 100 um cell strainer to
remove debris and large cell aggregates. The hepatocytes were then suspended in a
buffered wash medium (William’s Medium E + 10 mM HEPES pH 7.3), and collected by
centrifugation at approximately 50 x g for 4 min. Cells were then washed in the same
buffer two more times to improve cell purity. The final cell pellet was resuspended in
attachment medium (WME supplemented with 10% FBS, 100 nM insulin, 100 nM
Dexamethasone, 100 U/mL penicillin, 100 mg/mL streptomycin). An aliquot of the final
cell suspension was then removed, and placed into solution with Trypan Blue (Sigma
T8154) and PBS for determination of cell viability. Viability of the hepatocytes was then
assessed by Trypan Blue Exclusion as measured with a hemacytometer. Only
suspensions with > 90% viability were used in experiments. If viability was satisfactory,
cells were seeded at 2.5 x 105 cells/ well in 12-well, collagen coated culture plates (6-10

12
µg collagen/ well). After plating, the cells are allowed to attach for 3 h (37°C, 5% CO2).
Following selective attachment of parenchymal cells, medium in each well was
exchanged for fresh medium only, or fresh medium containing 40 µM zinc. These
culture conditions did not change for 18-22 h, when oxidant treatment began.
Treatment Period
Upon completion of the culture period, all media was removed and replaced with
medium containing 0-3 mM SNAP for 2 to 14 h, or 0-75 µM tBH for 2 h.
Cytotoxicity Assays
After the treatment period had ended, medium from each well was removed and set
aside. Cell monolayers were then washed twice with PBS, and cells were harvested in
1% Triton X-100 and disrupted by repeated passage through a 200 µL pipette tip. The
medium as well as cell extracts were then centrifuged (130 00 x g for 20s) to remove
cellular debris, stored at -20◦C for up to 48 h prior to enzyme analysis.
Lactate dehydrogenase activity was measured spectrophotometrically, for cell
viability determination, by three different methods. The first method involved incubating
aliquots of medium or cell extracts with a solution containing phosphate buffer (0.1 M,
pH 7.4), sodium pyruvate (9.1 mM), and β-NADH (3.3 mM). LDH in the sample
catalyzes the reaction of pyruvate to lactate, resulting in oxidation of NADH to NAD.
Absorbance at 340 nm is used to measure the rate of disappearance of NADH, and is
directly proportional to LDH in sample (Henry et al. 1960). The second method utilized
the lactate dehydrogenase (LD-L) assay kit (Sigma procedure 228-UV). This assay
procedure combined the same method of medium acquisition and cell harvesting as
above. However, the extracts were then incubated in a lactate dehydrogenase reagent
(50mM lactate, 7 mM NAD, 0.05% NaN3, and buffer, pH 8.9), and the reduction rate of

13
NAD was read immediately spectrophotometrically at 340 nm. The final method used to
analyze LDH activity was the Sigma TOX-7 assay. This assay involved reduction of
NAD to NADH, which was then used in a second coupled stoichiometric conversion of a
tetrazolium dye to a colored compound. The production of the colored compound was
then assessed spectrophotometrically at 590 nm. In all three methods a calculation of the
cell viability via percent LDH activity was made, by determining the LDH activity of
medium, and LDH activity in the cell extract (medium activity/ (medium activity + cell
activity)) (Jauregui et al. 1981).
Cell viability was also determined using cell proliferation reagent WST-1 (Roche
Cat. No. 1 644 807). The assay is based on the cleavage of the tetrazolium salt WST-1
by mitochondrial dehydrogenase through production of NADH in viable cells. Briefly,
hepatocytes were cultured in microtiter plates (tissue culture grade, 96 well, flat bottom
plate). After the treatment period, each well was brought to a final volume of 100 µl in a
humidified atmosphere (37◦C and 5% CO2). The WST-1 reagent was then added
(10µL/well). The plate was then incubated for 2 h in a humidified atmosphere (37◦C and
5% CO2). Before assaying, the plate was placed on a shaker for 5 min to ensure thorough
mixing. The absorbance at 450 nm was then measured spectrophotometrically with 690
nm as the reference wavelength. Culture medium served as the blank. Absorbance of
samples directly correlates to the viability of the cells.
The crystal violet assay was also used to determine cytotoxicity (Kim et al 1999).
Upon conclusion of the treatment period, medium was removed from each well, and the
cells were washed several times with PBS. The PBS was then removed and subsequently
replaced with a dye solution containing 0.1% v/v crystal violet, 4% v/v formaldehyde,

14
and 30% v/v ethanol diluted in PBS. The cells were then incubated in this solution for
approximately 15 min. After incubation, the free dye solution was removed, and the cells
were washed with PBS five times. The crystal violet dye within the cells was then
extracted using 0.1% wt/v SDS diluted in PBS. The resultant extract was centrifuged
(13,000xg for 20s) to remove cellular debris. Instead of simply diluting samples for
each assay, a standard procedure was used to bring the absorbance of control extracts to
an A590 between 0.650 and 0.750. First, the initial A590 of a control sample was read on
the Molecular Devices spectra max 340PC384 plate reader, with 0.1% wt/v SDS as the
blank. Then, a series of 4, 1: 10 dilutions were made, and a plot of volume percent crystal
violet versus absorbance was made. A linear least squares line of the absorbance at these
dilutions was then used to determine the molar absorptivity of crystal violet in the control
sample. Next, the volume percent crystal violet required to give an A590 of 0.6 for the
(untreated) control samples was determined. This absorbance was then used to represent
100% viability for a given group of hepatocytes. Finally, the same dilution used for
control samples was carried out for all the samples. The analytical absorbance directly
correlated to viable cell number.
Trypan blue exclusion was used as a final means of determining cell viability.
After treatment, cells were incubated for five min with dilute trypan blue (0.1 %). Dye
was then removed, and cells were washed five times with PBS. Cells were then
visualized on a Zeiss Axiovert S100 microscope (Carl Zeiss, Thornwood, NY) fitted with
a SPOT digital CCD camera (Diagnostics Instruments, Sterling Heights, MI). Digital
pictures were taken, and viability was determined by a simple picture grid.

