theiler’s virus infection of primary cultures of bone marrow
TRANSCRIPT

JOURNAL OF VIROLOGY, Dec. 2002, p. 12823–12833 Vol. 76, No. 240022-538X/02/$04.00�0 DOI: 10.1128/JVI.76.24.12823–12833.2002Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Theiler’s Virus Infection of Primary Cultures of BoneMarrow-Derived Monocytes/Macrophages
Cecile Martinat,† Ignacio Mena, and Michel Brahic*Unite des Virus Lents, CNRS URA 1930, Departement de Virologie, Institut Pasteur,
75724 Paris Cedex 15, France
Received 5 April 2002/Accepted 18 September 2002
Theiler’s virus, a murine picornavirus, causes a persistent infection of macrophage/microglial cells in thecentral nervous systems of SJL/J mice. Viral replication is restricted in the majority of infected cells, whereasa minority of them contain large amounts of viral RNA and antigens. For the present work, we infected primarycultures of bone marrow monocytes/macrophages from SJL/J mice with Theiler’s virus. During the first 10 hpostinfection (p.i.), infected monocytes/macrophages were round and covered with filopodia and containedlarge amounts of viral antigens throughout their cytoplasm. Later on, they were large, flat, and devoid offilopodia and they contained only small amounts of viral antigens distributed in discrete inclusions. These twotypes of infected cells were very reminiscent of the two types of infected macrophages found in the spinal cordsof SJL/J mice. At the peak of virus production, the viral yield per cell was approximately 200 times lower thanthat for BHK-21 cells. Cell death occurred in the culture during the first 24 h p.i. but not thereafter. No infectedcells could be detected after 4 days p.i., and the infection never spread to 100% of the cells. This restriction wasunchanged by treating the medium at pH 2 but was abolished by treating it with a neutralizing alpha/betainterferon antiserum, indicating a role for this cytokine in limiting virus expression in monocyte/macrophagecultures. The role of alpha/beta interferon was confirmed by the observation that monocytes/macrophages fromIFNA/BR�/� mice were fully permissive.
Theiler’s murine encephalomyelitis virus (TMEV), a mem-ber of the picornavirus family, is a naturally occurring entericpathogen of mice that causes a persistent infection of thecentral nervous system (CNS) and a chronic demyelinatingdisease resembling multiple sclerosis (19). Intracranial inocu-lation of TMEV in genetically susceptible mice results in abiphasic disease. The first phase is a mild encephalomyelitisduring which the virus is found in the gray matter of the brainand spinal cord. During the second phase, the virus disappearsfrom the gray matter and infects the white matter of the spinalcord where it persists for the lifetime of the animal. Viralpersistence results in chronic inflammation and primary demy-elination. Susceptibility to viral persistence and to the accom-panying pathologic changes varies greatly between inbredstrains of mice. Some are highly susceptible (e.g., the SJL/Jstrain), and others are completely resistant (e.g., the C57BL/6strain). Strains which are resistant to persistent infection clearthe infection after the early encephalomyelitis. Susceptibility toTMEV’s persistence and to the virus-induced demyelinatingdisease is multigenic and complex (for a review, see reference3).
TMEV-induced demyelination appears to be, at least inpart, immune mediated. Virus-specific CD4� T cells, respon-sible for a delayed-type hypersensitivity reaction, have beenimplicated (6, 10, 11, 12). Myelin epitope-specific CD4� T
cells, primed via epitope spreading, could also play a role in thelate stages of the disease (17, 24). According to the prevalentview, macrophages recruited and activated by virus- and mye-lin-specific CD4� T cells are the effectors of demyelination.Indeed, macrophages laden with myelin debris are common inwhite matter lesions, and the depletion of infiltrating macro-phages with mannosylated liposomes reduces demyelinatinglesions drastically (30).
Macrophages may play another important role in pathogen-esis, as reservoirs of viral persistence. Lipton et al. have shown,by double immunostaining, that the main burden of virus an-tigen resides in macrophages (21). This has been confirmed byPena-Rossi et al., using combined immunocytochemistry-insitu hybridization (30). The origin of these infected macro-phages, whether they are hematogenous infiltrating monocytesor microglial cells, remained an open question until Pena-Rossi et al. showed that depleting monocytes/macrophageswith liposomes cleared the CNS of persistent infection almostcompletely (30). More recently, Drescher et al. showed thattreating mice with transforming growth factor TGF-�2, a po-tent immunoregulatory mediator, decreased the virus load inthe white matter of the spinal cord by reducing the number ofinfiltrating macrophages (8). Taken together, these resultsstrongly indicate that blood-borne macrophages are the mainviral targets during persistent infection.
Two types of infected cells have been observed in the CNSduring viral persistence. Some of them contain large amountsof viral RNA and antigens, but the majority contain limitedamounts of viral products, indicating restricted viral replication(5, 21). These two types of cells could play different roles inviral persistence. The former might be important to propagatethe infection to new target cells, whereas the latter might serve
* Corresponding author. Mailing address: Unite des Virus Lents,CNRS URA 1930, Institut Pasteur, 28, rue du Docteur Roux, 75724Paris Cedex 15, France, Phone: (33-1) 45.68.87.70, Fax: (33-1)40.61.31.67. E-mail: [email protected].
† Present address: Taub Institute for the Aging Brain, Departmentsof Pathology and Neurology, Center for Neurobiology and Behavior,Columbia University, New York, NY 10032
12823
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

as a reservoir of persistent infection. Studies with macrophage-derived cell lines indicate that permissiveness to TMEV de-pends on the differentiation/activation status of the cell (14–16,32). Therefore, it has been hypothesized that the two types ofinfected cells observed in vivo could correspond to monocytes/macrophages at different stages of differentiation/activation.