15
Data Analysis
Data were analyzed by ANOVA for a three-way factorial design (2x2x2) to
determine significant main effects using genotype, zinc status, and oxidant treatment as
independent variables. Fisher’s LSD was used to determine significant differences
(designated by * p< 0.05, ** p< 0.01, and *** p<0.001).
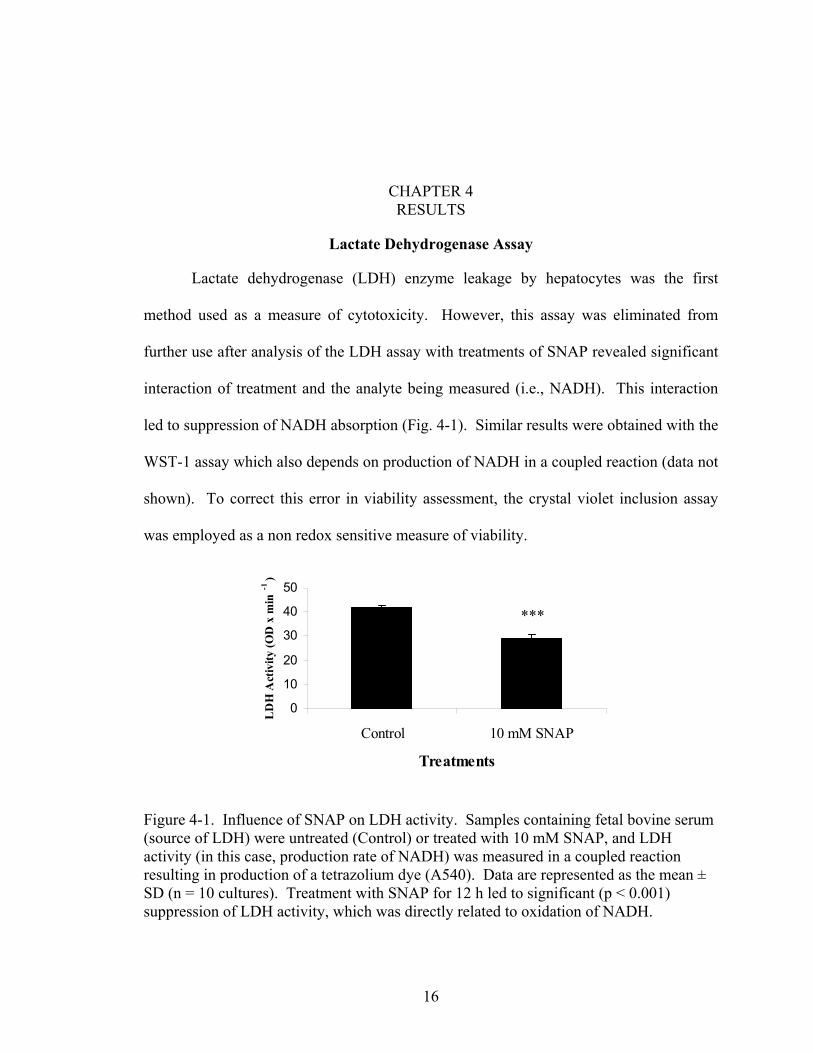
CHAPTER 4 RESULTS
Lactate Dehydrogenase Assay
Lactate dehydrogenase (LDH) enzyme leakage by hepatocytes was the first
method used as a measure of cytotoxicity. However, this assay was eliminated from
further use after analysis of the LDH assay with treatments of SNAP revealed significant
interaction of treatment and the analyte being measured (i.e., NADH). This interaction
led to suppression of NADH absorption (Fig. 4-1). Similar results were obtained with the
WST-1 assay which also depends on production of NADH in a coupled reaction (data not
shown). To correct this error in viability assessment, the crystal violet inclusion assay
was employed as a non redox sensitive measure of viability.
0
10
20
30
40
50
Control 10 m
Treatments
LD
H A
ctiv
ity (O
D x
min
-1 )
Figure 4-1. Influence of SNAP on LDH activity. Sample(source of LDH) were untreated (Control) or treated with activity (in this case, production rate of NADH) was measresulting in production of a tetrazolium dye (A540). DataSD (n = 10 cultures). Treatment with SNAP for 12 h led tsuppression of LDH activity, which was directly related to
16
***
M SNAP
s containing fetal bovine serum 10 mM SNAP, and LDH ured in a coupled reaction are represented as the mean ± o significant (p < 0.001) oxidation of NADH.

17
Metallothionein Genotype Differences
Hepatocyte viability due to genotype variation alone was investigated in hepatocyte
cultures from all four genotypes by the crystal violet assay. The consequences of tBH
and SNAP exposure on metallothionein absence or presence were determined by
utilization of KO and CK hepatocytes (Fig. 4-2A and Fig 4-2B). After 2 h tBH exposure
viability was drastically different between CK and KO cultures. At all concentrations
between 15 µM and 65 µM tBH CK hepatocytes were significantly more viable than KO
hepatocytes (p < 0.001). At concentrations of 0, 5, 10, and 75 µM tBH viability was not
significantly different between CK and KO hepatocytes. Viability was significantly
reduced at all but the lowest SNAP concentration. Effects of metallothionein
overexpression using CT and Tg genotypes are shown in Figures 4-2C and 4-2D.
Treatment with tBH led to no significant differences in viability between CT and Tg
genotypes at any concentraiton level, while treatment with SNAP at 0.5 mM was the only
significantly different concentration (p < 0.05). Examination of the concentrations of the
toxicants required to kill 50% of the hepatocytes revealed that, there is no benefit for
mice that overexpress metallothionein (Table 4-1). However, when KO mice are
compared to CK mice there is a large difference in LD50 (Table 4-1). The CK
hepatocytes required 56% more tBH, and 73% more SNAP, to reach the same level of
lethality as the KO hepatocytes. Therefore, CK cultures are more viable than KO
cultures under the same treatment conditions.

18
0 10 20 30 40 50 60 70 800
102030405060708090
100110
****** ***
*** ******
******
CK ZnN
KO ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
* CT ZnN
TG ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
CT ZnNTg ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
A
B
C
D
0.0 0.5 1.0 1.5 2.0 2.5 3.00
102030405060708090
100110
CK ZnN
KO ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol) ***
******
******
0 10 20 30 40 50 60 70 800
102030405060708090
100110
****** ***
*** ******
******
CK ZnN
KO ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
* CT ZnN
TG ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
CT ZnNTg ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
A
B
C
D
0 10 20 30 40 50 60 70 800
102030405060708090
100110
****** ***
*** ******
******
CK ZnN
KO ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
* CT ZnN
TG ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
CT ZnNTg ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
A
B
C
D
0.0 0.5 1.0 1.5 2.0 2.5 3.00
102030405060708090
100110
CK ZnN
KO ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol) ***
******
******
Figure 4-2. Viability due to altered metallothionein genotype. Hepatocyte cultures from KO and CK (left panels), and CT and Tg (right panels) mice were treated with 0-75 µM tBH for 2 h (A and C), or 0-3 mM SNAP for 12 h (B and D). Viability was assessed by the crystal violet assay. Data are represented as the mean ± SD (n = 3 cultures from a representative experiment). Statistically significant differences (designated by * for p < 0.05 and *** for p < 0.001) were determined by ANOVA and Fisher’s LSD.

19
Table 4-1. LD50 responses due to altered metallothionein genotype. MT Genotype tBH LD50 SNAP LD50
KO 20.0 ± 3 µM 0.75 ± 0.02 mM
CK 45.0 ± 2 µM*** 2.75 ± 0.13 mM***
CT 35.0 ± 2 µM 2.5 ± 0.3 mM
TG 37.0 ± 4 µM, ns 2.5 ± 0.2 mM, ns
Statistically significant differences (designated by *** for p < 0.001 and ns for not significant) were determined by ANOVA and Fisher’s LSD.
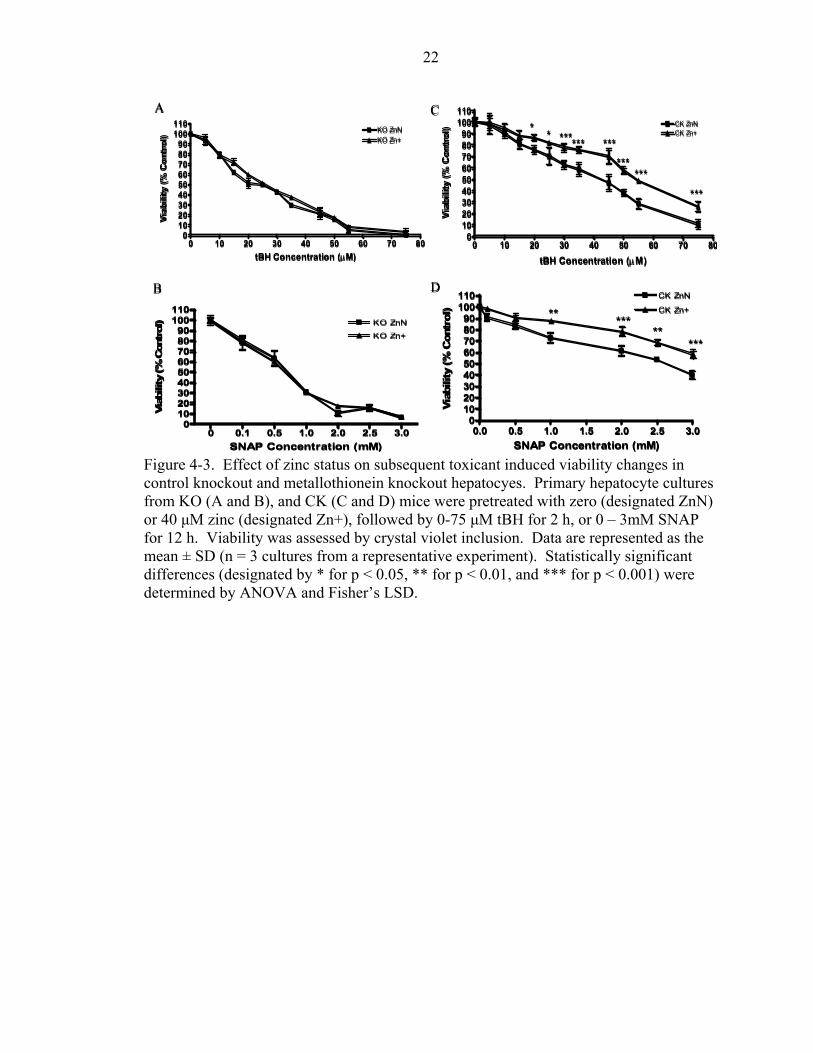
Zinc Supplementation
The effect of zinc supplementation on the individual metallothionein genotypes
was investigated using a dose of 40 µM zinc. Hepatocytes form KO, CK, CT, and Tg
mice were treated with 40 µM zinc for 18-22 h prior to addition of toxicants. Treatment
with either 0-75 µM tBH, or 0-3 mM SNAP for designatated amounts of time followed.
As expected, without the presence of metallothionein in the KO cells, there was no effect
produced by zinc on viability of tBH (Fig. 4-3A) or SNAP treated hepatocytes (Fig.
4-3B). On the contrary, zinc supplementation of CK hepatocytes prior to toxicant
exposure protected both tBH and SNAP treated cultures (Fig. 4-3C and Fig. 4-3D). The
LD50 response of CK hepatocytes supplemented with zinc to tBH and SNAP treatment
was 25% and 29% higher respectively, than zinc normal cultures (Table 4-2).
Control transgenic hepatocytes supplemented with 40 µM zinc were resistant to
tBH and SNAP over the majority of concentrations used (Fig. 4-4A and Fig. 4-4B), while
the un-supplemented hepatocytes were not. The tBH concentration required to kill 50
percent of the zinc supplemented CT hepatocytes was 18% higher than the ZnN cells, and
the LD50 for SNAP was 17% higher (Table 4-2). Zinc supplementation of