Although they offer obvious practical advantages, tissue cul-ture-adapted macrophage cell lines are significantly differentfrom the original cells. Furthermore, as already mentioned,susceptibility to persistence of TMEV depends critically on thegenetic background of the mouse, a parameter not taken intoaccount when using macrophage cell lines. Therefore, we de-cided to study the infection of primary cultures of monocytes/macrophages derived from the bone marrow of SJL/J mice, astrain susceptible to persistent infection. We report that, 10 hpostinfection (p.i.), the majority of infected cells were roundand covered with filopodia and that viral replication was activein these cells. The morphology of infected cells changed withtime. They became flat, with a large cytoplasm and no filopo-dia. At that stage, viral replication was restricted due to thesecretion of alpha/beta interferon (IFN-�/�).
MATERIALS AND METHODS
Viruses. The DA strain of TMEV was plaque purified on BHK-21 cells, andthe virus titers were determined by standard plaque assay on BHK-21 cells (20).
Vesicular stomatitis virus (VSV), strain Indiana, was kindly provided by ElianeMeurs (Pasteur Institute), and the virus titers were determined by conventionalplaque assay on NIH 3T3 cells.
Primary cultures of bone marrow-derived macrophages. SJL/J mice werepurchased from Janvier (Le Genest-St-Isle, France). IFNA/BR�/� mice werebred in the Institut Pasteur animal facility. Bone marrow cells were flushed fromfemurs and tibias of 8- to 9-week-old female mice. After depletion of red bloodcells by using Gey solution and low-speed centrifugation, the cell pellet wasresuspended in complete medium consisting of RPMI 1640 medium (Prolabo)supplemented with 1% penicillin-streptomycin (Gibco), 10% heat-inactivatedfetal bovine serum (FBS) (Gibco), and 10% L929 cell-conditioned medium as asource of macrophage colony-stimulating factor (M-CSF). The latter was pre-pared by plating 8 � 106 L929 cells in a T75 flask containing 40 ml of RPMI 1640medium supplemented with 10% FBS. After 1 week at 37°C in a 5% CO2
humidified atmosphere, the medium was harvested, filtered, and stored at �20°Cuntil use. Bone marrow cells were seeded at a density of 1.5 � 107 cells onto100-mm-diameter tissue culture dishes (Falcon). Every other day, one-thirdvolume of complete medium was added to the culture. After 6 days at 37°C in a5% CO2 humidified atmosphere, nonadherent cell clusters were gently flushedand centrifuged at low speed. The cell pellet was resuspended in a mixturecontaining RPMI 1640 medium, 1% penicillin-streptomycin, 10% FBS, and 2.5%L929 cell-conditioned medium (incomplete medium). Cells do not divide inincomplete medium. Cells were dissociated mechanically through needles, thenseeded onto 12-mm-diameter coverslips, in 24-well plates, at a density of 2 � 105
cells per well. Cells were cultured overnight at 37°C before being infected. Thecells were kept in incomplete medium throughout the experiment.
Infection of primary cultures of bone marrow-derived macrophages. Cellswere washed twice in RPMI 1640 medium to remove serum components, and0.35 ml of appropriately diluted virus stock solution was added to each well. Cellcultures were infected at a multiplicity of infection (MOI) of 5 PFU per cell.After adsorption of the virus for 2 h at 37°C, cells were washed twice with RPMI1640 medium and cultured with 0.5 ml of incomplete medium containing only2% FBS.
Infection of BHK-21 cells. BHK-21 cells were grown in six-well plates, washedtwice in PBS, and infected at an MOI of 5 PFU/cell in 0.5 ml. After adsorption
of the virus for 45 min at 25°C, 2.5 ml of medium containing 1% FBS was addedto each well.
Cell viability assays. The Alamar Blue assay (Interchim) consists of an oxi-doreduction indicator that changes color with cell growth. At different timespostinfection, one-tenth volume of Alamar Blue reagent was added to the me-dium of TMEV- and mock-infected cultures. After 3 h of incubation at 37°C, theabsorbance of the medium was measured at wavelengths of 570 and 600 nm (13).
To determine the number of macrophages in TMEV- and mock-infectedcultures, cells were lysed with 0.1 M citric acid containing 0.05% naphtol blueblack (Aldrich) and 1% cetrimide (Sigma). Under these conditions, only mac-rophage nuclei remain intact. Macrophage nuclei were counted in a hemacytom-eter (1, 28).
For both assays, the results for infected cultures were expressed as the per-centage of the number of viable cells in noninfected cultures treated in parallel.
Immunofluorescence labeling. Immunostaining was done directly on cellsseeded on glass coverslips. The cells were fixed with 4% (wt/vol) paraformalde-hyde (Electron Microscopy Sciences) for 15 min at room temperature and thenquenched with 50 mM NH4Cl in phosphate-buffered saline (PBS).
Cells were made permeable to reagents by incubating the coverslips for 20 minin PBS–2% (wt/vol) bovine serum albumin–0.1% saponin (Sigma) or for 5 min inPBS–0.1% Triton X-100 for the detection of, respectively, the intracellularMOMA-2 and TMEV antigens.
Nonspecific antibody-binding sites were blocked by incubating the coverslipsfor 30 min in PBS–2% normal goat serum (Sigma).
The following primary antibodies were used to characterize macrophages: ratmonoclonal MOMA-2 (Serotec), rat monoclonal FA/11 (kind gift of GenevieveMilon, Pasteur Institute), rat monoclonal anti-F4/80 (Serotec), biotinylated ratmonoclonal anti-Mac-1 (Pharmigen) and fluorescein isothiocyanate (FITC)-con-jugated mouse monoclonal anti-I-Ak, f, r, s (Pharmingen; clone 10-3.6). Cells werereacted with the primary antibody, diluted as indicated by the manufacturer for2 h at room temperature. After repeated washing steps in PBS–0.1 mg/ml sapo-nin–2% (wt/vol) bovine serum albumin, coverslips were incubated with Cy3-conjugated goat antirat immunoglobulin G (IgG) (Jackson Immuno ResearchLaboratories) (for MOMA-2, FA/11, and F4/80) or with Cy3-conjugated Strepta-vidin (Jackson Immuno Research Laboratories) (for Mac-1). To amplify thesignal given by the anti-I-A antibody, cells were incubated with peroxidase-conjugated anti-FITC (Boerhinger) and then with FITC-conjugated tyramide(NEN Life Science).