20
metallothionein transgenic hepatocytes led to no overall difference in viability (compared
to ZnN cultures) when tBH was used as a stressor (Fig. 4-4C). This was an unexpected
result considering the large difference in viabilities obtained when SNAP was used as a
stressor (Fig. 4-4D). The overall trend for SNAP treated Tg cultures was an increase in
viability with zinc treatment when compared to zinc normal cultures. Zinc treated
cultures from 1-3 mM SNAP were significantly different than the untreated counterparts.
The dose of SNAP required to kill 50% of the ZnN hepatocytes was 2.5 mM, while the
zinc supplemented hepatocytes required 3.5 mM (Fig. A-1). This represented a 29%
increase in protection of Tg hepatocytes due to zinc supplementation alone. Thus, zinc
increased viability within each genotype except for the KO group.
Altered Genotype and Zinc Supplementation
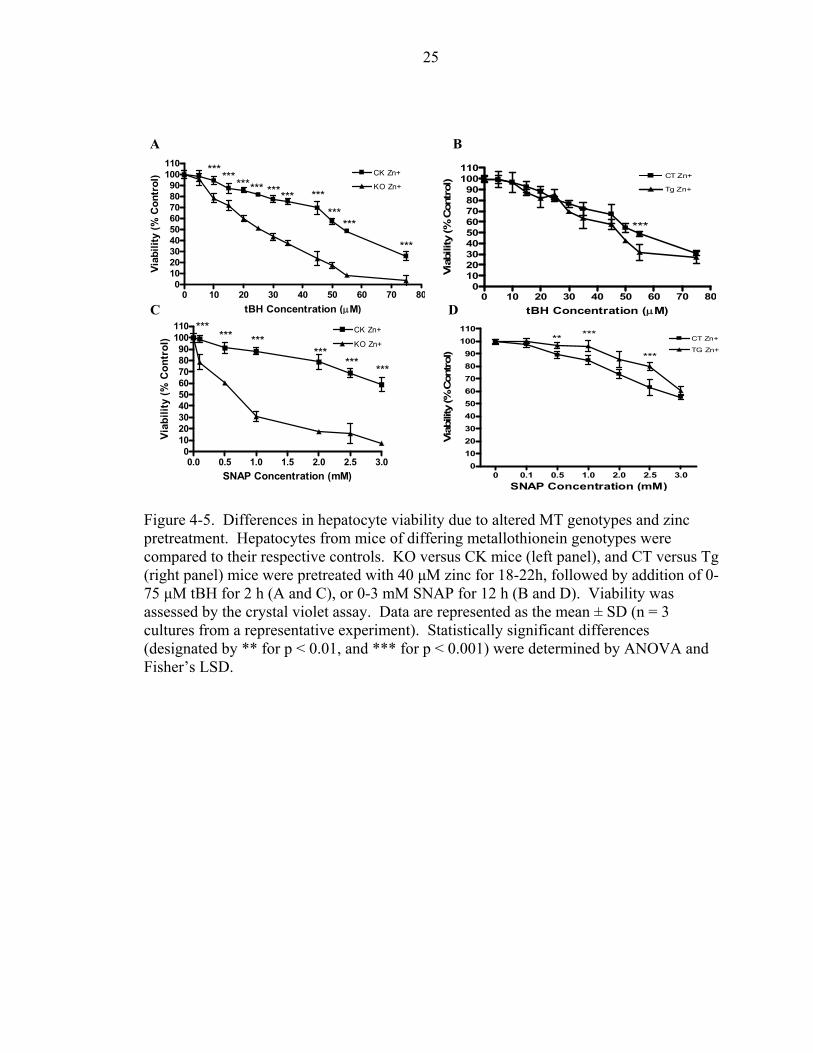
Zinc supplementation caused the greatest difference on subsequent toxicological
injury between the metallothionein genotypes. Pretreatment of CK hepatocytes with zinc
led to a large significant difference (p < 0.001) in viability from 10-75 µM tBH, when
compared to KO hepatocytes pretreated with zinc (Fig. 4-5A). The tBH concentration
required to kill 50% of CK hepatocytes was 58% higher than KO hepatocytes (Table 4-
3). The greatest difference in protection was realized by SNAP treated CK versus KO
hepatocytes. Pretreatment of CK hepatocytes with zinc led to 79% more protection
against SNAP (Table 4-3). Only one concentration of tBH (50 µM) caused a significant
difference between Tg and CT cells, but the overall trend in viability shows no difference
between the two genotypes (Fig. 4-5C). In cells treated with 0.5, 1.0, and 2.5 mM SNAP,
zinc pretreatment caused a significant increase in the viability of TG hepatocytes (Fig.
4-5D). The highest increase in viability occurred at 2.5 mM SNAP (40% increase in
viability due to TG genotype), but declined to no difference at 3.0 mM SNAP. The dose

21
of SNAP required to kill 50 percent of CT and Tg hepatocytes was the same (Fig. A-1).
As with tBH treatment, the trend in viability was the same for both CT and Tg
hepatocytes.
Metallothionein KO hepatocytes provided a means to determining the effect of
the presence of metallothionein. CK hepatocytes were significantly more viable than
their KO counterparts for both SNAP and tBH treatments. This difference was
exaggerated when zinc was supplemented in the medium (Fig. 4-5). Zinc caused an
increase in metallothionein expression which was directly related to increased CK
hepatocyte viability (Table 4-3). Hepatocytes of the Tg genotype gained no advantage in
viability under these treatment conditions (Table 4-1 and Table 4-3).
22
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
KO Zn+
KO ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
KO Zn+KO ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
A
B
C
0.0 0.5 1.0 1.5 2.0 2.5 3.00
102030405060708090
100110 CK ZnN
CK Zn+** *****
***
SNAP Concentration (mM)Vi
abili
ty (%
Con
trol
)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
**** *
*** ***
***
***
***
CK Zn+CK ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
D
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
KO Zn+
KO ZnN
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
KO Zn+KO ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
A
B
C
0.0 0.5 1.0 1.5 2.0 2.5 3.00
102030405060708090
100110 CK ZnN
CK Zn+** *****
***
SNAP Concentration (mM)Vi
abili
ty (%
Con
trol
)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
**** *
*** ***
***
***
***
CK Zn+CK ZnN
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
D
Figure 4-3. Effect of zinc status on subsequent toxicant induced viability changes in control knockout and metallothionein knockout hepatocyes. Primary hepatocyte cultures from KO (A and B), and CK (C and D) mice were pretreated with zero (designated ZnN) or 40 µM zinc (designated Zn+), followed by 0-75 µM tBH for 2 h, or 0 – 3mM SNAP for 12 h. Viability was assessed by crystal violet inclusion. Data are represented as the mean ± SD (n = 3 cultures from a representative experiment). Statistically significant differences (designated by * for p < 0.05, ** for p < 0.01, and *** for p < 0.001) were determined by ANOVA and Fisher’s LSD.

23
0 10 20 30 40 50 60 700
102030405060708090
100110
**** ****** ***
******
**
CT ZnN
CT Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
***Tg ZnN
Tg Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 0.1 0.5 1.0 2.0 2.5 3.00
1 02 03 04 05 06 07 08 09 0
1 0 01 1 0 T g Z n +
T g Z n N
*****
******
S N A P C o n c e n t r a t io n ( m M )Vi
abili
ty (%
Con
trol
)
A
B
C
D
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
****
*****
* CT ZnN
CT Zn+
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 700
102030405060708090
100110
**** ****** ***
******
**
CT ZnN
CT Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
***Tg ZnN
Tg Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 0.1 0.5 1.0 2.0 2.5 3.00
1 02 03 04 05 06 07 08 09 0
1 0 01 1 0 T g Z n +
T g Z n N
*****
******
S N A P C o n c e n t r a t io n ( m M )Vi
abili
ty (%
Con
trol
)
A
B
C
D
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
****
*****
* CT ZnN
CT Zn+
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol)
Figure 4-4. Effect of zinc status on subsequent toxicant induced viability changes in control transgenic and metallothionein transgenic hepatocyes. Primary hepatocyte cultures from CT (A and B), and Tg (C and D) mice, were pretreated with 0 (designated ZnN) or 40 µM zinc (designated Zn+), followed by 0-75 µM tBH for 2 h, or 0 – 3mM SNAP for 12 h. Viability was assessed by crystal violet inclusion. Data are represented as the mean ± SD (n = 3 cultures from a representative experiment). Statistically significant differences (designated by * for p < 0.05, ** for p < 0.01, and *** for p < 0.001) were determined by ANOVA and Fisher’s LSD.

24
Table 4-2. LD50 responses to toxicants of hepatocytes from MT KO, CK, CT, and MT Tg mice as a result of zinc status.
Genotype Toxicant Zinc status LD50 ZnN 25.0 ± 2 µM tBH Zn+ 25.0 ± 3 µM, ns ZnN 0.75 ± 0.2 mM
KO
SNAP Zn+ 0.75 ± 0.15 mM, ns ZnN 45.0 ± 2 µM tBH Zn+ 60.0 ± 4 µM*** ZnN 2.5 ± 0.3 mM
CK
SNAP Zn+ 3.5 ± 0.2 mM
ZnN 45 ± 3 µM tBH Zn+ 55 ± 1 µM*** ZnN 2.5 ± 0.4 mM
CT
SNAP Zn+ 3.0 ± 0.15 mM** ZnN 45 ± 5 µM tBH Zn+ 47 ± 3 µM, ns ZnN 2.5 ± 0.25 mM
TG
SNAP Zn+ 4.0 ± 0.11 mM***
Cultures supplemented with 40 µM zinc are labeled Zn+, unsupplemented cultures are labeled ZnN. Statistically significant differences (designated by * for p < 0.05, ** for p < 0.01, *** for p < 0.001, and ns for not significant) were determined by ANOVA and Fisher’s LSD.
25
A B
0 10 20 30 40 50 60 70 800
102030405060708090
100110 ************ ****** ***
******
***
CK Zn+
KO Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0 10 20 30 40 50 60 70 800
102030405060708090
100110
***
CT Zn+
Tg Zn+
tBH Concentration (µM)
Viab
ility
(% C
ontr
ol)
0.0 0.5 1.0 1.5 2.0 2.5 3.00
102030405060708090
100110 CK Zn+
KO Zn+
SNAP Concentration (mM)
Viab
ility
(% C
ontr
ol) *** ***
****** ***
***
0 0.1 0.5 1.0 2.0 2.5 3.00
102030405060708090
100110
** ***
***
CT Zn+
TG Zn+
SNAP Concentration (mM)
Viab
ility
(% C
ontrol
)
C D
Figure 4-5. Differences in hepatocyte viability due to altered MT genotypes and zinc pretreatment. Hepatocytes from mice of differing metallothionein genotypes were compared to their respective controls. KO versus CK mice (left panel), and CT versus Tg (right panel) mice were pretreated with 40 µM zinc for 18-22h, followed by addition of 0-75 µM tBH for 2 h (A and C), or 0-3 mM SNAP for 12 h (B and D). Viability was assessed by the crystal violet assay. Data are represented as the mean ± SD (n = 3 cultures from a representative experiment). Statistically significant differences (designated by ** for p < 0.01, and *** for p < 0.001) were determined by ANOVA and Fisher’s LSD.

26
Table 4-3. Summary of LD50 responses between the metallothionein genotypes after pretreatment by zinc.
MT Genotype tBH LD50 SNAP LD50
KO 25.0 ± 3 µM 0.75 ± 0.15 mM
CK 60.0 ± 4 µM*** 3.5 ± 0.2 mM***
CT 55.0 ± 1 µM 4.0 ± 0.25 mM
TG 50.0 ± 4 µM, ns 4.0 ± 0.33 mM, ns
Values obtained from representative experiments. Statistically significant differences (designated by * for p < 0.05, ** for p < 0.01, *** for p < 0.001, and ns for not significant) were determined by ANOVA and Fisher’s LSD.