TMEV antigens were detected with an anticapsid rabbit hyperimmune serum(4) followed by biotinylated antirabbit IgG (Diagnostic Pasteur) and Cy3-conju-gated streptavidin (Jackson Immuno Research Laboratories) or with an FITC-conjugated antirabbit IgG (Jackson Immuno Research Laboratories). In someexperiments, nuclei were counter-stained with 0.2 �M ethidium homodimer 1(Sigma).
RESULTS
Bone marrow-derived monocyte/macrophage cultures. Sin-gle-cell suspensions were obtained from the bone marrow ofSJL/J mice and grown in vitro with L929 cell-conditioned me-dium as a source of M-CSF. M-CSF is a growth factor that actsprimarily on bone marrow precursors committed to the mono-cytic lineage (23, 33). It induces their proliferation and differ-entiation in vitro into clusters of macrophages (25). After 6days, the cultures contained adherent cells composed mainly ofmature macrophages and fibroblasts and numerous clusters ofnonadherent cells. These clusters were gently harvested anddissociated. A sample was stained with May-Grunwald Giem-sa. More than 95% of the cells were phagocytic mononuclearcells and a small percentage were polymorphonuclear leuko-cytes (data not shown). The cells were plated on glass cover-slips and analyzed by indirect immunofluorescence for the
FIG. 1. Characterization of bone marrow-derived monocytes/macrophages by immunofluorescence using macrophage-specific antigens.(A) Rat monoclonal MOMA-2. (B) Rat monoclonal FA/11. (C and D) Mouse monoclonal anti-I-Ak, f, r, s. For each field, phase contrast is shownon the right. For panels A, B, and C, bars � 100 �m. For panel D, bar � 26.5 �m.
12824 MARTINAT ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

12825
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

expression of markers of macrophage differentiation. All thecells expressed MOMA-2, FA-11 (Fig. 1A and B), F4/80, andMac-1 (not shown). Only a few cells (approximately 3%) ex-pressed major histocompatibility complex (MHC) class II mol-ecules (Fig. 1C and D). Thus, these cultured cells had themorphology and the phenotype of phagocytic mononuclearcells differentiated into monocytes/resident macrophages (22).The few MHC class II-positive cells present in the culturecould have been dendritic cell precursors.
Infection of bone marrow-derived monocytes/macrophageswith TMEV. Monocytes/macrophages grown in vitro were in-fected with TMEV at an MOI of 5 PFU/cell, as described inMaterials and Methods, and observed for 13 days. Some cyto-pathic effect was observed 24 h after infection but not there-after. To confirm and quantify this observation, cell viabilitywas measured by using both the Alamar Blue and the naphtolblue black assays. The results are presented in Fig. 2 as per-centages of the number of viable cells in mock-infected cul-tures examined at the same time. As shown in the figure,viability was 100% at 10 h p.i. and then it decreased to about77% by 27 h p.i. and remained at that level until 13 days p.i.
The extent of viral infection in these cultures was examinedby detecting TMEV capsid antigens with immunofluorescence.The percentage of cells that contained TMEV antigen wasmeasured at different times p.i. by counting the total number ofcells and the number of antigen-positive cells in approximately10 randomly chosen fields per coverslip (between 400 and1,000 cells were examined for each coverslip) (Table 1). Ap-proximately 15% of the cells expressed viral antigens at 10 h
p.i. This percentage reached a maximum of about 25% be-tween 27 and 54 h p.i. After 27 h, the percentage of infectedcells decreased progressively and by 13 days p.i. no viral anti-gens could be detected in the cultures. To confirm the absenceof infection at 13 days p.i., the cells were hybridized in situ witha radioactive probe specific for TMEV RNA (18). No viralRNA could be detected (data not shown). It should be notedthat the disappearance of the infection was not due to a de-crease of the number of viable cells in the culture (Fig. 2).
Interestingly, two types of virus antigen-containing cellswere observed (Fig. 3). At 10 h p.i., approximately 60% ofinfected cells were round and covered with filopodia (Fig. 3Aand 3B). Viral antigens were uniformly distributed in the cy-toplasm of these cells, and fluorescent staining was intense,suggesting active virus replication. The other infected cellswere larger, flat, and devoid of filopodia. Viral antigens formedinclusions in them, often with a punctate appearance. Overall,fluorescence was weaker in these cells, suggesting restrictedviral replication. This second type predominated at 27 h p.i.(Fig. 3C) and was the only type present at 4 days p.i. (Fig. 3D).
Viral titers in the culture medium were measured at differ-ent times p.i. by using a plaque assay on BHK-21 cells. Theaverage titers obtained in four independent experiments areshown in Fig. 4. A peak of infectivity was observed at 27 h p.i.Interestingly, this is also the time at which cell death and thenumber of infected cells were at maximum levels (Fig. 2). Forcomparison, the figure shows the titers in the medium ofBHK-21 cells infected at the same MOI of 5 PFU/cell.
FIG. 2. Characterization of bone marrow-derived monocytes/macrophages by immunofluorescence using macrophage-specific antigens.(A) Rat monoclonal MOMA-2. (B) Rat monoclonal FA/11. (C and D) Mouse monoclonal anti-I-Ak, f, r, s. For each field, phase contrast is shownon the right. For panels A, B, and C, bars � 100 �m. For panel D, bar � 26.5 �m.
FIG. 3. Morphology of TMEV antigen-containing cells at different times p.i. (A and B) At 10 h p.i., the majority of infected cells were roundand covered with filopodia. For panel A, the same field is shown under phase contrast. Panel B shows two different fields. (C) At 27 h p.i., themajority of infected cells were spread out and had a large cytoplasm and no filopodia. (D) This kind of infected cell was the only one detected at4 days p.i. Bars � 26.5 �m.
12826 MARTINAT ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

12827
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

Characteristics of infected cells. We examined whether theinfection by TMEV altered the phagocytic activity of mono-cytes/macrophages and induced the expression of class II an-tigens.