CHAPTER 5 DISCUSSION
One common method for assessing cell viability is the lactate dehydrogenase
enzyme leakage assay. When perforation of the cellular plasma membrane occurs during
cell death, the cytosolic enzyme lactate dehydrogenase (among other enzymes) leaks out.
This leakage can be quantified by using the enzymatic conversion of lactate to pyruvate,
with subsequent formation of the analyte NADH. The highly reactive nature of NADH
(E◦ = -320 mV) suggests that in a solution with oxidative compounds, it will be oxidized.
As a consequence, an experiment was conducted to determine whether treatments to be
used in subsequent experiments would interact with (oxidize) NADH. As shown in
figure 4-1, addition of SNAP to the LDH assay led to suppression of absorbance from
NADH. Presumably, this was caused by NO production with concomitant oxidation of
NADH. Therefore, a redox-insensitive measure of viability was used for all subsequent
experiments.
Oxidative damage to cellular constituents plays a prominent role in the
pathogenesis of many diseases (Gutteridge and Halliwell 2000). Results of this study
demonstrate the importance of metallothionein in protection against cellular oxidants.
The absence of metallothionein in the knock-out hepatocytes proved to be deleterious
when cultures of CK and KO hepatocytes were treated with tertiary-butyl hydroperoxide
or S-nitroso n-acetyl penicillamine for 2 h, and 12 h respectively. The presence of
metallothionein in CK cultures provided more protection against toxicant injury than did
overexpression of metallothionein in transgenic cultures. However, because baseline
27

28
cellular metallothionein protein content of CT and Tg cultures was found to be similar
(Davis et al. unpublished data), it was not surprising to find a lack of difference between
the viabilities of CT and Tg cultures based on genotype difference alone. Therefore,
addition of 40 µM zinc prior to toxicant treatment was used to increase cellular
metallothionein levels (Cao et al. 2001). As a result, hepatocytes that were able to
increase metallothionein expression did so (data not shown).
Supplemental zinc increased viability within each genotype, except for the knock
out hepatocytes. The increase in viability of CK, CT, and Tg cells may thus be attributed
directly to zinc induced metallothionein expression. Even though zinc was able to
increase viability of CK, CT, and Tg cells when the genotypes were examined
individually, only the CK cells were more viable than the KO cultures when the
genotypes were compared to one another. Consequently, the presence of metallothionein
and its induction by zinc provided CK hepatocytes protection against oxidative insult,
while ablation of metallothionein in KO hepatocytes was deleterious (Table 4-5).
Contrary to the hypothesis, overexpression of metallothionein in transgenic cultures
provided no increased protection. These results are in agreement with previous findings
from this lab (Davis et al. 2001). Carbon tetrachloride had been used in the past to
examine the role of metallothionein in protection against hepatotoxicity. In those studies,
no combination of metallothionein gene expression (i.e., Tg, CT, CK, KO) and either
adequate or supplemental zinc provided hepatic protection. Even though, the mice had
drastically different levels of liver zinc and liver metallothionein.
Many studies have brought to light the role of metallothionein as an antioxidant.
Metallothionein has demonstrated the ability to quench reactive oxygen and reactive

29
nitrogen species at an even higher efficiency than the established cellular antioxidant
glutathione (Miura et al. 1997 and Quesada et al. 1996). The extensive network of zinc-
sulfur clusters in metallothionein provide the chemical basis for it’s antioxidant capacity
(reviewed in Maret 2000). The reduction potential of the zinc-sulfur clusters are
extremely low (E◦ < -340 mV) and thus provide a means to reduce a variety of cellular
oxidants. Even though zinc bound to metallothionein is extremely stable (pKd = 12.9),
zinc in metallothionein can readily exchange with free zinc in solution (Steinebach et al.
1992). Thus, metallothionein may play a critical role in redox sensing and zinc
distribution in the cell.
In fact, a variety of oxidative and nitrosative stressors have also been shown to
interact with zinc-sulfer clusters of zinc-finger proteins, and lead to concomitant loss of
zinc (Kroncke et al. 2002). One such nitrosative stressor is nitric oxide. NO has the
ability to release zinc from zinc dependant transcription factors, as well as
metallothionein (Schapiro et al. 2003, St. Croix et al. 2002). In vitro studies utilizing a
nitric oxide donor (DEA/ NO) have demonstrated that NO has the ability to liberate zinc
specifically from the β domain of metallothionein (Zangger et al 2001). More recent
studies in aortic endothelial cells have shown that iNOS derived NO has the ability to
increase labile zinc through interaction with metallothioenin (Spahl et al. 2003). The
current study utilized SNAP as a model for NO induced stress. The availability of
metallothionein to interact with nitric oxide in CK hepatocytes provided protection that
was not seen in KO hepatocytes. This protection was augmented when zinc increased
metallothionein expression in CK cells, but not in Tg hepatocytes (compared to CT cells).
A possible explanation for protection by the presence and induction of metallothionein by

30
zinc, but not the over-expression of MT, could be the release of zinc from MT and
subsequent signaling events such as induction of zinc dependent transcription factors or
zinc dependent proteins. In these cells, exposure to NO after zinc pretreatment caused
increased induction of metallothionein mRNA, with the largest increase in Tg
hepatocytes (Lichten et al. unpublished observations). NO mediated release of zinc from
metallothionein may be a primary means of increasing free zinc in the cell.
Consequently, the ratio of thionein (apo-metallothionein) to metallothionein may be one
mechanism with which the cell governs the levels of intracellular zinc (St. Croix et al.
2002). However, when metallothionein expression is altered, a shift in the MT/T ratio
may occur, causing increased production of thionein. Therefore, over-expression of
metallothionein may lead to production of large amounts of thionein that may sequester
zinc before it can be utilized by other apoproteins.
In these experiments, oxidative stress to the hepatocytes was produced by tBH.
The mitochondria are the primary target of this oxidative stressor (Mehendale et al.
1994). It is presumed that tBH causes mitochondrial permeability transition (MPT)
followed by mitochondrial depolarization, uncoupling of oxidative phosphorylation, ATP
depletion, and finally cell death. If metallothionein has the capacity to act as an
antioxidant, then MPT may be inhibited, and cell death (either necrosis or apoptosis) may
be prevented. Once again, the presence of metallothionein was beneficial to CK cultures,
but over expression of metallothionein in Tg cells was not (Table 4-3). However, the
magnitude of viability differences seen in the zinc supplemented CK and KO cultures
were not as great. This may be due to the difference between oxidative and nitrosative
stress towards the zinc-sulfur clusters of metallothionein. Zinc finger proteins have been

31
shown to react differently towards these stressors (Kroncke et al 2002). NO may be able
to release zinc from zinc-sulfur clusters without destroying them, while oxidative
stressors such as tBH could destroy the clusters. Thus, as with zinc finger proteins, a
reversible exchange of zinc may occur through NO interaction that does not occur when
an oxidant is applied. Therefore, zinc may be released from metallothionein in greater
amounts after interaction with tBH causing decreases in viability not only due to free
radical formation, but in the case of metallothionein transgenics, zinc toxicity.
In summary, the absence of metallothionein in MT KO hepatocytes led to
significant loss of viability after NO or tBH treatment. The presence, and induction of
metallothionein by zinc, increased the viability of CK hepatocytes, but not transgenic
hepatocytes. Therefore, there is a limit to the protection provided by metallothionein,
which directly questions the role of metallothionein as a cellular antioxidant. However,
the primary role of metallothionein may not be to act as an antioxidant, but to be a
transducer of zinc signaling.

APPENDIX CYTOTOXICITY EXPERIMENTS
0
20
40
60
80
100
120
0 1 2 3 4 5
SNAP Concentration (mM)
Viab
ility
(% c
ontr
ol) Tg Zn+
CT Zn+
Figure A-1. Differences in CT and Tg viability due to altered MT genotypes and zinc
pretreatment CT versus Tg (right panel) mice were pretreated with 40 µM zinc for 18-22h, followed by addition 0-4 mM SNAP for 12 h . Viability was assessed by the crystal violet assay
32

LIST OF REFERENCES
Andrews, G.K. (2000) Regulation of metallothionein gene expression by oxidative stressand metal ions. Biochem. Pharmacol. 59:95-104.
Andrews, G.K. (2001) Cellular zinc sensors: MTF-1 regulation of gene expression.Biometals. 14:223-37
Blain, D., Kubow, S., Chan, H.M. (1998) Zinc pretreatment inhibits isotretinoin teratogenicity and induces embryonic metallothionein in CD-1 mice. J. Nutr.128:1239-46.
Cai, L., Klein, J.B., Kang, Y.J. (2000) Metallothionein inhibits peroxynitrite-induced DNA and lipoprotein damage. J. Biol. Chem. 275:38957-60.
Cao, J., Bobo, J.A., Liuzzi, J.P., Cousins, R.J. (2001) Effects of intracellular zinc depletion on metallothionein and ZIP2 transporter expression and apoptosis. J. Leukoc. Biol. 70:559-66.
Chesters, J.K. (1992) Trace element gene interactions. Nutr. Rev. 50:21723.
Cousins, R.J. (1996) Zinc. In Present Knowledge in Nutrition, 7th ed., pp.293-306 (Filer, L.J., & Ziegler, E.E., editors). Washington, D.C.: International Life Sciences Institute-Nutrition Foundation.
Davis, S.R., Cousins, R.J. (2000) Metallothionein expression in animals: a physiological perspective on function. J. Nutr. 130:1085-8.
Davis, S.R., Samuelson, D.A., Cousins, R.J. (2001) Metallothionein expression protects against carbon tetrachloride-induced hepatotoxicity, but overexpression and dietary zinc supplementation provide no further protection in metallothionein transgenic and knockout mice. J. Nutr.131:215-22.
Dunn, M.A., Cousins, R.J. (1989) Kinetics of zinc metabolism in the rat: effect of dibutyryl cAMP. Am. J. Physiol. 256:420-30.
Farber, J.L. (1994) Mechanisms of cell injury by activated oxygen species. Environ. Health. Perspect. 102:17-24.
Fu, Y., Porres, J.M., Lei, X.G. (2001) Comparative impacts of glutathione peroxidase-1 gene knockout on oxidative stress induced by reactive oxygen and nitrogen species in mouse hepatocytes. Biochem. J. 359:687-95.
33

34
Geller, D.A., Di Silvio, M., Nussler, A.K., Wang, S.C., Shapiro, R.A., Simmons, R.L., Billiar, T.R. (1993) Nitric oxide synthase expression is induced in hepatocytes in vivo during hepatic inflammation. J. Surg. Res. 55:427-32.
Gutteridge, J.M., Halliwell, B. (2000) Free radicals and antioxidants in the year 2000. A historical look to the future. Ann. N. Y. Acad. Sci. 899:136-47.
Hyde, R.J., Hooper, N.M., Turner, A.J. (1996) Molecular cloning and expression in COS- 1 cells of pig kidney aminopeptidase P. Biochem. J.319:197-201.
Jacob, C., Maret, W., Vallee, B.L. (1998) Control of zinc transfer between thionein, metallothionein, and zinc proteins. Proc. Natl. Acad. Sci. 31:3489-94.
Jauregui, H.O., Hayner, N.T., Driscoll, J.L., Williams-Holland, R., Lipsky, M.H. & Galleti, P.M. (1981) Trypan blue uptake and lactate dehydrogenase in adult rat hepatocytes – freshly isolated cells, cell suspensions, and primary monolayer cultures. In Vitro. 17:1100-1110.
Kim, Y.M., Bombeck, C.A., Billiar, T.R. (1999) Nitric oxide as a bifunctional regulator of apoptosis. Circ. Res. 84:253-6.
Kroncke, K.D., Klotz, L.O., Suschek, C.V., Sies, H. (2002) Comparing nitrosative versus oxidative stress toward zinc finger-dependent transcription. Unique role for NO. J.Biol. Chem. 277:13294-301.
Lichtlen, P., Schaffner, W. (2001) The "metal transcription factor" MTF-1: biological facts and medical implications. Swiss Med. Wkly. 131:647-52.
Maret, W. (2000) The function of zinc metallothionein: a link between cellular zinc and redox state. J. Nutr. 130:1455-1458.
Masters, B.A., Kelly, E.J., Quaife, C.J., Brinster, R.L. & Palmiter RD. (1994) Targeted disruption of metallothionein I and II genes increases sensitivity to cadmium. Proc. Natl. Acad. Sci. U S A. ;91:584-8.
Matsubara, J. (1987) Alteration of radiosensitivity in metallothionein induced mice and a possible role of Zn-Cu-thionein in GSH-peroxidase system. Experientia. Suppl.;52:603-12.
Matsubara, J., Tajima, Y., Karasawa, M. (1987) Metallothionein induction as a potentmeans of radiation protection in mice. Radiat. Res. 111:267-75.
Miura, T., Muraoka, S., Ogiso, T. (1997) Antioxidant activity of metallothionein compared with reduced glutathione. Life Sci. 60:301-9.
Miura, T., Satoh, T., Takeuchi, H. (1988) Role of metal-ligand coordination in the folding pathway of zinc finger peptides. Biochim. Biophys. Acta. 1384:171-9.