FITC-labeled latex particles (2 �m diameter, Polysciences)were added to infected and mock-infected cultures at differenttimes p.i. at a particle-to-cell ratio of 50:1. After incubation at37°C for 1.5 h, the cells were washed several times with PBS,fixed with 4% paraformaldehyde, and stained for TMEV an-tigens as described in Materials and Methods. The number ofparticles per infected and uninfected cell was determined.Cells were also incubated with particles at 4°C, a temperaturewhich blocks phagocytosis, to determine the background ofadsorbed, as opposed to ingested, particles per cell (26, 29). Nostatistically significant difference in the number of ingestedparticles per cell was observed between infected and mock-infected cells at all times p.i. examined. Moreover, phagocyto-sis was not statistically different in the two types of infectedcells described above (data not shown).
The expression of MHC class II molecules on infected cellswas examined using double immunostaining for TMEV-anti-gens and MHC class II molecules. As described above, unin-fected cultures contained an average of 3.4% class II-positivecells. The number of class II-positive cells decreased progres-sively after infection. No class II-positive cells were detected ininfected cultures after day 4 p.i. However, class II-positive cellsdisappeared with time also in mock-infected cultures, indicat-
FIG. 4. Virus titers in the supernatants of infected cells, at differenttimes p.i. �, primary cultures of bone marrow-derived monocytes/mac-rophages; each point is the average of four independent experiments.{, BHK-21 cells; each point is the average of two independent exper-iments. Standard errors are shown.
FIG. 5. Detection of antiviral factors in supernatants of infected primary cultures of bone marrow-derived monocytes/macrophages. (A) Flowchart of the experiment. (B) Flow chart for neutralization assays with the anti-IFN-�/� polyclonal antibody. (C) Results obtained, in twoindependent experiments, for primary cultures treated with TMEV-infected (�), or mock-infected (■ ) supernatants harvested at 27 h p.i. Standarderrors are shown. �, P � 0,0001 as determined by a 2 test.
12828 MARTINAT ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.
ing that the decrease of expression of these molecules was notdue to the virus.
In summary, our data show that infection with TMEV didnot modify the capacity of bone marrow-derived monocytes/
macrophages to phagocytose latex beads and did not inducethe expression of MHC class II molecules.
Monocytes/macrophages infected with TMEV secrete a sol-uble antiviral factor. The fact that the infection did not spreadto 100% of the cells suggested that either there were twosubpopulations of monocytes/macrophages, and only one waspermissive to the virus, or that infected cells released a solublefactor which made the other cells in the culture resistant. Theexperiment outlined in Fig. 5A was designed to test the secondhypothesis. The medium of infected cultures (“conditionedmedium”) was harvested either at 10, 27, or 96 h p.i. and spunat 100,000 � g to remove viral particles as well as cell debris.Fresh primary cultures were incubated with conditioned me-dium for 24 h. This conditioned medium was then removedand kept at 37°C, and the cultures were infected with TMEV (5
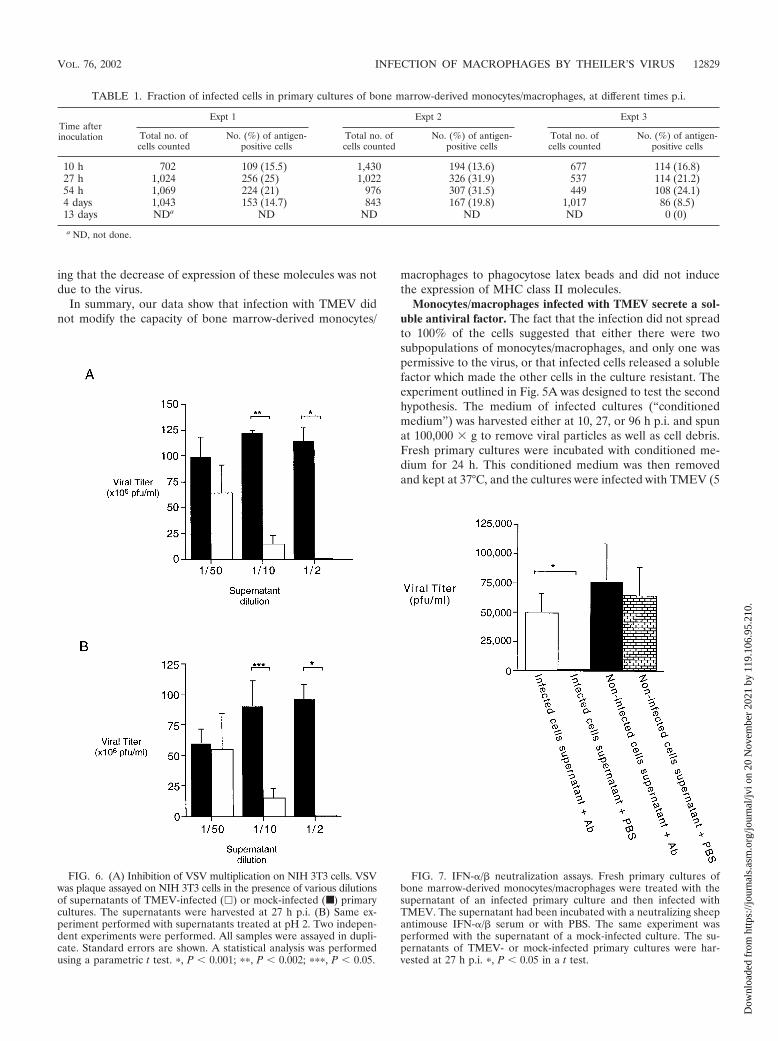
FIG. 6. (A) Inhibition of VSV multiplication on NIH 3T3 cells. VSVwas plaque assayed on NIH 3T3 cells in the presence of various dilutionsof supernatants of TMEV-infected (�) or mock-infected (■ ) primarycultures. The supernatants were harvested at 27 h p.i. (B) Same ex-periment performed with supernatants treated at pH 2. Two indepen-dent experiments were performed. All samples were assayed in dupli-cate. Standard errors are shown. A statistical analysis was performedusing a parametric t test. �, P � 0.001; ��, P � 0.002; ���, P � 0.05.