35
Nussler, A.K., Geller, D.A., Sweetland, M.A., Di Silvio, M., Billiar, T.R., Madariaga, J.B., Simmons, R.L., Lancaster, J.R. (1993) Induction of nitric oxide synthesis and its reactions in cultured human and rat hepatocytes stimulated with cytokines plus LPS. Biochem. Biophys. Res. Commun. 194:826-35.
Palmiter, R.D., Sandgren, E.P., Koeller, D.M. & Brinster, R.L. (1993) Distal regulatoryelements from the mouse metallothionein locus stimulate gene expression intransgenic mice. Mol. Cell Biol. ;13:5266-75.
Pearce, L.L., Gandley, R.E., Han, W., Wasserloos, K., Stitt, M., Kanai, A.J., McLaughlin,M.K., Pitt, B.R., Levitan, E.S. (2000) Role of metallothionein in nitric oxidesignaling as revealed by a green fluorescent fusion protein. Proc. Natl. Acad. Sci.4;97:477-82.
Pearce, L.L., Wasserloos, K., St. Croix, C.M., Gandley, R., Levitan, E.S., Pitt, B.R. (2000) Metallothionein, nitric oxide and zinc homeostasis in vascular endothelialcells. J. Nutr. 130:1467-70.
Philcox, J.C., Coyle, P., Michalska, A., Choo, A.K.H. & Rofe, A.M. (1995) Endotoxin-induced inflammation does not cause hepatic zinc accumulation in mice lacking metallothionein gene expression. Biochem. J. 308:543-546.
Powell, S.R. (2000) The antioxidant properties of zinc. J. Nutr. 130:1447-54.
Quesada, A.R., Byrnes, R.W., Krezoski, S.O., Petering, D.H. (1996) Direct reaction ofH2O2 with sulfhydryl groups in HL-60 cells: zinc-metallothionein and other sites. Arch. Biochem. Biophys. 334:241-50.
St. Croix, C.M., Wasserloos, K.J., Dineley, K.E., Reynolds, I.J., Levitan, E.S., Pitt, B.R.(2002) Nitric oxide-induced changes in intracellular zinc homeostasis are mediated by metallothionein/thionein. Am. J. Physiol. Lung Cell Mol. Physiol. 282:185-92.
Satoh, M., Naganuma, A., Imura, N. (1988) Involvement of cardiac metallothionein inprevention of adriamycin induced lipid peroxidation in the heart. Toxicology.53:231-7.
Schapiro, J.M., Libby, S.J., Fang, F.C. (2003) Inhibition of bacterial DNA replication by zinc mobilization during nitrosative stress. Proc. Natl. Acad. Sci. U. S. A. 100:8496-501.
Schroeder, J.J., Cousins, R.J. (1990) Interleukin 6 regulates metallothionein geneexpression and zinc metabolism in hepatocyte monolayer cultures. Proc. Natl.Acad. Sci. U. S. A. 87:3137-41.
Schroeder, J.J., Cousins, R.J. (1991a) Maintenance of zinc-dependent hepatic functions in rat hepatocytes cultured in medium without added zinc. J. Nutr. 121:844-53.

36
Schroeder, J.J., Cousins, R.J. (1991b) Metallothionein and zinc metabolism in hepatocytes. Methods. Enzymol. 205:575-84.
Schwarz, M.A., Lazo, J.S., Yalowich, J.C., Allen, W.P., Whitmore, M., Bergonia, H.A.,
Spahl, D.U., Berendji-Grun, D., Suschek, C.V., Kolb-Bachofen, V., Kroncke, K.D. (2003) Regulation of zinc homeostasis by inducible NO synthase-derived NO: nuclear metallothionein translocation and intranuclear Zn2+ release. Proc. Natl. Acad. Sci. U. S. A. 100:13952-7.
Steinebach, O.M., Wolterbeek, B.T. (1992) Metallothionein biodegradation in rathepatoma cells: a compartmental analysis aided 35S-radiotracer study. Biochim. Biophys. Acta. 1116:155-65.
Thornalley, P.J. & Vasak, M. (1985) Possible role for metallothionein in protectionagainst radiation-induced oxidative stress: Kinetics and mechanism of its reactionwith superoxide and hydroxyl radicals. Biochem. Biophys. Acta. 827:36-44.
Turner, J.S., Glands, P.D., Samson, A.C., Robinson, N.J. (1996) Zn2+-sensing by the cyanobacterial metallothionein repressor SmtB: different motifs mediate metal-induced protein-DNA dissociation. Nucleic Acids Res. 24:3714-21.
Tzeng, E., Billiar, T.R., Robbins, P.D., Lancaster, jr, J.R., & Pitt, B.R. (1995)Metallothionein Protects Against the Cytotoxic and DNA-Damaging Effects ofNitric Oxide. Proc. Natl. Acad. Sci. 92: 4452-4456.
Vallee, B.L., Coleman, J.E., Auld, D.S. (1991) Zinc fingers, zinc clusters, and zinc twists in DNA-binding protein domains. Proc. Natl. Acad. Sci. U. S. A. 88:999-1003.
Wink, D.A., Mitchell, J.B. (1998) Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic. Biol. Med. 25:434-56.
Yu, B.P. (1994) Cellular damage from reactive oxygen species. Physiol. Rev. 74:139-162.
Zago, M.P., Oteiza, P.I. (2001) The antioxidant properties of zinc: interactions with ironand antioxidants. Free Radic. Biol. Med. 31:266-74.
Zangger, K., Oz, G., Haslinger, E., Kunert, O., Armitage, I.M. (2001) Nitric oxideselectively releases metals from the amino-terminal domain of metallothioneins: potential role at inflammatory sites. FASEB J. 15:1303-5.
Zhou, Z., Sun, X., James, Kang, Y. (2002) Metallothionein protection against alcoholicliver injury through inhibition of oxidative stress. Exp. Biol. Med. 227:214-22.

BIOGRAPHICAL SKETCH
Louis Lichten was born in Miami, Florida on April 24, 1979. He received his
Bachelor of Science degree from the University of Florida in 2002, and plans to continue
on in the doctorate program for nutritional sciences.
37