FIG. 7. IFN-�/� neutralization assays. Fresh primary cultures ofbone marrow-derived monocytes/macrophages were treated with thesupernatant of an infected primary culture and then infected withTMEV. The supernatant had been incubated with a neutralizing sheepantimouse IFN-�/� serum or with PBS. The same experiment wasperformed with the supernatant of a mock-infected culture. The su-pernatants of TMEV- or mock-infected primary cultures were har-vested at 27 h p.i. �, P � 0.05 in a t test.
TABLE 1. Fraction of infected cells in primary cultures of bone marrow-derived monocytes/macrophages, at different times p.i.
Time afterinoculation
Expt 1 Expt 2 Expt 3
Total no. ofcells counted
No. (%) of antigen-positive cells
Total no. ofcells counted
No. (%) of antigen-positive cells
Total no. ofcells counted
No. (%) of antigen-positive cells
10 h 702 109 (15.5) 1,430 194 (13.6) 677 114 (16.8)27 h 1,024 256 (25) 1,022 326 (31.9) 537 114 (21.2)54 h 1,069 224 (21) 976 307 (31.5) 449 108 (24.1)4 days 1,043 153 (14.7) 843 167 (19.8) 1,017 86 (8.5)13 days NDa ND ND ND ND 0 (0)
a ND, not done.
VOL. 76, 2002 INFECTION OF MACROPHAGES BY THEILER’S VIRUS 12829
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.
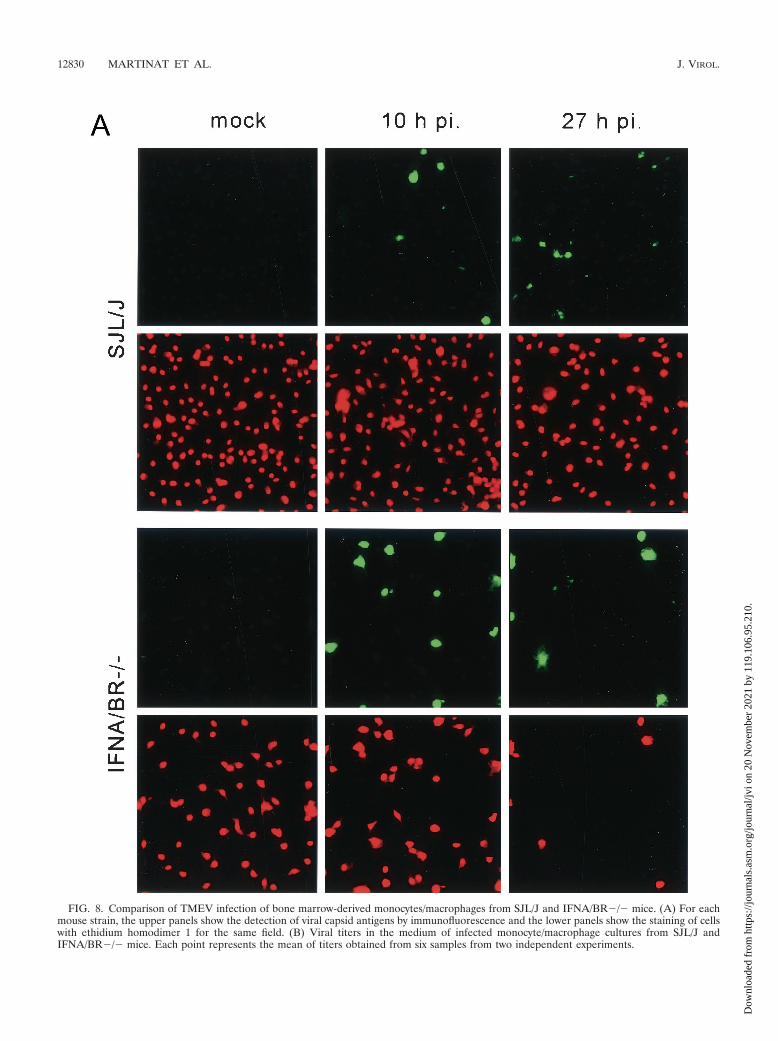
FIG. 8. Comparison of TMEV infection of bone marrow-derived monocytes/macrophages from SJL/J and IFNA/BR�/� mice. (A) For eachmouse strain, the upper panels show the detection of viral capsid antigens by immunofluorescence and the lower panels show the staining of cellswith ethidium homodimer 1 for the same field. (B) Viral titers in the medium of infected monocyte/macrophage cultures from SJL/J andIFNA/BR�/� mice. Each point represents the mean of titers obtained from six samples from two independent experiments.
12830 MARTINAT ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

PFU/cell) in a small volume. After adsorption of the virus, theconditioned medium was returned to the cultures. The cellswere examined for the presence of viral antigens by immuno-fluorescence at various times p.i. The experiment was alsoperformed by using medium from mock-infected cultures as acontrol.
As shown in Fig. 5C, there was a statistically significantdecrease in the number of infected cells in cultures treatedwith conditioned medium. Moreover, the number of infectedcells did not increase between 10 and 27 h p.i. in culturestreated with conditioned medium as opposed to culturestreated with control medium. In cultures treated with condi-tioned medium, all infected cells had large cytoplasms withpunctate inclusions of viral antigens. Figure 5C shows the re-sults obtained with conditioned medium harvested at 27 h p.i.Similar results were obtained with conditioned medium col-lected at 10 or 96 h p.i. (data not shown).
In summary, these data indicate that infected monocytes/macrophages release one or several antiviral soluble factors inthe medium.
Infected monocytes/macrophages secrete IFN-�/�. To de-termine if the antiviral effect was specific for TMEV, we testedthe effect of conditioned medium on the replication of VSV inNIH 3T3 cells. Conditioned medium was centrifuged as de-scribed above and diluted serially in Dulbecco’s modified Ea-gle medium (DMEM)–10% FBS. Monolayers of NIH 3T3 cellswere incubated for 24 h with dilutions of conditioned medium,rinsed with DMEM, and infected with 70 PFU of VSV. Afteradsorption, the cells were overlaid with agarose to perform astandard plaque assay. As shown in Fig. 6A, the multiplicationof VSV was inhibited in a dose-dependent manner by condi-
tioned medium. The drop in titer was no longer statisticallysignificant for a 1:50 dilution.
IFN-�/� is the only cytokine that is stable after treatment atpH 2 (34). To determine if the inhibitory effect of conditionedmedium was due to IFN-�/�, the experiment was repeatedafter treating the medium at pH 2 for 48 h at 4°C followed byneutralization to pH 7.0. As shown in Fig. 6B, treatment at pH2 did not impair the inhibitory effect, strongly suggesting thatinhibition was due to IFN-�/�.
Neutralization assays with a specific hyperimmune serumwere performed to confirm that the inhibition was due toIFN-�/� (Fig. 5B). Conditioned medium, centrifuged at highspeed as described above, was incubated for 1 h at 37°C fol-lowed by 1 h at 4°C with an amount of hyperimmune sheepantiserum able to neutralize 5,000 U of IFN-�/� (a kind gift ofI. Gresser, Institut Curie, Paris) (2, 31). Conditioned mediumwas also incubated with PBS as control. Fresh monocyte/mac-rophage cultures were incubated with these media for 24 h at37°C. The media were removed, and the cells were washed withRPMI and infected with TMEV at an MOI of 5 PFU/cell.After adsorption, the cells were washed again with RPMI andcultured with fresh RPMI–2% FBS–2.5% M-CSF. After 27 h,viral production was measured with a plaque assay usingBHK-21 cells.
As shown in Fig. 7, no infectivity was detected when the cellshad been treated with nonneutralized conditioned medium,confirming the presence of an inhibitory factor in the medium.Approximately 5 � 104 PFU/ml was detected when the condi-tioned medium had been treated with the neutralizing anti-interferon serum or when the conditioned medium was frommock-infected cultures. This result confirmed that the antiviralfactor released by TMEV-infected monocytes/macrophageswas IFN-�/�.
Last, we tested the role of IFN-�/� directly by using primarycultures of bone marrow monocytes/macrophages derivedfrom mice with an inactivated gene for the IFN-�/� receptor(IFNA/BR�/�) (27). The procedure for culturing and infectingthese cells was identical to that used for SJL/J mice. Staining byimmunofluorescence for viral antigens showed that, in contrastto SJL/J macrophages, 100% of IFNA/BR�/� macrophagesbecame infected between 10 and 24 h p.i. Infected cells wereround and contained large amounts of viral antigens (Fig. 8A).This infection resulted in 100% cell death. As shown in Fig. 8B,viral titers released in the medium were more than 10 timeshigher than for SJL/J macrophages, although they were stillapproximately 100 times lower than for BHK-21 cells.
DISCUSSION
Results from several laboratories indicate that infiltratingCNS macrophages are the main viral reservoir in SJL/J micepersistently infected with TMEV (21, 30). A dichotomy hasbeen observed among these cells (5, 21). Some contain largeamounts of viral RNA and antigens, whereas viral expression isrestricted in others. The former could be important in spread-ing the infection within the CNS, and indeed it has been shownthat macrophages isolated from persistently infected spinalcords can release infectious virus (7).
In the present study, we observed two kinds of TMEV-infected cells in primary cultures of bone marrow-derived mono-
FIG. 8—Continued.
VOL. 76, 2002 INFECTION OF MACROPHAGES BY THEILER’S VIRUS 12831
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

cytes/macrophages from SJL/J mice. At early times p.i., themajority of infected cells were round, covered with filopodia,and contained large amounts of viral antigens diffusely distrib-uted in the cytoplasm (Fig. 3A). At the same time, there was acytopathic effect in the culture and the production of infectiousparticles in the medium, suggestive of active viral replication.Later on, infected cells were flat and without filopodia and theycontained punctate inclusions of viral antigens. At that timethere was no cytopathic effect and little or no infectivity re-leased in the culture medium, although a small amount ofinfectivity (approximately 102 PFU/104 cells) was found asso-ciated with the cells (not shown). These observations suggestthat replication becomes restricted at this stage and that thereis sequestration of infectious particles in intracellular sites.Cell-associated infectivity in the absence of release of infec-tious particles could contribute to the persistence of the infec-tion in vivo. Clearly, these two types of infected macrophagesare very reminiscent of the two types of infected cells describedin vivo in SJL/J mice, using immunofluorescence (21). Exper-iments are under way, using a newly developed quantitativeassay in which immunofluorescence is coupled to fluorescent insitu hybridization, to compare the ratio of expression of viralRNA to that of viral capsid proteins in cultured macrophagesand in cells in vivo during persistent infection.
The cells that we infected in vitro were differentiated in thepresence of M-CSF (L929 conditioned medium). Importantly,they had the same phenotype (FA-11�, MOMA2�, F4/80�,and Mac-1�) as the macrophages, including the infected onesthat are present in vivo in chronic CNS lesions of SJL/J mice(21). It has been shown that growth of TMEV in macrophagecell lines depends on their state of differentiation, suggestingthe existence in vivo of a window of susceptibility to TMEV formonocytes entering chronic inflammatory infiltrates (14). In-terestingly, we observed in preliminary experiments that in-creasing the treatment of bone marrow-derived macrophageswith M-CSF (increasing the time of treatment or the concen-tration of M-CSF) diminishes their permissiveness to TMEV(not shown), an observation which is consistent with the above-mentioned hypothesis.
Most of the late restriction observed in cultured macro-phages could be attributed to the secretion of IFN-�/�. Thisraises the possibility that IFN-�/� plays an important role invivo in restricting viral replication during persistent infection.Unfortunately, this cannot be tested using IFNA/BR�/� micesince these animals die early of overwhelming meningoen-cephalomyelitis (9).
At a maximum, 32% of cultured macrophages were infectedin our experiments, although the cells were inoculated at anMOI of 5 PFU/cell. Increasing the MOI 10-fold raised the levelof infection to only 50% (data not shown). Since the macro-phage cultures were not clonal, this result could be due to thepresence of a subpopulation of resistant cells. However, theMOI used to infect macrophages corresponds to titers ob-tained by plaque assay on highly susceptible BHK-21 cells. It ismore likely that limited infection of macrophages simply re-flects the lower permissiveness of macrophages to TMEV in-fection. Furthermore, the secretion of IFN-�/� prevented thespread of the infection in the culture, as shown by the fact thatthe virus did spread to the entire culture when the macro-phages were from IFNA/BR�/� mice. Viral replication was
also slower in macrophages than in BHK-21 cells, and viralyield was lower (Fig. 4). Taking into account that approxi-mately 30% of macrophages were viral antigen positive at thepeak of viral production, the yield from infected macrophageswas approximately 0.5 PFU/cell, whereas it was on the order of500 PFU/cell for BHK-21 cells (Fig. 4). Interestingly, theseyields are very similar to those reported by Trottier et al. formacrophages infected in vivo and for BHK-21 cells (35). Incomparison, the yield for IFNA/BR�/� macrophages was ap-proximately 40 PFU/cell.
Genetically resistant inbred mouse strains clear TMEV in-fection after 2 to 3 weeks (3). If the infection of macrophagesis central to TMEV’s persistence, one expects some of theresistance genes to be expressed in these cells. Interestingly, weobserved that bone marrow monocytes/macrophages fromB10.S mice, a strain that is resistant although it bears the sameH-2s haplotype as the susceptible SJL/J strain, cannot be in-fected in vitro with the DA strain of Theiler’s virus (data notshown). It will be important to determine the origin of thisblock.
Infiltrating monocytes/macrophages are continuously re-cruited into white matter perivascular cuffs during persistentinfection. It is conceivable that some of these cells, which arepermissive to viral replication, although less so than BHK-21cells, become infected. The secretion of IFN-�/� will restrictreplication in the cells and limit diffusion of the infection in thetissues, but recruitment from the periphery will continuouslyreplenish the population of permissive cells. According to thisreconstruction of pathogenesis, a combination of replenish-ment of target cells and of interferon-mediated restriction ofviral replication could form the basis of TMEV persistence inthe CNS of SJL/J mice. Obviously this view might be an over-simplification. Further work may well reveal a more complexreality.
In summary, we report that primary cultures of mouse mono-cytes/macrophages could be infected by the DA strain ofTMEV. Our results, in particular the detection of two kinds ofinfected cells, are very reminiscent of observations made invivo. To our knowledge, we described here the first tissueculture system which mimics in vivo observations. This systemmakes it possible to study viral persistence at the molecularlevel in relevant cells.
ACKNOWLEDGMENTS
We thank Genevieve Milon, Institut Pasteur, for teaching us toculture bone marrow macrophages and for helpful advice and for giftsof reagents. We thank Eliane Meurs (Institut Pasteur) and Ion Gresser(Institut Curie) for advice, discussions, and gifts of reagents; Jean-Pierre Roussarie for performing some of the antigen characterizationdescribed in this work; and Mireille Gau for secretarial assistance.
Work on Theiler’s virus in M.B.’s laboratory is supported by grantsfrom Institut Pasteur, CNRS, ARSEP, and the National Multiple Scle-rosis Society.
REFERENCES
1. Antoine, J. C., C. Jouanne, T. Lang, E. Prina, C. De Chastellier, and C.Frehel. 1991. Localization of major histocompatibility complex class II mol-ecules in phagolysosomes of murine macrophages infected with Leishmaniaamazonensis. Infect. Immun. 59:764–775.
2. Belardelli, F., F. Vignaux, E. Proietti, and I. Gresser. 1984. Injection of micewith antibody to interferon renders peritoneal macrophages permissive forvesicular stomatitis virus and encephalomyocarditis virus. Proc. Natl. Acad.Sci. USA 81:602–606.
12832 MARTINAT ET AL. J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.

3. Brahic, M., and J.-F. Bureau. 1998. Genetics of susceptibility to Theiler’svirus infection. Bioessays 20:627–633.
4. Brahic, M., A. T. Haase, and E. Cash. 1984. Simultaneous in situ detectionof viral RNA and antigens. Proc. Natl. Acad. Sci. USA 81:5445–5448.
5. Cash, E., M. Chamorro, and M. Brahic. 1985. Theiler’s virus RNA andprotein synthesis in the central nervous system of demyelinating mice. Vi-rology 144:290–294.
6. Clatch, R. J., H. L. Lipton, and S. D. Miller. 1986. Characterization ofTheiler’s murine encephalomyelitis virus (TMEV)-specific delayed-type hy-persensitivity responses in TMEV-induced demyelinating disease: correla-tion with clinical signs. J. Immunol. 136:920–927.
7. Clatch, R. J., S. D. Miller, R. Metzner, M. C. Dal Canto, and H. L. Lipton.1990. Monocytes/macrophages isolated from the mouse central nervous sys-tem contain infectious Theiler’s murine encephalomyelitis virus (TMEV).Virology 176:244–254.
8. Drescher, K., P. Murray, X. Lin, J. Carlino, and M. Rodriguez. 2000.TGF-�2 reduces demyelination, virus antigen expression, and macrophagerecruitment in a viral model of multiple sclerosis. J. Immunol. 164:3207–3213.
9. Fiette, L., C. Aubert, U. Muller, S. Huang, M. Aguet, M. Brahic, and J.-F.Bureau. 1995. Theiler’s virus infection of 129Sv mice that lack the interferon�/� or interferon receptors. J. Exp. Med. 181:2069–2076.
10. Gerety, S. J., R. J. Clatch, H. L. Lipton, R. G. Goswami, M. K. Rundell, andS. D. Miller. 1991. Class II-restricted T cell responses in Theiler’s murineencephalomyelitis virus-induced demyelinating disease. IV. Identification ofan immunodominant T cell determinant on the N-terminal end of the VP2capsid protein in susceptible SJL/J mice. J. Immunol. 146:2401–2408.
11. Gerety, S. J., W. J. Karpus, A. R. Cubbon, R. G. Goswami, M. K. Rundell,J. D. Peterson, and S. D. Miller. 1994. Class II-restricted T cell responses inTheiler’s murine encephalomyelitis virus-induced demyelinating disease. V.Mapping of a dominant immunopathologic VP2 T cell epitope in susceptibleSJL/J mice. J. Immunol. 152:908–918.
12. Gerety, S. J., M. K. Rundell, M. C. Dal Canto, and S. D. Miller. 1994. ClassII-restricted T cell responses in Theiler’s murine encephalomyelitis virus-induced demyelinating disease. VI. Potentiation of demyelination with andcharacterization of an immunopathologic CD4� T cell line specific for animmunodominant VP2 epitope. J. Immunol. 152:919–929.
13. Jarousse, N., S. Syan, C. Martinat, and M. Brahic. 1998. The neurovirulenceof the DA and GDVII strains of Theiler’s virus correlates with their abilityto infect cultured neurons. J. Virol. 72:7213–7220.
14. Jelachich, M. L., P. Bandyopadhyay, K. Blum, and H. L. Lipton. 1995.Theiler’s virus growth in murine macrophage cell lines depends on the stateof differentiation. Virology 209:437–444.
15. Jelachich, M. L., C. Bramlage, and H. L. Lipton. 1999. Differentiation of M1myeloid precursor cells into macrophages results in binding and infection byTheiler’s murine encephalomyelitis virus and apoptosis. J. Virol. 73:3227–3335.
16. Jelachich, M. L., and H. Lipton. 1999. Restricted Theiler’s murine enceph-alomyelitis virus infection in murine macrophages induces apoptosis. J. Gen.Virol. 80:1701–1705.
17. Katz-Levy, Y., K. L. Neville, J. Padilla, S. Rahbe, W. S. Begolka, A. M.Girvin, J. K. Olson, C. L. Vanderlugt, and S. D. Miller. 2000. Temporal
development of autoreactive Th1 responses and endogenous presentation ofself myelin epitopes by central nervous system-resident APCs in Theiler’svirus-infected mice. J. Immunol. 165:5304–5314.
18. Levy, M., C. Aubert, and M. Brahic. 1992. Theiler’s virus replication in brainmacrophages cultured in vitro. J. Virol. 66:3188–3193.
19. Lipton, H. L. 1975. Theiler’s virus infection in mice: an unusual biphasicdisease process leading to demyelination. Infect. Immun. 11:1147–1155.
20. Lipton, H. L., and M. C. Dal Canto. 1979. The TO strains of Theiler’s virusescause “slow virus-like” infections in mice. Ann. Neurol. 6:25–28.
21. Lipton, H. L., G. Twaddle, and M. L. Jelachich. 1995. The predominant virusantigen burden is present in macrophages in Theiler’s murine encephalomy-elitis virus-induced demyelinating disease. J. Virol. 69:2525–2533.
22. McKnight, A. J., and S. Gordon. 1998. Membrane molecules as differenti-ation antigens of murine macrophages. Adv. Immunol. 68:271–314.
23. Metcalf, D. 1989. The molecular control of cell division, differentiationcommitment and maturation in haematopoietic cells. Nature 339:27–30.
24. Miller, S. D., C. L. Vanderlugt, W. S. Begolka, W. Pao, R. L. Yauch, K. L.Neville, Y. Katz-Levy, A. Carrizosa, and B. S. Kim. 1997. Persistent infectionwith Theiler’s virus leads to CNS autoimmunity via epitope spreading. Na-ture Med. 3:1133–1136.
25. Morioka, Y., M. Naito, T. Sato, and K. Takahashi. 1994. Immunophenotypicand ultrastructural heterogeneity of macrophage differentiation in bone mar-row and fetal hematopoiesis of mouse in vitro and in vivo. J. Leukoc. Biol.55:642–651.
26. Muller, C. D., and F. Schuber. 1986. Fluorometric determination of poly-styrene latex: application to the measurement of phagosomes and phagocy-tosis. Anal. Biochem. 152:167–171.
27. Muller, U., U. Steinhoff, L. F. L. Reis, S. Hemmi, J. Pavlovic, R. M. Zink-ernagel, and M. Aguet. 1994. Functional role of type I and type II interferonsin antiviral defense. Science 264:1918–1921.
28. Nakagawara, A., and C. F. Nathan. 1983. A simple method for countingadherent cells: application to cultured human monocytes, macrophages andmultinucleated giant cells. J. Immunol. Methods 56:262–268.
29. Oda, T., and H. Maeda. 1986. A new simple fluorometric assay for phago-cytosis. J. Immunol. Methods 88:175–183.
30. Pena Rossi, C., M. Delcroix, I. Huitinga, A. McAllister, N. van Rooijen, E.Claassen, and M. Brahic. 1997. Role of macrophages during Theiler’s virusinfection. J. Virol. 71:3336–3340.
31. Proietti, E., S. Gessani, F. Belardelli, and I. Gresser. 1986. Mouse peritonealcells confer an antiviral state on mouse cell monolayers: role of interferon.J. Virol. 57:456–463.
32. Shaw-Jackson, C., and T. Michiels. 1997. Infection of macrophages by Thei-ler’s murine encephalomyelitis virus is highly dependent on their activationor differentiation state. J. Virol. 71:8864–8867.
33. Stanley, E. R., K. L. Berg, D. B. Einstein, P. S. Lee, F. L. Pixley, Y. Wang, andY. G. Yeung. 1997. Biology and action of colony stimulating factor-1. Mol.Reprod. Dev. 46:4–10.
34. Stewart, W. E., E. D. De Clerqc, and P. De Somer. 1974. Stabilization ofinterferons by defensive reversible denaturation. Nature 249:460–461.
35. Trottier, M., P. Kallio, W. Wang, and H. L. Lipton. 2001. High numbers ofviral RNA copies in the central nervous system of mice during persistentinfection with Theiler’s virus. J. Virol. 75:7420–7428.
VOL. 76, 2002 INFECTION OF MACROPHAGES BY THEILER’S VIRUS 12833
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 20
Nov
embe
r 20
21 b
y 11
9.10
6.95
.210
.